
UHPLC-MS Characterization and Biological Insights of Different Solvent Extracts of Two Achillea Species (A. aleppica and A. santolinoides) from Turkey


 , , ,
, , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Collection and Extract Preparation
2.2. Total Phenolic and Flavonoid Content
2.3. Ultra-High Performance Liquid Chromatography Coupled with High Resolution Mass Spectrometry (UHPLC-HRMS)
2.4. Determination of Antioxidant and Enzyme Inhibitory Effects
2.5. Statistical Analysis
2.6. Bioinformatics Analysis
3. Results and Discussion
3.1. Chemical Profile
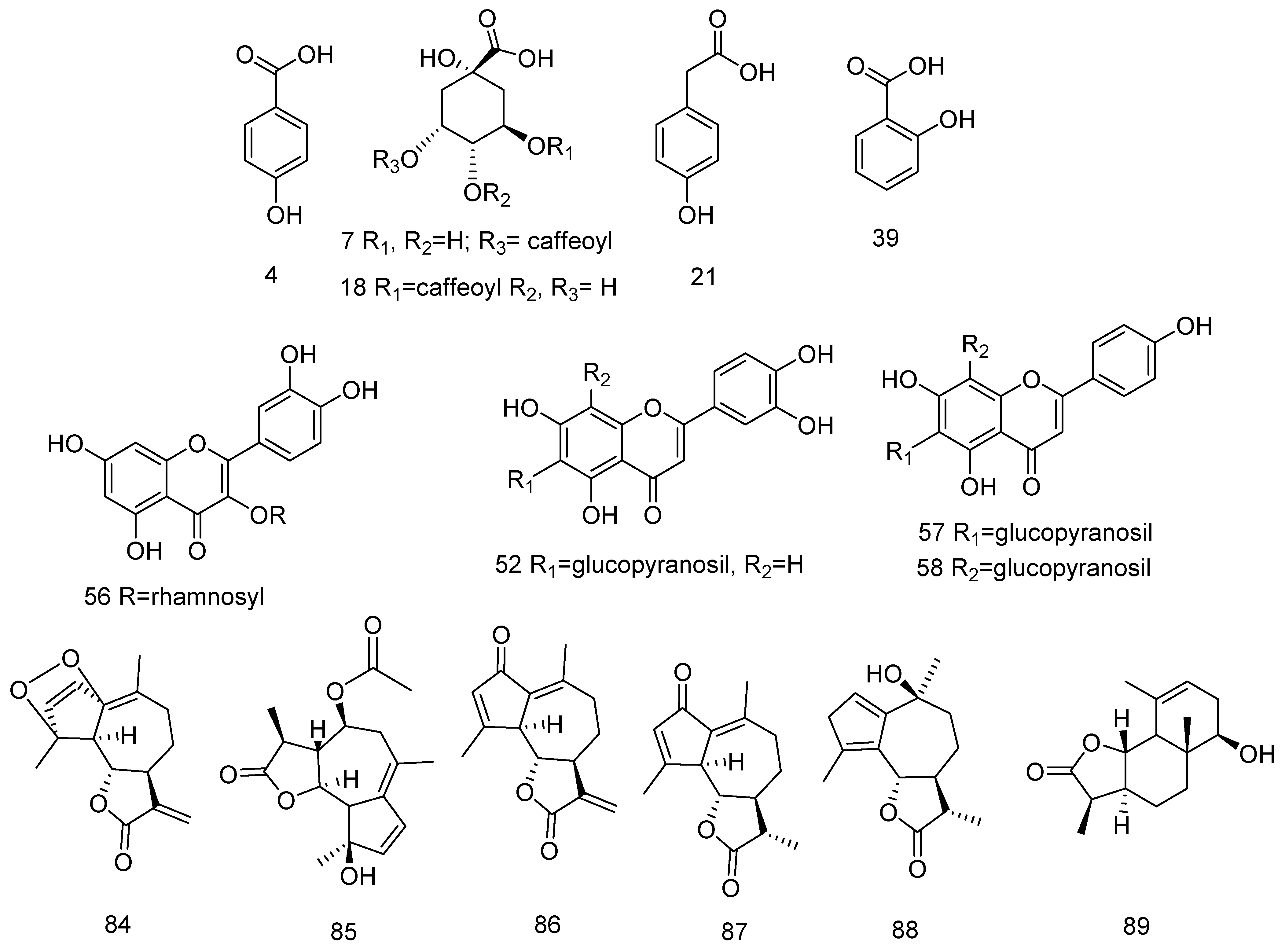
3.1.1. Hydroxybenzoic, Hydroxycinnamic and their Glycosides, and Sugar Esters
3.1.2. Acylquinic Acids
3.1.3. Flavonoids
C-, C,O- and O-Flavonoid Glycosides
6-Methoxyflavonoids
Sesquiterpene Lactones (STLs)
Fatty Acids Amides
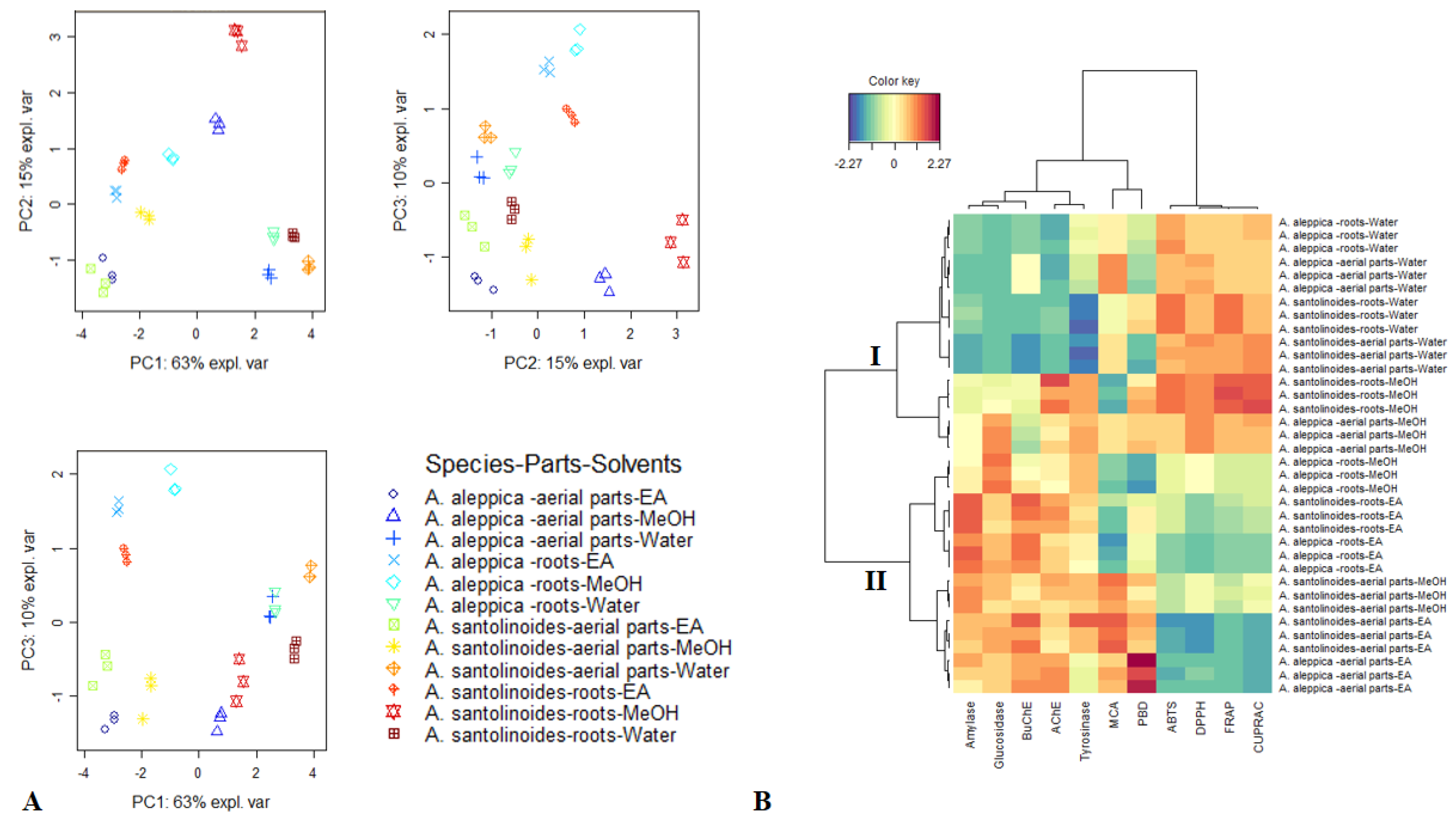
3.2. Antioxidant Effects
3.3. Enzyme Inhibitory Effects
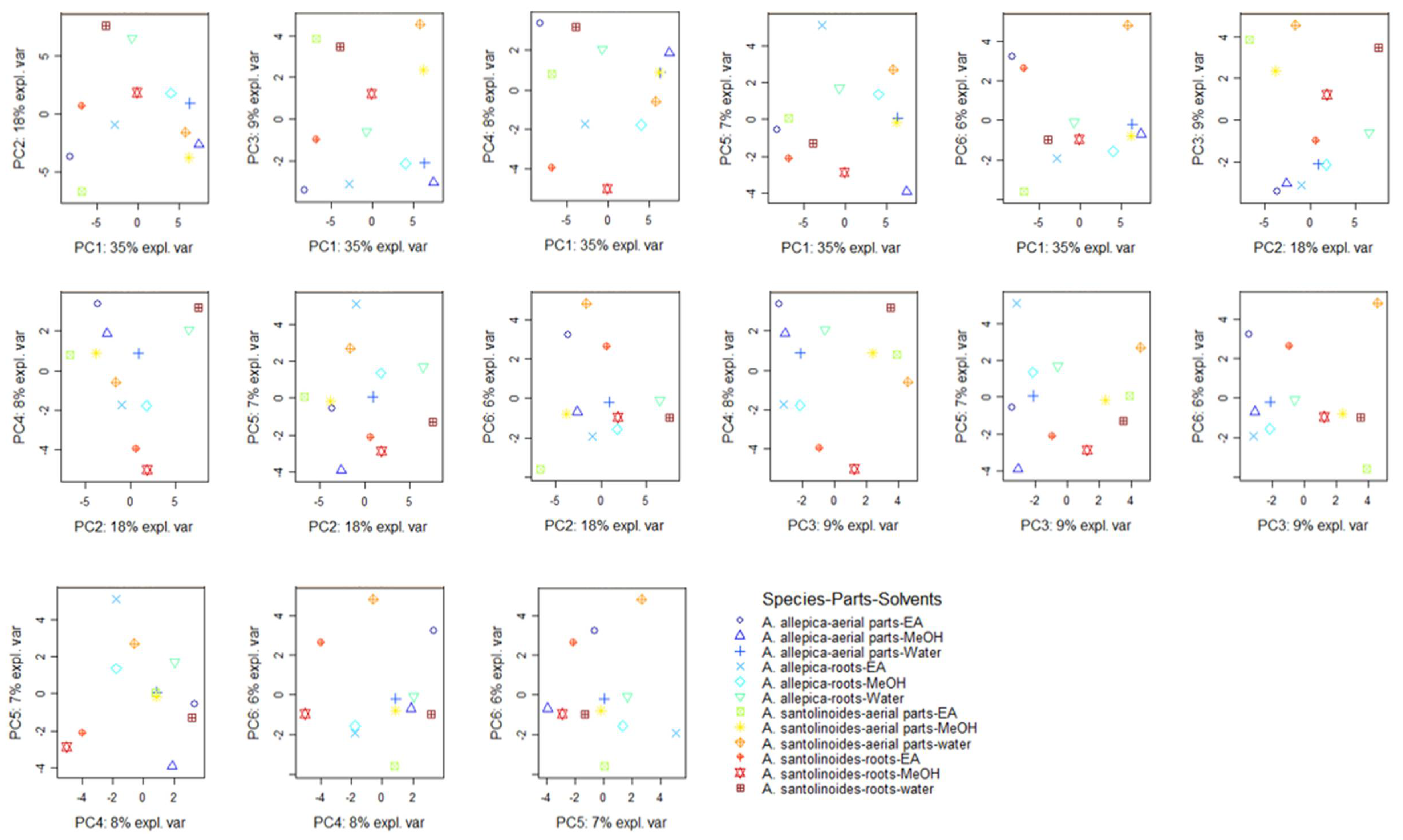
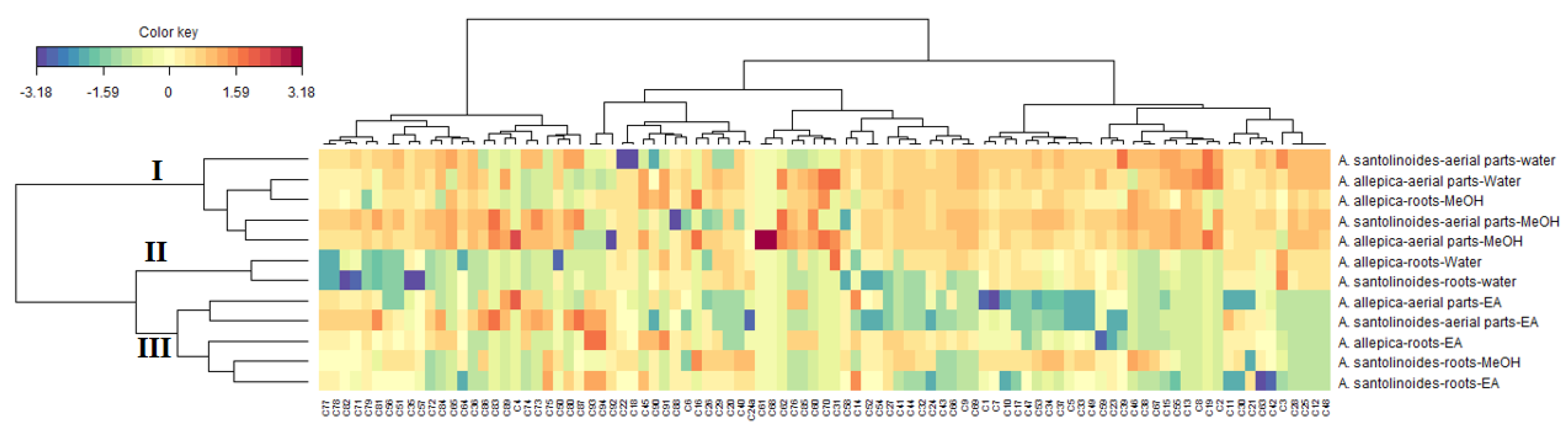
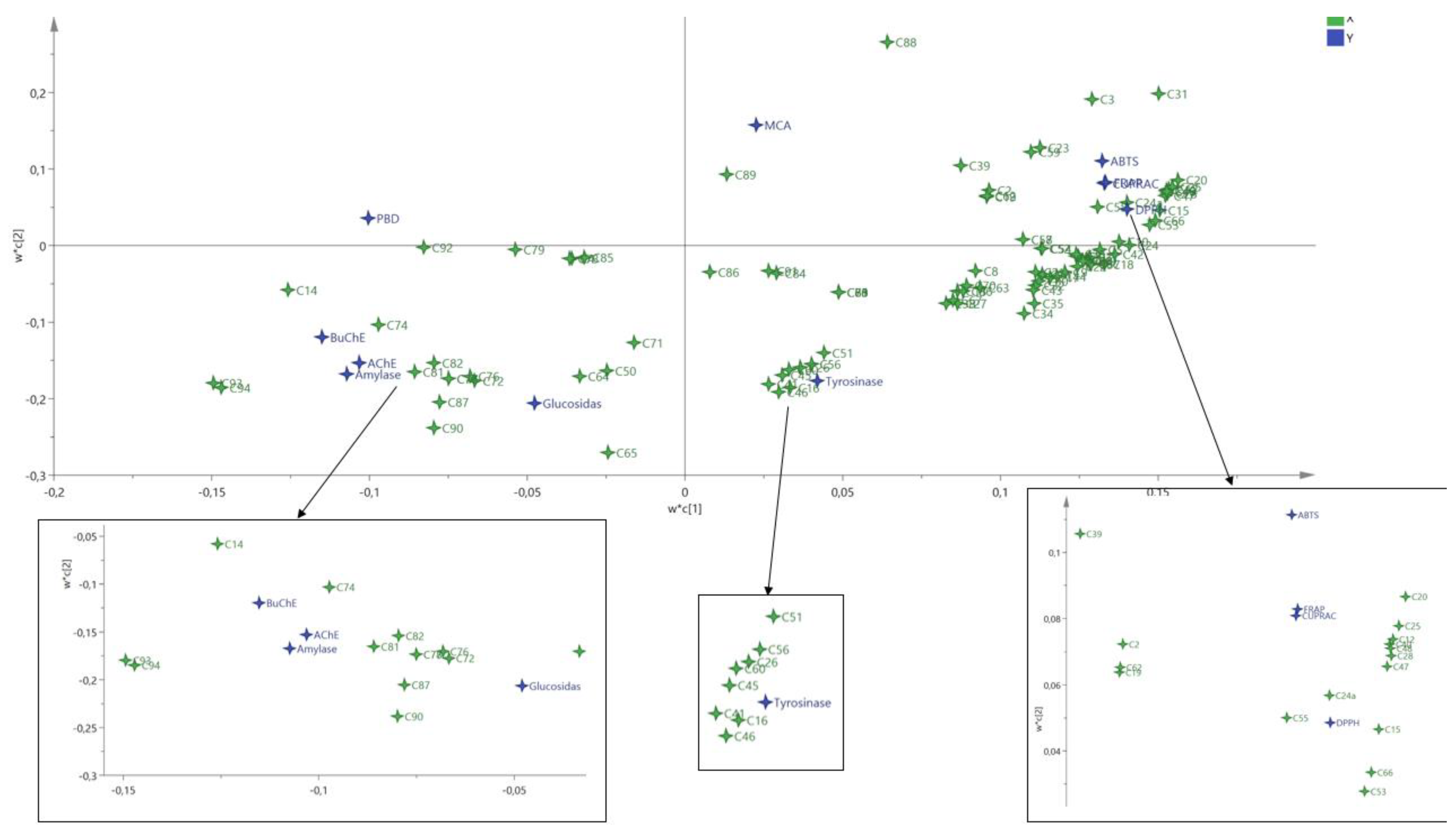
3.4. Data Mining
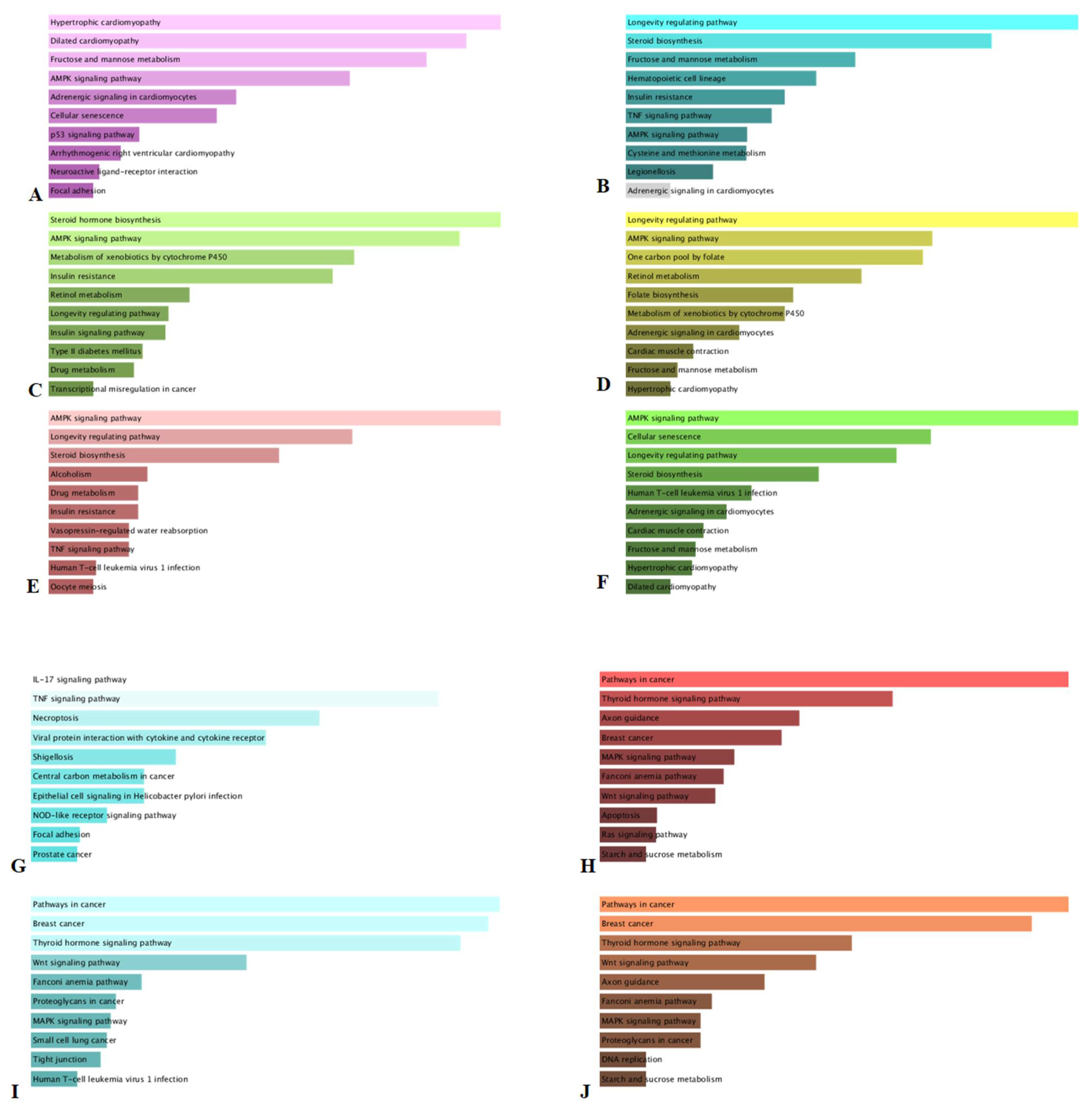
3.5. KEGG Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cam, M.E.; Cesur, S.; Taskin, T.; Erdemir, G.; Kuruca, D.S.; Sahin, Y.M.; Kabasakal, L.; Gunduz, O. Fabrication, characterization and fibroblast proliferative activity of electrospun Achillea lycaonica-loaded nanofibrous mats. Eur. Polym. J. 2019, 120, 109239. [Google Scholar] [CrossRef]
- Kaczorová, D.; Karalija, E.; Dahija, S.; Bešta-Gajević, R.; Parić, A.; Ćavar Zeljković, S. Influence of extraction solvent on the phenolic profile and bioactivity of two Achillea species. Molecules 2021, 26, 1601. [Google Scholar] [CrossRef] [PubMed]
- Kuropka, G.; Glombitza, K.W. Further polyenic and polyynic carboxamides and sesamin from Achillea ptarmica. Planta Med. 1987, 53, 440–442. [Google Scholar] [CrossRef]
- Yassa, N.; Saeidnia, S.; Pirouzi, R.; Shafiee, A. Three phenolic glycosides and immunological properties of Achillea millefolium from Iran, population of Golestan. DARU J. Pharm. Sci. 2007, 1, 49–52. [Google Scholar]
- Glasl, S.; Presser, A.; Gunbilig, D.; Werner, I.; Narantuya, S.; Haslinger, E.; Jurenitsch, J.; Kubelka, W. Highly hydroxylated guaianolides of Achillea asiatica and Middle European Achillea species. Phytochemistry 2001, 58, 1189–1194. [Google Scholar] [CrossRef]
- Trifunovic, S.; Vlatka, V.; Tešević, V.; Dejan, D.; Slobodan, M. Lignans from the plant species Achillea lingulata. J. Serb. Chem. Soc. 2003, 68, 277–280. [Google Scholar] [CrossRef]
- Manayi, A.; Kurepaz-Mahmoodabadi, M.; Gohari, A.R.; Ajani, Y.; Saeidnia, S. Presence of phthalate derivatives in the essential oils of a medicinal plant Achillea tenuifolia. DARU J. Pharm. Sci. 2014, 22, 78. [Google Scholar] [CrossRef] [Green Version]
- Shah, R.M.; Patel, T.; Tettamanzi, C.M.; Rajan, J.; Shah, M.; Peethambaran, B. Isolation of a novel piperidide from Achillea moonshine using bioactivity guided fractionation for the treatment of acne. J. Med. Plant. Res. 2016, 10, 495–504. [Google Scholar]
- Farhadi, N.; Babaei, K.; Farsaraei, S.; Moghaddam, M.; Pirbalouti, A.G. Changes in essential oil compositions, total phenol, flavonoids and antioxidant capacity of Achillea millefolium at different growth stages. Ind. Crop. Prod. 2020, 152, 6. [Google Scholar] [CrossRef]
- Konovalov, D.; Chelombit’ko, V. Sesquiterpene lactones from Achillea millefolium. Chem. Nat. Compd. 1991, 27, 640–641. [Google Scholar] [CrossRef]
- Li, H.L.; Li, J.; Liu, M.Q.; Xie, R.R.; Zang, Y.; Li, J.; Aisa, H.A. Guaianolide sesquiterpene lactones from Achillea millefolium L. Phytochemistry 2021, 186, 9. [Google Scholar] [CrossRef] [PubMed]
- Konovalov, D. Polyacetylene compounds of plants of the Asteraceae Family (Review). Pharm. Chem. J. 2015, 48, 36–53. [Google Scholar] [CrossRef]
- Todorova, M.N.; Tsankova, E.T.; Mustakerova, E.I. Spirodepressolide: An unusual bis-norsesquiterpene lactone from Achillea depressa Janka. Nat. Prod. Res. 2004, 18, 461–464. [Google Scholar] [CrossRef]
- Asghari, B.; Mafakheri, S.; Zengin, G.; Dinparast, L.; Bahadori, M.B. In-depth study of phytochemical composition, antioxidant activity, enzyme inhibitory and antiproliferative properties of Achillea filipendulina: A good candidate for designing biologically-active food products. J. Food Meas. Charact. 2020, 14, 2196–2208. [Google Scholar] [CrossRef]
- Si, X.T.; Zhang, M.L.; Shi, Q.W.; Kiyota, H. Chemical constituents of the plants in the genus Achillea. Chem. Biodiv. 2006, 3, 1163–1180. [Google Scholar] [CrossRef]
- Mohammadhosseini, M.; Sarker, S.D.; Akbarzadeh, A. Chemical composition of the essential oils and extracts of Achillea species and their biological activities: A review. J. Ethnopharmacol. 2017, 199, 257–315. [Google Scholar] [CrossRef]
- Ali, N.; Shah, S.W.; Ahmed, G.; Shah, I.; Shoaib, M.; Junaid, M.; Ali, W. Acute toxicity and antispasmodic activities of Achillea wilhelmsii C. Koch. Pak. J. Pharm. Sci. 2014, 27, 309–315. [Google Scholar] [PubMed]
- Ashtiani, M.; Nabatchian, F.; Galavi, H.R.; Saravani, R.; Farajian-Mashhadi, F.; Salimi, S. Effect of Achillea wilhelmsii extract on expression of the human telomerase reverse transcriptase mRNA in the PC3 prostate cancer cell line. Biomed. Rep. 2017, 7, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Majnooni, M.B.; Mohammadi-Farani, A.; Gholivand, M.B.; Nikbakht, M.R.; Bahrami, G.R. Chemical composition and anxiolytic evaluation of Achillea wilhelmsii C. Koch essential oil in rat. Res. Pharm. Sci. 2013, 8, 269–275. [Google Scholar] [PubMed]
- Asgary, S.; Naderi, G.H.; Sarrafzadegan, N.; Mohammadifard, N.; Mostafavi, S.; Vakili, R. Antihypertensive and antihyperlipidemic effects of Achillea wilhelmsii. Drugs Exp. Clin. Res. 2000, 26, 89–93. [Google Scholar]
- Barış, D.; Kızıl, M.; Aytekin, Ç.; Kızıl, G.; Yavuz, M.; Çeken, B.; Ertekin, A.S. In vitro antimicrobial and antioxidant activity of ethanol extract of three Hypericum and three Achillea species from Turkey. Int. J. Food Prop. 2011, 14, 339–355. [Google Scholar] [CrossRef]
- Marucci, G.; Buccioni, M.; Ben, D.D.; Lambertucci, C.; Volpini, R.; Amenta, F. Efficacy of acetylcholinesterase inhibitors in Alzheimer’s disease. Neuropharmacology 2021, 190, 108352. [Google Scholar] [CrossRef] [PubMed]
- Papoutsis, K.; Zhang, J.; Bowyer, M.C.; Brunton, N.; Gibney, E.R.; Lyng, J. Fruit, vegetables, and mushrooms for the preparation of extracts with α-amylase and α-glucosidase inhibition properties: A review. Food Chem. 2021, 338, 128119. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Biswas, R.; Sharma, A.; Banerjee, S.; Biswas, S.; Katiyar, C.K. Validation of medicinal herbs for anti-tyrosinase potential. J. Herb. Med. 2018, 14, 1–16. [Google Scholar] [CrossRef]
- Bernardo, J.; Malheiro, I.; Videira, R.A.; Valentão, P.; Santos, A.C.; Veiga, F.; Andrade, P.B. Trichilia catigua and Turnera diffusa extracts: In vitro inhibition of tyrosinase, antiglycation activity and effects on enzymes and pathways engaged in the neuroinflammatory process. J. Ethnopharmacol. 2021, 271, 113865. [Google Scholar] [CrossRef] [PubMed]
- Spínola, V.; Castilho, P.C. Evaluation of Asteraceae herbal extracts in the management of diabetes and obesity. Contribution of caffeoylquinic acids on the inhibition of digestive enzymes activity and formation of advanced glycation end-products (in vitro). Phytochemistry 2017, 143, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Suliman, S.; Yagi, S.; Elbashir, A.A.; Mohammed, I.; Hussein, A.; Ak, G.; Zengin, G.; Orlando, G.; Ferrante, C. Phenolic profile, enzyme inhibition and antioxidant activities and bioinformatics analysis of leaf and stem bark of Ficus sycomorus L. Process Biochem. 2021, 101, 169–178. [Google Scholar] [CrossRef]
- Slinkard, K.; Singleton, V.L. Total phenol analysis: Automation and comparison with manual methods. Am. J. Enol. Viticul. 1977, 28, 49–55. [Google Scholar]
- Zengin, G.; Nithiyanantham, S.; Locatelli, M.; Ceylan, R.; Uysal, S.; Aktumsek, A.; Selvi, P.K.; Maskovic, P. Screening of in vitro antioxidant and enzyme inhibitory activities of different extracts from two uninvestigated wild plants: Centranthus longiflorus subsp. longiflorus and Cerinthe minor subsp. auriculata. Eur. J. Integ. Med. 2016, 8, 286–292. [Google Scholar] [CrossRef]
- Uysal, S.; Zengin, G.; Locatelli, M.; Bahadori, M.B.; Mocan, A.; Bellagamba, G.; De Luca, E.; Mollica, A.; Aktumsek, A. Cytotoxic and enzyme inhibitory potential of two Potentilla species (P. speciosa L. and P. reptans Willd.) and their chemical composition. Front. Pharmacol. 2017, 8, 290. [Google Scholar] [CrossRef] [PubMed]
- Zengin, G. A study on in vitro enzyme inhibitory properties of Asphodeline anatolica: New sources of natural inhibitors for public health problems. Ind. Crop. Prod. 2016, 83, 39–43. [Google Scholar] [CrossRef]
- Lagunin, A.; Ivanov, S.; Rudik, A.; Filimonov, D.; Poroikov, V. DIGEP-Pred: Web service for in silico prediction of drug-induced gene expression profiles based on structural formula. Bioinformatics 2013, 29, 2062–2063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene Set Knowledge Discovery with Enrichr. Curr. Prot. 2021, 1, e90. [Google Scholar]
- Kaiser, H.F. The application of electronic computers to factor analysis. Educ. Psychocol. Meas. 1960, 20, 141–151. [Google Scholar] [CrossRef]
- Ak, G.; Gevrenova, R.; Sinan, K.I.; Zengin, G.; Zheleva, D.; Mahomoodally, M.F.; Senkardes, I.; Brunetti, L.; Leone, S.; Di Simone, S.C. Tanacetum vulgare L. (Tansy) as an effective bioresource with promising pharmacological effects from natural arsenal. Food Chem. Toxicol. 2021, 153, 112268. [Google Scholar] [CrossRef]
- Gevrenova, R.; Zheleva-Dimitrova, D.; Balabanova, V.; Voynikov, Y.; Sinan, K.I.; Mahomoodally, M.F.; Zengin, G. Integrated phytochemistry, bio-functional potential and multivariate analysis of Tanacetum macrophyllum (Waldst. & Kit.) Sch. Bip. and Telekia speciosa (Schreb.) Baumg.(Asteraceae). Ind. Crop. Prod. 2020, 155, 112817. [Google Scholar]
- Zheleva-Dimitrova, D.; Sinan, K.I.; Etienne, O.K.; Zengin, G.; Gevrenova, R.; Mahomoodally, M.F.; Lobine, D.; Mollica, A. Chemical composition and biological properties of Synedrella nodiflora (L.) Gaertn: A comparative investigation of different extraction methods. Process. Biochem. 2020, 96, 202–212. [Google Scholar] [CrossRef]
- Zheleva-Dimitrova, D.; Zengin, G.; Balabanova, V.; Voynikov, Y.; Lozanov, V.; Lazarova, I.; Gevrenova, R. Chemical characterization with in vitro biological activities of Gypsophila species. J. Pharm. Biomed. Anal. 2018, 155, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Ferreres, F.; Gil-Izquierdo, A.; Andrade, P.B.; Valentão, P.; Tomás-Barberán, F. Characterization of C-glycosyl flavones O-glycosylated by liquid chromatography—Tandem mass spectrometry. J. Chrom. A 2007, 1161, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.; Ran, L.; Yang, C.; Xu, M.; Yi, L. Integrated strategy for identifying minor components in complex samples combining mass defect, diagnostic ions and neutral loss information based on ultra-performance liquid chromatography-high resolution mass spectrometry platform: Folium Artemisiae Argyi as a case study. J. Chrom. A 2018, 1550, 35–44. [Google Scholar]
- Bakr, R.O.; Arafa, R.K.; Al-Abd, A.M.; Elshishtawy, H.M. Phenolics of Achillea fragrantissima growing in Egypt and its cytotoxic activity. J. Med. Plant Res. 2014, 8, 763–771. [Google Scholar]
- Abd-Alla, H.I.; Shalaby, N.M.; Hamed, M.A.; El-Rigal, N.S.; Al-Ghamdi, S.N.; Bouajila, J. Phytochemical composition, protective and therapeutic effect on gastric ulcer and α-amylase inhibitory activity of Achillea biebersteinii Afan. Arch. Pharm. Res. 2016, 39, 10–20. [Google Scholar] [CrossRef]
- Yao, D.; Li, Z.; Huo, C.; Wang, Y.; Wu, Y.; Zhang, M.; Li, L.; Shi, Q.; Kiyota, H.; Shi, X. Identification of in vitro and in vivo metabolites of alantolactone by UPLC-TOF-MS/MS. J. Chrom. B 2016, 1033, 250–260. [Google Scholar] [CrossRef]
- Kastner, U.; Breuer, J.; Glasl, S.; Baumann, A.; Robien, W.; Jurenitsch, J.; Rücker, G.; Kubelka, W. Guaianolide-endoperoxide and monoterpene-hydroperoxides from Achillea nobilis. Planta Med. 1995, 61, 83–85. [Google Scholar] [CrossRef]
- Todorova, M.; Trendafilova, A.; Mikhova, B.; Vitkova, A.; Duddeck, H. Chemotypes in Achillea collina based on sesquiterpene lactone profile. Phytochemistry 2007, 68, 1722–1730. [Google Scholar] [CrossRef]
- Castillo-Peinado, L.; López-Bascón, M.; Mena-Bravo, A.; de Castro, M.L.; Priego-Capote, F. Determination of primary fatty acid amides in different biological fluids by LC–MS/MS in MRM mode with synthetic deuterated standards: Influence of biofluid matrix on sample preparation. Talanta 2019, 193, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Semaming, Y.; Pannengpetch, P.; Chattipakorn, S.C.; Chattipakorn, N. Pharmacological properties of protocatechuic Acid and its potential roles as complementary medicine. Evid.-Based Complement. Altern. Med. 2015, 2015, 593902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakkar, S.; Bais, S. A review on protocatechuic acid and its pharmacological potential. ISRN Pharmacol. 2014, 2014, 952943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Li, K.; Xie, H.; Xie, Y.; Li, Y.; Zhao, X.; Jiang, X.; Chen, D. Antioxidant and cytoprotective effects of the Di-O-Caffeoylquinic acid family: The mechanism, structure-activity relationship, and conformational effect. Molecules 2018, 23, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andjelković, M.; Van Camp, J.; De Meulenaer, B.; Depaemelaere, G.; Socaciu, C.; Verloo, M.; Verhe, R. Iron-chelation properties of phenolic acids bearing catechol and galloyl groups. Food Chem. 2006, 98, 23–31. [Google Scholar] [CrossRef]
- Hosseini, M.; Harandizadeh, F.; Niazamand, S.; Soukhtanloo, M.; Mahmoudabady, M. Antioxidant effect of Achillea wilhelmsii extract on pentylenetetrazole (seizure model)—Induced oxidative brain damage in Wistar rats. Ind. J. Physiol. Pharmacol. 2013, 57, 418–424. [Google Scholar]
- Saeidi, K.; Moosavi, M.; Lorigooini, Z.; Maggi, F. Chemical characterization of the essential oil compositions and antioxidant activity from Iranian populations of Achillea wilhelmsii K.Koch. Ind. Crop. Prod. 2018, 112, 274–280. [Google Scholar] [CrossRef]
- Rasool, M.; Malik, A.; Waquar, S.; Tul-Ain, Q.; Jafar, T.H.; Rasool, R.; Kalsoom, A.; Ghafoor, M.A.; Sehgal, S.A.; Gauthaman, K.; et al. In-silico characterization and in-vivo validation of albiziasaponin-a, iso-orientin, and salvadorin using a rat model of Alzheimer’s disease. Front. Pharmacol. 2018, 9, 730. [Google Scholar] [CrossRef] [PubMed]
- Sheeja Malar, D.; Beema Shafreen, R.; Karutha Pandian, S.; Pandima Devi, K. Cholinesterase inhibitory, anti-amyloidogenic and neuroprotective effect of the medicinal plant Grewia tiliaefolia—An in vitro and in silico study. Pharm. Biol. 2017, 55, 381–393. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.R.; Men, X.; Gao, X.H.; Liu, L.B.; Fan, H.Q.; Xia, X.H.; Wang, Q.A. Discovery of potent and selective acetylcholinesterase (AChE) inhibitors: Acacetin 7-O-methyl ether Mannich base derivatives synthesised from easy access natural product naringin. Nat. Prod. Res. 2018, 32, 743–747. [Google Scholar] [CrossRef] [PubMed]
- Toublet, F.-X.; Lalut, J.; Hatat, B.; Lecoutey, C.; Davis, A.; Since, M.; Corvaisier, S.; Freret, T.; Sopková-de Oliveira Santos, J.; Claeysen, S.; et al. Pleiotropic prodrugs: Design of a dual butyrylcholinesterase inhibitor and 5-HT6 receptor antagonist with therapeutic interest in Alzheimer’s disease. Eur. J. Med. Chem. 2021, 210, 113059. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.-J.; Yen, F.-L.; Chen, P.-C.; Wang, M.-C.; Lin, C.-N.; Lee, C.-W.; Ko, H.-H. HPLC-Fingerprints and antioxidant constituents of Phyla nodiflora. Sci. World J. 2014, 2014, 528653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihaylova, M.M.; Shaw, R.J. The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nat. Cell Biol. 2011, 13, 1016–1023. [Google Scholar] [CrossRef]
- Umezawa, S.; Higurashi, T.; Nakajima, A. AMPK: Therapeutic target for diabetes and cancer prevention. Curr. Pharm. Des. 2017, 23, 3629–3644. [Google Scholar] [CrossRef]
- Babaei, G.; Aliarab, A.; Abroon, S.; Rasmi, Y.; Aziz, S.G.-G. Application of sesquiterpene lactone: A new promising way for cancer therapy based on anticancer activity. Biomed. Pharmacother. 2018, 106, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Wang, W.; Wang, Y.; Li, J. Anti-proliferative and apoptosis-inducing effects of matricin on human non-small cell lung cancer H1299 cells via MAPK pathway activation. Eur. J. Inflamm. 2020, 18, 2058739220942335. [Google Scholar] [CrossRef]
- Shang, Y.; Li, X.F.; Jin, M.J.; Li, Y.; Wu, Y.L.; Jin, Q.; Zhang, Y.; Li, X.; Jiang, M.; Cui, B.W.; et al. Leucodin attenuates inflammatory response in macrophages and lipid accumulation in steatotic hepatocytes via P2x7 receptor pathway: A potential role in alcoholic liver disease. Biomed. Pharmacother. 2018, 107, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Ye, T.; Su, J.; Huang, C.; Yu, D.; Dai, S.; Huang, X.; Chen, B.; Zhou, M. Isoorientin induces apoptosis, decreases invasiveness, and downregulates VEGF secretion by activating AMPK signaling in pancreatic cancer cells. OncoTargets Ther. 2016, 9, 7481–7492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, M.; Min, J.W.; Kong, W.L.; He, X.H.; Li, J.X.; Peng, B.W. A review on the pharmacological effects of vitexin and isovitexin. Fitoterapia 2016, 115, 74–85. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Parts | Solvents | Yields | TPC (mg GAE/g) | TFC (mg RE/g) | PBD (mmol TE/g) |
|---|---|---|---|---|---|---|
| A. aleppica | Aerial parts | EA | 4.61 | 20.77 ± 0.83 e | 13.23 ± 0.52 c | 2.33 ± 0.09 a |
| MeOH | 10.01 | 41.41 ± 0.88 b | 49.18 ± 0.98 a | 1.92 ± 0.04 b | ||
| Water | 9.88 | 36.56 ± 0.01 c | 16.62 ± 0.17 b | 1.45 ± 0.05 d | ||
| Roots | EA | 1.88 | 22.41 ± 0.59 d | 3.95 ± 0.11 e | 1.65 ± 0.04 c | |
| MeOH | 2.85 | 23.83 ± 0.24 d | 5.93 ± 0.04 d | 1.27 ± 0.03 e | ||
| Water | 4.58 | 43.24 ± 0.19 a | 4.12 ± 0.05 e | 1.57 ± 0.04 cd | ||
| A. santolinoides | Aerial parts | EA | 7.40 | 20.69 ± 0.32 f | 19.58 ± 0.32 a | 1.95 ± 0.05 a |
| MeOH | 16.04 | 32.20 ± 0.22 d | 8.42 ± 0.63 d | 1.90 ± 0.09 ab | ||
| Water | 19.05 | 44.97 ± 0.49 c | 18.08 ± 0.23 b | 1.33 ± 0.03 c | ||
| Roots | EA | 1.41 | 26.27 ± 0.90 e | 5.07 ± 0.23 e | 1.73 ± 0.10 b | |
| MeOH | 6.92 | 52.07 ± 1.58 a | 11.09 ± 0.18 c | 1.93 ± 0.12 ab | ||
| Water | 4.91 | 47.39 ± 0.05 b | 3.59 ± 0.27 f | 1.88 ± 0.04 ab |
| No. | Identified/Tentatively Annotated Compound | Molecular Formula | Exact Mass [M − H]− | Fragmentation Pattern In (−) ESI-MS/MS | tR (Min) | Δ ppm | Distribution |
|---|---|---|---|---|---|---|---|
| Hydroxybenzoic, hydroxycinnamic and acylquinic acids, and derivatives | |||||||
| 1 | protocatechuic acid-O-hexoside | C13H16O9 | 315.0722 | 315.0725 (100), 153.0179 (30.5), 109.0279 (99.3) | 1.71 | 0.385 | 2,3,4,5,6,7,8,9,10,11,12 |
| 2 | caffeoylgluconic acid | C15H18O10 | 357.0827 | 357.0827 (8.1), 195.0503 (100), 179.0340 (27.2), 177.0397 (18.2), 135.0440 (25.0), 87.0073 (3.6), 59.0121 (11.1) | 2.01 | −0.020 | 2,3,8,9 |
| 3 | protocatechuic acid a | C7H6O4 | 153.0182 | 153.0180 (14.6), 123.0435 (100), 109.0278 (40.8) | 2.03 | −1.362 | 1,2,3,4,5,6,7,8,9,10,11,12 |
| 4 | p-hydroxyphenylacetic acid-O-hexoside | C14H18O8 | 313.0929 | 313.0923 (2.7), 151.0387 (100), 123.0071 (0.9) | 2.67 | −0.591 | 1,2 |
| 5 | protocatechuic acid-O-hexoside isomer | C13H16O9 | 315.0722 | 315.0723 (100), 153.0180 (60.3), 123.0437 (17.1), 109.0279 (75.9) | 2.14 | 0.145 | 2,3,4,5,6,8,9,10,11,12 |
| 6 | syringic acid 4-O-hexoside | C15H20O10 | 359.0984 | 359.0984 (9.1), 197.0445 (100), 182.0210 (19.2), 166.9974 (7.6), 153.0544 (14.8), 138.0307 (28.5), 123.0072 (32.0) | 2.28 | −0.010 | 1,2,3,4,5,6,9,10 |
| 7 | neochlorogenic (3-caffeoylquinic) acid a | C16H18O9 | 353.0867 | 353.0879 (42.4), 191.0551 (100), 179.0339 (60.4), 173.0444 (3.7), 161.0236 (4.2), 135.0437 (53.1), 127.0387 (2.4), 93.0331 (4.9), 85.0277 (9.9) | 2.31 | 0.115 | 2,3,4,5,6,7,8,9,10,11,12 |
| 8 | caffeoylgluconic acid isomer | C15H18O10 | 357.0827 | 357.0810 (4.8), 195.0500 (72.1), 179.0338 (100), 177.0395 (7.3), 161.0234 (1.1), 135.0437 (77.2), 129.0177 (2.2), 87.0070 (5.6), 59.0124 (2.4) | 2.40 | −1.730 | 2,3,5,8,9 |
| 9 | caffeic acid-O-hexoside | C15H18O9 | 341.0867 | 341.0880 (5.0), 179.0338 (100), 135.0436 (62.0) | 2.40 | 0.195 | 2,3,4,5,6,8,9 |
| 10 | gentisic acid-O-hexoside | C13H16O9 | 315.0722 | 315.0724 (33.5), 153.0180 (70.9), 135.0072 (4.3), 109.0279 (100), 91.0171 (0.4) | 2.58 | 0.205 | 2,3,4,5,6,7,8,9,11,12 |
| 11 | vanillic acid 4-O-hexoside | C14H18O9 | 329.0878 | 329.0878 (27.1), 197.0446 (100), 182.0210 (15.5), 167.0335 (5.5), 153.0544 (28.7), 123.0073 (19.0), | 2.69 | -0.035 | 2,3,4,5,6,7,8,9,11,12 |
| 12 | caffeoylgluconic acid isomer | C15H18O10 | 357.0827 | 357.0828 (23.9), 339.0726 (11.2), 195.0500 (100), 179.0339 (18.9), 177.0392 (16.9), 161.0235 (3.4), 135.0437 (22.5), 129.0174 (9.7), 87.0071 (10.6), 59.0124 (1.4) | 2.81 | 0.044 | 2,3,5,6,8,9,12 |
| 13 | O-caffeoyl hexose isomer | C15H18O9 | 341.0867 | 341.0869 (23.7), 281.0665 (94.7), 251.0557(54.2), 221.0448 (44.0), 179.0339 (100), 161.0231 (56.9), 135.0437 (72.4) | 2.82 | −0.955 | 2,5,8,9 |
| 14 | 4-hydroxybenzoic acid a | C7H6O3 | 137.0244 | 137.0229 (12.6), 108.0208 (0.1), 93.0329 (100) | 2.86 | −1.527 | 1,2,3,4,5,6,7,8,9,10,11,12 |
| 15 | 4-hydroxybenzoic acid-hexoside | C13H16O8 | 299.0772 | 299.0773 (1.5), 137.0230 (100), 93.0330 (54.3) | 3.00 | 0.029 | 1,2,3,4,5,6,7,8,9,10,11,12 |
| 16 | p-hydroxyphenylacetic acid O-hexoside | C14H18O8 | 313.0929 | 313.0932 (13.4), 151.0386 (100), 123.0070 (0.9) | 3.00 | 0.309 | 2,5,11 |
| 17 | quinic acid | C7H12O6 | 191.0561 | 191.0550 (100), 173.0444 (2.0), 155.0332 (0.3), 127.0386 (4.0), 111.0436 (1.6), 93.0330 (6.2), 85.0279 (19.1) | 3.16 | −1.101 | 2,3,4,5,6,8,9,11,12 |
| 18 | chlorogenic (5-caffeoylquinic) acid a | C16H18O9 | 353.0867 | 353.0857 (1.9), 191.0550 (100), 161.0230 (1.5), 93.0331 (1.5), 85.0278 (8.8) | 3.19 | 0.835 | 1,2,3,4,5,6,7,8,10,11,12 |
| 19 | p-coumaric acid a | C9H8O3 | 163.0389 | 163.0385 (12.4), 135.0438 (4.2), 119.0487 (100) | 3.35 | −1.527 | 2,3,9 |
| 20 | 3-feruloylquinic acid b | C17H20O9 | 367.1035 | 367.1035 (22.0), 193.0496 (100), 191.0550 (2.6), 173.0443 (3.9), 134.0358 (64.7), 93.0329 (1.4), 85.0281 (0.9) | 3.44 | −0.005 | 2,3,5,6,8,10,11,12 |
| 21 | p-hydroxyphenylacetic acid a | C8H8O3 | 151.0401 | 151.0387 (100), 107.0486 (1.4), 136.0154 (1.5), 123.0072 (4.2) | 3.48 | 1.397 | 2,3,4,5,6,7,8,9,10,12 |
| 22 | caffeic acid a | C9H8O4 | 179.0338 | 179.0339 (3.6), 135.0437 (100), 151.0754 (2.4), 107.0489 (1.6) | 3.56 | −1.092 | 1,2,3,4,5,6,7,8,10,11,12 |
| 23 | gentisic acid a | C7H6O4 | 153.0182 | 153.0180 (73.8), 135.0073 (31.4), 109.0279 (100), 91.0173 (6.3) | 3.87 | −1.372 | 1,23,5,6,8,9,10,11,12 |
| 24 | 5-p-coumaroylquinic acid | C16H18O8 | 337.0929 | 337.0933 (9.3), 191.0550 (100), 173.0444 (6.8), 163.0388 (6.1), 161.0229 (0.2), 127.0385 (1.2), 119.0487 (5.3), 93.0329 (17.9), 85.0278 (5.1) | 3.96 | 0.369 | 1,2,3,4,5,6,8,9,11,12 |
| 24a | 1,3-dicaffeoylquinic acid | C25H24O12 | 515.1195 | 115.1199 (78.8), 353.0873 (36.6), 335.0779 (10.9), 191.0550 (100), 179.0338 (73.2), 173.0452 (5.6), 161.0232 (9.9), 135.0435 (58.7), 111.0434 (1.7) | 4.13 | 0.401 | 1,2,3,4,5,6,8,9,11,12 |
| 25 | caffeic acid-O-hexoside isomer | C15H18O9 | 341.0867 | 341.0830 (5.5), 179.0335 (6.2), 161.0230 (39.2), 135.0436 (63.6) | 4.34 | −4.765 | 2,3,5,6,8,9,10,12 |
| 26 | 5-feruloylquinic acid | C17H20O9 | 367.1035 | 367.1035 (18.5), 193.0498 (8.3), 191.0552 (100), 173.0444 (24.1), 134.0359 (12.2), 127.0382 (1.0), 111.0436 (5.0), 93.0329 (30.0), 85.0278 (6.0) | 4.42 | −0.015 | 2,3,4,5,7,9,10,11,12 |
| 27 | m-coumaric acid a | C9H8O3 | 163.0389 | 163.0387 (9.0), 135.0434 (1.8), 119.0486 (100) | 4.57 | −1.367 | 2,3,4,5,6,8,9,10,11 |
| 28 | 5-p-coumaroylquinic acid isomer | C16H18O8 | 337.0929 | 337.0934 (7.0), 191.0550 (100), 173.0444 (1.9), 163.0387 (2.2), 127.0385 (2.2), 119.0487 (1.6), 111.0434 (1.3), 93.0332 (5.0), 85.0278 (8.1) | 4.62 | 0.489 | 2,3,5,6,8,9,12 |
| 29 | 4-feruloylquinic acid | C17H20O9 | 367.1035 | 367.1034 (89.4), 193.0497 (9.8), 173.0443 (63.1), 155.0338 (4.1), 134.0358 (21.8), 111.0436 (14.7), 93.0329 (100), 85.0276 (0.5) | 4.66 | −0.055 | 2,3,4,5,6,10,11,12 |
| 30 | vanillic acid a | C8H8O4 | 167.0350 | 167.0338 (100), 152.0101 (27.8), 123.0071 (4.8), 95.0124 (3.4) | 4.79 | −1.232 | 2,3,4,5,6,7,8,9,11,12 |
| 31 | o-coumaric acid a | C9H8O3 | 163.0389 | 163.0387 (19.5), 135.0441 (4.1), 119.0487 (100) | 4.84 | −1.367 | 2,3,6 |
| 32 | vanillic acid-4-O-(6-O-caffeoyl)-hexoside b | C23H24O12 | 491.1195 | 491.1209 (100.0), 323.0774 (23.3), 221.0458 (4.6), 179.0343 (10.6), 167.0338 (16.2), 161.0231 (38.9), 152.0101 (18.3), 135.0437 (14.8), 123.0436 (1.2) | 5.52 | 0.928 | 2,3,4,5,6,8,9,11 |
| 33 | 3,4-dicaffeoylquinic acid a | C25H24O12 | 515.1195 | 515.1179 (94.1), 353.0875 (62.2), 335.0771 (6.7), 299.0573 (13.6), 203.0339 (41.1), 191.0548 (32.6), 179.0339(76.0), 173.0444 (100), 161.0233 (13.6), 135.0437 (77.0), 111.0436 (4.2), 93.0330 (38.4), 85.0278 (3.9) | 5.60 | −1.579 | 2,3,4,5,6,8,9,10,11,12 |
| 34 | 1,5-dicaffeoylquinic acid a | C25H24O12 | 515.1195 | 515.1189 (15.1), 353.0878 (33.0), 335.0774 (2.2), 191.0550 (100), 179.0338(6.2), 173.0446 (3.1), 161.0231 (5.0), 135.0436 (6.6), 127.0382 (1.8), 111.0433 (1.1), 93.0331 (4.5), 85.0278 (7.6) | 5.70 | −0.599 | 1,2,3,4,5,6,7,8,9,10,11,12 |
| 35 | 3,5-dicaffeoylquinic acid a | C25H24O12 | 515.1195 | 515.1204 (22.6), 353.0878 (100), 191.0551 (96.5), 179.0338 (53.1), 173.0441 (5.3), 161.0229 (7.9), 135.0437 (52.7), 111.0433 (1.1), 93.0328 (4.7), 85.0279 (9.1) | 5.86 | 0.921 | 1,2,3,4,5,6,7,8,9,10,11,12 |
| 36 | dicaffeoyl-tetrahydroxy-pentanoic acid | C23H22O12 | 489.1038 | 489.1030 (43.3), 327.0720 (40.6), 165.0392 (100), 179.0341 (17.2), 161.0231 (3.2), | 6.12 | −0.849 | 2,3,5,8,9 |
| 37 | 4,5-dicaffeoylquinic acid a | C25H24O12 | 515.1195 | 515.1204 (84.6), 353.0877 (76.6), 191.0549 (50.2), 179.0338 (72.4), 173.0443 (100), 161.0232 (8.0), 111.0435 (2.0), 93.0330 (27.1), 85.0278 (4.2) | 6.23 | 0.901 | 1,2,3,4,5,6,7,8,9,10,11,12 |
| 38 | shikimic acid | C7H10O5 | 173.0455 | 173.0444 (100), 155.0335 (2.0), 137.0232 (1.4), 127.0390 (0.5), 111.0437 (10.0), 93.0330 (68.4) | 6.23 | −6.453 | 2,3,5,8,9,11 |
| 39 | salicylic acid a | C7H6O3 | 137.0244 | 137.0228 (15.2), 109.0279 (0.7), 93.0330 (100) | 6.29 | −1.467 | 1,2,3,4,5,6,7,8,9,10,11,12 |
| 40 | 3-feruloyl-4-caffeoylquinic acid | C26H26O12 | 529.1351 | 529.1339 (100), 335.075 (9.3), 193.0498 (60.1), 191.0558 (7.6), 179.0336 (29.5), 173.0441 (29.3), 161.0231 (8.7), 135.0434 (32.1), 134.0357 (49.5), 93.0330 (6.8) | 6.49 | −1.299 | 2,3,5,6,9,11,12 |
| 41 | 3-p-coumaroyl-5-caffeoylquinic acid | C25H24O11 | 499.1246 | 499.1388 (44.0), 353.0885 (5.4), 337.0932 (83.6), 191.0553 (31.7), 173.0443 (7.0), 163.0388 (100.0), 135.0429 (1.7), 93.0326 (7.6), 85.0278 (1.8) | 6.53 | 14.205 | 2,3,4,5,8,9,11 |
| 42 | caffeic acid-O-(salicyl)-hexoside | C22H21O11 | 461.1089 | 461.1093 (49.1), 371.0756 (0.5), 341.0656 (1.9), 323.0774 (24.5), 299.0767 (1.6), 179.0340 (5.1), 161.0231 (23.1), 137.0229 (100), 93.0330 (61.3) | 6.56 | 0.405 | 1,2,3,4,5,6,7,8,9,11,12 |
| 43 | 3-feruloyl-5-caffeoylquinic acid | C26H26O12 | 529.1351 | 529.1355 (17.3), 367.1033(3.0), 353.2703 (1.6), 335.0754 (1.5), 193.0496 (100), 191.0554 (8.0), 173.0450 (9.0), 161.0230 (10.50), 134.0359 (74.4), 93.0331 (2.6) | 6.82 | 0.351 | 2,3,4,5,6,8,9,10,12 |
| 44 | 4-p-coumaroyl-5-caffeoylquinic acid | C25H24O11 | 499.1246 | 499.1218 (-0.871), 337.0932 (61.7), 179.0339 (9.0), 173.0442 (100), 163.0390 (21.2), 135.0437 (4.7), 119.0487 (8.7), 111.0437 (2.8) | 6.90 | −2.755 | 2,3,4,5,6,8,9,11 |
| 45 | 1-caffeoyl-3-feruloylquinic acid | C26H26O12 | 529.1351 | 529.1355 (17.3), 367.1033(3.0), 353.2703 (1.6), 335.0754 (1.5), 193.0496 (100), 191.0554 (8.0), 173.0450 (9.0), 161.0230 (10.50), 134.0359 (74.4), 93.0331 (2.6) | 7.23 | 0.651 | 2,4,5,10 |
| 46 | 3,4,5-tricaffeoylquinic acid | C34H30O15 | 677.1512 | 677.1509 (100). 515.1186 (32.4), 353.0879 (40.3), 335.0752 (13.7), 191.0552 (36.9), 179.0338 (70.6), 173.0444 (69.4), 161.0231 (19.8), 135.0437 (74.8), 111.0442 (3.6), 93.0330 (17.5) | 7.80 | −0.253 | 2,5,8,9,11 |
| Flavonoids | |||||||
| 47 | 6, 8-diC-hexosidyl-luteolin | C27H30O16 | 609.1461 | 609.1467 (100), 519.1136 (4.1), 489.1045 (14.6), 471.0941 (0.9), 429.0831 (6.1), 399.0722 (24.4), 369.0617 (26.2), 339.0507 (3.5), 311.0547 (5.2), 175.0387 (1.2), 133.0283 (6.7) | 3.64 | 0.622 | 2,3,5,6,8,9,10,11,12 |
| 48 | O,C-dihexosyl-luteolin | C27H30O16 | 609.1461 | 609.1469 (100), 447.0930 (24.2), 387.0808 (1.1), 369.0595 (1.6), 357.0616 (16.3), 327.0509 (54.5), 299.0557 (10.2), 298.0480 (6.7), 297.0403 (5.5), 175.0386 (1.6), 133.0283 (6.8) | 3.87 | 0.742 | 2,3,5,6,8,9,12 |
| 49 | diC-hexosyl-apigenin | C27H30O15 | 593.1512 | 593.1518 (100), 503.1205 (3.9), 473.1089 (14.9), 455.0996 (1.5), 413.0878 (1.9), 383.0773 (14.7), 353.0667 (31.6), 325.0723 (2.4), 309.0763 (1.8), 297.0769 (12.4), 175.0389 (1.5), 117.0331 (4.4) | 4.03 | 0.597 | 2,3,4,5,6,8,9,10,11,12 |
| 50 | 6-C-hexosyl-8-C-pentosyl-luteolin | C26H28O15 | 579.1355 | 579.1362 (100), 519.1219 (1.4), 489.1044 (10.7), 471.0909 (2.3), 459.0936 (9.1), 441.0836 (4.1), 429.0844 (7.1), 411.0721 (2.6), 399.0720 (26.7), 381.0613 (2.3), 369.0617 (24.6), 339.0504 (4.7), 311.0559 (4.3), 298.0483 (4.5), 175.0390 (0.9), 133.0280 (5.5) | 4.12 | 0.627 | 1,2,3,4,5,7,8,9,10,11,12 |
| 51 | 2”-O-pentosyl-6-C-hexosyl-luteolin | C26H28O15 | 579.1355 | 579.1362 (100), 459.0923 (9.6), 429.0820 (3.7), 399.0729 (1.4), 369.0618 (3.7), 357.0618 (25.8), 327.0514 (48.0), 309.0394 (7.2), 299.0558 (6.5), 298.0485 (17.0), 297.0400 (10.8), 175.0392 (1.4), 133.0278 (9.7) | 4.47 | 0.687 | 2,3,4,5,7,8,9,10,11 |
| 52 | Homoorientin a | C21H20O11 | 447.0933 | 447.0935 (100), 429.0830 (3.6), 411.0712 (0.9), 399.0707 (1.1), 387.0720 (0.5), 381.0609 (0.5), 369.0618 (2.8), 357.0617 (47.4), 327.0511 (68.9), 299.0559 (11.8), 298.0477 (9.1), 297.0403 (12.0), 285.0402 (8.6), 269.0449 (1.8), 199.0391 (1.5), 133.0280 (15.0), 107.0119 (0.7) | 4.53 | 0.225 | 1,2,3,4,5,6,8,9,10,11 |
| 53 | 6-C-hexosyl-8-C-pentosyl apigenin | C26H28O14 | 563.1406 | 563.1412 (100), 503.1214 (3.6), 473.1079 (6.7), 443.0990 (7.5), 425.0868 (2.9), 413.0889 (2.9), 383.0774 (20.1), 365.0665 (2.0), 353.0668 (25.9), 325.0715 (2.1), 324.0595 (0.4), 323.0562 (1.4), 297.0765 (9.3), 283.0611 (2.2), 175.0393 (1.3), 135.0434 (2.0), 117.0330 (3.1) | 4.53 | 0.541 | 1,2,3,4,5,6,7,8,9,10,11,12 |
| 54 | orientin (luteolin-8-C-glucoside) | C21H20O11 | 447.0933 | 447.0935 (89.2), 369.0621 (3.2), 357.0616 (31.4), 327.0511 (100), 299.0560 (10.7), 298.0476 (7.0), 297.0404 (13.5), 285.0397 (6.4), 269.0457 (1.6), 133.03 (19.4), 119.0485 (1.4), 151.022 (0.7), 107.0121 (0.6) | 4.68 | 1,2,4,5,6,8,9,10,11 | |
| 55 | C-hexosyl-C-pentosyl methylluteolin | C27H30O15 | 593.1512 | 593.1520 (100), 503.1203 (9.8), 473.1092 (11.1), 443.0963 (2.7), 425.0835 (1.0), 413.0881 (14.7), 395.0765 (0.6), 383.0775 (24.8), 341.0677 (1.1), 323.0550 (1.1), 313.0683 (1.9), 312.0639 (18.5), 299.0557 (0.5), 298.0476 (2.4), 283.0614 (1.8) | 4.75 | 0.787 | 1,2,3,4,5,6,7,8,9,10,11,12 |
| 56 | Rutin a | C27H30O16 | 609.1461 | 609.1464 (100), 301.0347 (39.7), 300.0274 (70.1), 271.0247 (39.2), 255.0297 (18.2), 243.0294 (9.3), 227.0342 (2.7), 211.0394 (0.9), 178.9977 (3.2), 163.0027 (1.6), 151.0024 (7.1), 121.0278 (1.1), 107.0121 (2.2) | 5.06 | 0.512 | 2,3,4,5,7,8,9,10,11,12 |
| 57 | Vitexin a | C21H20O10 | 431.0984 | 431.0986 (95.1), 341.0666 (0.5), 311.0562 (100), 293.0452 (2.0), 283.0610 (30.3), 117.0330 (15.0) | 5.15 | 0.200 | 1,2,3,4,5,6,7,8,9,10,11,12 |
| 58 | Isovitexin a | C21H20O10 | 431.0984 | 431.0986 (100), 413.0880 (1.7), 341.0666 (32.8), 311.0562 (69.1), 283.0610 (22.1), 269.0447 (4.1), 239.0706 (1.3), 117.0330 (8.7) | 5.30 | 0.200 | 1,2,4,5,6,7,9,10,11 |
| 59 | 2”-O-pentosyl-6-C-hexosyl-methylluteolin | C27H30O15 | 593.1512 | 593.1500 (100), 473.1140 (1.7), (443.0968 (6.1), 383.0750 (7.7), 371.0761 (16.5), 341.0664 (36.4), 323.0566 (20.7), 308.0315 (6.7), 299.0526 (2.7), 298.0486 (15.9) | 5.36 | 0.002 | 1,2,3,5,6,7,8,9,10,11,12 |
| 60 | Luteolin-7-O-glucoside a | C21H20O11 | 447.0933 | 447.0935 (100), 285.0404 (82.4), 133.0283 (11.8) | 5.39 | 0.437 | 2,4,5,8 |
| 61 | chrysoeriol-6-C-hexoside | C22H22O11 | 461.1078 | 461.1095 (100), 371.0774 (24.0), 341.0667 (73.2), 298.0481 (44.3), 296.0324 (0.9), 297.0403 (14.6) | 5.43 | 0.535 | 2 |
| 62 | nepetin-O-hexuronide | C22H20O13 | 491.0832 | 491.0829 (72.9), 315.0511 (100), 300.0275 (54.3), 272.0326 (8.6), 243.0297 (0.9), 227.0347 (0.5), 133.0284 (2.1) | 5.47 | 0.335 | 2,3,8 |
| 63 | 6-methoxykaempferol-O-hexoside | C22H22O12 | 477.1042 | 477.1041 (100), 315.0512 (56.5), 300.0272 (16.4), 299.0197 (18.2), 271.0247 (52.2), 243.0292 (0.7), 227.0344 (0.4), 151.0020 (1.6), 107.0122 (0.3) | 5.48 | 0.251 | 1,2,3,4,5,6,7,8,9,10,11,12 |
| 64 | nepetin-O-hexoside | C22H22O12 | 477.1038 | 477.1033 (100), 315.0486 (29.7), 300.0269 (15.9), 299.0197 (20.5), 271.0244 (3.6), 255.0307 (1.8), 243.0303 (2.6), 227.0344 (3.5), 165.8804 (0.5), 136.9889 (1.7), 133.0279 (10.0) | 5.67 | −0.549 | 1,2,4,5,7,8,9,11,12 |
| 65 | kaempferol-3-O-glucoside a | C21H20O11 | 447.0933 | 447.0935 (100), 285.0397 (22.0), 284.0324 (55.3), 255.0294 (41.8), 227.0341 (35.0), 151.0023 (1.6) | 5.86 | 0.195 | 2,3,4,5,8,9 |
| 66 | isorhamnetin 3-O-glucoside a | C22H22O12 | 477.1042 | 477.1041 (100), 315.0493 (10.1), 314.0433 (49.0), 300.0279 (3.1), 299.0212 (4.6), 271.0245 (18.5), 255.0300 (0.8), 243.0291 (19.6), 227.0347 (2.8), 215.0350 (3.8), 151.0022 (2.42) | 6.02 | 0.251 | 1,2,3,4,5,6,8,9 |
| 67 | apigenin-7-O-glucoside a | C21H20O10 | 431.0984 | 431.0986 (100), 269.0450 (27.7), 151.0019 (1.0), 107.0121 (1.4) | 6.06 | 0.200 | 2,3,5,8,9,11 |
| 68 | cirsiliol-O-hexoside | C23H24O12 | 491.1184 | 491.1198 (0.3), 476.0963 (26.6), 461.0726 (9.7), 329.0664 (5.0), 314.0425 (5.5), 313.0355 (13.1), 299.0197 (4.3), 285.0402 (11.8), 271.0245 (9.6), 243.0292 (9.0), | 6.31 | 0.311 | 2 |
| 69 | chrysoeriol-O-hexuronide | C23H22O12 | 475.0882 | 475.0884 (83.8), 299.0560 (100), 284.0325 (65.6), 256.0373 (6.5), 227.0347 (1.1), 175.0237 (15.3), 151.0024 (3.3), 113.0228 (37.6), 85.0278 (22.7), | 6.34 | 0.181 | 2,3,4,5,6,8,9 |
| 70 | jaceosidin-O-hexuronide | C23H22O13 | 505.0988 | 505.0993 (88.3), 329.0667 (100), 314.0432 (18.7), 299.0197 (36.3), 271.0247 (36.7), 243.0290 (0.6), 227.0342 (0.6), 175.0237 (13.2), 161.0229 (0.6), 113.0227 (34.1), 85.0278 (22.6) | 6.34 | 0.566 | 2,3,5 |
| 71 | Luteolin a | C15H10O6 | 285.0405 | 285.0403 (100), 241.0975 (21.4), 226.075 (8.4) | 7.59 | −0.181 | 1,2,3,4,5,6,7,8,9,10,11 |
| 72 | Quercetin a | C15H10O7 | 301.0354 | 301.0353 (100), 273.0405 (1.5), 178.9975 (22.7), 151.0023 (51.2), 121.0281 (12.7), 107.0123 (13.4) | 7.62 | −0.036 | 1,2,3,4,5,7,8,9 |
| 73 | patuletin (6-methoxyquercetin) b | C16H12O8 | 331.0464 | 331.0458 (100), 316.0024 (65.9), 287.0190 (14.1), 271.0246 (3.5), 259.0238 (3.1), 243.0285 (2.7), 181.0132 (7.1),165.9885 (19.2), 139.0023 (11.2), 109.9994 (9.6) | 7.72 | −0.161 | 1,2,7,8,9 |
| 74 | axillarin | C17H14O8 | 345.0616 | 345.0615 (99.2), 330.0381 (100), 315.0147 (48.0), 287.0196 (12.3), 243.0227 (2.6), 231.0295 (5.8), 215.0342 (4.1), 165.9897 (4.9), 149.0230 (1.2), 139.0385 (4.2), 136.9861 (1.3), 121.0280 (1.6) | 8.24 | −0.101 | 1,2,4,7,8,9 |
| 75 | Apigenin a | C15H10O5 | 269.0457 | 269.0453 (100), 225.0553 (1.6), 201.0546 (0.5), 151.0023 (5.4), 149.0239 (4.4), 117.0331 (18.4), 107.0124 (4.8) | 8.62 | 0.870 | 2,7,8,10,11 |
| 76 | Kaempferol a | C15H10O6 | 285.0405 | 285.0402 (100), 178.9938 (0.9), 151.0026 (1.0), 107.0121 (1.4) | 8.83 | −0.161 | 1,2,3,4,5,8 |
| 77 | hispidulin (scutellarein-6-methyl ether) a | C16H12O6 | 299.0563 | 299.0559 (62.4), 284.0324 (100), 255.0303 (1.3), 227.0471 (3.4), 212.0471 (3.2), 211.0389 (2.6), 164.9812 (2.0), 163.0005 (0.3), 149.9963 (1.1), 136.9865 (14.6), 117.0324 (1.5) | 8.92 | −.201 | 1,2,3,4,5,7,8,9,10,11 |
| 78 | Chrysoeriol a | C16H12O6 | 299.0562 | 299.0560 (93.1), 284.0324 (100), 256.0372 (6.4), 227.0344 (3.3), 211.0392 (1.8), 151.0024 (5.2), 133.0280 (1.6), 107.0122 (4.6) | 8.97 | −0.141 | 1,2,3,4,5,7,8,9,10,11 |
| 79 | cirsiliol | C17H14O7 | 329.0677 | 329.0667(100), 314.0432 (32.6), 299.0160 (21.2), 271.0248 (7.2), 255.0294 (1.0), 243.0294 (2.7), 230.1474 (11.8), 227.0344 (2.3), 163.0024 (2.0), 136.9874 (0.4), 135.0074 (1.4), 133.0282 (8.0) | 9.16 | 0.034 | 1,2,3,4,7,8,9,10,11 |
| 80 | quercetagetin-3,6,3’(4’)-trimethyl ether | C18H16O8 | 359.0772 | 359.0773 (100), 344.0536 (90.3), 329.0304 (49.3), 314.0068 (7.9), 301.0343 (3.5), 286.0118 (34.9), 258.0168 (10.9), 230.0214 (8.3), 202.0263 (10.1), 164.9807 (1.6), 148.0146 (1.6), 136.9854 (0.4), | 9.74 | 0.059 | 2,7,8,9,10 |
| 81 | cirsimaritin (6-hydroxyapigenin-6,7-dimethyl ether) | C17H14O6 | 313.0719 | 313.0822 (100), 298.0481 (56.4), 283.0246 (57.8), 269.0455 (2.8), 255.0299 (17.8), 227.0333 (5.8), 211.0333 (2.6), 163.0024 (19.5), 117.0326 (10.6) | 10.38 | −0.411 | 1,2,3,4,5,6,7,8,9,10,11,12 |
| 82 | santin/eupatilin | C18H16O7 | 343.0812 | 343.0822 (76.5), 328.0588 (100), 313.0355 (23.9), 298.0119 (19.4), 285.0402 (7.2), 270.0168 (24.2), 257.0085 (1.8), 254.0224 (0.7), 242.0218 (3.1), 226.0267 (1.6), 214.0266 (3.3), 198.0314 (2.3), 165.9895 (1.1), 164.9812 (0.3), 163.0020 (0.2), 136.9866 (1.9), 132.0201 (1.4) | 10.68 | −0.086 | 1,2,3,4,5,6,7,8,9,10,11 |
| 83 | acacetin | C16H12O5 | 283.0612 | 283.0610 (100), 268.0375 (72.4), 240.0425 (5.4), 239.0342 (4.8), 151.0026 (5.2), 107.0122 (3.1) | 11.44 | 1.036 | 2,7,8,9 |
| Tentatively Annotated Compound | Molecular Formula | Exact Mass [M + H]+ | Fragmentation Pattern in (+) ESI-MS/MS | tR (min) | Δ ppm | Distribution | |
| Sesquiterpene lactones and derivatives | |||||||
| 84 | tanaparthin-peroxide | C15H18O5 | 279.1226 | 279.1213 (2.82), 261.1115 (39.19), 237.1117 (100), 243.1015 (12.03), 233.1169 (38.41), 221.0806 (77.86), 215.1064 (29.10), 203.0699 (79.07), 193.0857 (48.82), 187.1112 (16.07), 175.0752 (85.47), 165.0909 (47.60), 147.0802 (45.29), 123.0441 (37.47), 105.0701 (37.62), 91.0547 (36.27), 79.0548 (21.43), 67.0550 (13.10) | 6.41 | −0.395 | 3,4,7,8,9 |
| 85 | achillicin/matricin | C17H22O5 | 307.1537 | 307.1530 (56.90), 265.1427 (14.78), 247.1324 (100), 229.1220 (34.74), 219.1376 (28.47), 201.1272 (41.37), 173.0956 (29.40), 147.0802 (52.01), 131.0852 (31.79), 105.0700 (25.38), 91.0545 (21.60), 79.0549 (16.19) | 8.04 | −0.913 | 1,2,3,4,5,6,8 |
| 86 | dehydroachillin/ dehydroleucodin | C15H16O3 | 245.1170 | 245.1166 (100), 227.1064 (6.46), 209.0956 (6.71), 199.1115 (18.76), 181.1010 (4.52), 156.0932 (3.66), 143.0852 (3.98), 123.0804 (5.57), 105.0701 (4.23), 91.0548 (3.25), 79.0548 (1.97), 69.0341 (10.05) | 9.37 | −0.860 | 1,2,3,5,7,8,11 |
| 87 | achillin/leucodin | C15H18O2 | 247.1326 | 247.1323 (100), 229.1213 (1.49), 219.1374 (4.62), 201.1272 (5.13), 191.1426 (4.81), 173.0959 (33.47), 158.0725 (5.02), 145.1009 (9.93), 135.0803 (3.08), 117.0699 (1.99), 107.0858 (3.94), 97.0651 (2.56), 79.0547 (2.01), 69.0341 (6.12), 55.0550 (0.66) | 9.55 | −1.412 | 1,2,3,4,5,6,7,8,9,10,11,12 |
| 88 | artabsin | C15H20O3 | 249.1482 | 249.1479 (73.40), 231.1375 (57.58), 221.1530 (6.61), 213.1268 (8.55), 203.1428 (100), 185.1322 (37.65), 175.1116 (75.95), 161.0958 (11.74), 157.1010 (78.89), 147.1166 (52.45), 133.1012 (32.49), 119.0857 (54.68), 10.0702 (59.52), 93.0703 (28.58), 81.0704 (10.70), 67.0550 (5.91), 55.0551 (8.37) | 11.30 | −1.409 | 1,2,3,4,5,6,7,9,10,11,12 |
| 89 | dihydrosantamarin | C15H22O3 | 251.1638 | 251.1635 (31.06), 233.1530 (42.34), 215.1428 (5.23), 205.1584 (100), 187.1478 (52.95), 177.1272 (47.19), 159.1165 (68.27), 147.1166 (20.87), 133.1010 (16.22), 119.0856 (16.77), 105.0701 (27.07), 97.0652 (17.16), 81.0704 (17.26), 67.0549 (5.80) | 12.64 | −1.597 | 1,2,3,7,8 |
| Fatty acids amides | |||||||
| 90 | tetradecenoic acid amide | C14H25NO | 224.2006 | 224.2004 (100), 196.2052 (0.09), 182.1537 (0.24), 168.1380 (6.51), 151.1115 (6.61), 123.1168 (2.56), 109.1014 (3.28), 95.0495 (6.80), 81.0340 (9.65), 69.0705 (11.84), 57.0707 (14.36) | 15.09 | −1.387 | 1,2,3,4,5,6,8,10,11,12 |
| 91 | linolenamide | C18H31NO | 278.2473 | 278.2472 (100), 261.2202 (0.89), 243.2098 (1.08), 219.1740 (0.69), 167.1302 (18.76), 152.1069 (6.86), 135.1169 (1.06), 109.1009 (4.05), 95.0859 (6.54), 81.0703 (9.21), 67.0549 (15.81) | 19.82 | −1.801 | 1,3,4,5,6,12 |
| 92 | linoleamide | C18H33NO | 280.2631 | 280.2628 (100), 263.2361 (82.19), 245.2258 (64.41), 221.2253 (3.56), 189.1632 (4.79), 179.1793 (9.84), 165.1634 (15.59), 147.1167 (8.77), 133.1011 (15.10), 123.1167 (23.89), 109.1013 (44.22), 95.0859 (71.60), 81.0704 (66.64), 69.0705 (50.93), 57.0706 (23.67) | 20.43 | −1.432 | 1,2,3,4,5,6,7,8,9,10,11,12 |
| 93 | palmitamide | C16H33NO | 256.2631 | 256.2627 (100), 214.2169 (0.26), 130.1227 (0.29), 116.1070 (1.43), 102.0916 (4.15), 88.0710 (0.33), 74.0607 (2.39) | 21.33 | −1.683 | 1,2,3,4,5,6,7,8,9,10,11,12 |
| 94 | oleamide | C18H35NO | 282.2786 | 282.2784 (100), 265.2520 (29.11), 247.2415 (25.90), 226.2156 (1.06), 212.2007 (3.29), 191.1790 (5.05), 177.1636 (4.54), 163.1478 (7.65), 149.1321 (12.53), 135.1167 (15.92), 121.1013 (14.04), 97.1015 (34.81), 83.0860 (35.84), 69.0706 (48.28) | 21.75 | −2.059 | 1,2,3,4,5,6,7,8,9,10,11,12 |
| Species | Parts | Solvents | DPPH (mg TE/g) | ABTS (mg TE/g) | CUPRAC (mg TE/g) | FRAP (mg TE/g) | MCA (mg EDTAE/g) |
|---|---|---|---|---|---|---|---|
| A. aleppica | Aerial parts | EA | 13.83 ± 0.07 e | 22.04 ± 1.31 e | 50.49 ± 3.22 e | 27.29 ± 0.21 e | 23.55 ± 1.37 ab |
| MeOH | 55.15 ± 0.05 a | 88.93 ± 0.79 b | 151.21 ± 5.64 a | 101.38 ± 1.79 a | 21.51 ± 0.09 bc | ||
| Water | 49.71 ± 1.17 b | 90.83 ± 0.12 b | 138.34 ± 1.94 b | 95.19 ± 0.62 b | 25.37 ± 0.33 a | ||
| Roots | EA | 12.44 ± 0.14 e | 30.38 ± 1.31 d | 66.55 ± 3.46 d | 34.75 ± 1.12 d | 10.28 ± 1.37 d | |
| MeOH | 35.66 ± 0.29 d | 56.23 ± 0.79 c | 88.69 ± 0.57 c | 54.62 ± 1.10 c | 12.03 ± 0.76 d | ||
| Water | 43.44 ± 0.35 c | 101.88 ± 0.98 a | 143.53 ± 0.75 ab | 93.79 ± 0.99 b | 20.25 ± 0.52 c | ||
| A. santolinoides | Aerial parts | EA | 6.57 ± 0.15 f | 15.31 ± 0.96 f | 51.59 ± 0.11 f | 25.96 ± 0.39 f | 27.37 ± 0.46 a |
| MeOH | 30.49 ± 0.30 d | 42.06 ± 0.40 e | 104.45 ± 3.32 d | 50.42 ± 1.61 d | 26.06 ± 1.20 a | ||
| Water | 51.90 ± 0.67 b | 95.34 ± 1.15 c | 164.05 ± 1.57 b | 105.24 ± 1.07 c | 21.33 ± 0.16 b | ||
| Roots | EA | 15.93 ± 0.07 e | 50.47 ± 1.33 d | 85.86 ± 2.57 e | 43.33 ± 3.63 e | 12.25 ± 1.70 d | |
| MeOH | 54.11 ± 0.03 a | 112.53 ± 0.18 a | 183.55 ± 1.68 a | 129.92 ± 3.18 a | 10.72 ± 0.42 d | ||
| Water | 47.59 ± 0.07 c | 109.04 ± 0.20 b | 151.23 ± 0.28 c | 118.50 ± 0.41 b | 17.59 ± 0.08 c |
| Species | Parts | Solvents | AChE (mg GALAE/g) | BuChE (mg GALAE/g) | Tyrosinase (mg KAE/g) | Amylase (mmol ACAE/g) | Glucosidase (mmol ACAE/g) |
|---|---|---|---|---|---|---|---|
| A. aleppica | Aerial parts | EA | 2.63±0.03 a | 6.07±0.14 a | 57.63±1.17 cd | 0.29±0.04 b | 0.64±0.01 d |
| MeOH | 2.01±0.21 bc | 2.12±0.25 c | 71.22±0.57 a | 0.22±0.01 c | 0.78±0.01 b | ||
| Water | 0.48±0.04 d | 3.83±0.01 b | 54.86±2.16 d | 0.07±0.01 d | na | ||
| Roots | EA | 2.21±0.10 b | 6.73±0.25 a | 63.26±0.93 b | 0.37±0.02 a | 0.70±0.04 c | |
| MeOH | 1.83±0.04 c | 3.92±0.54b | 70.36±0.30 a | 0.24±0.01 c | 0.85±0.01 a | ||
| Water | 0.50±0.02 d | 1.25±0.04 d | 58.83±0.74 c | 0.10±0.01 d | na | ||
| A. santolinoides | Aerial parts | EA | 2.02±0.18 c | 6.76±0.77 a | 73.00±4.87 a | 0.30±0.01 c | 0.74±0.02 a |
| MeOH | 2.32±0.23 bc | 4.74±0.41 b | 69.02±0.86 a | 0.35±0.01 b | 0.66±0.08 ab | ||
| Water | 0.55±0.04 d | na | 40.32±1.40 b | 0.04±0.01 f | na | ||
| Roots | EA | 2.61±0.04 ab | 6.70±0.72 a | 66.99±1.98 a | 0.40±0.01 a | 0.60±0.01 b | |
| MeOH | 2.83±0.32 a | 3.28±0.17 c | 72.60±0.34 a | 0.19±0.01 d | 0.38±0.07 c | ||
| Water | 0.70±0.07 d | 0.78±0.02 d | 39.23±0.78 b | 0.10±0.01 e | na |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gevrenova, R.; Zengin, G.; Sinan, K.I.; Yıldıztugay, E.; Zheleva-Dimitrova, D.; Picot-Allain, C.; Mahomoodally, M.F.; Imran, M.; Dall’Acqua, S. UHPLC-MS Characterization and Biological Insights of Different Solvent Extracts of Two Achillea Species (A. aleppica and A. santolinoides) from Turkey. Antioxidants 2021, 10, 1180. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10081180
Gevrenova R, Zengin G, Sinan KI, Yıldıztugay E, Zheleva-Dimitrova D, Picot-Allain C, Mahomoodally MF, Imran M, Dall’Acqua S. UHPLC-MS Characterization and Biological Insights of Different Solvent Extracts of Two Achillea Species (A. aleppica and A. santolinoides) from Turkey. Antioxidants. 2021; 10(8):1180. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10081180
Chicago/Turabian StyleGevrenova, Reneta, Gokhan Zengin, Kouadio Ibrahime Sinan, Evren Yıldıztugay, Dimitrina Zheleva-Dimitrova, Carene Picot-Allain, Mohamad Fawzi Mahomoodally, Muhammad Imran, and Stefano Dall’Acqua. 2021. "UHPLC-MS Characterization and Biological Insights of Different Solvent Extracts of Two Achillea Species (A. aleppica and A. santolinoides) from Turkey" Antioxidants 10, no. 8: 1180. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10081180