
Microbial Biosurfactant: A New Frontier for Sustainable Agriculture and Pharmaceutical Industries

 ,
,  , , , ,
, , , ,  ,
, 
Abstract
:1. Introduction
2. Microorganisms and Biosurfactants
3. Factors Affecting Biosurfactant Production
4. Biosurfactant Applications in Improving the Soil Quality
5. Biosurfactant Application in Plant Disease Management
6. Biosurfactant Application in Pharmaceutical Industries
6.1. Antioxidant Properties of Biosurfactants
6.2. Antimicrobial Properties of Biosurfactants
7. Future Directions and Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Popp, J.; Pető, K.; Nagy, J. Pesticide productivity and food security: A review. Agron. Sustain. Dev. 2013, 33, 243–255. [Google Scholar] [CrossRef]
- Muthusamy, K.; Gopalakrishnan, S.; Ravi, T.K.; Sivachidambaram, P. Biosurfactants: Properties, commercial production and application. Curr. Sci. 2008, 94, 736–747. [Google Scholar]
- Singh, S.K.; Singh, M.K.; Verma, H.; Singh, P.P.; Singh, A.V.; Rashmi, K.; Kumar, A. Biosurfactant producing microbes for clean-up of soil contaminants. In Microbe Mediated Remediation of Environmental Contaminants; Woodhead Publishing: Sawston, UK, 2021; pp. 89–93. [Google Scholar]
- Rahman, P.K.; Gakpe, E. Production, characterisation and applications of biosurfactants-Review. Biotechnology 2008, 7, 360–370. [Google Scholar] [CrossRef] [Green Version]
- Varjani, S.J.; Upasani, V.N. Critical review on biosurfactant analysis, purification and characterization using rhamnolipid as a model biosurfactant. Bioresour. Technol. 2017, 232, 389–397. [Google Scholar] [CrossRef]
- Vaidya, S.; Ganguli, A.K. Microemulsion Methods for Synthesis of Nanostructured Materials; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Sáenz-Marta, C.I.; de Lourdes Ballinas-Casarrubias, M.; Rivera-Chavira, B.E.; Nevárez-Moorillón, G.V. Biosurfactants as useful tools in bioremediation. In Advances in Bioremediation of Wastewater and Polluted Soil; InTechopen: London, UK, 2015; Volume 5. [Google Scholar]
- Mohanty, S.S.; Koul, Y.; Varjani, S.; Pandey, A.; Ngo, H.H.; Chang, J.S.; Wong, J.W.; Bui, X.T. A critical review on various feedstocks as sustainable substrates for biosurfactants production: A way towards cleaner production. Microb. Cell Fact. 2021, 20, 1–13. [Google Scholar] [CrossRef]
- Kuyukina, M.S.; Ivshina, I.B.; Makarov, S.O.; Litvinenko, L.V.; Cunningham, C.J.; Philp, J.C. Effect of biosurfactants on crude oil desorption and mobilization in a soil system. Environ. Int. 2005, 31, 155–161. [Google Scholar] [CrossRef]
- Shekhar, S.; Sundaramanickam, A.; Balasubramanian, T. Biosurfactant producing microbes and their potential applications: A review. Crit. Rev. Environ. Sci. Technol. 2015, 45, 1522–1554. [Google Scholar] [CrossRef]
- Bhatt, P.; Verma, A.; Gangola, S.; Bhandari, G.; Chen, S. Microbial glycoconjugates in organic pollutant bioremediation: Recent advances and applications. Microb. Cell Fact. 2021, 20, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.K.; Das, K. Microbial Surfactants and Their Potential Applications: An Overview. In Biosurfactants; Sen, R., Ed.; Springer: New York, NY, USA, 2010; Volume 672. [Google Scholar]
- Mulligan, C.N. Environmental applications for biosurfactants. Environ. Pollut. 2005, 133, 183–198. [Google Scholar] [CrossRef]
- Vatsa, P.; Sanchez, L.; Clement, C.; Baillieul, F.; Dorey, S. Rhamnolipid biosurfactants as new players in animal and plant defense against microbes. Int. J. Mol. Sci. 2010, 11, 5095–5108. [Google Scholar] [CrossRef] [Green Version]
- Tahir, Z.; Nazir, M.S.; Aslam, A.A.; Bano, S.; Ali, Z.; Akhtar, M.N.; Azam, K.; Abdullah, M.A. Active metabolites and biosurfactants for utilization in environmental remediation and eco-restoration of polluted soils. In Green Sustainable Process for Chemical and Environmental Engineering and Science; Elsevier: Amsterdam, The Netherlands, 2021; pp. 31–51. [Google Scholar]
- Prasad, N.K.; Jayakumar, R.; Wagdevi, P.; Ramesh, S. Application of biosurfactants as bioabsorption agents of heavily contaminated soil and water. In Green Sustainable Process for Chemical and Environmental Engineering and Science; Elsevier: Alpharetta, GA, USA, 2021; pp. 21–30. [Google Scholar]
- Santos, D.K.F.; Rufino, R.D.; Luna, J.M.; Santos, V.A.; Sarubbo, L.A. Biosurfactants: Multifunctional biomolecules of the 21st century. Int. J. Mol. Sci. 2016, 17, 401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patowary, K.; Patowary, R.; Kalita, M.C.; Deka, S. Characterization of biosurfactant produced during degradation of hydrocarbons using crude oil as sole source of carbon. Front. Microbiol. 2017, 8, 279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saharan, B.S.; Sahu, R.K.; Sharma, D. A review on biosurfactants: Fermentation, current developments and perspectives. Genet. Eng. Biotechnol. J. 2011, 1, 1–14. [Google Scholar]
- Chen, C.Y.; Baker, S.C.; Darton, R.C. The application of a high throughput analysis method for the screening of potential biosurfactants from natural sources. J. Microbiol. Methods 2007, 70, 503–510. [Google Scholar] [CrossRef]
- Bushnell, L.D.; Haas, H.F. The utilization of certain hydrocarbons by microorganisms. J. Bacteriol. 1941, 41, 653–673. [Google Scholar] [CrossRef] [Green Version]
- Ilori, M.O.; Adebusoye, S.A.; Ojo, A.C. Isolation and characterization of hydrocarbon-degrading and biosurfactant-producing yeast strains obtained from a polluted lagoon water. World J. Microbiol. Biotechnol. 2008, 24, 2539–2545. [Google Scholar] [CrossRef]
- Balan, S.S.; Kumar, C.G.; Jayalakshmi, S. Pontifactin, a new lipopeptide biosurfactant produced by a marine Pontibacter korlensis strain SBK-47: Purification, characterization and its biological evaluation. Process Biochem. 2016, 51, 2198–2207. [Google Scholar] [CrossRef]
- Coronel-León, J.; de Grau, G.; Grau-Campistany, A.; Farfan, M.; Rabanal, F.; Manresa, A.; Marqués, A.M. Biosurfactant production by AL 1.1, a Bacillus licheniformis strain isolated from Antarctica: Production, chemical characterization and properties. Ann. Microbiol. 2015, 65, 2065–2078. [Google Scholar] [CrossRef]
- Xu, M.; Fu, X.; Gao, Y.; Duan, L.; Xu, C.; Sun, W.; Li, Y.; Meng, X.; Xiao, X. Characterization of a biosurfactant-producing bacteria isolated from Marine environment: Surface activity, chemical characterization and biodegradation. J. Environ. Chem. Eng. 2020, 8, 104277. [Google Scholar] [CrossRef]
- Silva, S.N.; Farias, C.B.; Rufino, R.D.; Luna, J.M.; Sarubbo, L.A. Glycerol as substrate for the production of biosurfactant by Pseudomonas aeruginosa UCP0992. Colloids Surf. B Biointerfaces 2010, 1, 174–183. [Google Scholar] [CrossRef]
- Santa Anna, L.M.; Sebastian, G.V.; Menezes, E.P.; Alves, T.L.M.; Santos, A.S.; Pereira, N., Jr.; Freire, D.M.G. Production of biosurfactants from Pseudomonas aeruginosa PA 1 isolated in oil environments. Braz. J. Chem. Eng. 2002, 19, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Jadhav, M.; Kalme, S.; Tamboli, D.; Govindwar, S. Rhamnolipid from Pseudomonas desmolyticum NCIM-2112 and its role in the degradation of Brown 3REL. J. Basic Microbiol. 2011, 51, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.H.; Lai, H.C.; Chen, S.Y.; Yeh, M.S.; Chang, J.S. Biosurfactant production by Serratia marcescens SS-1 and its isogenic strain SMΔR defective in SpnR, a quorum-sensing LuxR family protein. Biotechnol. Lett. 2004, 26, 799–802. [Google Scholar] [CrossRef] [PubMed]
- Revathi, K.; Chandrasekaran, R.; Thanigaivel, A.; Kirubakaran, S.A.; Sathish-Narayanan, S.; Senthil-Nathan, S. Effects of Bacillus subtilis metabolites on larval Aedes aegypti L. Pestic. Biochem. Physiol. 2013, 107, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Naruse, N.; Tenmyo, O.; Kobaru, S.; Kamei, H.; Miyaki, T.; Konishi, M.; Oki, T. Pumilacidin, a complex of new antiviral antibiotics: Production, isolation, chemical properties, structure and biological activity. J. Antibiot. 1990, 43, 267–280. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Wang, Y.; Zang, T.; Wei, J.; Wu, H.; Wei, C.; Qiu, G.; Li, F. A biosurfactant-producing Pseudomonas aeruginosa S5 isolated from coking wastewater and its application for bioremediation of polycyclic aromatic hydrocarbons. Bioresour. Technol. 2019, 281, 421–428. [Google Scholar] [CrossRef]
- Jang, J.Y.; Yang, S.Y.; Kim, Y.C.; Lee, C.W.; Park, M.S.; Kim, J.C.; Kim, I.S. Identification of orfamide A as an insecticidal metabolite produced by Pseudomonas protegens F6. J. Agric. Food Chem. 2013, 61, 6786–6791. [Google Scholar] [CrossRef]
- Goswami, D.; Handique, P.J.; Deka, S. Rhamnolipid biosurfactant against Fusarium sacchari the causal organism of pokkah boeng disease of sugarcane. J. Basic Microbiol. 2014, 54, 548–555. [Google Scholar] [CrossRef]
- Janek, T.; Lukaszewicz, M.; Krasowska, A. Antiadhesive activity of the biosurfactant pseudofactin II secreted by the Arctic bacterium Pseudomonas fluorescens BD5. BMC Microbiol. 2012, 12, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Donio, M.B.; Ronica, S.F.; Viji, V.T.; Velmurugan, S.; Jenifer, J.A.; Michaelbabu, M.; Citarasu, T. Isolation and characterization of halophilic Bacillus sp. BS3 able to produce pharmacologically important biosurfactants. Asian Pac. J. Trop. Med. 2013, 6, 876–883. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Shi, R.; Zhao, J.; Li, G.; Bai, X.; Han, S.; Zhang, Y. Heterologous production of Pseudomonas aeruginosa rhamnolipid under anaerobic conditions for microbial enhanced oil recovery. J. Appl. Microbiol. 2015, 118, 379–389. [Google Scholar] [CrossRef]
- Singh, A.K.; Cameotra, S.S. Efficiency of lipopeptide biosurfactants in removal of petroleum hydrocarbons and heavy metals from contaminated soil. Environ. Sci. Pollut. Res. 2013, 20, 7367–7376. [Google Scholar] [CrossRef]
- Solaiman, D.K.; Ashby, R.D.; Crocker, N.V. High-titer production and strong antimicrobial activity of sophorolipids from Rhodotorula bogoriensis. Biotechnol. Prog. 2015, 31, 867–874. [Google Scholar] [CrossRef]
- Campos, J.M.; Stamford, T.L.; Sarubbo, L.A. Production of a bioemulsifier with potential application in the food industry. Appl. Biochem. Biotechnol. 2014, 172, 3234–3252. [Google Scholar] [CrossRef]
- Rufino, R.D.; De Luna, J.M.; De Campos Takaki, G.M.; Sarubbo, L.S. Characterization and properties of the biosurfactant produced by Candida lipolytica UCP 0988. Electron. J. Biotechnol. 2014, 17, 6. [Google Scholar] [CrossRef] [Green Version]
- Gautam, G.; Mishra, V.; Verma, P.; Pandey, A.K.; Negi, S.A. Cost Effective Strategy for Production of Bio-surfactant from Locally Isolated Penicillium chrysogenum SNP5 and its Applications. J. Bioprocess. Biotechnol. 2014, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Andrade, S.; Luna, M.A.; Santiago, A.L.; Franco, L.O.; Silva, G.K.; de Souza, P.M.; Okada, K.; Albuquerque, C.D.; da Silva, C.A.; Campos-Takaki, G.M. Biosurfactant-and-bioemulsifier produced by a promising Cunninghamella echinulata isolated from Caatinga soil in the northeast of Brazil. Int. J. Mol. Sci. 2014, 15, 15377–15395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitamoto, D.; Haneishi, K.; Nakahara, T.; Tabuchi, T. Production of mannosylerythritol lipids by Candida antarctica from vegetable oils. Agric. Biol. Chem. 1990, 54, 37–40. [Google Scholar] [CrossRef] [Green Version]
- Zinjarde, S.; Pant, A. Emulsifier from tropical marine yeast, Yarrowia lipolytica NCIM 3589. J. Basic Microbiol. 2002, 42, 67–73. [Google Scholar] [CrossRef]
- Holler, U.; Konig, G.M.; Wright, A. Three new metabolites from marine derived fungi of the genera Coniothyrium and Microsphaeropsis. J. Nat. Prod. 1999, 62, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Amaral, P.F.F.; Da Silva, J.M.; Lehocky, B.M.; Barros-Timmons, A.M.V.; Coelho, M.A.Z.; Marrucho, I.M.; Coutinho, J.A.P. Production and characterization of a bioemulsifier from Yarrowia lipolytica. Process Biochem. 2006, 41, 1894–1898. [Google Scholar] [CrossRef]
- Teichmann, B.; Linne, U.; Hewald, S.; Marahiel, M.A.; Bölker, M. A biosynthetic gene cluster for a secreted cellobiose lipid with antifungal activity from Ustilago maydis. Mol. Microbiol. 2007, 66, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Oh, D.K.; Lee, K.H.; Kim, J.H. Effect of soybean oil and glucose on sophorose lipid fermentation by Torulopsis bombicola in continuous culture. Appl. Microbiol. Biotechnol. 1997, 48, 23–26. [Google Scholar] [CrossRef] [PubMed]
- Kiran, G.S.; Hema, T.A.; Gandhimathi, R.; Selvin, J.; Thomas, T.A.; Ravji, T.R.; Natarajaseenivasan, K. Optimization and production of a biosurfactant from the sponge-associated marine fungus Aspergillus ustus MSF3. Colloids Surf. B Biointerfaces 2009, 73, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Felse, P.A.; Shah, V.; Chan, J.; Rao, K.J.; Gross, R.A. Sophorolipid biosynthesis by Candida bombicola from industrial fatty acid residues. Enzyme Microb. Technol. 2007, 40, 316–323. [Google Scholar] [CrossRef]
- Alej, C.S.; Humberto, H.S.; Maria, J.F. Production of glycolipids with antimicrobial activity by Ustilago maydis FBD12 in submerged culture. Afr. J. Microbiol. Res. 2011, 5, 2512–2523. [Google Scholar]
- Ribeiro, I.A.; Faustino, C.M.; Guerreiro, P.S.; Frade, R.F.; Bronze, M.R.; Castro, M.F.; Ribeiro, M.H. Development of novel sophorolipids with improved cytotoxic activity toward MDA-MB-231 breast cancer cells. J. Mol. Recognit. 2015, 28, 155–165. [Google Scholar] [CrossRef]
- Cooper, D.G.; Paddock, D.A. Torulopsis petrophilum and surface activity. Appl. Environ. Microbiol. 1983, 46, 1426–1429. [Google Scholar] [CrossRef] [Green Version]
- Lukondeh, T.; Ashbolt, N.J.; Rogers, P.L. Evaluation of Kluyveromyces marxianus FII 510700 grown on a lactose-based medium as a source of a natural bioemulsifier. J. Ind. Microbiol. Biotechnol. 2003, 30, 715–720. [Google Scholar] [CrossRef]
- Rau, U.; Nguyen, L.A.; Roeper, H.; Koch, H.; Lang, S. Fed-batch bioreactor production of mannosylerythritol lipids secreted by Pseudozyma aphidis. Appl. Microbiol. Biotechnol. 2005, 68, 607–613. [Google Scholar] [CrossRef]
- Morita, T.; Takashima, M.; Fukuoka, T.; Konishi, M.; Imura, T.; Kitamoto, D. Isolation of basidiomycetous yeast Pseudozyma tsukubaensis and production of glycolipid biosurfactant, a diastereomer type of mannosylerythritol lipid-B. Appl. Microbiol. Biotechnol. 2010, 88, 679–688. [Google Scholar] [CrossRef]
- Ribeiro, B.G.; Guerra, J.M.C.; Sarubbo, L.A. Potential food application of a biosurfactant produced by Saccharomyces cerevisiae URM 6670. Front. Bioeng. Biotechnol. 2020, 8, 434. [Google Scholar] [CrossRef] [PubMed]
- Chandran, P.R.; Das, N.I. Biosurfactant production and diesel oil degradation by yeast species Trichosporon asahii isolated from petroleum hydrocarbon contaminated soil. Int. J. Eng. Sci. Technol. 2010, 2, 6942–6953. [Google Scholar]
- Sharma, P.; Sangwan, S.; Kaur, H. Process parameters for biosurfactant production using yeast Meyerozyma guilliermondii YK32. Environ. Monit. Assess. 2019, 191, 531. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.; Borah, S.N.; Bora, A.; Deka, S. Production, characterization, and antifungal activity of a biosurfactant produced by Rhodotorula babjevae YS3. Microb. Cell Fact. 2017, 16, 95. [Google Scholar] [CrossRef] [Green Version]
- Joshi-Navare, K.; Singh, P.K.; Prabhune, A.A. New yeast isolate Pichia caribbica synthesizes xylolipid biosurfactant with enhanced functionality. Eur. J. Lipid Sci. Technol. 2014, 116, 1070–1079. [Google Scholar] [CrossRef]
- Thanomsub, B.; Watcharachaipong, T.; Chotelersak, K.; Arunrattiyakorn, P.; Nitoda, T.; Kanzaki, H. Monoacylglycerols: Glycolipid biosurfactants produced by a thermotolerant yeast, Candida ishiwadae. J. Appl. Microbiol. 2004, 96, 588–592. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Das, P.; Sen, R. Towards commercial production of microbial surfactants. Trends Biotechnol. 2006, 24, 509–515. [Google Scholar] [CrossRef]
- McInerney, M.J.; Duncan, K.E.; Youssef, N.; Fincher, T.; Maudgalya, S.K.; Folmsbee, M.J.; Knapp, R.; Simpson, R.R.; Ravi, N.; Nagle, D. Development of Microorganisms with Improved Transport and Biosurfactant Activity for Enhanced oil Recovery; University of Oklahoma: Norman, OK, USA, 2005. [Google Scholar]
- Sheppard, J.D.; Mulligan, C.N. The production of surfactin by Bacillus subtilis grown on peat hydrolysate. Appl. Microbiol. Biotechnol. 1987, 27, 110–116. [Google Scholar] [CrossRef]
- Singh, P.; Patil, Y.; Rale, V. Biosurfactant production: Emerging trends and promising strategies. J. Appl. Microbiol. 2019, 126, 2–13. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Xu, J.; Xie, W.; Yao, Z.; Yang, H.; Sun, C.; Li, X. Pseudomonas aeruginosa L10: A hydrocarbon-degrading, biosurfactant-producing, and plant-growth-promoting endophytic bacterium isolated from a reed (Phragmites australis). Front. Microbiol. 2018, 9, 1087. [Google Scholar] [CrossRef]
- Pansiripat, S.; Pornsunthorntawee, O.; Rujiravanit, R.; Kitiyanan, B.; Somboonthanate, P.; Chavadej, S. Biosurfactant production by Pseudomonas aeruginosa SP4 using sequencing batch reactors: Effect of oil-to-glucose ratio. Biochem. Eng. J. 2010, 49, 185–191. [Google Scholar] [CrossRef]
- Hassan, M.; Essam, T.; Yassin, A.S.; Salama, A. Optimization of rhamnolipid production by biodegrading bacterial isolates using Plackett-Burman design. Int. J. Biol. Macromol. 2016, 8, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Onwosi, C.O.; Odibo, F.J.C. Effects of carbon and nitrogen sources on rhamnolipid biosurfactant production by Pseudomonas nitroreducens isolated from soil. World J. Microbiol. Biotechnol. 2012, 28, 937–942. [Google Scholar] [CrossRef]
- Moussa, T.A.A.; Mohamed, M.S.; Samak, N. Production and characterization of di-rhamnolipid produced by Pseudomonas aeruginosa TMN. Braz. J. Chem. Eng. 2014, 31, 867–880. [Google Scholar] [CrossRef] [Green Version]
- Yeh, M.; Wei, Y.; Chang, J. Enhanced Production of Surfactin from Bacillus subtilis by addition of solid carriers. Biotechnol. Prog. 2005, 21, 1329–1334. [Google Scholar] [CrossRef]
- Ghribi, D.; Ellouze-Chaabouni, S. Enhancement of Bacillus subtilis lipopeptide biosurfactants production through optimization of medium composition and adequate control of aeration. Biotechnol. Res. Int. 2011, 2011, 653654. [Google Scholar] [CrossRef] [Green Version]
- Fooladi, T.; Bin, A.; Hamid, A.; Mohtar, W.; Yusoff, W. Production of Biosurfactant by Indigenous Isolated Bacteria in Fermentation System. AIP Conf. Proc. 2013, 197, 197–201. [Google Scholar]
- Tomar, G.S.; Srinikethan, G. Studies on production of biosurfactant from Pseudomonas aeruginosa (MTCC7815) & its application in microbial enhanced oil recovery. Res. J. Chem. Environ. Sci. 2016, 4, 84–91. [Google Scholar]
- Willenbacher, J.; Rau, J.T.; Rogalla, J.; Syldatk, C.; Hausmann, R. Foam-free production of Surfactin via anaerobic fermentation of Bacillus subtilis DSM 10T. AMB Express 2015, 5, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ejike Ogbonna, K.; Victor Agu, C.; Okonkwo, C.C.; Tochukwu Ughamba, K.; Akor, J.; Njoku, O.U. Use of Spondias Mombin fruit pulp as a substrate for biosurfactant production. Bioengineered 2021, 12, 1–12. [Google Scholar] [CrossRef]
- Kanna, R.; Gummadi, S.N.; Kumar, G.S. Production and characterization of biosurfactant by Pseudomonas putida MTCC 2467. J. Biol. Sci. 2014, 14, 436–445. [Google Scholar] [CrossRef] [Green Version]
- de França, Í.W.L.; de Oliveira, D.W.F.; Giro, M.E.A.; Melo, V.M.M.; Gonçalves, L.R.B. Production of surfactin by Bacillus subtilis LAMI005 and evaluation of its potential as tensoactive and emulsifier. Can. J. Chem. Eng. 2021. [Google Scholar] [CrossRef]
- Agarry, S.E.; Salam, K.K.; Arinkoola, A.; Aremu, M.O. Biosurfactant production by indigeneous Pseudomonas and Bacillus species isolated from auto-mechanic soil environment toward microbial enhanced oil recovery. Eur. J. Eng. Technol. 2015, 3, 27–39. [Google Scholar]
- Cherif, N.; Tifrit, A.; Larbi Daouadji, K.; Mezouari, S.; Chama, Z.; Abbouni, B. Effect of carbon and nitrogen source on the microbial production of biosurfactants by Pseudomonas aeruginosa. Pharm. Lett. 2015, 7, 42–48. [Google Scholar]
- Abouseoud, M.; Maachi, R.; Amrane, A.; Boudergua, S.; Nabi, A. Evaluation of different carbon and nitrogen sources in production of biosurfactant by Pseudomonas fluorescens. Desalination 2008, 223, 143–151. [Google Scholar] [CrossRef]
- Abdel-Mawgoud, A.M.; Aboulwafa, M.M.; Hassouna, N.A.H. Optimization of surfactin production by Bacillus subtilis isolate BS5. Appl. Biochem. Biotechnol. 2008, 150, 305–325. [Google Scholar] [CrossRef]
- Liu, C.; You, Y.; Zhao, R.; Sun, D.; Zhang, P.; Jiang, J.; Zhu, A.; Liu, W. Biosurfactant production from Pseudomonas taiwanensis L1011 and its application in accelerating the chemical and biological decolorization of azo dyes. Ecotoxicol. Environ. Saf. 2017, 145, 8–15. [Google Scholar] [CrossRef]
- Baskaran, S.M.; Zakaria, M.R.; Sabri, A.S.M.A.; Mohamed, M.S.; Wasoh, H.; Toshinari, M.; Hassan, M.A.; Banat, I.M. Valorization of biodiesel side stream waste glycerol for rhamnolipids production by Pseudomonas aeruginosa RS6. Environ. Pollut. 2021, 276, 116742. [Google Scholar] [CrossRef]
- Almansoory, A.F.; Hasan, H.A.; Idris, M.; Abdullah, S.R.S.; Anuar, N. Biosurfactant production by the hydrocarbon-degrading bacteria (HDB) Serratia marcescens: Optimization using central composite design (CCD). J. Ind. Eng. Chem. 2017, 47, 272–280. [Google Scholar] [CrossRef]
- Patel, P.; Modi, A.; Minipara, D.; Kumar, A. Microbial biosurfactants in management of organic waste. In Sustainable Environmental Clean-Up; Elsevier: Amsterdam, The Nederland, 2021; pp. 211–230. [Google Scholar]
- Almeida, P.F.; Moreira, R.S.; Almeida, R.C.C.; Guimardes, A.K.; Carvalho, A.S.; Quintella, C. Selection and application of microorganisms to improve oil recovery. Eng. Life Sci. 2004, 4, 319–324. [Google Scholar] [CrossRef]
- De Lima, C.J.B.; Ribeiro, E.J.; Servulo, E.F.C.; Resende, M.M.; Cardoso, V.L. Biosurfactant production by Pseudomonas aeruginosa grown in residual soybean oil. Appl. Biochem. Biotechnol. 2009, 152, 156–168. [Google Scholar] [CrossRef]
- Sivapathasekaran, C.; Sen, R. Origin, properties, production and purification of microbial surfactants as molecules with immense commercial potential. Tenside Surfactants Deterg. 2017, 54, 92–107. [Google Scholar]
- Agarwal, P.; Sharma, D.K. Studies on the production of biosurfactant for the microbial enhanced oil recovery by using bacteria isolated from oil contaminated wet soil. Pet. Sci. Technol. 2009, 27, 1880–1893. [Google Scholar] [CrossRef]
- Al-Bahry, S.N.; Al-Wahaibi, Y.M.; Elshafie, A.E.; Al-Bemani, A.S.; Joshi, S.J.; Al-Makhmari, H.S.; Al-Sulaimani, H.S. Biosurfactant production by Bacillus subtilis B20 using date molasses and its possible application in enhanced oil recovery. Int. Biodeterior. Biodegrad. 2013, 81, 141–146. [Google Scholar] [CrossRef]
- Hentati, D.; Chebbi, A.; Hadrich, F.; Frikha, I.; Rabanal, F.; Sayadi, S.; Manresa, A.; Chamkha, M. Production, characterization and biotechnological potential of lipopeptide biosurfactants from a novel marine Bacillus stratosphericus strain FLU5. Ecotoxicol. Environ. Saf. 2019, 167, 441–449. [Google Scholar] [CrossRef]
- Souza, K.S.T.; Gudiña, E.J.; Schwan, R.F.; Rodrigues, L.R.; Dias, D.R.; Teixeira, J.A. Improvement of biosurfactant production by Wickerhamomyces anomalus CCMA 0358 and its potential application in bioremediation. J. Hazard. Mater. 2018, 346, 152–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazareth, T.C.; Zanutto, C.P.; Maass, D.; de Souza, A.A.U.; Ulson, S.M.D.A.G. A low-cost brewery waste as a carbon source in bio-surfactant production. Bioprocess Biosyst. Eng. 2021, 1–8. [Google Scholar] [CrossRef]
- De Lima, A.M.; De Souzaa, R.R. Use of sugar cane vinasse as substrate for biosurfactant production using Bacillus subtilis PC. Chem. Eng. 2014, 37, 673–678. [Google Scholar]
- Gudiña, E.J.; Fernandes, E.C.; Rodrigues, A.I.; Teixeira, J.A.; Rodrigues, L.R. Biosurfactant production by Bacillus subtilis using corn steep liquor as culture medium. Front. Microbiol. 2015, 6, 59. [Google Scholar] [PubMed] [Green Version]
- Gurjar, J.; Sengupta, B. Production of surfactin from rice mill polishing residue by submerged fermentation using Bacillus subtilis MTCC 2423. Bioresour. Technol. 2015, 189, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Coronel-León, J.; Marqués, A.M.; Bastida, J.; Manresa, A. Optimizing the production of the biosurfactant lichenysin and its application in biofilm control. J. Appl. Microbiol. 2016, 120, 99–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Deng, M.; Wang, Y.; Chen, W. Production and characteristics of biosurfactant produced by Bacillus pseudomycoides BS6 utilizing soybean oil waste. Int. Biodeterior. Biodegrad. 2016, 112, 72–79. [Google Scholar] [CrossRef]
- Maass, D.; Moya Ramirez, I.; Garcia Roman, M.; Jurado Alameda, E.; Ulson de Souza, A.A.; Borges Valle, J.A.; Altmajer Vaz, D. Two-phase olive mill waste (alpeorujo) as carbon source for biosurfactant production. J. Chem. Technol. Biotechnol. 2016, 91, 1990–1997. [Google Scholar] [CrossRef]
- Jajor, P.; Piłakowska-Pietras, D.; Krasowska, A.; Łukaszewicz, M. Surfactin analogues produced by Bacillus subtilis strains grown on rapeseed cake. J. Mol. Struct. 2016, 1126, 141–146. [Google Scholar] [CrossRef]
- Zhi, Y.; Wu, Q.; Xu, Y. Production of surfactin from waste distillers’ grains by co-culture fermentation of two Bacillus amyloliquefaciens strains. Bioresour. Technol. 2017, 235, 96–103. [Google Scholar] [CrossRef]
- Phulpoto, I.A.; Yu, Z.; Hu, B.; Wang, Y.; Ndayisenga, F.; Li, J.; Liang, H.; Qazi, M.A. Production and characterization of surfactin-like biosurfactant produced by novel strain Bacillus nealsonii S2MT and it’s potential for oil contaminated soil remediation. Microb. Cell Fact. 2020, 19, 1–12. [Google Scholar] [CrossRef]
- Hentati, D.; Cheffi, M.; Hadrich, F.; Makhloufi, N.; Rabanal, F.; Manresa, A.; Sayadi, S.; Chamkha, M. Investigation of halotolerant marine Staphylococcus sp. CO100, as a promising hydrocarbon-degrading and biosurfactant-producing bacterium, under saline conditions. J. Environ. Manag. 2021, 277, 111480. [Google Scholar] [CrossRef]
- Cheffi, M.; Maalej, A.; Mahmoudi, A.; Hentati, D.; Marques, A.M.; Sayadi, S.; Chamkha, M. Lipopeptides production by a newly Halomonas venusta strain: Characterization and biotechnological properties. Bioorg. Chem. 2021, 109, 104724. [Google Scholar] [CrossRef]
- Derguine-Mecheri, L.; Kebbouche-Gana, S.; Djenane, D. Biosurfactant production from newly isolated Rhodotorula sp. YBR and its great potential in enhanced removal of hydrocarbons from contaminated soils. World J. Microbiol. Biotechnol. 2021, 37, 1–8. [Google Scholar] [CrossRef]
- Ahmadi, N.; Shafaati, M.; Mirgeloybayat, M.; Lavasani, P.S.; Khaledi, M.; Afkhami, H. Molecular identification and primary evaluation of bio-surfactant production in edible paddy straw mushroom. Gene Rep. 2021, 24, 101258. [Google Scholar] [CrossRef]
- Auhim, H.S.; Mohamed, A.I. Effect of different environmental and nutritional factors on biosurfactant production from Azotobacter chroococcum. Int. J. Adv. Pharm. Biol. Chem. 2013, 2, 477–481. [Google Scholar]
- Joaad, A.I.; Hassan, S.S. Effect of Different Environmental and Nutritional Factors on Biosurfactant Production from Candida guilliermondii. Iraqi J. Sci. 2015, 56, 329–336. [Google Scholar]
- Robinson, B.H.; Bañuelos, G.; Conesa, H.M.; Evangelou, M.W.; Schulin, R. The phytomanagement of trace elements in soil. Crit. Rev. Plant Sci. 2009, 28, 240–266. [Google Scholar] [CrossRef]
- Roy, A. Review on the biosurfactants: Properties, types and its applications. J. Fundam. Renew. Energy Appl. 2017, 8, 1–14. [Google Scholar] [CrossRef]
- Singh, R.; Glick, B.R.; Rathore, D. Biosurfactants as a biological tool to increase micronutrient availability in soil: A review. Pedosphere 2018, 28, 170–189. [Google Scholar] [CrossRef]
- Pacwa-Płociniczak, M.; Płaza, G.A.; Piotrowska-Seget, Z.; Cameotra, S.S. Environmental applications of biosurfactants: Recent advances. Int. J. Mol. Sci. 2011, 12, 633–654. [Google Scholar] [CrossRef]
- Mnif, I.; Ghribi, D. Review lipopeptides biosurfactants: Mean classes and new insights for industrial, biomedical, and environmental applications. Pept. Sci. 2015, 104, 129–147. [Google Scholar] [CrossRef]
- Rufino, R.D.; Luna, J.M.; Sarubbo, L.A.; Rodrigues, L.R.M.; Teixeira, J.A.C.; Campos-Takaki, G.M. Antimicrobial and anti-adhesive potential of a biosurfactant Rufisan produced by Candida lipolytica UCP 0988. Colloids Surf. B Biointerfaces 2011, 84, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Cameotra, S.S. Potential applications of microbial surfactants in biomedical sciences. Trends Biotechnol. 2004, 22, 142–146. [Google Scholar] [CrossRef]
- Olaniran, A.O.; Balgobind, A.; Pillay, B. Bioavailability of heavy metals in soil: Impact on microbial biodegradation of organic compounds and possible improvement strategies. Int. J. Mol. Sci. 2013, 14, 10197–10228. [Google Scholar] [CrossRef] [Green Version]
- Bustamante, M.; Duran, N.; Diez, M.C. Biosurfactants are useful tools for the bioremediation of contaminated soil: A review. J. Soil Sci. Plant Nutr. 2012, 12, 667–687. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Mulligan, C.N. An evaluation of surfactant foam technology in remediation of contaminated soil. Chemosphere 2004, 57, 1079–1089. [Google Scholar] [CrossRef]
- Sheng, X.; He, L.; Wang, Q.; Ye, H.; Jiang, C. Effects of inoculation of biosurfactant-producing Bacillus sp. J119 on plant growth and cadmium uptake in a cadmium-amended soil. J. Hazard. Mater. 2008, 155, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Stacey, S.P.; McLaughlin, M.J.; Çakmak, I.; Hettiarachchi, G.M.; Scheckel, K.G.; Karkkainen, M. Root uptake of lipophilic zinc- rhamnolipid complexes. J. Agric. Food Chem. 2008, 56, 2112–2117. [Google Scholar] [CrossRef]
- Singha, L.P.; Pandey, P. Rhizosphere assisted bioengineering approaches for the mitigation of petroleum hydrocarbons contamination in soil. Crit. Rev. Biotechnol. 2021, 41, 1–26. [Google Scholar] [CrossRef]
- Liao, C.; Xu, W.; Lu, G.; Deng, F.; Liang, X.; Guo, C.; Dang, Z. Biosurfactant-enhanced phytoremediation of soils contaminated by crude oil using maize (Zea mays. L). Ecol. Eng. 2016, 92, 10–17. [Google Scholar] [CrossRef]
- Marchut-Mikołajczyk, O.; Drożdżyński, P.; Polewczyk, A.; Smułek, W.; Antczak, T. Biosurfactant from endophytic Bacillus pumilus 2A: Physicochemical characterization, production and optimization and potential for plant growth promotion. Microb. Cell Factories 2021, 20, 1–11. [Google Scholar] [CrossRef]
- Chopra, A.; Bobate, S.; Rahi, P.; Banpurkar, A.; Mazumder, P.B.; Satpute, S. Pseudomonas aeruginosa RTE4: A Tea Rhizobacterium With Potential for Plant Growth Promotion and Biosurfactant Production. Front. Bioeng. Biotechnol. 2020, 8, 861. [Google Scholar] [CrossRef]
- Alsohim, A.S.; Taylor, T.B.; Barrett, G.A.; Gallie, J.; Zhang, X.X.; Altamirano-Junqueira, A.E.; Johnson, L.J.; Rainey, P.B.; Jackson, R.W. The biosurfactant viscosin produced by Pseudomonas fluorescens SBW 25 aids spreading motility and plant growth promotion. Environ. Microbiol. 2014, 16, 2267–2281. [Google Scholar] [CrossRef]
- Droby, S. (Ed.) Food Security and Plant Disease Management; Elsevier Science & Technology: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Gill, H.K.; Garg, H. Pesticide: Environmental impacts and management strategies. Pestic. Toxic Asp. 2014, 8, 187. [Google Scholar]
- Mostafalou, S.; Abdollahi, M. Pesticides and human chronic diseases: Evidences, mechanisms, and perspectives. Toxicol. Appl. Pharmacol. 2013, 268, 157–177. [Google Scholar] [CrossRef]
- Preininger, C.; Sauer, U.; Bejarano, A.; Berninger, T. Concepts and applications of foliar spray for microbial inoculants. Appl. Microbiol. Biotechnol. 2018, 102, 7265–7282. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Yu, Q.; Ren, H.; Geng, J.; Xu, K.; Zhang, Y.; Ding, L. Towards physicochemical and biological effects on detachment and activity recovery of aging biofilm by enzyme and surfactant treatments. Biores. Technol. 2018, 247, 319–326. [Google Scholar] [CrossRef]
- Palma-Bautista, C.; Tataridas, A.; Kanatas, P.; Travlos, I.S.; Bastida, F.; Domínguez-Valenzuela, J.A.; De Prado, R. Can Control of Glyphosate Susceptible and Resistant Conyza sumatrensis Populations Be Dependent on the Herbicide Formulation or Adjuvants? Agronomy 2020, 10, 1599. [Google Scholar] [CrossRef]
- Buffington, E.J.; McDonald, S.K. Adjuvants & Surfactants; Colorado Environmental Pesticide Education Program: Fort Collins, CO, USA, 2006; Volume 301. [Google Scholar]
- Rebello, S.; Asok, A.K.; Mundayoor, S.; Jisha, M.S. Surfactants: Chemistry, Toxicity and Remediation. In Pollutant Diseases, Remediation and Recycling; Lichtfouse, E., Schwarzbauer, J., Robert, D., Eds.; Springer: Cham, Switzerland, 2013; Volume 4. [Google Scholar]
- Ryckaert, B.; Spanoghe, P.; Haesaert, G.; Heremans, B.; Isebaert, S.; Steurbaut, W. Quantitative determination of the influence of adjuvants on foliar fungicide residues. Crop Prot. 2007, 26, 1589–1594. [Google Scholar] [CrossRef]
- Jibrin, M.O.; Liu, Q.; Jones, J.B.; Zhang, S. Surfactants in plant disease management: A brief review and case studies. Plant Pathol. 2021, 70, 495–510. [Google Scholar] [CrossRef]
- Sachdev, D.P.; Cameotra, S.S. Biosurfactants in agriculture. Appl. Microbiol. Biotechnol. 2013, 97, 1005–1016. [Google Scholar] [CrossRef] [Green Version]
- Mizumoto, S.; Shoda, M. Medium optimization of antifungal lipopeptide, iturin A, production by Bacillus subtilis in solid-state fermentation by response surface methodology. Appl. Microbiol. Biotechnol. 2007, 76, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Crouzet, J.; Arguelles-Arias, A.; Dhondt-Cordelier, S.; Cordelier, S.; Pršić, J.; Hoff, G.; Mazeyrat-Gourbeyre, F.; Baillieul, F.; Clément, C.; Ongena, M.; et al. Biosurfactants in plant protection against diseases: Rhamnolipids and lipopeptides case study. Front. Bioeng. Biotechnol. 2020, 8, 8. [Google Scholar] [CrossRef]
- Kumar, A.; Zhimo, Y.; Biasi, A.; Salim, S.; Feygenberg, O.; Wisniewski, M.; Droby, S. Endophytic Microbiome in the Carposphere and Its Importance in Fruit Physiology and Pathology. In Postharvest Pathology; Spadaro, D., Droby, S., Gullino, M.L., Eds.; Springer: Cham, Switzerland, 2021; Volume 11. [Google Scholar]
- Singh, V.K.; Singh, A.K.; Kumar, A. Disease management of tomato through PGPB: Current trends and future perspective. 3 Biotech 2017, 7, 255. [Google Scholar] [CrossRef]
- Varnier, A.L.; Sanchez, L.; Vatsa, P.; Boudesocque, L.; Garcia-Brugger, A.N.G.E.L.A.; Rabenoelina, F.; Sorokin, A.; Renault, J.H.; Kauffmann, S.; Pugin, A.; et al. Bacterial rhamnolipids are novel MAMPs conferring resistance to Botrytis cinerea in grapevine. Plant Cell Environ. 2009, 32, 178–193. [Google Scholar] [CrossRef] [PubMed]
- Kruijt, M.; Tran, H.; Raaijmakers, J.M. Functional, genetic and chemical characterization of biosurfactants produced by plant growth-promoting Pseudomonas putida 267. J. Appl. Microbiol. 2009, 107, 546–556. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, T.H.; Sørensen, J. Production of cyclic lipopeptides by Pseudomonas fluorescens strains in bulk soil and in the sugar beet rhizosphere. Appl. Environ. Microbiol. 2003, 69, 861–868. [Google Scholar] [CrossRef] [Green Version]
- Perneel, M.; D’hondt, L.; De Maeyer, K.; Adiobo, A.; Rabaey, K.; Höfte, M. Phenazines and biosurfactants interact in the biological control of soil-borne diseases caused by Pythium spp. Environ. Microbiol. 2008, 10, 778–788. [Google Scholar] [CrossRef]
- D’aes, J.; Hua, G.K.H.; De Maeyer, K.; Pannecoucque, J.; Forrez, I.; Ongena, M.; Dietrich, L.E.; Thomashow, L.S.; Mavrodi, D.V.; Höfte, M. Biological control of Rhizoctonia root rot on bean by phenazine-and cyclic lipopeptide-producing Pseudomonas CMR12a. Phytopathology 2011, 101, 996–1004. [Google Scholar] [CrossRef] [Green Version]
- Velho, R.V.; Medina, L.F.C.; Segalin, J.; Brandelli, A. Production of lipopeptides among Bacillus strains showing growth inhibition of phytopathogenic fungi. Folia Microbiol. 2011, 56, 297. [Google Scholar] [CrossRef]
- Akladious, S.A.; Gomaa, E.Z.; El-Mahdy, O.M. Efficiency of bacterial biosurfactant for biocontrol of Rhizoctonia solani (AG-4) causing root rot in faba bean (Vicia faba) plants. Eur. J. Plant Pathol. 2019, 153, 15–35. [Google Scholar] [CrossRef]
- Hussain, T.; Haris, M.; Shakeel, A.; Ahmad, G.; Khan, A.A.; Khan, M.A. Bio-nematicidal activities by culture filtrate of Bacillus subtilis HussainT-AMU: New promising biosurfactant bioagent for the management of Root Galling caused by Meloidogyne incognita. Vegetos 2020, 33, 229–238. [Google Scholar] [CrossRef]
- Shalini, D.; Benson, A.; Gomathi, R.; Henry, A.J.; Jerritta, S.; Joe, M.M. Isolation, characterization of glycolipid type biosurfactant from endophytic Acinetobacter sp. ACMS25 and evaluation of its biocontrol efficiency against Xanthomonas oryzae. Biocatal. Agric. Biotechnol. 2017, 11, 252–258. [Google Scholar] [CrossRef]
- Haddad, N.I.; Wang, J.; Mu, B. Isolation and characterization of a biosurfactant producing strain, Brevibacilis brevis HOB1. J. Ind. Microbiol. Biotechnol. 2008, 35, 1597–1604. [Google Scholar] [CrossRef]
- Radhakrishnan, E.K.; Mathew, J. Characterization of biosurfactant produced by the endophyte Burkholderia sp. WYAT7 and evaluation of its antibacterial and antibiofilm potentials. J. Biotechnol. 2020, 313, 1–10. [Google Scholar]
- Kim, S.K.; Kim, Y.C.; Lee, S.; Kim, J.C.; Yun, M.Y.; Kim, I.S. Insecticidal activity of rhamnolipid isolated from Pseudomonas sp. EP-3 against green peach aphid (Myzus persicae). J. Agric. Food. Chem. 2011, 59, 934–938. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.E.; Caiazza, N.C.; O’Toole, G.A. Rhamnolipid surfactant production affects biofilm architecture in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2003, 185, 1027–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jonghe, K.; De Dobbelaere, I.; Sarrazyn, R.; Höfte, M. Control of Phytophthora cryptogea in the hydroponic forcing of witlo of chicory with the rhamnolipid-based biosurfactant formulation PRO1. Plant Pathol. 2005, 54, 219–222. [Google Scholar] [CrossRef] [Green Version]
- Hultberg, M.; Bengtsson, T.; Liljeroth, E. Late blight on potato is suppressed by the biosurfactant-producing strain Pseudomonas koreensis 2.74 and its biosurfactant. BioControl 2010, 55, 543–550. [Google Scholar] [CrossRef]
- Fernandes, P.A.; Arruda, I.R.; Santos, A.F.; Araújo, A.A.; Maior, A.M.; Ximenes, E.A. Antimicrobial activity of surfactants produced by Bacillus subtilis R14 against multidrug-resistant bacteria. Braz. J. Microbiol. 2007, 38, 704–709. [Google Scholar] [CrossRef] [Green Version]
- Kim, P.I.; Ryu, J.; Kim, Y.H.; Chi, Y.T. Production of biosurfactant lipopeptides Iturin A, fengycin and surfactin A from Bacillus subtilis CMB32 for control of Colletotrichum gloeosporioides. J. Microbiol. Biotechnol. 2010, 20, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Edosa, T.T.; Hun Jo, Y.; Keshavarz, M.; Soo Han, Y. Biosurfactants: Production and potential application in insect pest management. Trends Entomol. 2018, 14, 79–87. [Google Scholar] [CrossRef]
- Yun, D.C.; Yang, S.Y.; Kim, Y.C.; Kim, I.S.; Kim, Y.H. Identification of surfactin as an aphicidal metabolite produced by Bacillus amyloliquefaciens G1. J. Korean Soc. Appl. Biol. Chem. 2013, 56, 751–753. [Google Scholar] [CrossRef]
- Khedher, S.B.; Boukedi, H.; Dammak, M.; Kilani-Feki, O.; Sellami-Boudawara, T.; Abdelkefi-Mesrati, L.; Tounsi, S. Combinatorial effect of Bacillus amyloliquefaciens AG1 biosurfactant and Bacillus thuringiensis Vip3Aa16 toxin on Spodoptera littoralis larvae. J. Invertebr. Pathol. 2017, 144, 11–17. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Abdollahi, S.; Tofighi, Z.; Babaee, T.; Shamsi, M.; Rahimzadeh, G.; Rezvanifar, H.; Saeidi, E.; Mohajeri Amiri, M.; Saffari Ashtiani, Y.; Samadi, N. Evaluation of Antioxidant and Anti-biofilm Activities of Biogenic Surfactants Derived from Bacillus amyloliquefaciens and Pseudomonas aeruginosa. Iran. J. Pharm. Res. 2020, 19, 115–126. [Google Scholar] [PubMed]
- Amaro da Silva, I.; Gercyane Oliveira Bezerra, K.; Batista Durval, I.J.; Bronzo Barbosa Farias, C.; Galdino Junior, C.J.; Mendes Da Silva Santos, E.; Moura De Luna, J.; Asfora Sarubbo, L. Evaluation of the Emulsifying and Antioxidant Capacity of the Biosurfactant Produced by Candida Bombicola Urm 3718. Chem. Eng. 2020, 79, 67–72. [Google Scholar]
- Palanisamy, S.; Vinosha, M.; Marudhupandi, T.; Rajasekar, P.; Prabhu, N.M. Isolation of fucoidan from Sargassum polycystum brown algae: Structural characterization, in vitro antioxidant and anticancer activity. Int. J. Biol. Macromol. 2017, 102, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Markande, A.R.; Patel, D.; Varjani, S. A review on biosurfactants: Properties, applications and current developments. Biores. Technol. 2021, 330, 124963. [Google Scholar] [CrossRef]
- Sharma, D.; Singh Saharan, B. Simultaneous Production of Biosurfactants and Bacteriocins by Probiotic Lactobacillus casei MRTL3. Int. J. Microbiol. 2014, 2014, 698713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basit, M.; Rasool, M.H.; Naqvi, S.A.R.; Waseem, M.; Aslam, B. Biosurfactants production potential of native strains of Bacillus cereus and their antimicrobial, cytotoxic and antioxidant activities. Pak. J. Pharm. Sci. 2018, 31, 251–256. [Google Scholar]
- Ekramul, H.; Kayalvizhi, K.; Saqib, H. Biocompatibility, Antioxidant and Anti-Infective Effect of Biosurfactant Produced by Marinobacter litoralis MB15. Int. J. Pharm. Investig. 2020, 10, 173–178. [Google Scholar]
- Merghni, A.; Dallel, I.; Noumi, E.; Kadmi, Y.; Hentati, H.; Tobji, S.; Ben Amor, A.; Mastouri, M. Antioxidant and antiproliferative potential of biosurfactants isolated from Lactobacillus casei and their anti-biofilm effect in oral Staphylococcus aureus strains. Microb. Pathog. 2017, 104, 84–89. [Google Scholar] [CrossRef]
- Giri, S.S.; Ryu, E.C.; Sukumaran, V.; Park, S.C. Antioxidant, antibacterial, and anti-adhesive activities of biosurfactants isolated from Bacillus strains. Microb. Pathog. 2019, 132, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Ohadi, M.; Forootanfar, H.; Rahimi, H.R.; Jafari, E.; Shakibaie, M.; Eslaminejad, T.; Dehghannoudeh, G. Antioxidant Potential and Wound Healing Activity of Biosurfactant Produced by Acinetobacter junii B6. Curr. Pharm. Biotechnol. 2017, 18, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Morita, T.; Fukuoka, T.; Imura, T.; Kitamoto, D. Glycolipid biosurfactants, mannosylerythritol lipids, show antioxidant and protective effects against H2O2-induced oxidative stress in cultured human skin fibroblasts. J. Oleo Sci. 2012, 61, 457–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zouari, R.; Moalla-Rekik, D.; Sahnoun, Z.; Rebai, T.; Ellouze-Chaabouni, S.; Ghribi-Aydi, D. Evaluation of dermal wound healing and in vitro antioxidant efficiency of Bacillus subtilis SPB1 biosurfactant. Biomed. Pharmacother. 2016, 84, 878–891. [Google Scholar] [CrossRef]
- Mouafo, H.T.; Mbawala, A.; Somashekar, D.; Tchougang, H.M.; Harohally, N.V.; Ndjouenkeu, R. Biological properties and structural characterization of a novel rhamnolipid like-biosurfactants produced by Lactobacillus casei subsp. casei TM1B. Biotechnol. Appl. Biochem. 2021, 68, 585–596. [Google Scholar] [CrossRef]
- Li, S.; Huang, R.; Shah, N.P.; Tao, X.; Xiong, Y.; Wei, H. Antioxidant and antibacterial activities of exopolysaccharides from Bifidobacterium bifidum WBIN03 and Lactobacillus Plantarum R315. J. Dairy Sci. 2014, 97, 7334–7343. [Google Scholar] [CrossRef] [Green Version]
- Jemil, N.; Ben Ayed, H.; Manresa, A.; Nasri, M.; Hmidet, N. Antioxidant properties, antimicrobial and anti-adhesive activities of DCS1 lipopeptides from Bacillus methylotrophicus DCS1. BMC Microbiol. 2017, 17, 144. [Google Scholar] [CrossRef] [Green Version]
- Ceresa, C.; Fracchia, L.; Fedeli, E.; Porta, C.; Banat, I.M. Recent Advances in Biomedical, Therapeutic and Pharmaceutical Applications of Microbial Surfactants. Pharmaceutics 2021, 13, 466. [Google Scholar] [CrossRef]
- Fariq, A.; Yasmin, A. Production, characterization and bioactivities of biosurfactants from newly isolated strictly halophilic bacteria. Process. Biochem. 2020, 98, 1–10. [Google Scholar] [CrossRef]
- Kharat, M.; McClements, D.J. Fabrication and characterization of nanostructured lipid carriers (NLC) using a plant-based emulsifier: Quillaja saponin. Food Res. Int. 2019, 126, 108601. [Google Scholar] [CrossRef]
- DOH and DEFRA. UK Five-Year Antimicrobial Resistance Strategy 2013 to 2018. 2013. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/244058/20130902_UK_5_year_AMR_strategy.pdf (accessed on 12 February 2019).
- Dıaz De Rienzo, M.A.; Banat, I.M.; Dolman, B.; Winterburn, J.; Martin, P.J. Sophorolipid biosurfactants: Possible uses as antibacterial and antibiofilm agent. New Biotechnol. 2015, 32, 720–726. [Google Scholar] [CrossRef]
- Foschi, C.; Salvo, M.; Cevenini, R.; Parolin, C.; Vitali, B.; Marangoni, A. Vaginal Lactobacilli reduce Neisseria gonorrhoeae viability through multiple strategies: An in vitro study. Front. Cell. Infect. Microbiol. 2017, 7, 502. [Google Scholar] [CrossRef]
- Morais, I.M.C.; Cordeiro, A.L.; Teixeira, G.S.; Domingues, V.S.; Nardi, R.M.D.; Monteiro, A.S.; Alves, R.J.; Siqueira, E.P.; Santos, V.L. Biological and physicochemical properties of biosurfactants produced by Lactobacillus jensenii P 6A and Lactobacillus gasseri P 65. Microb. Cell Fact. 2017, 16, 1–15. [Google Scholar] [CrossRef]
- Dusane, D.H.; Nancharaiah, Y.V.; Zinjarde, S.S.; Venugopalan, V.P. Rhamnolipid mediated disruption of marine Bacillus pumilus biofilms. Colloids Surf. B Biointerfaces 2010, 81, 242–248. [Google Scholar] [CrossRef]
- Kaczorek, E. Effect of external addition of rhamnolipids biosurfactant on the modification of gram positive and gram negative bacteria cell surfaces during biodegradation of hydrocarbon fuel contamination. Pol. J. Environ. Stud. 2012, 21, 901–909. [Google Scholar]
- Bharali, P.; Konwar, B.K. Production and physicochemical characterization of abiosurfactant produced by Pseudomonas aeruginosa OBP1 isolated from petroleum sludge. Appl. Biochem. Biotechnol. 2011, 164, 1444–1460. [Google Scholar] [CrossRef]
- Naughton, P.J.; Marchant, R.; Naughton, V.; Banat, I.M. Microbial biosurfactants: Current trends and applications in agricultural and biomedical industries. J. Appl. Microbiol. 2019, 127, 12–28. [Google Scholar] [CrossRef] [Green Version]
- Gudina, E.J.; Rangarajan, V.; Sen, R.; Rodrigues, L.R. Potential therapeutic applications of biosurfactants. Trends Pharmacol. Sci. 2013, 34, 667–675. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Yan, L.; Xu, X.; Jiang, C.; Shi, J.; Zhang, Y.; Liu, L.; Lei, S.; Shao, D.; Huang, Q. Potential of Bacillus subtilis lipopeptides in anti-cancer I: Induction of apoptosis and paraptosis and inhibition of autophagy in K562 cells. AMB Express 2018, 8, 1–16. [Google Scholar] [CrossRef]
- Dey, G.; Bharti, R.; Dhanarajan, G.; Das, S.; Dey, K.K.; Kumar, B.N.; Sen, R.; Mandal, M. Marine lipopeptide Iturin A inhibits Akt mediated GSK3b and FoxO3asignalling and triggers apoptosis in breast cancer. Sci. Rep. 2015, 5, 10316. [Google Scholar] [CrossRef] [Green Version]





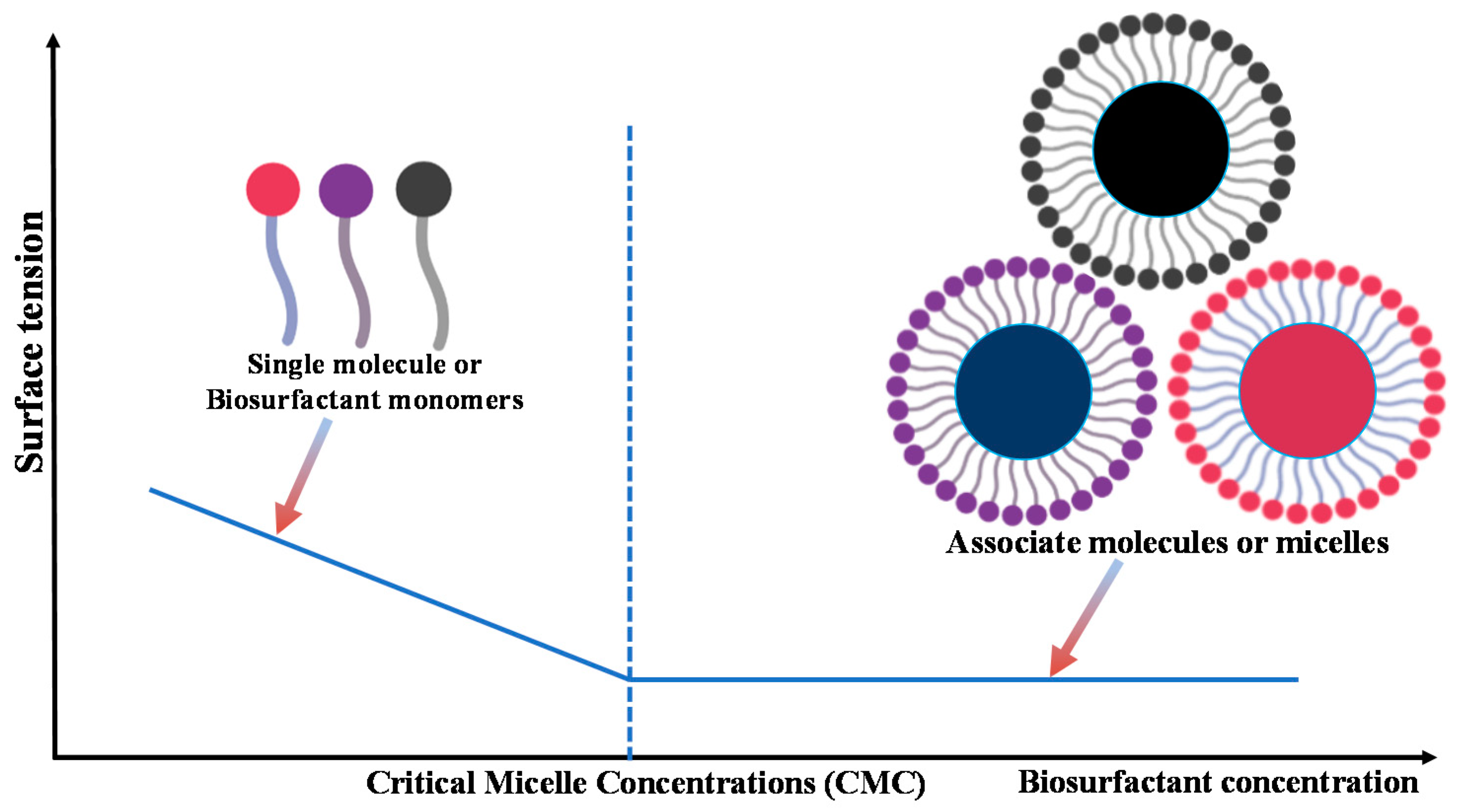{kind=link}
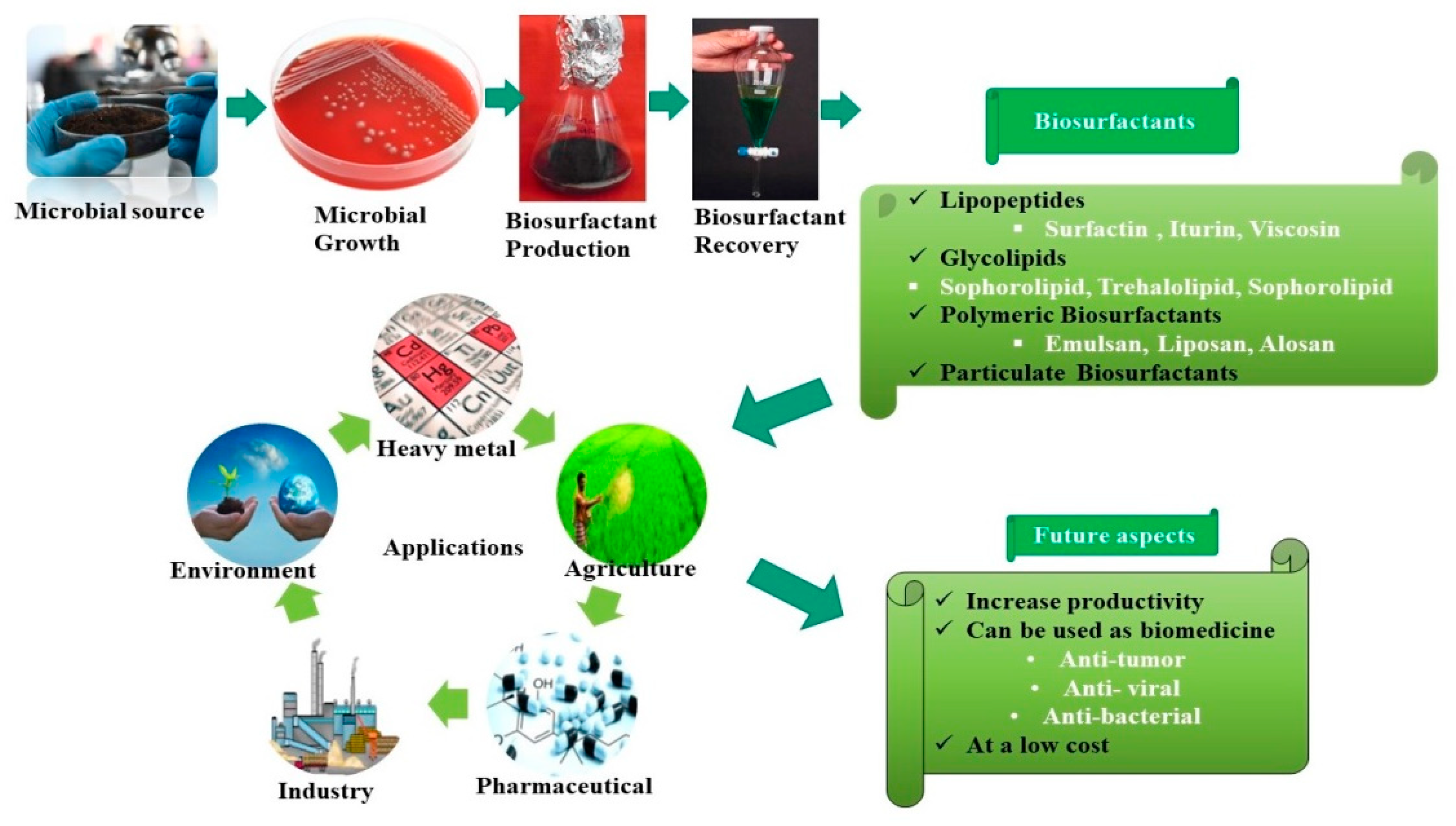{kind=link}
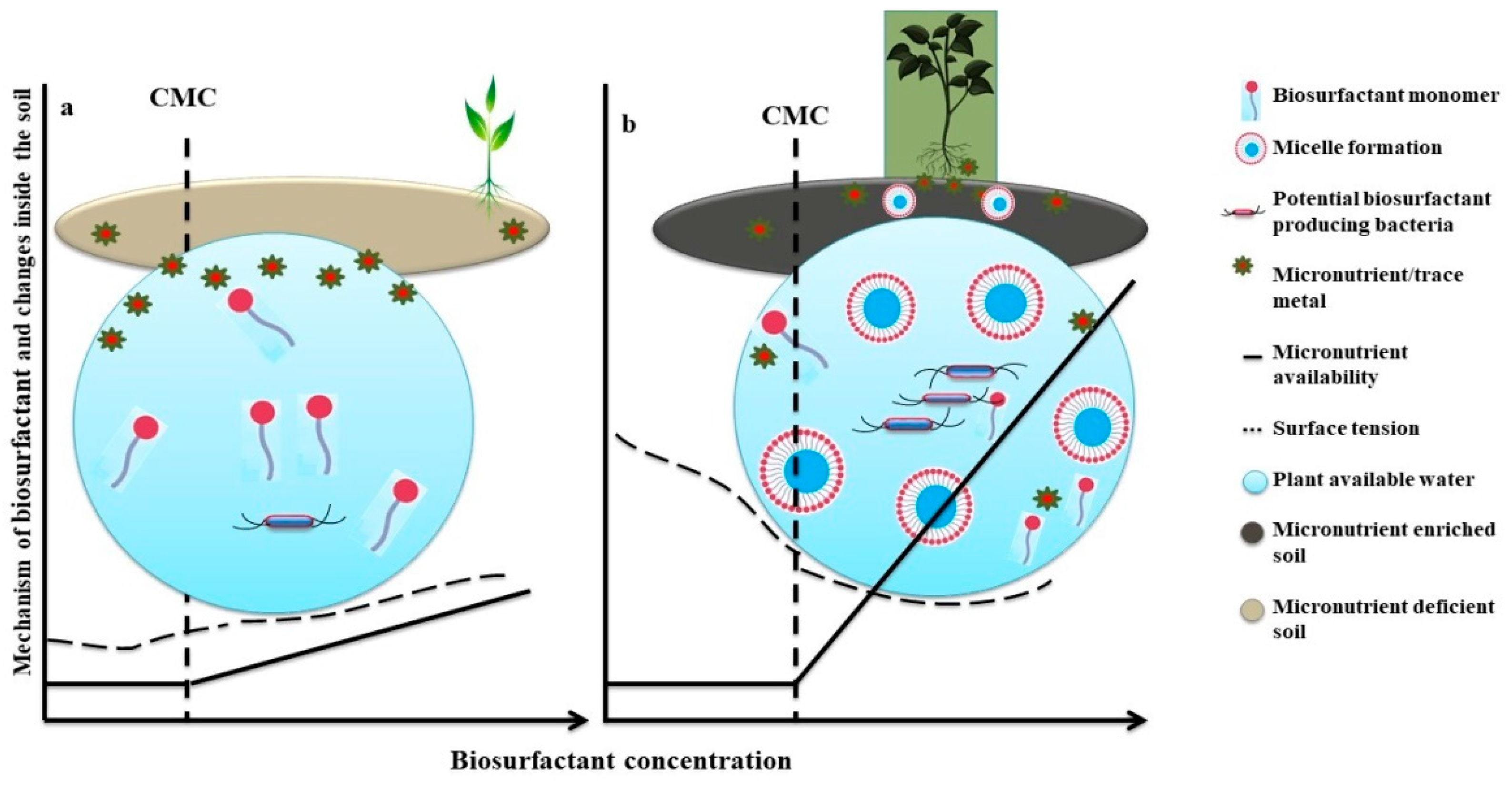{kind=link}
{kind=link}
| Types of Biosurfactants | Common Examples | |
|---|---|---|
| Low molecular mass | Glycolipids | Rhamnolipids |
| Sophorolipids | ||
| Trehalose lipids | ||
| Phospholipids | Phospholipids | |
| Corinomiocolic acid | ||
| Fatty acids | ||
| Lipopeptides | Surfactin | |
| Wisconsin | ||
| Gramicidin | ||
| Subtilisin | ||
| Peptide lipid | ||
| Lichenysin | ||
| High molecular mass | Polymeric | Liposan |
| Emulsan | ||
| Biodispersion | ||
| Mannan-lipid protein | ||
| Carbohydrate lipid-protein | ||
| Particulate | Vesicles |
| (A). Different Types of Biosurfactants Produced from Bacterial Strains | ||||
| Bacterial Strains | Biosurfactants | Properties | Isolation Source | References |
| Pontibacter korlensis strain SBK-47 | Pontifactin | Surface-active, antimicrobial, and antibiofilm activities | Coastal waters of Karaikal, Puducherry, India | [23] |
| Bacillus licheniformis | Lipopeptides | Heat resistance and capacity to emulsify oils used in cosmetics | Deception Island (Antarctica) | [24] |
| Paracoccus sp. MJ9 | Rhamnolipid | Enhance solubility of hydrophobic compounds | Jiaozhou Bay in Qingdao, Shandong Province | [25] |
| Pseudomonas aeruginosa UCP0992 | Rhamnolipids | High emulsifying activities against different oils, capacity to remove hydrophobic contaminants, and did not show toxicity | Centre of Research in Environmental Sciences, Catholic University of Pernambuco, Brazil | [26] |
| Pseudomonas aeruginosa PA1 | Rhamnolipid | Capacity to use as carbon sources | Oil production wastewater in the northeast of Brazil | [27] |
| Pseudomonas desmolyticum NCIM 2112 | Rhamnolipid | Degradation of textile dye | National Center for Industrial Microorganisms (NCIM), Pune, India | [28] |
| Serratia marcescens SS-1 | Serrawettins | Produces lipopeptide surfactants, having the capability to reduce surface tension | Taiwan | [29] |
| Bacillus subtilis | Cyclic lipopeptides | A significant reduction in the activities of acetylcholinesterase, a-carboxylesterase, and acid phosphatases | Namakkal and Tirunelveli district, Tamil Nadu, India | [30] |
| Bacillus subtilis | Pumilacidin | Antiviral activity against Herpes simplex virus 1 (HSV-1) | Tree trunk near lake Yamanaka, Japan | [31] |
| Pseudomonas aeruginosa S5 | Glycolipid | Removal of polycyclic aromatic hydrocarbons | Supelco (Bellefonte, PA, USA) | [32] |
| Pseudomonas protegens F6 | Orfamide A | Insecticidal against Myzus persicae | Soil from previously reported diesel oil-contaminated site | [33] |
| Pseudomonas aeruginosa DS9 | Rhamnolipid | Antifungal agents against F. sacchari in pokkah boeng Disease | Lakota oil-field of Sivsagar district, Assam, India | [34] |
| Pseudomonas fluorescens BD5 | Pseudofactin II | Antiadhesive activity and disinfectant | Freshwater from the Arctic Archipelago of Svalbard | [35] |
| Bacillus sp. BS3 | Lipopeptide | Anticancer activity and antiviral properties | Solar salt works in Tamilnadu, India | [36] |
| Pseudomonas aeruginosa | Rhamnolipid | Enhanced oil recovery through anaerobic production of Rhamnolipid | Daqing oilfield-produced water | [37] |
| Bacillus subtilis A21 | Lipopeptide | Removal of petroleum hydrocarbons, heavy metals | Adityapur Industrial Area, Jharkhand | [38] |
| Rhodotorula bogoriensis | Sophorolipid | Antimicrobial property against Propionibacterium acnes | American Type Culture Collection | [39] |
| (B). Different Types of Biosurfactants Produced from Fungal Strains | ||||
| Fungi | Biosurfactants | Properties | Isolation Source | References |
| Candida utilis | Emulsifiers | Emulsifiers | Culture collection from the Department of Antibiotics of the Universidade Federal de, Pernambuco, Brazil | [40] |
| Candida lipolytica UCP 0988 | Lipopeptide | Not toxic against different vegetable seed | Culture collection of Nucleus of Research in Environmental Sciences, Catholic University of Pernambuco, Recife-PE, Brazil | [41] |
| Penicillium chrysogenum SNP5 | Lipopeptide | Role in pharmaceuticals as well as in the petroleum and oil industry | Soil-contaminated grease waste | [42] |
| Cunninghamella echinulata | Complex Carbohydrate/protein/lipid | Reduce and increase the viscosity of hydrophobic substrates and their molecules | Caatinga soil of Pernambuco, Northeast of Brazil | [43] |
| Candida Antarctica | Mixtures of 4 mannosylerythritol lipids | Produced the lipids from different vegetable oils | Centraalbureau voor Schimmelcultures, the Netherlands | [44] |
| Microsphaeropsis sp. | Eremophilane derivative | Antimicrobial properties | Waters around the Caribbean Island of Dominica | [45] |
| Yarrowia lipolytica NCIM 3589 | Bioemulsifier | Increased the hydrophobicity of the cells during the growth phase | Seawater near Mumbai, India | [46] |
| Yarrowia lipolytica IMUFRJ50682 | Carbohydrate protein complex | Capable of stabilizing oil-in-water emulsions | Guanabara Bay in Rio de Janeiro | [47] |
| Ustilago maydis | Cellobiose lipids | Secreted cellobiose lipid having antifungal activity | - | [48] |
| Torulopsis bombicola | Sophorose lipid | Sophorose lipid fermentation | American Type Culture Collection | [49] |
| Aspergillus ustus | Glycolipoprote | Antimicrobial activity | Peninsular coast of India | [50] |
| Candida bombicola ATCC 22214 | Sophorolipid | Used in low-end consumer products and household application | American Type Culture Collection | [51] |
| Ustilago maydis FBD12 | Glycolipids | Antimicrobial activity | American Type Culture Collection | [52] |
| (C). Different Types of Biosurfactants Produced from Yeast Strains | ||||
| Yeast | Biosurfactants | Properties | Isolation source | References |
| Starmerella bombicola | Sophorolipids | Cytotoxic effect on MDA-MB-321 breast cancer cell line | Fungal Biodiversity Centre | [53] |
| Torulopsis Petrophilum ATCC 20225 | Glycolipids | Protein emulsifier | American Type Culture Collection | [54] |
| Kluyveromyces marxianus FII 510700 | Mannanoprotein | Source of emulsifier in the food industry | Culture Collection of the University of New South Wales, UNSW | [55] |
| Pseudozyma aphids, DSM 70725 and DSM 14930 | Mannosylerythritol lipids | Foam formation | Deutsche Stammsammlung von Mikroorganismen und Zellkulturen (DSMZ), Braunschweig, Germany | [56] |
| Pseudozyma tsukubaensis | Glycolipid | Producing diastereomer MEL-B from vegetable oils | Leaves of Perilla frutescens on Ibaraki in Japan | [57] |
| Saccharomyces cerevisiae URM 6670 | Glycolipid | Antioxidant activity and cytotoxic potential | Culture Collection of the Department of Antibiotics of the Federal University of Pernambuco (Brazil) | [58] |
| Trichosporon asahii | Sophorolipid | Efficient degrader of diesel oil, higher hydrophobicity, emulsification activity, and surface tension reduction | Petroleum hydrocarbon-contaminated soil in India | [59] |
| Meyerozyma guilliermondii YK32 | Sophorolipid | Emulsification properties | Soil samples collected from hydrocarbon-polluted locations of Hisar, Haryana | [60] |
| Rhodotorula babjevae YS3 | Sophorolipid | Antimicrobial activity | Agricultural field in Assam, Northeast India | [61] |
| Pichia caribbica | Xylolipid | Reduced the surface the tension of distilled water | Microbial type culture collection, India | [62] |
| Candida ishiwadae Y12 | Monoacylglycerols: Glycolipid | Exhibited high surfactant activities | Plant material in Thailand | [63] |
| Substrate | Conc. (gL−1) | Microorganisms | Yield (gL−1) | References |
|---|---|---|---|---|
| Glucose | 40 | P. aeruginosa | 0.3 | [72] |
| 40 | B. subtilis | 3.6 | [73] | |
| 40 | B. subtilis | 0.72 | [74] | |
| 30 | B. pumilus | 0.72 | [75] | |
| 20 | P. aeruginosa | 3.88 | [76] | |
| 10 | B. subtilis | 0.16 | [77] | |
| - | Pseudomonas sp. | 0.35 | [78] | |
| Sucrose | 20 | P. putida | 1.30 | [79] |
| Glucose and fructose | 16.55 | B. subtilis | 0.93 | [80] |
| Glucose + Yeast extract | 1:3 | Bacillus sp. | 2.56 | [81] |
| Glycerol + yeast extract | 30:5 | P. aeruginosa | 2.7 | [82] |
| Yeast extract | 1 | P. taiwanensis | 1.12 | [83] |
| Yeast extract | 2 | Bacillus sp. | 2.5 | [84] |
| NaNO3 | 0.2 M | P. aeruginosa | 2.73 | [85] |
| NaNO3 | 5 | B. subtilis | 1.12 | [86] |
| Peptone | 4 | Serratia marcescens | 1.2 | [81] |
| NH4NO3 | 1 | P. fluorescens | 2 | [87] |
| Microorganism | Alternative Media Source | Yield and Properties | References |
|---|---|---|---|
| Bacillus subtilis ATCC 6051 | Brewery waste (trub) | The product yield of 100.76 mgL−1 | [96] |
| Bacillus subtilis PC | Sugar cane vinasse | Able to reduce surface tension 32 mN m−1 and the E24 to 51.10%. | [97] |
| Bacillus subtilis | Corn steep liquor | Biosurfactant yields 1.3 gL−1; the different yields increased (up to 4.1, 4.4, and 3.5 g/L for iron, manganese, and magnesium supplements, respectively). However, at the optimum concentration, the yield of these three metals increased up to 4.8 gL−1. | [98] |
| Bacillus subtilis MTCC 2423 | Rice mill polishing residue | Surfactin yield 4.17 g kg−1 residue | [99] |
| Bacillus licheniformis AL1.1 | Molasses | Lichenysin yield of 3·2 gL−1 | [100] |
| Bacillus pseudomycoides | Soybean oil waste | C.M.C. of lipopeptide 56 mgL−1 and able to reduce the surface tension of water from 71.6 mN m−1 to 30.2 mN m−1 | [101] |
| Bacillus subtilis DSM 3256 | Two-phase olive mill waste | Surfactin yields 0.068 g g−1, and the surface tension of the culture medium is reduced to 30.1 ± 0.9 mN m−1. | [102] |
| Bacillus subtilis | Rapeseed cake | Surfactin analogues | [103] |
| Bacillus amyloliquefaciens | Distillers’ grains | Surfactin yield 1.04 gL−1 | [104] |
| Bacillus nealsonii S2MT | Glycerol 2% (v/v) and NH4NO3 0.1% (w/v) | The maximum biosurfactant yield was 1300 mg/L and reduced the surface tension (34.15 ± 0.6 mN/m). Additionally, highly stable at environmental factors such as salinity, pH and temperature variations. | [105] |
| Staphylococcus sp. | Residual frying oil, expired milk | The C.M.C. of the purified lipopeptides was 65–750 mg/L, depending upon carbon source. Additionally, it was stable within a broad range of pH, temperature, and salinity values. | [106] |
| Halomonas venusta PHKT | Glycerol | Surfactin, Pumilacidin, and Bios-PHKT have a critical micelle concentration (C.M.C.) of 125 mgL−1 and showed a high steadiness against a broad spectrum of salinity (0–120 gL−1 NaCl), temperature (4–121 °C), and pH values (2–12). | [107] |
| Rhodotorula sp. | Olive oil mills | Potent biosurfactant producer with E24 = 69% and a significant reduction in S.T. from 72 to 35 mN m−1. In addition, it showed stability over a wide range of pH (2–12), temperature (4–100 °C), and salinity values (1–10%). | [108] |
| Volvariella volvacea | Edible paddy straw mushroom | Biosurfactant effectively showed a reduction in the surface tension, emulsification index, and oil spreading activity of 35.15 dyne/cm, 80%, and 11 cm, respectively. | [109] |
| Microorganism | Biosurfactant | Properties | Reference |
|---|---|---|---|
| Pseudomonas sp. EP-3 | Rhamnolipid | Insecticidal activity | [155] |
| Pseudomonasaeruginosa PAO1 | Rhamnolipid | Biofilm formation | [156] |
| Pseudomonas aeruginosa | Rhamnolipids | Control of Phytophthora cryptogea | [157] |
| Pseudomonas aeruginosa | Rhamnolipids | Resistance to Botrytis cinerea in grapevine | [144] |
| Pseudomonas putida | Biosurfactants | Zoospores of the oomycete pathogen Phytophthora capsici | [145] |
| Pseudomonas koreensis | Biosurfactant | Late blight on potato | [158] |
| Acinetobacter sp. ACMS25 | Glycolipid | Biocontrol of Xanthomonas oryzae | [152] |
| Burkholderia sp. WYAT7 | Glycolipid | Antibacterial and ant- biofilm potentials | [154] |
| Bacillus licheniformis | Biosurfactant | Biocontrol of Rhizoctonia solani causing root rot in faba bean | [150] |
| Pseudomonas CMR12a | Lipopeptides | Biological control of Rhizoctonia root rot on bean | [148] |
| Brevibacillus brevis | Lipopeptides | Antibacterial and Antifungal properties | [153] |
| Bacillus sp. | Lipopeptides | Growth inhibition of Fusarium spp., Aspergillus spp., and Biopolaris sorokiniana | [149] |
| Bacillus subtilis R14 | Lipopeptide | Antimicrobial activity | [159] |
| Bacillus subtilis | Lipopeptides Iturin A, fengycin, and surfactin | Colletotrichum gloeosporioides, the causative agent for anthracnose on papaya leaves | [160] |
| Source | Chemical Nature of Biosurfactant | Antioxidant Activity Assessment | Antioxidant | Antibacterial | Antiproliferative | Reference |
|---|---|---|---|---|---|---|
| Lactobacillus casei subsp. casei TM1B | Rhamnolipid-like biosurfactant | DPPH (1-diphenyl-2-picrylhydrazyl) assay, ABTS (2.2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) assay | yes | yes | not tested | [177] |
| Pseudomonas aeruginosa MN1 | Rhamnolipid | FRAP and DPPH assay | yes | yes | not tested | [165] |
| Bacillus amyloliquefaciens NS6 | Surfactin | Ferric reducing antioxidant power (FRAP) and DPPH assay | yes | yes | not tested | [165] |
| Marinobacter litoralis MB15 | Rhamnolipid | DPPH assay | yes | yes | yes | [171] |
| Halomonas elongata, Halobacillus karajiensis and Alkalibacillus almallahensis | Glycolipid | DPPH assay | yes | yes | not tested | [178] |
| Bacillus subtilis VSG4 | Lipopeptide | DPPH assay | yes | yes | not tested | [173] |
| Bacillus licheniformis VS16 | Phospholipopeptide | DPPH assay | yes | yes | not tested | [173] |
| Lactobacillus casei (BS-LBl) | Not mentioned | DPPH assay | yes | yes | yes | [172] |
| Acinetobacter junii B6 | Lipopeptide | DPPH and FRAP assay | yes | yes | not tested | [174] |
| Bifidobacterium bifidum WBIN03 and Lactobacillus plantarum R315 | Exo polysaccharides | DPPH assay and superoxide and hydroxy radical estimation | yes | yes | not tested | [180] |
| Bacillus methylotrophicus DCS1 | Lipopeptide | DPPH assay | yes | yes | not tested | [179] |
| Pseudozyma hubeiensis | Mannosylerythritol lipids | DPPH assay | yes | not tested | yes | [175] |
| Bacillus subtilis SPB1 | Lipopeptide | DPPH assay | yes | not tested | yes | [176] |
| Bacillus cereus MMIC | Lipopeptide | DPPH assay | yes | yes | yes | [170] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, A.; Singh, S.K.; Kant, C.; Verma, H.; Kumar, D.; Singh, P.P.; Modi, A.; Droby, S.; Kesawat, M.S.; Alavilli, H.; et al. Microbial Biosurfactant: A New Frontier for Sustainable Agriculture and Pharmaceutical Industries. Antioxidants 2021, 10, 1472. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10091472
Kumar A, Singh SK, Kant C, Verma H, Kumar D, Singh PP, Modi A, Droby S, Kesawat MS, Alavilli H, et al. Microbial Biosurfactant: A New Frontier for Sustainable Agriculture and Pharmaceutical Industries. Antioxidants. 2021; 10(9):1472. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10091472
Chicago/Turabian StyleKumar, Ajay, Sandeep Kumar Singh, Chandra Kant, Hariom Verma, Dharmendra Kumar, Prem Pratap Singh, Arpan Modi, Samir Droby, Mahipal Singh Kesawat, Hemasundar Alavilli, and et al. 2021. "Microbial Biosurfactant: A New Frontier for Sustainable Agriculture and Pharmaceutical Industries" Antioxidants 10, no. 9: 1472. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10091472