
Sitagliptin Modulates Oxidative, Nitrative and Halogenative Stress and Inflammatory Response in Rat Model of Hepatic Ischemia-Reperfusion


 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Chemicals
2.3. IR Procedure
2.4. Analytical Methods
2.4.1. Transcriptomic Analysis
2.4.2. Liver Homogenates and Protein Quantification
2.4.3. Cytoprofiling
2.4.4. Metabolomic Analysis
2.5. Statistical Analysis
3. Results
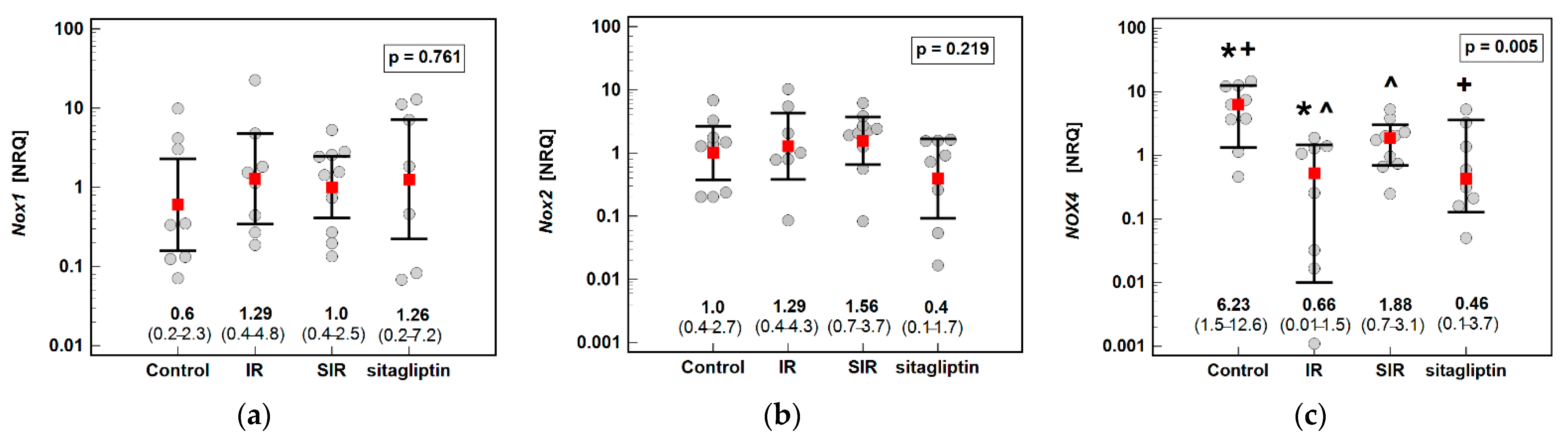
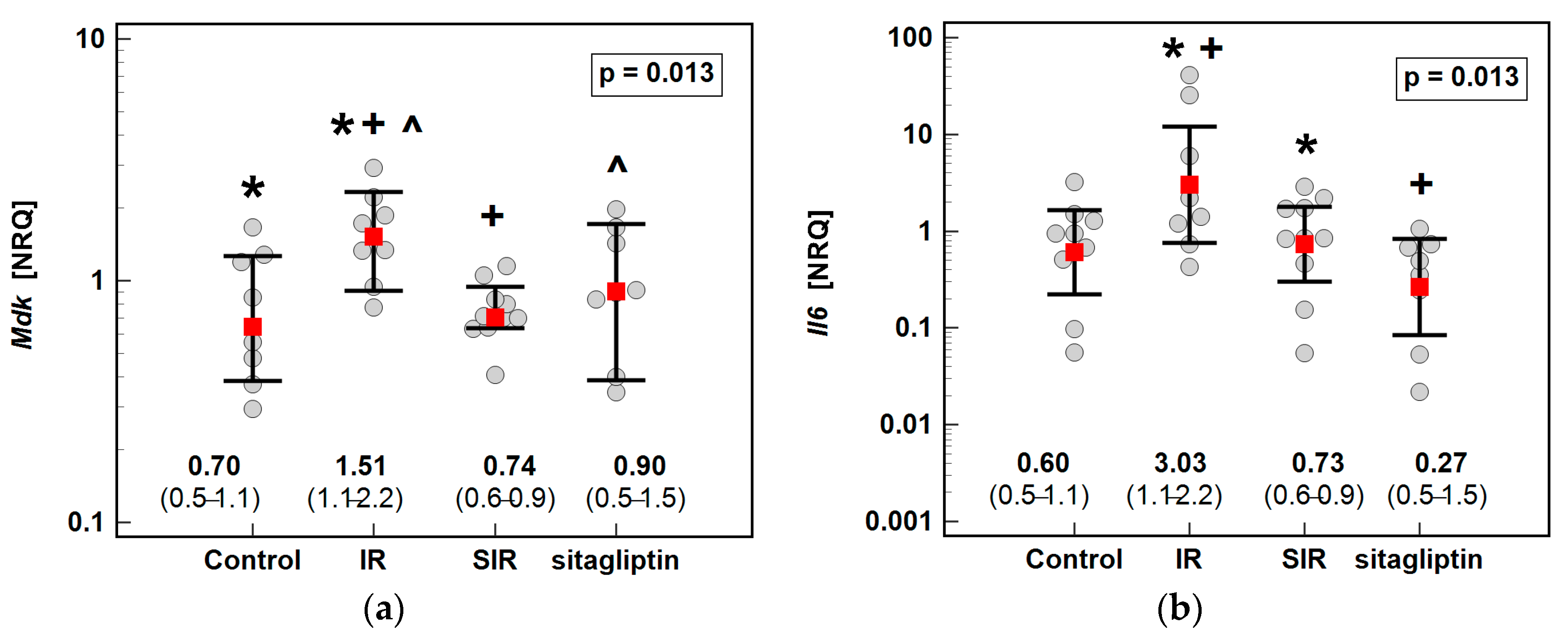
3.1. Transcriptomic Analysis
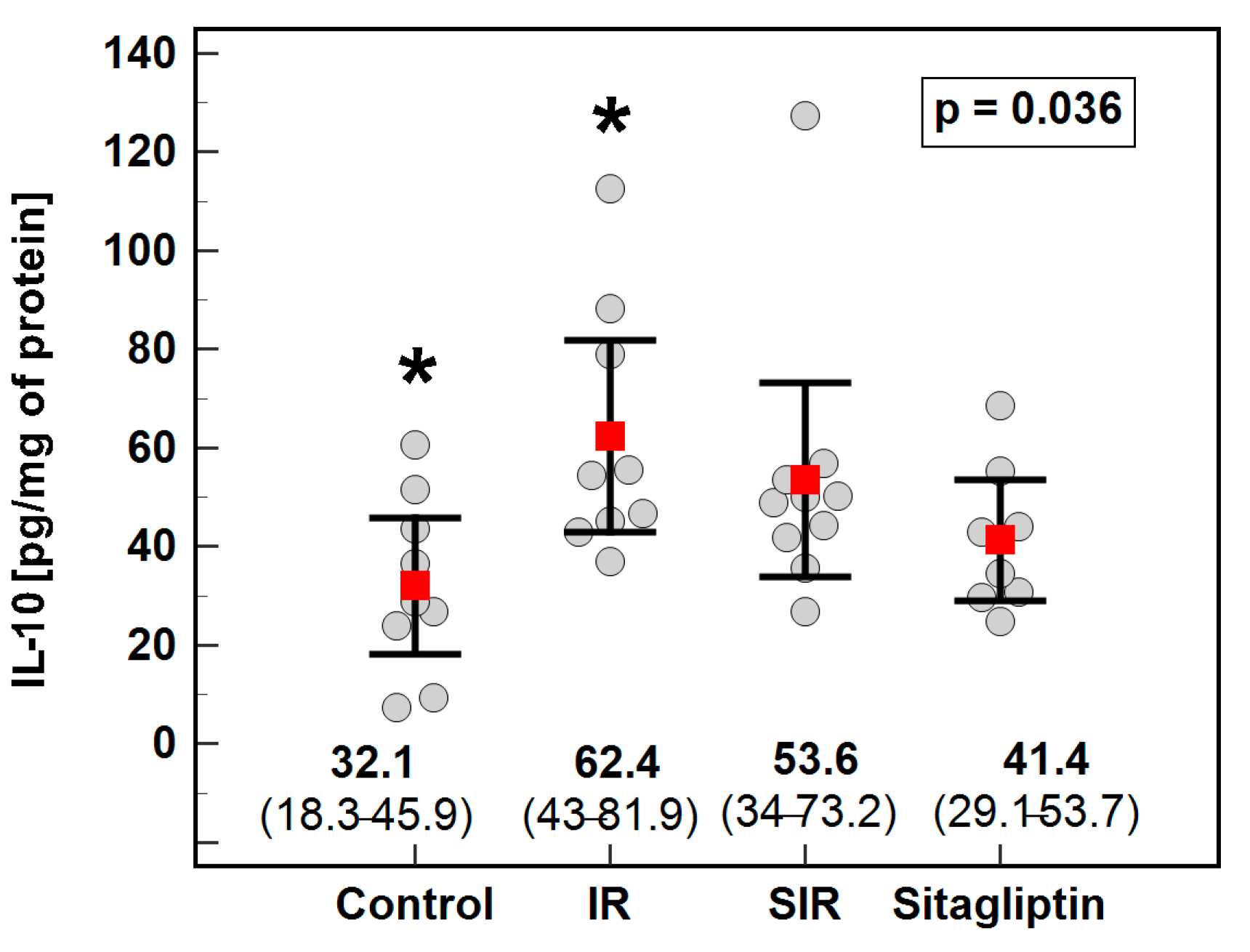
3.2. Cytoprofiling
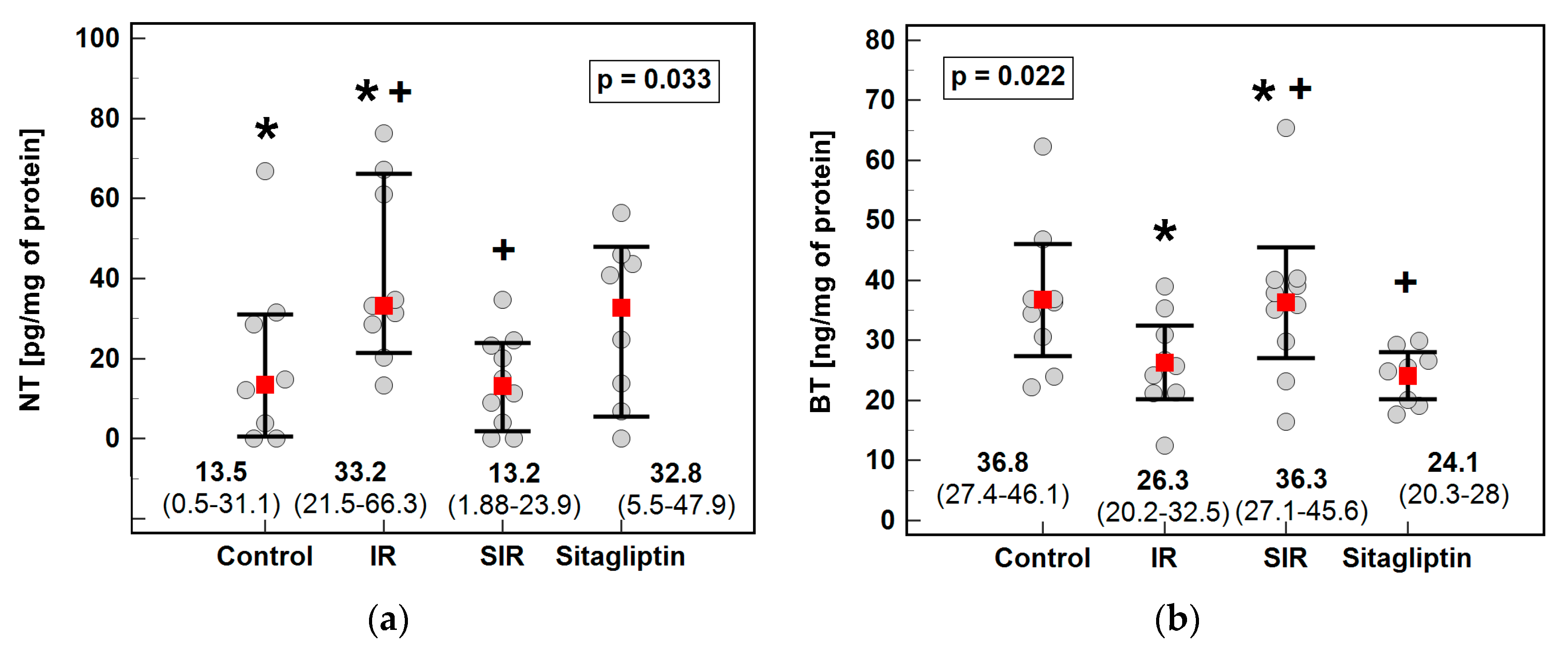
3.3. 3-Nitro- (NT), 3-Chloro- (CT) and 3-Bromotyrosine (BT)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rampes, S.; Ma, D. Hepatic ischemia-reperfusion injury in liver transplant setting: Mechanisms and protective strategies. J. Biomed. Res. 2019, 33, 221–234. [Google Scholar] [CrossRef]
- Fan, C.; Zwacka, R.M.; Engelhardt, J.F. Therapeutic approaches for ischemia/reperfusion injury in the liver. J. Mol. Med. 1999, 77, 577–596. [Google Scholar] [CrossRef] [PubMed]
- Hines, I.N.; Harada, H.; Flores, S.; Gao, B.; McCord, J.M.; Grisham, M.B. Endothelial nitric oxide synthase protects the post-ischemic liver: Potential interactions with superoxide. Biomed. Pharmacother. 2005, 59, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Jaeschke, H.; Woolbright, B.L. Current strategies to minimize hepatic ischemia-reperfusion injury by targeting reactive oxygen species. Transplant. Rev. 2012, 26, 103–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moldogazieva, N.T.; Mokhosoev, I.M.; Mel’nikova, T.I.; Zavadskiy, S.P.; Kuz’menko, A.N.; Terentiev, A.A. Dual character of reactive oxygen, nitrogen, and halogen species: Endogenous sources, interconversions and neutralization. Biochemistry 2020, 85 (Suppl. 1), S56–S78. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Chen, Y.; d’Avignon, A.; Hazen, S.L. 3-Bromotyrosine and 3,5-dibromotyrosine are major products of protein oxidation by eosinophil peroxidase: Potential markers for eosinophil-dependent tissue injury in vivo. Biochemistry 1999, 38, 3538–3548. [Google Scholar] [CrossRef] [PubMed]
- Hazen, S.L.; Heinecke, J.W. 3-Chlorotyrosine, a specific marker of myeloperoxidase-catalyzed oxidation, is markedly elevated in low density lipoprotein isolated from human atherosclerotic intima. J. Clin. Investig. 1997, 99, 2075–2081. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, G.A.; Cheng, R.Y.S.; Ridnour, L.A.; Basudhar, D.; Somasundaram, V.; McVicar, D.W.; Monteiro, H.P.; Wink, D.A. Inducible nitric oxide synthase in the carcinogenesis of gastrointestinal cancers. Antioxid. Redox Signal. 2017, 26, 1059–1077. [Google Scholar] [CrossRef] [PubMed]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug repurposing: Progress, challenges and recommendations. Nat. Rev. Drug Discov. 2019, 18, 41–58. [Google Scholar] [CrossRef]
- Sherif, I.O.; Al-Shaalan, N.H. Vildagliptin attenuates hepatic ischemia/reperfusion Iinjury via the TLR4/NF-κB signaling pathway. Oxid. Med. Cell. Longev. 2018, 2018, 3509091. [Google Scholar] [CrossRef] [Green Version]
- Bradic, J.; Milosavljevic, I.; Bolevich, S.; Litvitskiy, P.F.; Jeremic, N.; Bolevich, S.; Zivkovic, V.; Srejovic, I.; Jeremic, J.; Jovicic, N.; et al. Dipeptidyl peptidase 4 inhibitors attenuate cardiac ischaemia-reperfusion injury in rats with diabetes mellitus type 2. Clin. Exp. Pharmacol. Physiol. 2021, 48, 575–584. [Google Scholar] [CrossRef]
- Glorie, L.L.; Verhulst, A.; Matheeussen, V.; Baerts, L.; Magielse, J.; Hermans, N.; D’Haese, P.C.; De Meester, I.; De Beuf, A. DPP4 inhibition improves functional outcome after renal ischemia-reperfusion injury. Am. J. Physiol. Renal Physiol. 2012, 303, F681–F688. [Google Scholar] [CrossRef]
- Rankovic, M.; Jeremic, N.; Srejovic, I.; Radonjic, K.; Stojanovic, A.; Glisic, M.; Bolevich, S.; Bolevich, S.; Jakovljevic, V. Dipeptidyl peptidase-4 inhibitors as new tools for cardioprotection. Heart Fail. Rev. 2021, 26, 437–450. [Google Scholar] [CrossRef]
- Nam, D.H.; Park, J.; Park, S.H.; Kim, K.S.; Baek, E.B. Effect of gemigliptin on cardiac ischemia/reperfusion and spontaneous hypertensive rat models. Korean J. Physiol. Pharmacol. 2019, 23, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Mulvihill, E.E.; Drucker, D.J. Pharmacology, physiology, and mechanisms of action of dipeptidyl peptidase-4 inhibitors. Endocr. Rev. 2014, 35, 992–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, S.; Xu, Q.; Yu, X.; Pan, R.; Chen, Y. Dipeptidyl peptidase 4 inhibitors and their potential immune modulatory functions. Pharmacol. Ther. 2020, 209, 107503. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Watanabe, A.; Tanaka, M.; Shiota, M.; Osada-Oka, M.; Sano, S.; Yoshiyama, M.; Miura, K.; Kitajima, S.; Matsunaga, S.; et al. A dipeptidyl peptidase-4 (DPP-4) inhibitor, linagliptin, attenuates cardiac dysfunction after myocardial infarction independently of DPP-4. J. Pharmacol. Sci. 2019, 139, 112–119. [Google Scholar] [CrossRef]
- Vaghasiya, J.; Sheth, N.; Bhalodia, Y.; Manek, R. Sitagliptin protects renal ischemia reperfusion induced renal damage in diabetes. Regul. Pept. 2011, 166, 48–54. [Google Scholar] [CrossRef]
- Chen, Y.T.; Tsai, T.H.; Yang, C.C.; Sun, C.K.; Chang, L.T.; Chen, H.H.; Chang, C.L.; Sung, P.H.; Zhen, Y.Y.; Leu, S.; et al. Exendin-4 and sitagliptin protect kidney from ischemia-reperfusion injury through suppressing oxidative stress and inflammatory reaction. J. Transl. Med. 2013, 11, 270–289. [Google Scholar] [CrossRef] [Green Version]
- Scott, L.J. Sitagliptin: A review in type 2 diabetes. Drugs 2017, 77, 209–224. [Google Scholar] [CrossRef]
- Trocha, M.; Nowak, B.; Merwid-Ląd, A.; Szuba, A.; Dzięgiel, P.; Pieśniewska, M.; Gomułkiewicz, A.; Wiśniewski, J.; Piasecki, T.; Gziut, M.; et al. The impact of sitagliptin, inhibitor of dipeptidyl peptidase-4 (DPP-4), on the ADMA-DDAH-NO pathway in ischemic and reperfused rat livers. Adv. Clin. Exp. Med. 2018, 27, 1483–1490. [Google Scholar] [CrossRef]
- Trocha, M.; Krzystek-Korpacka, M.; Merwid-Ląd, A.; Nowak, B.; Pieśniewska, M.; Dzięgiel, P.; Gomułkiewicz, A.; Kowalski, P.; Diakowska, D.; Szeląg, A.; et al. Sitagliptin-dependent differences in the intensity of oxidative sStress in rat livers subjected to ischemia and reperfusion. Oxid. Med. Cell. Longev. 2019, 2019, 2738605. [Google Scholar] [CrossRef] [Green Version]
- Al-Awar, A.; Almási, N.; Szabó, R.; Takacs, I.; Murlasits, Z.; Szűcs, G.; Török, S.; Pósa, A.; Varga, C.; Kupai, K. Novel potentials of the DPP-4 inhibitor sitagliptin against ischemia-reperfusion (I/R) injury in rat ex-vivo heart model. Int. J. Mol. Sci. 2018, 19, 3226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khedr, R.M.; Ahmedt, A.A.E.; Kamel, R.; Raafat, E.M. Sitagliptin attenuates intestinal ischemia/reperfusion injury via cAMP/PKA, PI3K/Akt pathway in a glucagon-like peptide 1 receptor-dependent manner. Life Sci. 2018, 211, 1–39. [Google Scholar] [CrossRef]
- Abdel-Gaber, S.A.; Geddawy, A.; Moussa, R.A. The hepatoprotective effect of sitagliptin against hepatic ischemia reperfusion-induced injury in rats involves Nrf-2/HO-1 pathway. Pharmacol. Rep. 2019, 71, 1044–1049. [Google Scholar] [CrossRef]
- Song-Chol, M.; Hye-Sun, H. Dipeptidyl peptidase-4 inhibitor sitagliptin prevents hepatic injury via liver ischemia/reperfusion in rats. Int. J. Clin. Exp. Physiol. 2018, 5, 123–126. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, C.; Ma, H.; Huang, Z.; Li, J.; Lou, J.; Li, B.; Tu, Q.; Gao, W. Detrimental effect of sitagliptin induced autophagy on multiterritory perforator flap survival. Front. Pharmacol. 2020, 11, 951. [Google Scholar] [CrossRef] [PubMed]
- Nuransoy, A.; Beytur, A.; Polat, A.; Samdanci, E.; Sagir, M.; Parlakpinar, H. Protective effect of sitagliptin against renal ischemia reperfusion injury in rats. Ren. Fail. 2015, 37, 687–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, M.W.; Chen, C.H.; Chen, Y.C.; Wu, Y.C.; Zhen, Y.Y.; Leu, S.; Tsai, T.H.; Ko, S.F.; Sung, P.H.; Yang, C.C.; et al. Sitagliptin protects rat kidneys from acute ischemia-reperfusion injury via upregulation of GLP-1 and GLP-1 receptors. Acta Pharmacol. Sin. 2015, 36, 119–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Sahar, A.E.; Safar, M.M.; Zaki, H.F.; Attia, A.S.; Ain-Shoka, A.A. Sitagliptin attenuates transient cerebral ischemia/reperfusion injury in diabetic rats: Implication of the oxidative-inflammatory-apoptotic pathway. Life Sci. 2015, 126, 81–86. [Google Scholar] [CrossRef]
- Abdel-Aziz, A.M.; Naguib Abdel Hafez, S.M. Sitagliptin protects male albino rats with testicular ischaemia/reperfusion damage: Modulation of VCAM-1 and VEGF-A. Andrologia 2020, 52, e13472. [Google Scholar] [CrossRef] [PubMed]
- Khedr, R.; Ahmed, A.; Kamel, R.; Rafaat, E. Antioxidant effects of sitagliptin in a rat model of intestinal ischemia/reperfusion injury. J. Adv. Pharm. Res. 2021, 5, 234–240. [Google Scholar] [CrossRef]
- Song, R.; Ren, J.; Sun, J.; Li, M. Melatonin postconditioning combined with sitagliptin exerts full cardioprotection in diabetic hearts of aged rats through an AMPK-dependent mechanism. Arch. Biol. Sci. 2021, 73, 83–92. Available online: https://www.serbiosoc.org.rs/arch/index.php/abs/article/view/6144 (accessed on 9 June 2021). [CrossRef]
- Hellemans, J.; Vandesompele, J. qPCR data analysis—Unlocking the secret to successful results. In PCR Troubleshooting and Optimization: The Essential Guide, 1st ed.; Kennedy, S., Oswald, N., Eds.; Caister Academic Press: Poole, UK, 2011; pp. 1–13. [Google Scholar]
- Fleszar, M.G.; Fortuna, P.; Zawadzki, M.; Kosyk, B.; Krzystek-Korpacka, M. Simultaneous LC-MS/MS-based quantification of free 3-nitro-L-tyrosine, 3-chloro-L-tyrosine, and 3-bromo-L-tyrosine in plasma of colorectal cancer patients during early postoperative period. Molecules 2020, 25, 5158. [Google Scholar] [CrossRef]
- Hewedy, W.A. Effects of treatment with sitagliptin on hepatotoxicity induced by acetaminophen in mice. Braz. J. Pharm. Sci. 2020, 56, 13. Available online: https://www.scielo.br/j/bjps/a/RjzXGxphxnzrZ3MxxwKdCWD/ (accessed on 9 June 2021). [CrossRef]
- He, F.; Ru, X.; Wen, T. NRF2, a transcription factor for stress response and beyond. Int. J. Mol. Sci. 2020, 21, 4777. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.X.; Török, N.J. NADPH oxidases in chronic liver diseases. Adv. Hepatol. 2014, 2014, 742931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortezaee, K. Nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX) and liver fibrosis: A review. Cell Biochem. Funct. 2018, 36, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hausding, M.; Weng, S.Y.; Kim, Y.O.; Steven, S.; Klein, T.; Daiber, A.; Schuppan, D. Gliptins suppress inflammatory macrophage activation to mitigate inflammation, fibrosis, oxidative stress, and vascular dysfunction in models of nonalcoholic steatohepatitis and liver fibrosis. Antioxid. Redox Signal. 2018, 28, 87–109. [Google Scholar] [CrossRef]
- Eun, H.S.; Cho, S.Y.; Joo, J.S.; Kang, S.H.; Moon, H.S.; Lee, E.S.; Kim, S.H.; Lee, B.S. Gene expression of NOX family members and their clinical significance in hepatocellular carcinoma. Sci. Rep. 2017, 7, 11060. [Google Scholar] [CrossRef]
- Buelna-Chontal, M.; García-Niño, W.R.; Silva-Palacios, A.; Enríquez-Cortina, C.; Zazueta, C. Implications of oxidative and nitrosative post-translational modifications in therapeutic strategies against reperfusion damage. Antioxidants 2021, 10, 749. [Google Scholar] [CrossRef] [PubMed]
- Abo-Haded, H.M.; Elkablawy, M.A.; Al-johani, Z.; Al-ahmadi, O.; El-Agamy, D.S. Hepatoprotective effect of sitagliptin against methotrexate induced liver toxicity. PLoS ONE 2017, 12, e0174295. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Williams, V.; Liu, L.; Chen, H.; Sawamura, T.; Antakli, T.; Mehta, J.L. LOX-1 inhibition in myocardial ischemia-reperfusion injury: Modulation of MMP-1 and inflammation. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H1795–H1801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klune, J.R.; Bartels, C.; Luo, J.; Yokota, S.; Du, Q.; Geller, D.A. IL-23 mediates murine liver transplantation ischemia-reperfusion injury via IFN-γ/IRF-1 pathway. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G991–G1002. [Google Scholar] [CrossRef] [PubMed]
- Khodeert, D.M.; Bilasy, S.E.; Farag, N.E.; Mehana, A.E.; Elbaz, A.A. Sitagliptin protects diabetic rats with acute myocardial infarction through induction of angiogenesis: Role of IGF-1 and VEGF. Can. J. Physiol. Pharmacol. 2019, 97, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Cornide-Petronio, M.E.; Jiménez-Castro, M.B.; Gracia-Sancho, J.; Peralta, C. New insights into the liver-visceral adipose aAxis during hepatic resection and liver transplantation. Cells 2019, 8, 1100. [Google Scholar] [CrossRef] [Green Version]
- Yu, A.; Zhou, R.; Xia, B.; Dang, W.; Yang, Z.; Chen, X. NAMPT maintains mitochondria content via NRF2-PPARα/AMPKα pathway to promote cell survival under oxidative stress. Cell. Signal. 2020, 66, 109496. [Google Scholar] [CrossRef] [PubMed]
- Bala, M.; Martin, J.; Kopp, A.; Hanses, F.; Buechler, C.; Schäffler, A. In vivo suppression of visfatin by oral glucose uptake: Evidence for a novel incretin-like effect by glucagon-like peptide-1 (GLP-1). J. Clin. Endocrinol. Metab. 2011, 96, 2493–2501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashima, E. Visfatin and resistin: Mediators of the pleiotropic effects of incretins? J. Diabetes Investig. 2012, 3, 427–428. [Google Scholar] [CrossRef] [Green Version]
- Sorrelle, N.; Dominguez, A.T.A.; Brekken, R.A. From top to bottom: Midkine and pleiotrophin as emerging players in immune regulation. J. Leukoc. Biol. 2017, 102, 277–286. [Google Scholar] [CrossRef]
- Miao, J.; Ding, M.; Zhang, A.; Xiao, Z.; Qi, W.; Luo, N.; Di, W.; Tao, Y.; Fang, Y. Pleiotrophin promotes microglia proliferation and secretion of neurotrophic factors by activating extracellular signal-regulated kinase 1/2 pathway. Neurosci. Res. 2012, 74, 269–276. [Google Scholar] [CrossRef]
- Christman, K.L.; Fang, Q.; Kim, A.J.; Sievers, R.E.; Fok, H.H.; Candia, A.F.; Colley, K.J.; Herradon, G.; Ezquerra, L.; Deuel, T.F.; et al. Pleiotrophin induces formation of functional neovasculature invivo. Biochem. Biophys. Res. Commun. 2005, 332, 1146–1152. [Google Scholar] [CrossRef] [PubMed]
- Horiba, M.; Kadomatsu, K.; Yasui, K.; Lee, J.K.; Takenaka, H.; Sumida, A.; Kamiya, K.; Chen, S.; Sakuma, S.; Muramatsu, T.; et al. Midkine plays a protective role against cardiac ischemia/reperfusioninjury through a reduction of apoptotic reaction. Circulation 2006, 114, 1713–1720. [Google Scholar] [CrossRef] [Green Version]
- Sato, W.; Kadomatsu, K.; Yuzawa, Y.; Muramatsu, H.; Hotta, N.; Matsuo, S.; Muramatsu, T. Midkine is involved in neutrophil infiltrationinto the tubulointerstitium in ischemic renal injury. J. Immunol. 2001, 167, 3463–3469. [Google Scholar] [CrossRef] [Green Version]
- Weckbach, L.T.; Groesser, L.; Borgolte, J.; Pagel, J.I.; Pogoda, F.; Schymeinsky, J.; Muller-Hocker, J.; Shakibaei, M.; Muramatsu, T.; Deindl, E.; et al. Midkine acts as proangiogenic cytokine inhypoxia-induced angiogenesis. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, H429–H438. [Google Scholar] [CrossRef] [Green Version]
- Ochiai, K.; Muramatsu, H.; Yamamoto, S.; Ando, H.; Muramatsu, T. The role of midkine and pleiotrophin in liver regeneration. Liver Int. 2004, 24, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Yazihan, N.; Kocak, M.K.; Akcil, E.; Erdem, O.; Sayal, A. Role of midkine in cadmium-induced liver, heart and kidney damage. Hum. Exp. Toxicol. 2011, 30, 391–397. [Google Scholar] [CrossRef]
- Tang, J.; Yan, Z.; Feng, Q.; Yu, L.; Wang, H. The roles of neutrophils in the pathogenesis of liver diseases. Front. Immunol. 2021, 12, 625472. [Google Scholar] [CrossRef]
- Hilscher, M.B.; Shah, V.H. Neutrophil extracellular traps and liver disease. Semin. Liver Dis. 2020, 40, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Weckbach, L.T.; Grabmaier, U.; Uhl, A.; Gess, S.; Boehm, F.; Zehrer, A.; Pick, R.; Salvermoser, M.; Czermak, T.; Pircher, J.; et al. Midkine drives cardiac inflammation by promoting neutrophil trafficking and NETosis in myocarditis. J. Exp. Med. 2019, 216, 350–368. [Google Scholar] [CrossRef]
- Santocki, M.; Kolaczkowska, E. On neutrophil extracellular trap (NET) removal: What we know thus far and why so little. Cells 2020, 9, 2079. [Google Scholar] [CrossRef] [PubMed]
- Lee, V.G.; Johnson, M.L.; Baust, J.; Laubach, V.E.; Watkins, S.C.; Billiar, T.R. The roles of iNOS in liver ischemia-reperfusion injury. Shock 2001, 16, 355–360. [Google Scholar] [CrossRef]
- Li Volti., G.; Sorrenti, V.; Acquaviva, R.; Murabito, P.; Gullo, A.; Barcellona, M.L.; Galvano, F.; Rodella, L.; Rezzani, R.; Vanella, L.; et al. Effect of ischemia-reperfusion on renal expression and activity of N(G)-N(G)-dimethylarginine dimethylaminohydrolases. Anesthesiology 2008, 109, 1054–1062. [Google Scholar] [CrossRef] [Green Version]
- Siriussawakul, A.; Zaky, A.; Lang, J.D. Role of nitric oxide in hepatic ischemia-reperfusion injury. World J. Gastroenterol. 2010, 16, 6079–6086. [Google Scholar] [CrossRef] [PubMed]
- Hide, D.; Ortega-Ribera, M.; Garcia-Pagan, J.C.; Peralta, C.; Bosch, J.; Gracia-Sancho, J. Effects of warm ischemia and reperfusion on the liver microcirculatory phenotype of rats: Underlying mechanisms and pharmacological therapy. Sci. Rep. 2016, 6, 22107. [Google Scholar] [CrossRef] [Green Version]
- Keshet, R.; Erez, A. Arginine and the metabolic regulation of nitric oxide synthesis in cancer. Dis. Models Mech. 2018, 11, dmm033332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Yang, Y.; Wang, M.; Wang, S.; Jeong, J.M.; Xu, L.; Wen, Y.; Emontzpohl, C.; Atkins, C.L.; Duong, K.; et al. Eosinophils attenuate hepatic ischemia-reperfusion injury in mice through ST2-dependent IL-13 production. Sci. Transl. Med. 2021, 13, eabb6576. [Google Scholar] [CrossRef]
- Senthilmohan, R.; Kettle, A.J. Bromination and chlorination reactions of myeloperoxidase at physiological concentrations of bromide and chloride. Arch. Biochem. Biophys. 2006, 445, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Kagiyama, T.; Glushakov, A.V.; Sumners, C.; Roose, B.; Dennis, D.M.; Phillips, M.I.; Ozcan, M.S.; Seubert, C.N.; Martynyuk, A.E. Neuroprotective action of halogenated derivatives of L-phenylalanine. Stroke 2004, 35, 1192–1196. [Google Scholar] [CrossRef] [Green Version]
- Hollande, C.; Boussier, J.; Ziai, J.; Nozawa, T.; Bondet, V.; Phung, W.; Lu, B.; Duffy, D.; Paradis, V.; Mallet, V.; et al. Inhibition of the dipeptidyl peptidase DPP4 (CD26) reveals IL-33-dependent eosinophil-mediated control of tumor growth. Nat. Immunol. 2019, 20, 257–264. [Google Scholar] [CrossRef]
- Sun, Z.; Su, Q.; Rokita, S.E. The distribution and mechanism of iodotyrosine deiodinase defied expectations. Arch. Biochem. Biophys. 2017, 632, 77–87. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Sequence | Reverse Sequence |
|---|---|---|
| Gapdh | TGACTCTACCCACGGCAAGTTCAA | ACGACATACTCAGCACCAGCATCA |
| Il6 | ACAGCGATGATGCACTGTCAG | ATGGTCTTGGTCCTTAGCCAC |
| Tnfa | GCCCAGACCCTCACACTC | CCACTCCAGCTGCTCCTCT |
| Mmp1 | CCACTAACATTCGAAAGGGTTT | GGTCCATCAAATGGGTTATTG |
| Nampt | TCTGGAAATCCGCTCGACAC | TATCCACTCCGTCCCCTTGA |
| Mdk | TGGAGCCGACTGCAAATAC | TGTACCGAGCCTTCTTCAGG |
| Nox1 | TTCCCTGGAACAAGAGATGG | GACGTCAGTGGCTCTGTCAA |
| Nox2 | CTGCCAGTGTGTCGGAATCT | TGTGAATGGCCGTGTGAAGT |
| Nox4 | GGATCACAGAAGGTCCCTAGC | AGAAGTTCAGGGCGTTCACC |
| Ptn | TGGAGCTGAGTGCAAATAC | TGTGCAGAGCTCTCTTCAGA |
| Gene | Mean (95% CI) (NRQ) | p | |||
|---|---|---|---|---|---|
| Control | IR | SIR | Sitagliptin | ||
| Mmp1 | 0.37 (0.1–1.0) | 1.09 (0.1–9.4) | 0.68 (0.2–2.9) | 3.36 (0.3–38.7) | 0.244 |
| Nampt | 1.12 (0.5–2.5) | 1.25 (0.5–3.0) | 1.39 (0.8–2.4) | 0.50 (0.2–1.3) | 0.158 |
| Ptn | 1.25 (0.7–2.4) | 1.0 (0.6–1.6) | 1.21 (0.8–1.8) | 0.66 (0.4–1.1) | 0.172 |
| Tnfa | 0.77 (0.4–1.5) | 1.07 (0.3–4.2) | 1.21 (0.6–2.6) | 0.81 (0.3–2.6) | 0.839 |
| Gene | Nampt | Mmp1 | Nox2 | Il6 | Nox1 | Mdk | Tnfa | Nox4 |
|---|---|---|---|---|---|---|---|---|
| Ptn | 0.76 3 | ns | 0.52 2 | ns | ns | ns | ns | 0.47 2 |
| Nampt | ns | 0.60 3 | 0.34 1 | −0.38 1 | ns | −0.41 1 | 0.45 2 | |
| Mmp1 | −0.41 1 | ns | 0.73 3 | 0.41 1 | ns | ns | ||
| Nox2 | ns | −0.43 2 | ns | ns | 0.35 1 | |||
| Il6 | ns | ns | ns | ns | ||||
| Nox1 | 0.49 2 | 0.53 3 | −0.56 3 | |||||
| Mdk | ns | −0.49 2 | ||||||
| Tnfa | −0.44 2 |
| Cytokine | Mean (95% CI) (pg/mg of Protein) | p | |||
|---|---|---|---|---|---|
| Control | IR | SIR | Sitagliptin | ||
| IL-1β | 46.7 (27.3–66.1) | 48.6 (23.2–74) | 50.8 (44.3–57.2) | 62.1 (37.4–86.8) | 0.613 |
| IFN-γ | 211.8 (123–300) | 201.5 (136–267) | 207.1 (138–276) | 209.7 (173–246) | 0.995 |
| MIP-2 | 104.6 (63–146) | 91.4 (75–108) | 103 (77–129) | 90.7 (51–130) | 0.825 |
| TNF-α | 0.90 (0.2–1.6) | 0.49 (0.2–0.8) | 0.73 (0.2–1.3) | 0.57 (0.4–0.8) | 0.551 |
| VEGF-A | 31.7 (22.9–40.5) | 34.6 (26.9–42.4) | 39.0 (32.1–46) | 37.3 (30.2–44.4) | 0.426 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trocha, M.; Fleszar, M.G.; Fortuna, P.; Lewandowski, Ł.; Gostomska-Pampuch, K.; Sozański, T.; Merwid-Ląd, A.; Krzystek-Korpacka, M. Sitagliptin Modulates Oxidative, Nitrative and Halogenative Stress and Inflammatory Response in Rat Model of Hepatic Ischemia-Reperfusion. Antioxidants 2021, 10, 1168. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10081168
Trocha M, Fleszar MG, Fortuna P, Lewandowski Ł, Gostomska-Pampuch K, Sozański T, Merwid-Ląd A, Krzystek-Korpacka M. Sitagliptin Modulates Oxidative, Nitrative and Halogenative Stress and Inflammatory Response in Rat Model of Hepatic Ischemia-Reperfusion. Antioxidants. 2021; 10(8):1168. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10081168
Chicago/Turabian StyleTrocha, Małgorzata, Mariusz G. Fleszar, Paulina Fortuna, Łukasz Lewandowski, Kinga Gostomska-Pampuch, Tomasz Sozański, Anna Merwid-Ląd, and Małgorzata Krzystek-Korpacka. 2021. "Sitagliptin Modulates Oxidative, Nitrative and Halogenative Stress and Inflammatory Response in Rat Model of Hepatic Ischemia-Reperfusion" Antioxidants 10, no. 8: 1168. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10081168