
Impact of Selective Renal Afferent Denervation on Oxidative Stress and Vascular Remodeling in Spontaneously Hypertensive Rats

 and
and 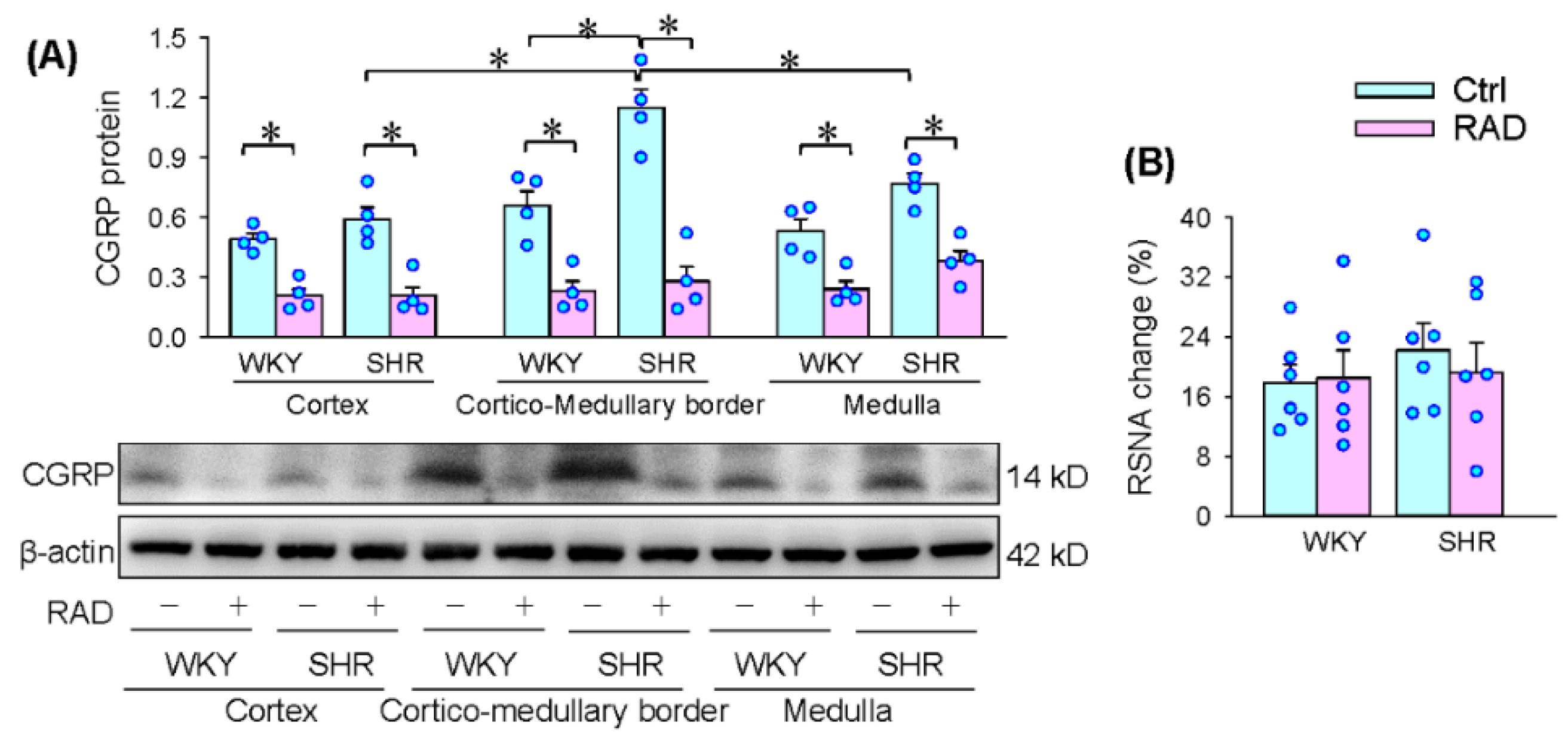{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Renal Afferent Denervation
2.3. Identification of RAD
2.4. Measurement of Blood Pressure and Heart Rate
2.5. Examination of Plasma Norepinephrine Level and Heart Rate Variability
2.6. Measurement of NOX Activity and Superoxide Production
2.7. In Situ Detection of Superoxide Anions in the PVN
2.8. Masson’s Staining
2.9. Western Blot and Antibodies
2.10. Examination of Aldosterone, Potassium and Sodium Levels in Serum and Urine
2.11. Statistics
3. Results
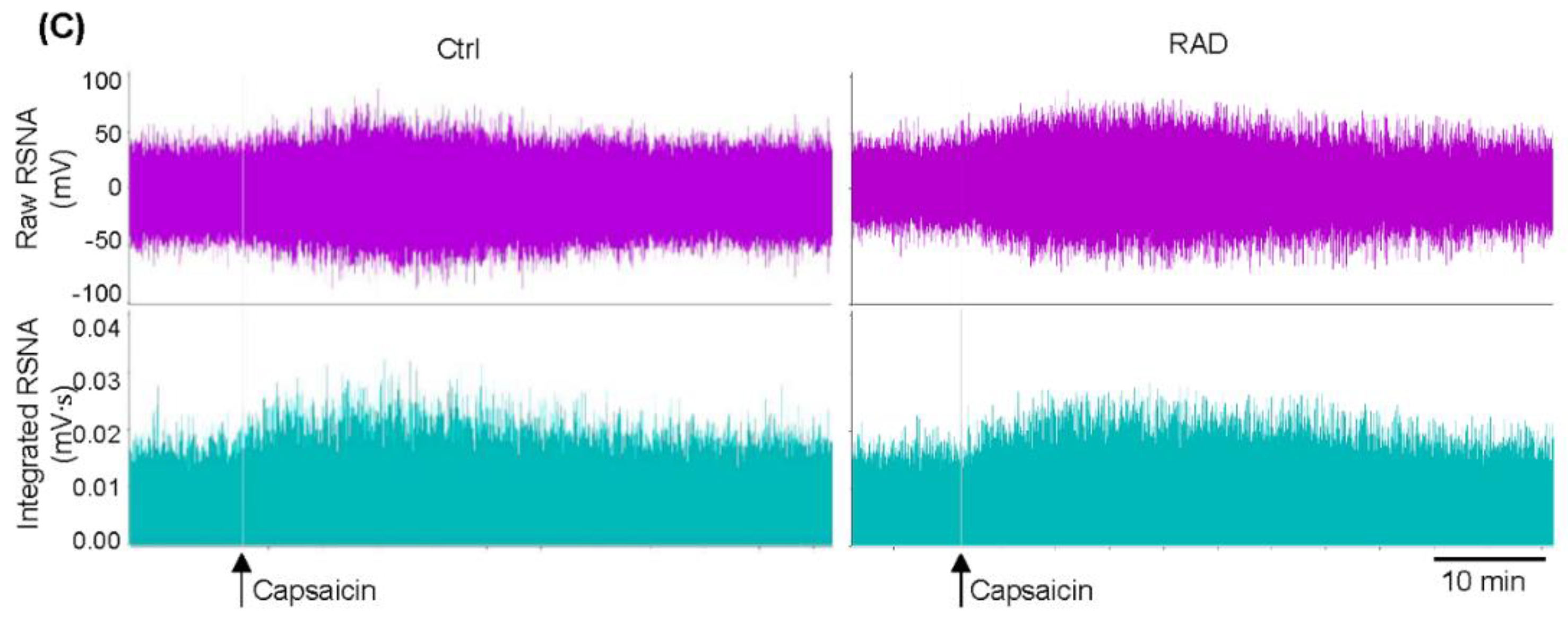
3.1. Identification of the RAD Reliability
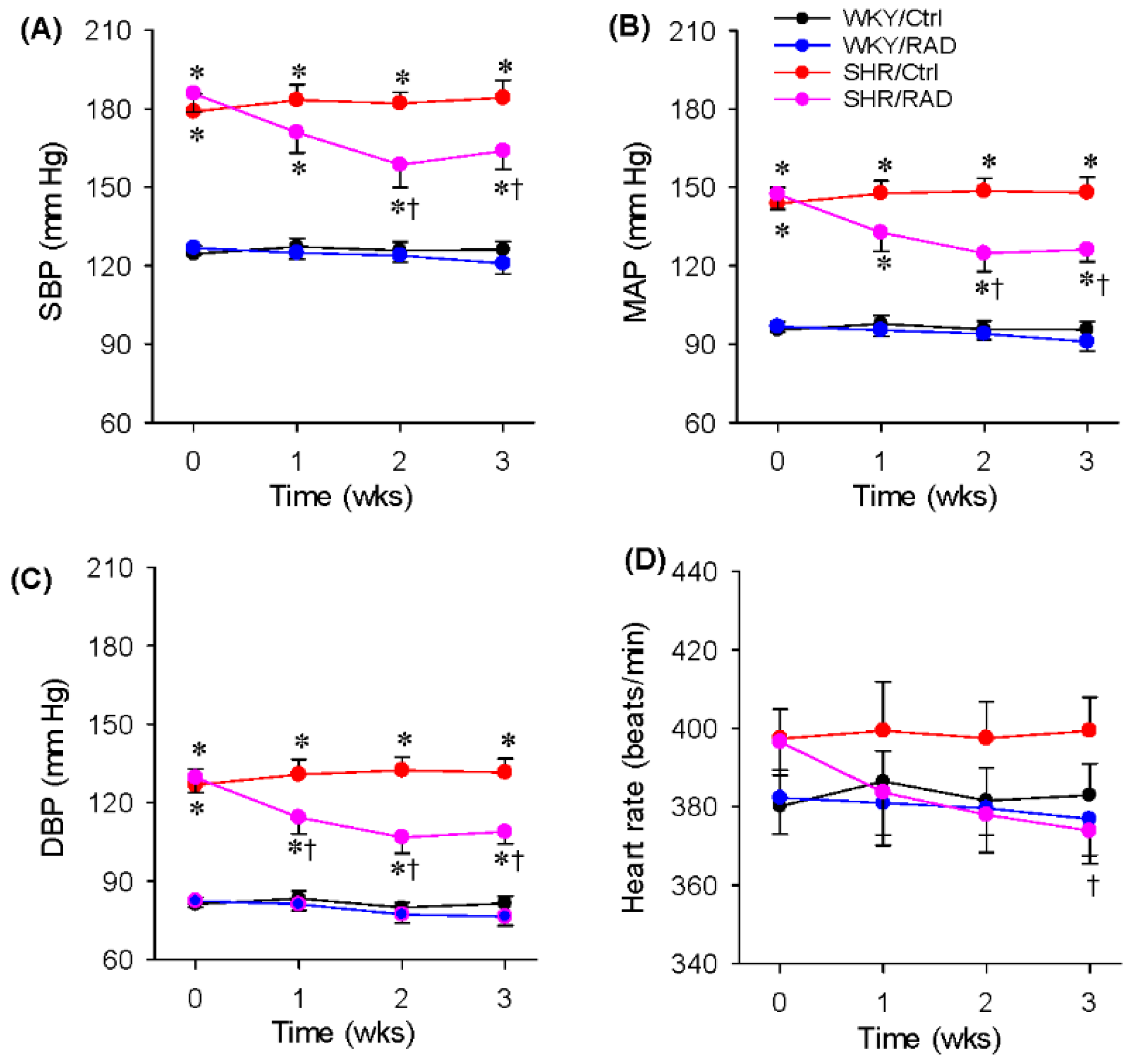
3.2. Blood Pressure and Heart Rate
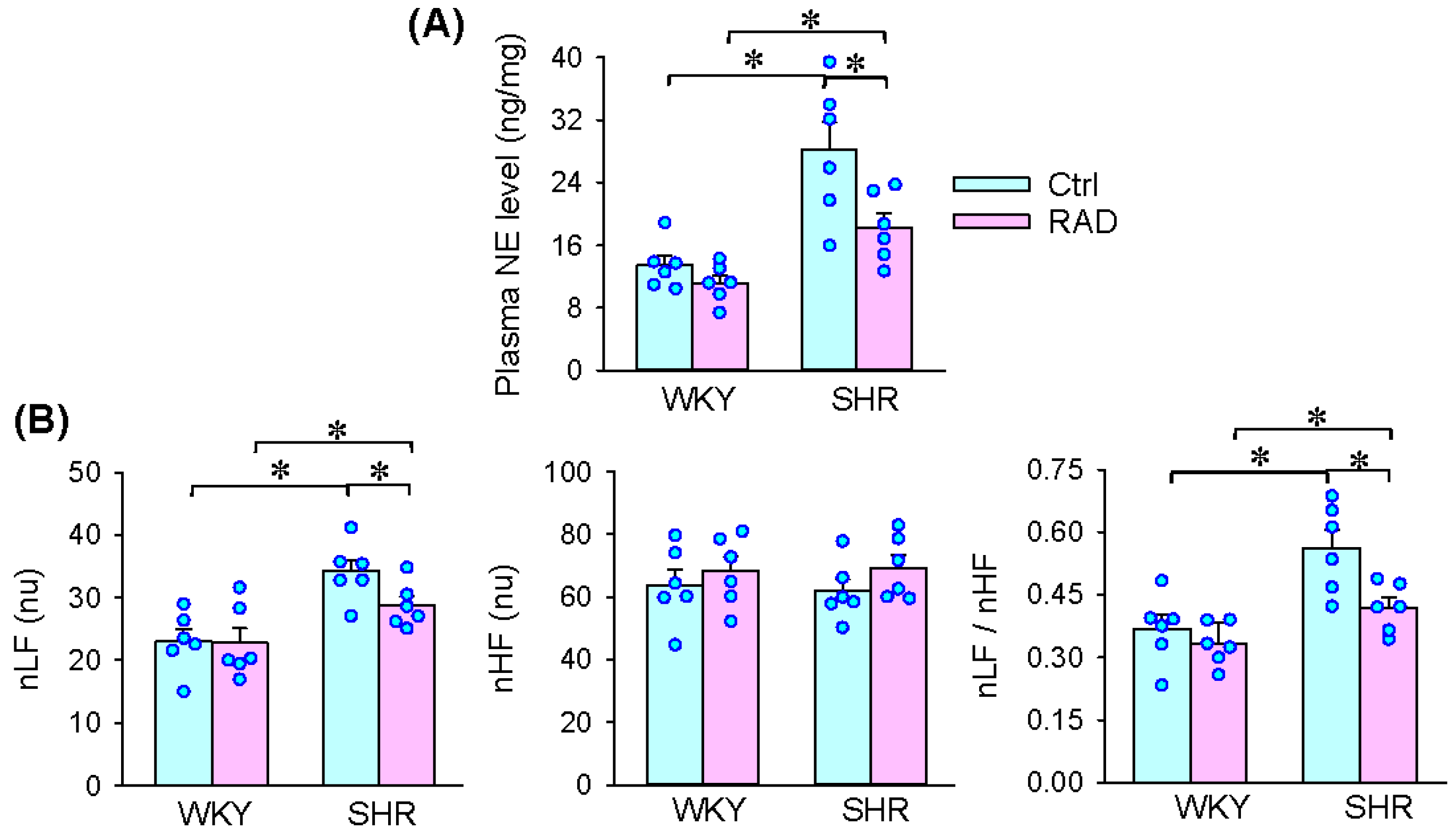
3.3. Plasma NE Level and HRV
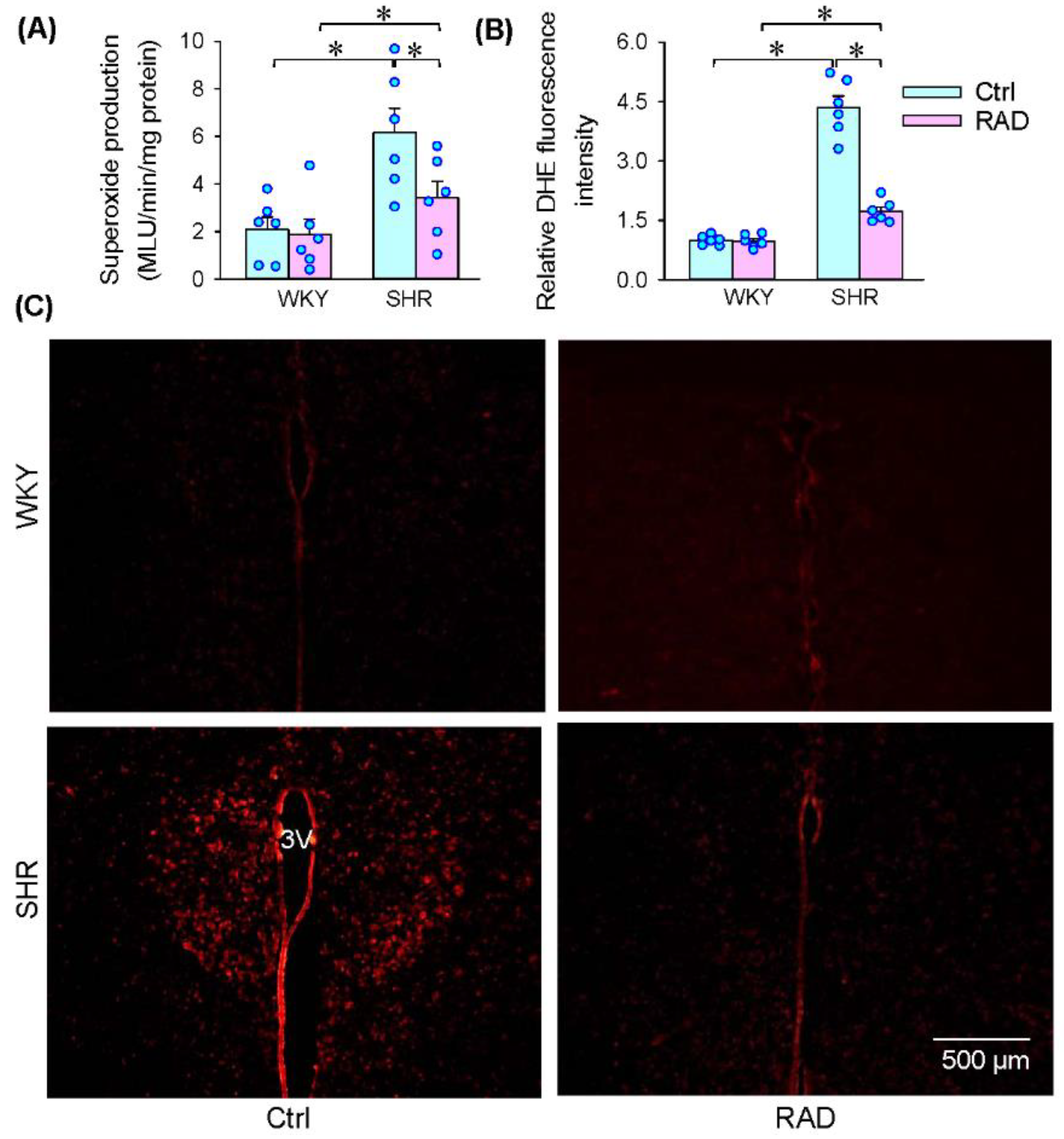
3.4. Superoxide Production in the PVN
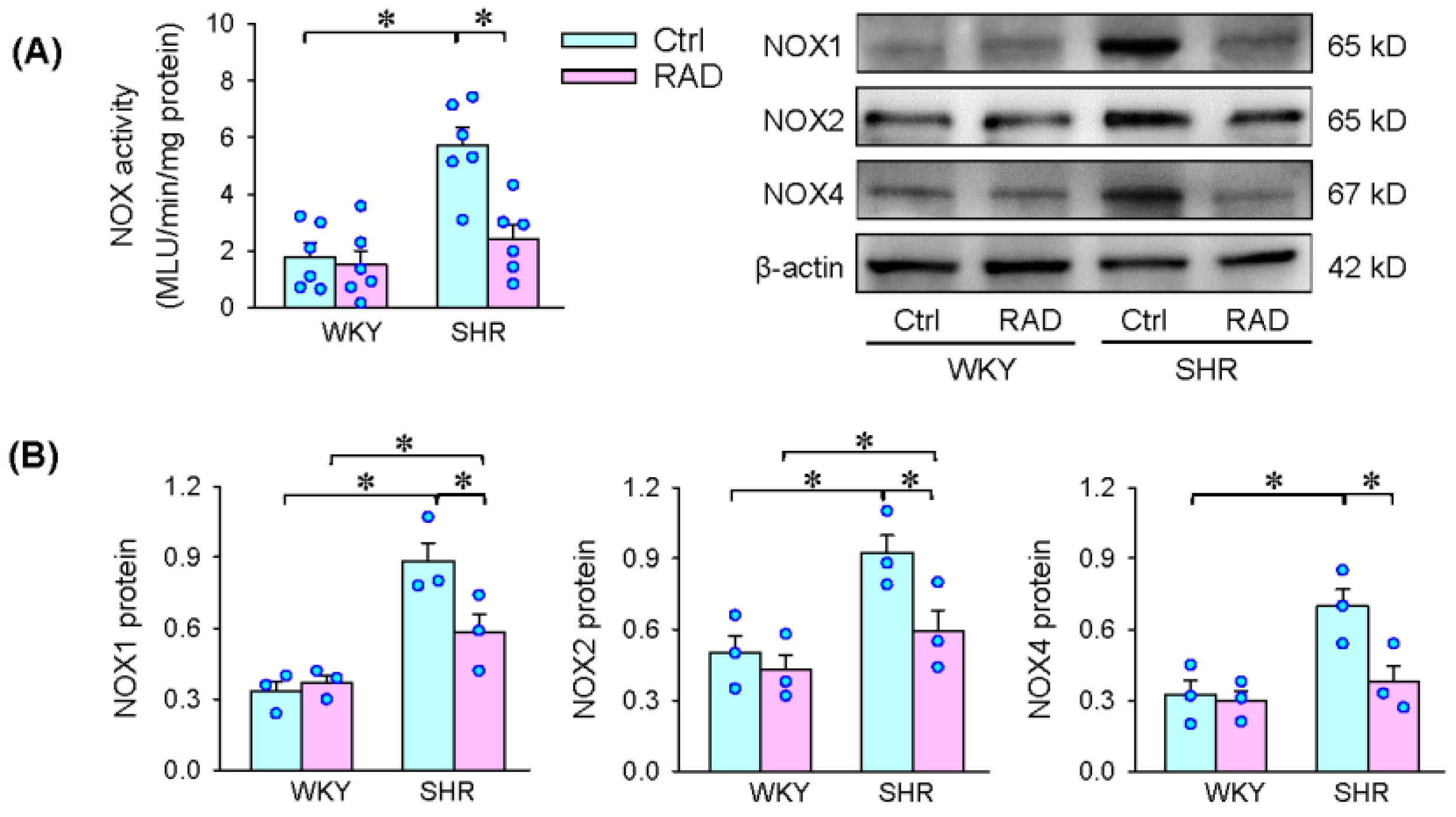
3.5. NOX Activity and Expression in the PVN
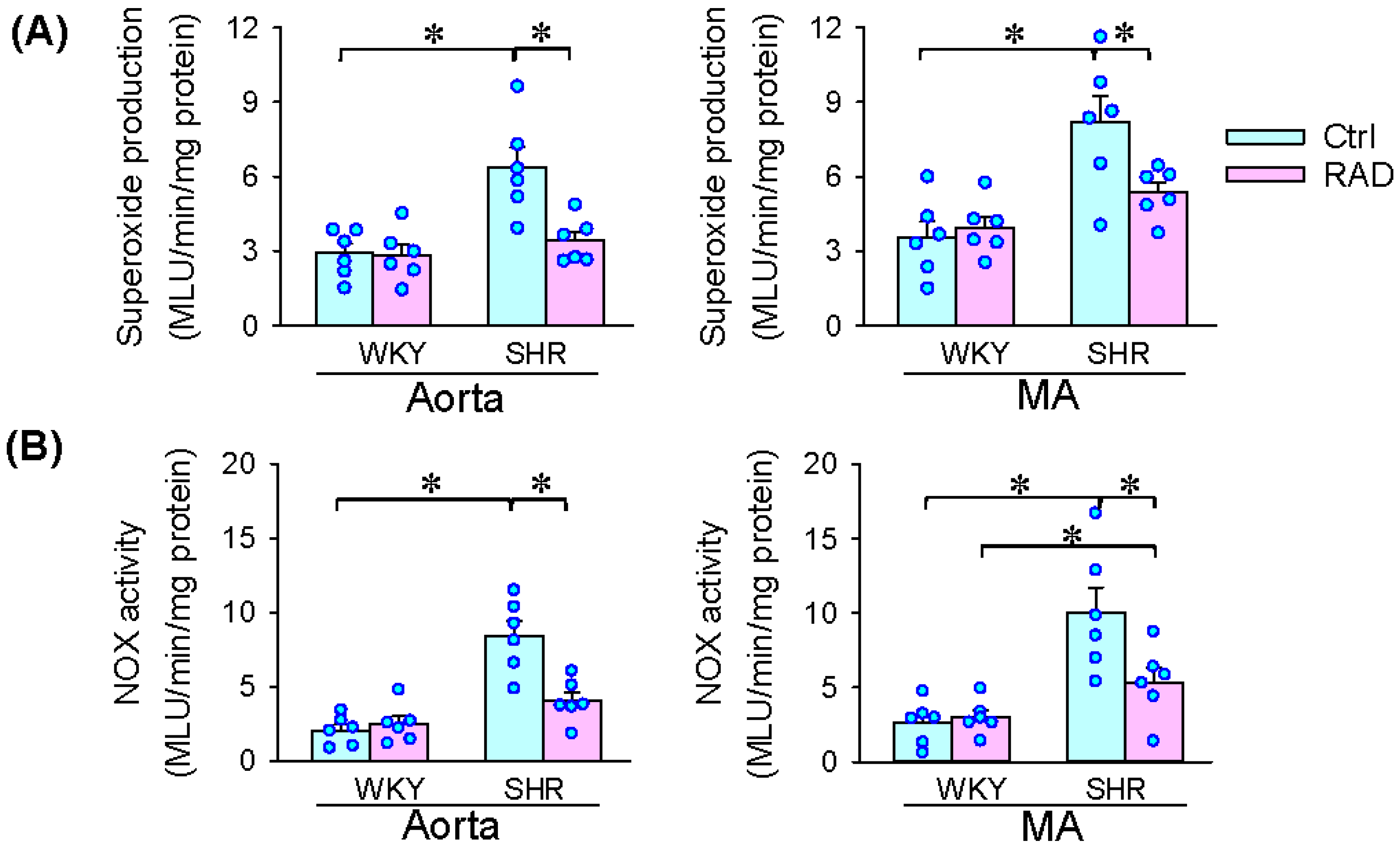
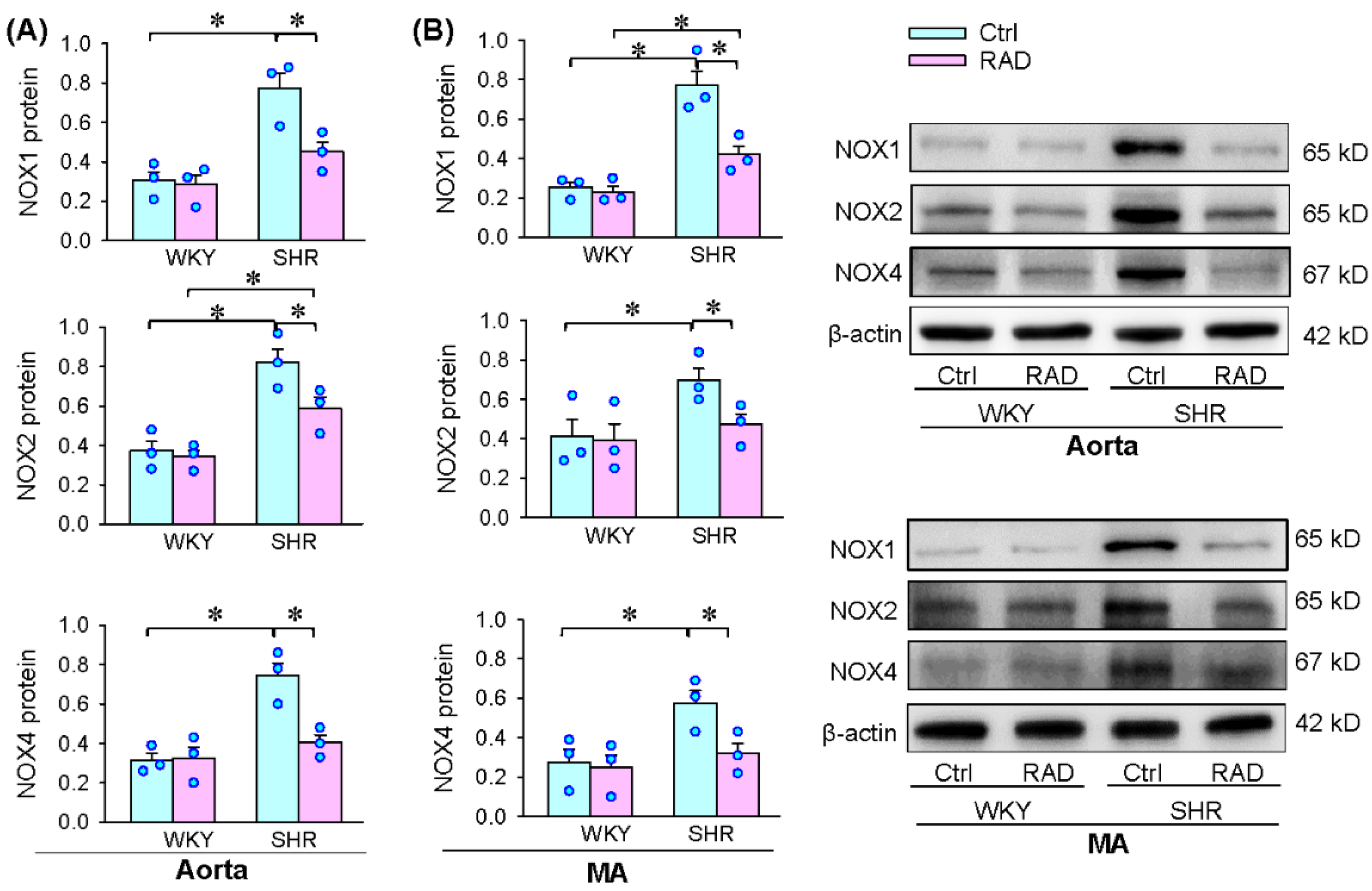
3.6. Oxidative Stress in Arteries
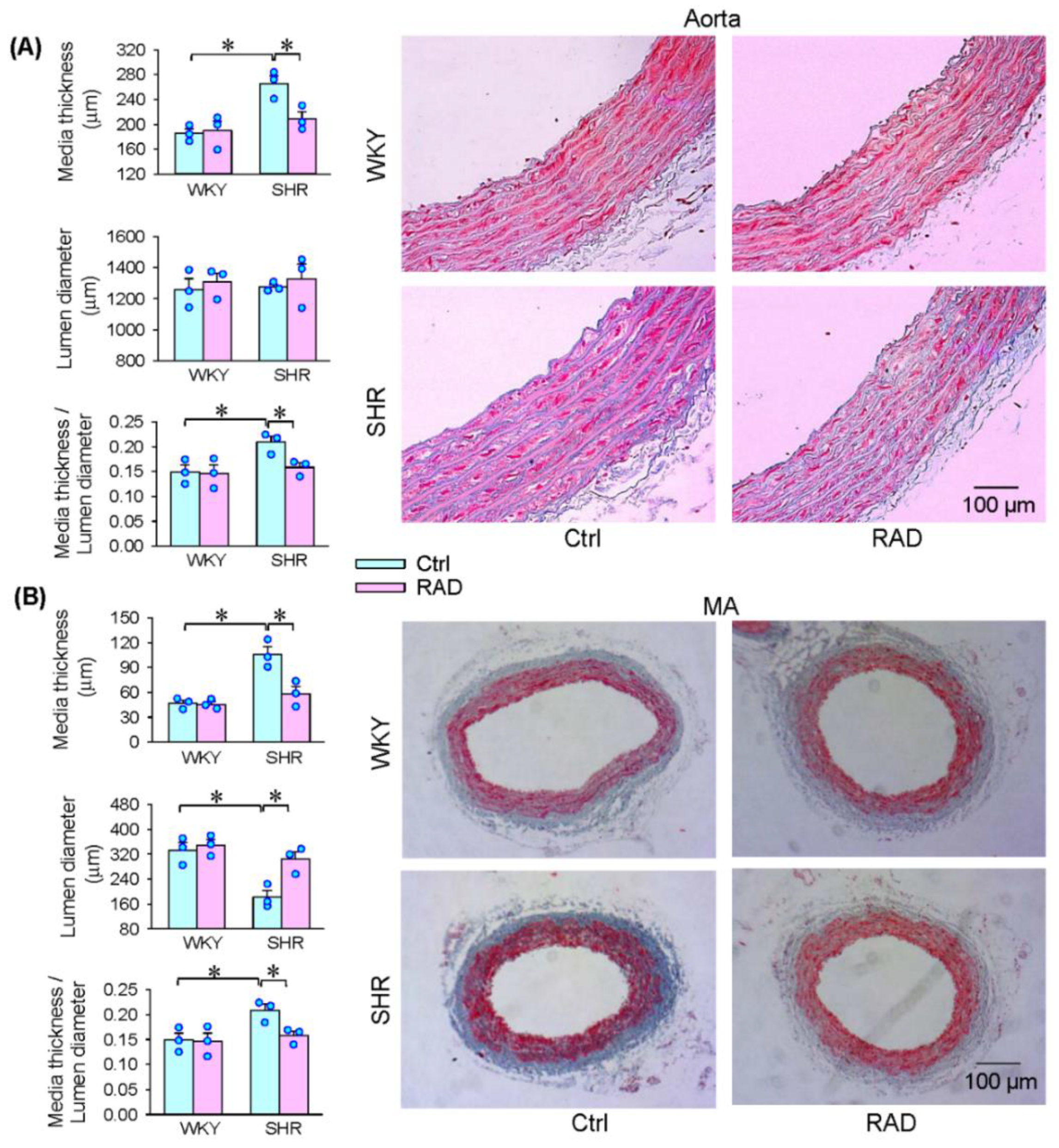
3.7. Vascular Remodeling
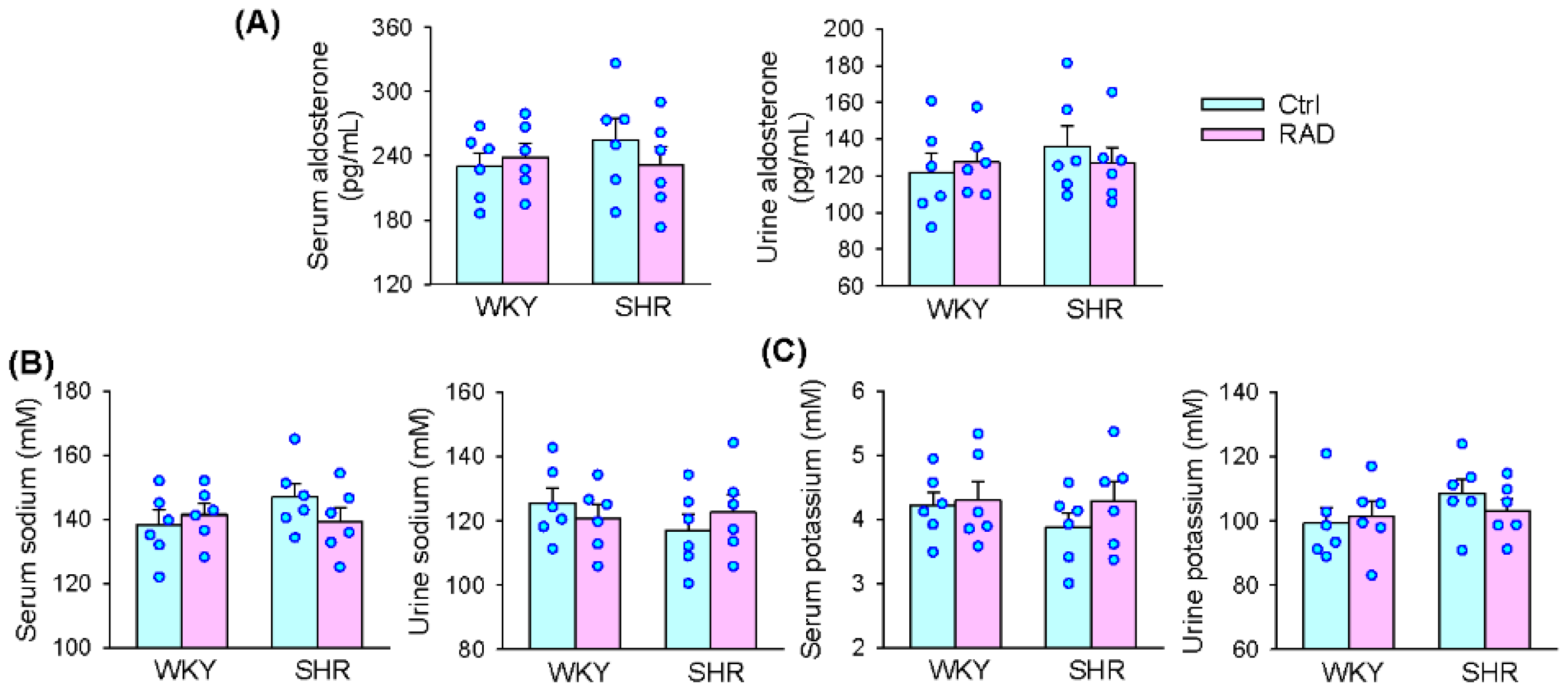
3.8. Serum and Urine Aldosterone, K+ and Na+ Levels
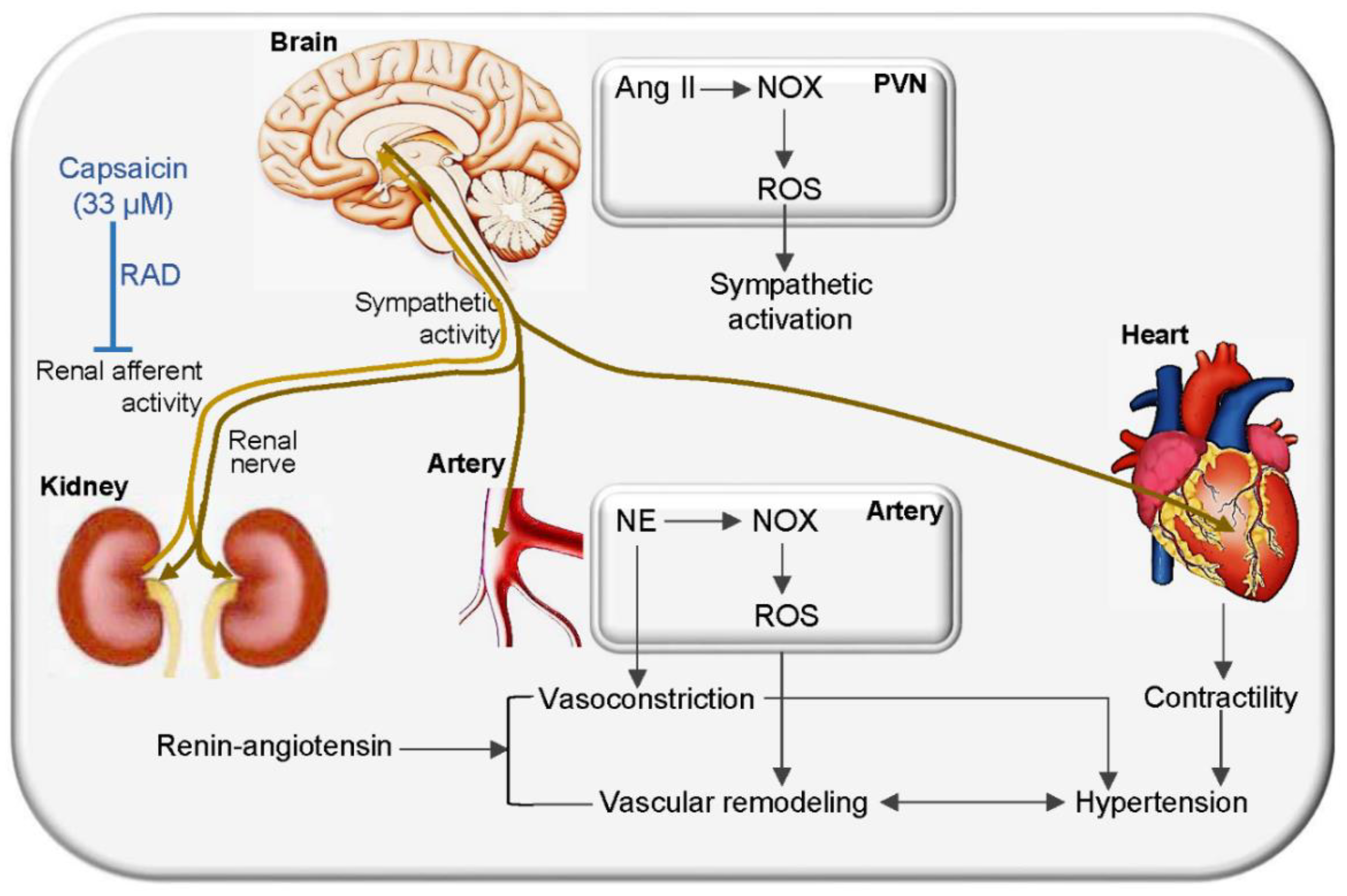
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Garcia-Redondo, A.B.; Aguado, A.; Briones, A.M.; Salaices, M. NADPH oxidases and vascular remodeling in cardiovascular diseases. Pharmacol. Res. 2016, 114, 110–120. [Google Scholar] [CrossRef] [PubMed]
- La, M.S.; Lopez-Sanz, L.; Bernal, S.; Jimenez-Castilla, L.; Prieto, I.; Morelli, G.; Gomez-Guerrero, C.; Marasco, D. Antioxidant effects of PS5, a peptidomimetic of suppressor of cytokine signaling 1, in experimental atherosclerosis. Antioxidants 2020, 9, 754. [Google Scholar]
- Ho, C.C.; Chen, Y.C.; Tsai, M.H.; Tsai, H.T.; Weng, C.Y.; Yet, S.F.; Lin, P. Ambient particulate matter induces vascular smooth muscle cell phenotypic changes via NOX1/ROS/NF-kB dependent and independent pathways: Protective effects of polyphenols. Antioxidants 2021, 10, 782. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Ye, C.; Zheng, F.; Wan, G.W.; Wu, L.L.; Chen, Q.; Li, Y.H.; Kang, Y.M.; Zhu, G.Q. MiR155-5p inhibits cell migration and oxidative stress in vascular smooth muscle cells of spontaneously hypertensive rats. Antioxidants 2020, 9, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obradovic, M.; Essack, M.; Zafirovic, S.; Sudar-Milovanovic, E.; Bajic, V.P.; Van, N.C.; Trpkovic, A.; Stanimirovic, J.; Bajic, V.B.; Isenovic, E.R. Redox control of vascular biology. Biofactors 2020, 46, 246–262. [Google Scholar] [CrossRef]
- Touyz, R.M.; Rios, F.J.; ves-Lopes, R.; Neves, K.B.; Camargo, L.L.; Montezano, A.C. Oxidative Stress: A Unifying Paradigm in Hypertension. Can. J. Cardiol. 2020, 36, 659–670. [Google Scholar] [CrossRef] [Green Version]
- Krzeminska, J.; Wronka, M.; Mfynarska, E.; Franczyk, B.; Rysz, J. Arterial Hypertension-Oxidative Stress and Inflammation. Antioxidants 2022, 11, 172. [Google Scholar] [CrossRef]
- Zhou, B.; Wu, L.L.; Zheng, F.; Wu, N.; Chen, A.D.; Zhou, H.; Chen, J.Y.; Chen, Q.; Li, Y.H.; Kang, Y.M.; et al. miR-31-5p Promotes Oxidative Stress and Vascular Smooth Muscle Cell Migration in Spontaneously Hypertensive Rats via Inhibiting FNDC5 Expression. Biomedicines 2021, 9, 1009. [Google Scholar] [CrossRef]
- Chen, W.W.; Xiong, X.Q.; Chen, Q.; Li, Y.H.; Kang, Y.M.; Zhu, G.Q. Cardiac sympathetic afferent reflex and its implications for sympathetic activation in chronic heart failure and hypertension. Acta Physiol. 2015, 213, 778–794. [Google Scholar] [CrossRef]
- Cheng, Z.J.; Wang, R.; Chen, Q.H. Autonomic rregulation of the cardiovascular system: Diseases, treatments, and novel approaches. Neurosci. Bull. 2019, 35, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.; Zheng, F.; Nan, W.; Zhu, G.Q.; Li, X.Z. Extracellular vesicles in vascular remodeling. Acta Pharmacol. Sin. 2022; Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Grassi, G.; Mark, A.; Esler, M. The sympathetic nervous system alterations in human hypertension. Circ. Res. 2015, 116, 976–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lalio, L.J.; Sved, A.F.; Stocker, S.D. Sympathetic Nervous System Contributions to Hypertension: Updates and Therapeutic Relevance. Can. J. Cardiol. 2020, 36, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Osborn, J.W.; Tyshynsky, R.; Vulchanova, L. Function of Renal Nerves in Kidney Physiology and Pathophysiology. Annu. Rev. Physiol. 2021, 83, 429–450. [Google Scholar] [CrossRef] [PubMed]
- Kaur, J.; Young, B.E.; Fadel, P.J. Sympathetic overactivity in chronic kidney disease: Consequences and mechanisms. Int. J. Mol. Sci. 2017, 18, 1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, C.; Qiu, Y.; Zhang, F.; Chen, A.D.; Zhou, H.; Wang, J.J.; Chen, Q.; Li, Y.H.; Kang, Y.M.; Zhu, G.Q. Chemical stimulation of renal tissue induces sympathetic activation and pressor response via hypothalamic paraventricular nucleus. Neurosci. Bull. 2020, 36, 143–152. [Google Scholar] [CrossRef]
- Qiu, Y.; Zheng, F.; Ye, C.; Chen, A.D.; Wang, J.J.; Chen, Q.; Li, Y.H.; Kang, Y.M.; Zhu, G.Q. Angiotensin type 1 receptors and superoxide anion production in hypothalamic paraventricular nucleus contribute to capsaicin-induced excitatory renal reflex and sympathetic activation. Neurosci. Bull. 2020, 36, 463–474. [Google Scholar] [CrossRef]
- Zheng, F.; Ye, C.; Wan, G.W.; Zhou, B.; Tong, Y.; Lei, J.Z.; Chen, Q.; Li, Y.H.; Kang, Y.M.; Zhu, G.Q. Interleukin-1b in hypothalamic paraventricular nucleus mediates excitatory renal reflex. Pflugers Arch. 2020, 472, 1577–1586. [Google Scholar] [CrossRef]
- Xu, B.; Zheng, H.; Liu, X.; Patel, K.P. Activation of afferent renal nerves modulates RVLM-projecting PVN neurons. Am. J. Physiol. Heart Circ. Physiol. 2015, 308, H1103–H1111. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.; Zheng, F.; Wang, J.X.; Wang, X.L.; Chen, Q.; Li, Y.H.; Kang, Y.M.; Zhu, G.Q. Dysregulation of the excitatory renal reflex in the sympathetic activation of spontaneously hypertensive rat. Front. Physiol. 2021, 12, 673950. [Google Scholar] [CrossRef]
- Kopp, U.C. Role of renal sensory nerves in physiological and pathophysiological conditions. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, R79–R95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milanez, M.I.O.; Veiga, A.C.; Martins, B.S.; Pontes, R.B.; Bergamaschi, C.T.; Campos, R.R.; Nishi, E.E. Renal sensory activity regulates the g-aminobutyric acidergic inputs to the paraventricular nucleus of the hypothalamus in Goldblatt hypertension. Front. Physiol. 2020, 11, 601237. [Google Scholar] [CrossRef] [PubMed]
- Lauder, L.; Bohm, M.; Mahfoud, F. The current status of renal denervation for the treatment of arterial hypertension. Prog. Cardiovasc. Dis. 2021, 65, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.A.; Osborn, J.W. Improved Understanding of Renal Nerve Anatomy: An Opportunity to Enhance Denervation Treatment of Hypertension. JACC Cardiovasc. Interv. 2021, 14, 316–318. [Google Scholar] [CrossRef]
- Lopes, N.R.; Milanez, M.I.O.; Martins, B.S.; Veiga, A.C.; Ferreira, G.R.; Gomes, G.N.; Girardi, A.C.; Carvalho, P.M.; Nogueira, F.N.; Campos, R.R.; et al. Afferent innervation of the ischemic kidney contributes to renal dysfunction in renovascular hypertensive rats. Pflugers Arch. 2020, 472, 325–334. [Google Scholar] [CrossRef]
- Veiga, A.C.; Milanez, M.I.O.; Ferreira, G.R.; Lopes, N.R.; Santos, C.P.; De, A.K.; Garcia, M.L.; Oyama, L.M.; Gomes, G.N.; Nogueira, F.N.; et al. Selective afferent renal denervation mitigates renal and splanchnic sympathetic nerve overactivity and renal function in chronic kidney disease-induced hypertension. J. Hypertens 2020, 38, 765–773. [Google Scholar] [CrossRef]
- Graham, D.; McBride, M.W.; Brain, N.J.; Dominiczak, A.F. Congenic/consomic models of hypertension. Methods Mol Med 2005, 108, 3–15. [Google Scholar]
- Bell, D.; Kelso, E.J.; Argent, C.C.; Lee, G.R.; Allen, A.R.; McDermott, B.J. Temporal characteristics of cardiomyocyte hypertrophy in the spontaneously hypertensive rat. Cardiovasc. Pathol. 2004, 13, 71–78. [Google Scholar] [CrossRef]
- Foss, J.D.; Wainford, R.D.; Engeland, W.C.; Fink, G.D.; Osborn, J.W. A novel method of selective ablation of afferent renal nerves by periaxonal application of capsaicin. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, R112–R122. [Google Scholar] [CrossRef] [Green Version]
- Kondo, H.; Kondo, M.; Hayashi, K.; Kusafuka, S.; Hamamura, K.; Tanaka, K.; Kodama, D.; Hirai, T.; Sato, T.; Ariji, Y.; et al. Orthodontic tooth movement-activated sensory neurons contribute to enhancing osteoclast activity and tooth movement through sympathetic nervous signalling. Eur. J. Orthod. 2021; Online ahead of print. [Google Scholar] [CrossRef]
- Giancola, F.; Gentilini, F.; Romagnoli, N.; Spadari, A.; Turba, M.E.; Giunta, M.; Sadeghinezhad, J.; Sorteni, C.; Chiocchetti, R. Extrinsic innervation of ileum and pelvic flexure of foals with ileocolonic aganglionosis. Cell Tissue Res. 2016, 366, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.Q.; Chen, W.W.; Zhu, G.Q. Adipose afferent reflex: Sympathetic activation and obesity hypertension. Acta Physiol. 2014, 210, 468–478. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Chen, W.W.; Xiong, X.Q.; Han, Y.; Zhou, Y.B.; Zhang, F.; Gao, X.Y.; Zhu, G.Q. Sympathetic activation by chemical stimulation of white adipose tissues in rats. J. Appl. Physiol. 2012, 112, 1008–1014. [Google Scholar] [CrossRef] [Green Version]
- Xiong, X.Q.; Chen, W.W.; Han, Y.; Zhou, Y.B.; Zhang, F.; Gao, X.Y.; Zhu, G.Q. Enhanced adipose afferent reflex contributes to sympathetic activation in diet-induced obesity hypertension. Hypertension 2012, 60, 1280–1286. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.L.; Bo, J.H.; Zheng, F.; Zhang, F.; Chen, Q.; Li, Y.H.; Kang, Y.M.; Zhu, G.Q. Salusin-b in intermediate dorsal motor nucleus of the vagus regulates sympathetic-parasympathetic balance and blood pressure. Biomedicines 2021, 9, 1118. [Google Scholar] [CrossRef] [PubMed]
- Task Force of the European Society of Cardiology the North American Society of Pacing Electrophysiology. Heart rate variability: Standards of measurement, physiological interpretation and clinical use. Task Force of the European Society of Cardiology and the North American Society of Pacing and Electrophysiology. Circulation 1996, 93, 1043–1065. [Google Scholar] [CrossRef] [Green Version]
- Garde, A.H.; Laursen, B.; Jorgensen, A.H.; Jensen, B.R. Effects of mental and physical demands on heart rate variability during computer work. Eur. J. Appl. Physiol. 2002, 87, 456–461. [Google Scholar] [CrossRef]
- Wu, N.; Zheng, F.; Li, N.; Han, Y.; Xiong, X.Q.; Wang, J.J.; Chen, Q.; Li, Y.H.; Zhu, G.Q.; Zhou, Y.B. RND3 attenuates oxidative stress and vascular remodeling in spontaneously hypertensive rat via inhibiting ROCK1 signaling. Redox Biol. 2021, 48, 102204. [Google Scholar] [CrossRef]
- Sun, H.J.; Ren, X.S.; Xiong, X.Q.; Chen, Y.Z.; Zhao, M.X.; Wang, J.J.; Zhou, Y.B.; Han, Y.; Chen, Q.; Li, Y.H.; et al. NLRP3 inflammasome activation contributes to VSMC phenotypic transformation and proliferation in hypertension. Cell Death Dis. 2017, 8, e3074. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.S.; Tong, Y.; Qiu, Y.; Ye, C.; Wu, N.; Xiong, X.Q.; Wang, J.J.; Han, Y.; Zhou, Y.B.; Zhang, F.; et al. MiR155-5p in adventitial fibroblasts-derived extracellular vesicles inhibits vascular smooth muscle cell proliferation via suppressing angiotensin-converting enzyme expression. J. Extracell Vesicles 2020, 9, 1698795. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.Y.; Shi, C.X.; Gao, R.; Sun, H.J.; Xiong, X.Q.; Ding, L.; Chen, Q.; Li, Y.H.; Wang, J.J.; Kang, Y.M.; et al. Irisin inhibits hepatic gluconeogenesis and increases glycogen synthesis via the PI3K/Akt pathway in type 2 diabetic mice and hepatocytes. Clin. Sci. 2015, 129, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, M. Capsaicin and sensory neurones—A review. Pain 1983, 15, 109–130. [Google Scholar] [CrossRef]
- Wang, D.H.; Li, J.; Qiu, J. Salt-sensitive hypertension induced by sensory denervation: Introduction of a new model. Hypertension 1998, 32, 649–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.H.; Wu, W.; Lookingland, K.J. Degeneration of capsaicin-sensitive sensory nerves leads to increased salt sensitivity through enhancement of sympathoexcitatory response. Hypertension 2001, 37 Pt 2, 440–443. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, A.F.; Wang, D.H. ET(A) receptor blockade prevents renal dysfunction in salt-sensitive hypertension induced by sensory denervation. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H2005–H2011. [Google Scholar] [CrossRef] [Green Version]
- Kopp, U.C.; Cicha, M.Z.; Smith, L.A. Dietary sodium loading increases arterial pressure in afferent renal-denervated rats. Hypertension 2003, 42, 968–973. [Google Scholar] [CrossRef] [Green Version]
- Kopp, U.C.; Jones, S.Y.; DiBona, G.F. Afferent renal denervation impairs baroreflex control of efferent renal sympathetic nerve activity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R1882–R1890. [Google Scholar] [CrossRef] [Green Version]
- Holzer, P. Capsaicin: Cellular targets, mechanisms of action, and selectivity for thin sensory neurons. Pharmacol Rev 1991, 43, 143–201. [Google Scholar]
- Ulrich-Lai, Y.M.; Fraticelli, A.I.; Engeland, W.C. Capsaicin-sensitive nerve fibers: A potential extra-ACTH mechanism participating in adrenal regeneration in rats. Microsc Res Tech 2003, 61, 252–258. [Google Scholar] [CrossRef]
- Wang, H.; Wang, D.H.; Galligan, J.J. P2Y2 receptors mediate ATP-induced resensitization of TRPV1 expressed by kidney projecting sensory neurons. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R1634–R1641. [Google Scholar] [CrossRef] [Green Version]
- Knuepfer, M.M.; Schramm, L.P. The conduction velocities and spinal projections of single renal afferent fibers in the rat. Brain Res. 1987, 435, 167–173. [Google Scholar] [CrossRef]
- Szallasi, A.; Blumberg, P.M. Vanilloid (Capsaicin) receptors and mechanisms. Pharmacol. Rev. 1999, 51, 159–212. [Google Scholar]
- Sun, H.J.; Zhang, L.L.; Fan, Z.D.; Chen, D.; Zhang, L.; Gao, X.Y.; Kang, Y.M.; Zhu, G.Q. Superoxide anions involved in sympathoexcitation and pressor effects of salusin-beta in paraventricular nucleus in hypertensive rats. Acta Physiol. 2014, 210, 534–545. [Google Scholar] [CrossRef] [PubMed]
- Yuan, N.; Zhang, F.; Zhang, L.L.; Gao, J.; Zhou, Y.B.; Han, Y.; Zhu, G.Q. SOD1 gene transfer into paraventricular nucleus attenuates hypertension and sympathetic activity in spontaneously hypertensive rats. Pflugers Arch. 2013, 465, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Schraml, E.; Quan, P.; Stelzer, I.; Fuchs, R.; Skalicky, M.; Viidik, A.; Schauenstein, K. Norepinephrine treatment and aging lead to systemic and intracellular oxidative stress in rats. Exp. Gerontol. 2007, 42, 1072–1078. [Google Scholar] [CrossRef] [PubMed]
- Deo, S.H.; Jenkins, N.T.; Padilla, J.; Parrish, A.R.; Fadel, P.J. Norepinephrine increases NADPH oxidase-derived superoxide in human peripheral blood mononuclear cells via a-adrenergic receptors. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R1124–R1132. [Google Scholar] [CrossRef]
- Hering, L.; Rahman, M.; Potthoff, S.A.; Rump, L.C.; Stegbauer, J. Role of a-adrenoceptors in hypertension: Focus on renal sympathetic neurotransmitter release, inflammation, and sodium homeostasis. Front. Physiol. 2020, 11, 566871. [Google Scholar] [CrossRef]
- Kato, T.; Mizuguchi, N.; Ito, A. Blood pressure, renal biochemical parameters and histopathology in an original rat model of essential hypertension (SHRSP/Kpo strain). Biomed. Res. 2015, 36, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Qiu, L.; Zhang, J.; Yang, Y.; Zhang, H.; Lee, F.F.; He, Q.; Huang, C.; Huang, L.; Qian, L.; Luo, J. In vivo assessment of hypertensive nephrosclerosis using ultrasound localization microscopy. Med. Phys. 2022, 49, 2295–2308. [Google Scholar] [CrossRef]
- Naruse, M.; Tanabe, A.; Naruse, K.; Adachi, C.; Yoshimoto, T.; Seki, T.; Takagi, S.; Imaki, T.; Watanabe, T.; Takano, K. Hemodynamic and biochemical effects of endothelin-A- and -B-receptor antagonist TAK-044 in stroke-prone spontaneously hypertensive rats. J. Cardiovasc. Pharmacol. 2000, 36 (Suppl. S1), S334–S336. [Google Scholar] [CrossRef]
- Wang, X.; Zhu, Y.; Wang, S.; Wang, Z.; Sun, H.; He, Y.; Yao, W. Effects of eplerenone on cerebral aldosterone levels and brain lesions in spontaneously hypertensive rats. Clin. Exp. Hypertens 2020, 42, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Qin, F.; Li, J.; Dai, Y.F.; Zhong, X.G.; Pan, Y.J. Renal denervation inhibits the renin-angiotensin-aldosterone system in spontaneously hypertensive rats. Clin. Exp. Hypertens 2022, 44, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Naruse, M.; Tanabe, A.; Sato, A.; Takagi, S.; Tsuchiya, K.; Imaki, T.; Takano, K. Aldosterone breakthrough during angiotensin II receptor antagonist therapy in stroke-prone spontaneously hypertensive rats. Hypertension 2002, 40, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Derderian, D.T.; Vendramini, R.C.; Menani, J.V.; De Luca, L.A., Jr. Water deprivation-induced sodium appetite and differential expression of encephalic c-Fos immunoreactivity in the spontaneously hypertensive rat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R1298–R1309. [Google Scholar] [CrossRef] [PubMed]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, L.-L.; Zhang, Y.; Li, X.-Z.; Du, X.-L.; Gao, Y.; Wang, J.-X.; Wang, X.-L.; Chen, Q.; Li, Y.-H.; Zhu, G.-Q.; et al. Impact of Selective Renal Afferent Denervation on Oxidative Stress and Vascular Remodeling in Spontaneously Hypertensive Rats. Antioxidants 2022, 11, 1003. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11051003
Wu L-L, Zhang Y, Li X-Z, Du X-L, Gao Y, Wang J-X, Wang X-L, Chen Q, Li Y-H, Zhu G-Q, et al. Impact of Selective Renal Afferent Denervation on Oxidative Stress and Vascular Remodeling in Spontaneously Hypertensive Rats. Antioxidants. 2022; 11(5):1003. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11051003
Chicago/Turabian StyleWu, Lu-Lu, Yue Zhang, Xiu-Zhen Li, Xin-Li Du, Ying Gao, Jing-Xiao Wang, Xiao-Li Wang, Qi Chen, Yue-Hua Li, Guo-Qing Zhu, and et al. 2022. "Impact of Selective Renal Afferent Denervation on Oxidative Stress and Vascular Remodeling in Spontaneously Hypertensive Rats" Antioxidants 11, no. 5: 1003. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11051003