
Evaluation of Antioxidant Capacity and Gut Microbiota Modulatory Effects of Different Kinds of Berries

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fruit Materials
2.2. Extraction and Identification of Berry Anthocyanins
2.3. In Vitro Chemical Antioxidant Capacity Evaluation of Berry Extract
2.4. Animals and Treatment
2.5. 16 S rDNA Amplicon Sequencing and Analysis
2.6. In Vivo Antioxidant Capacity Evaluation
2.7. Statistical Analysis
3. Results
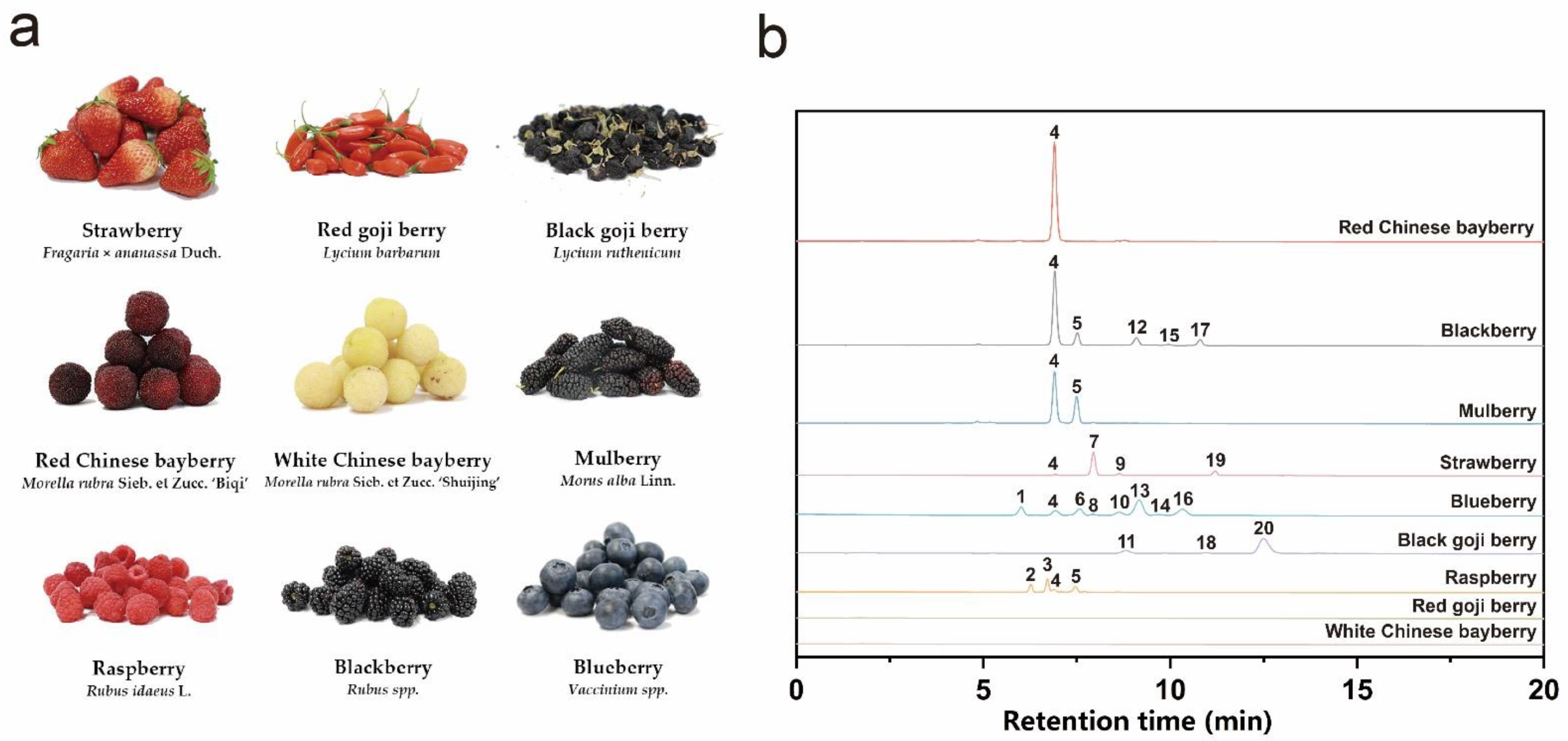
3.1. Anthocyanin Profiles of Different Kinds of Berries
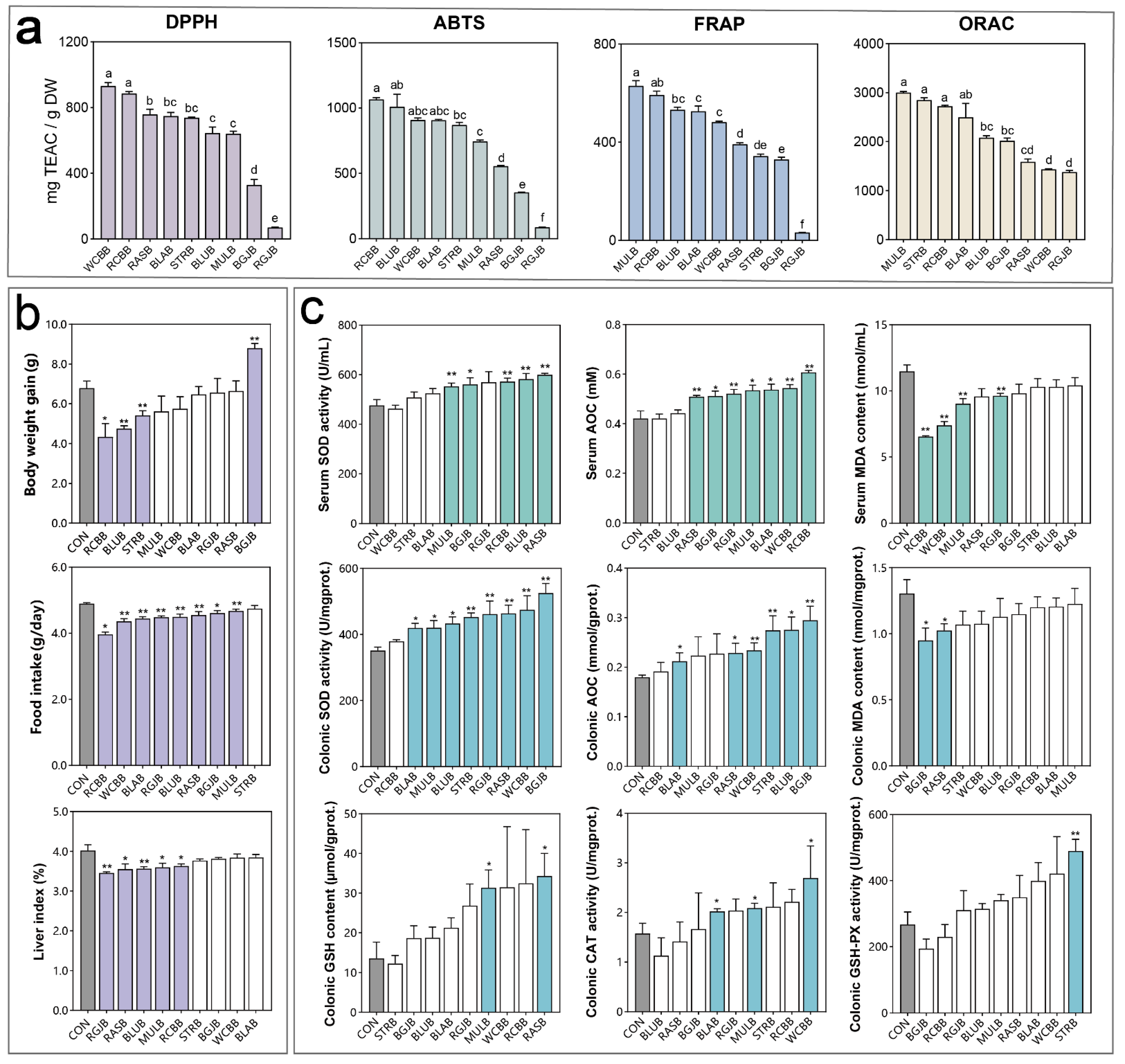
3.2. Antioxidant Capacity Evaluation of Different Kinds of Berries
3.3. Influence of Berry Extracts on the Antioxidant Capacities of Healthy Objects
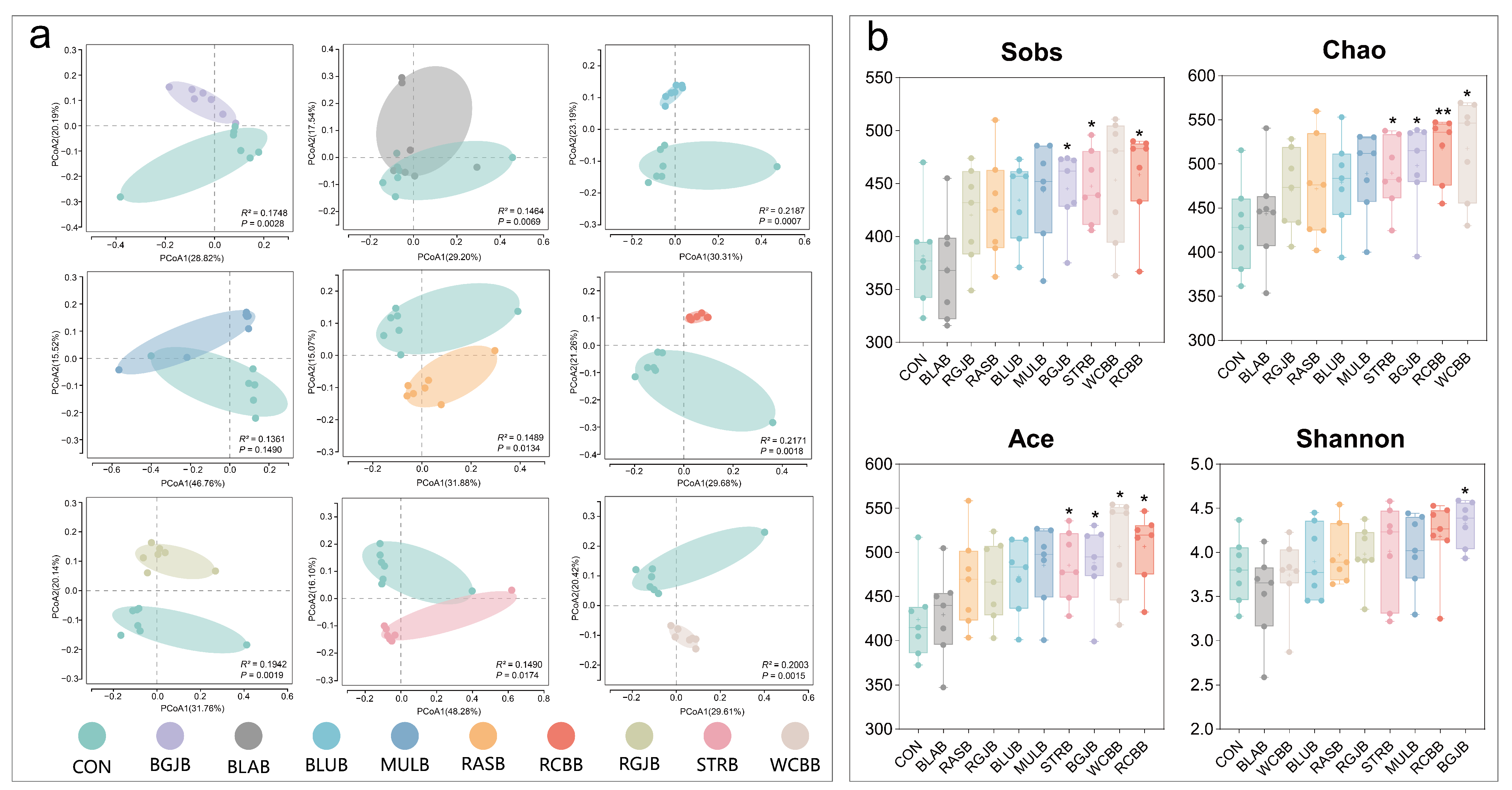
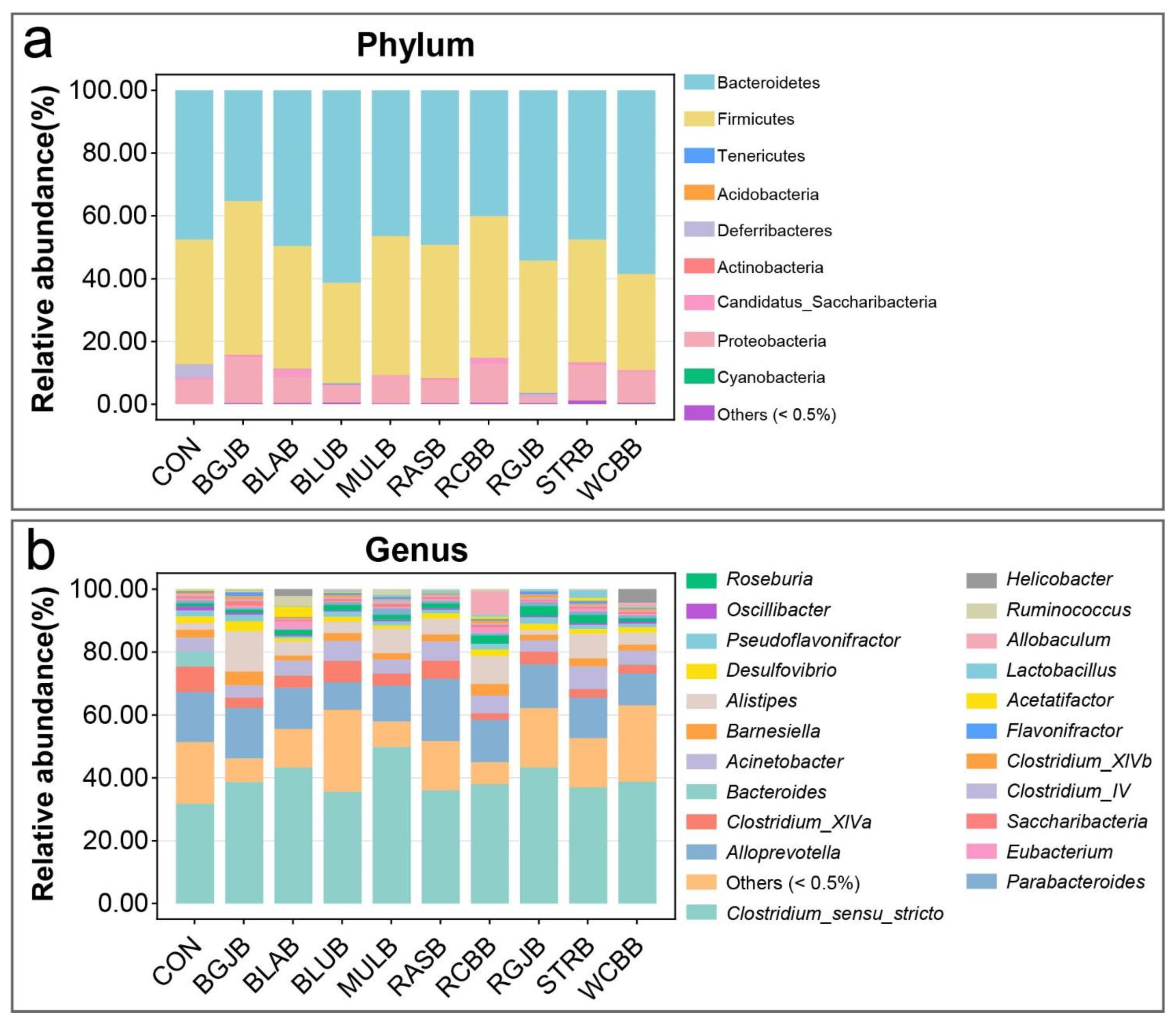
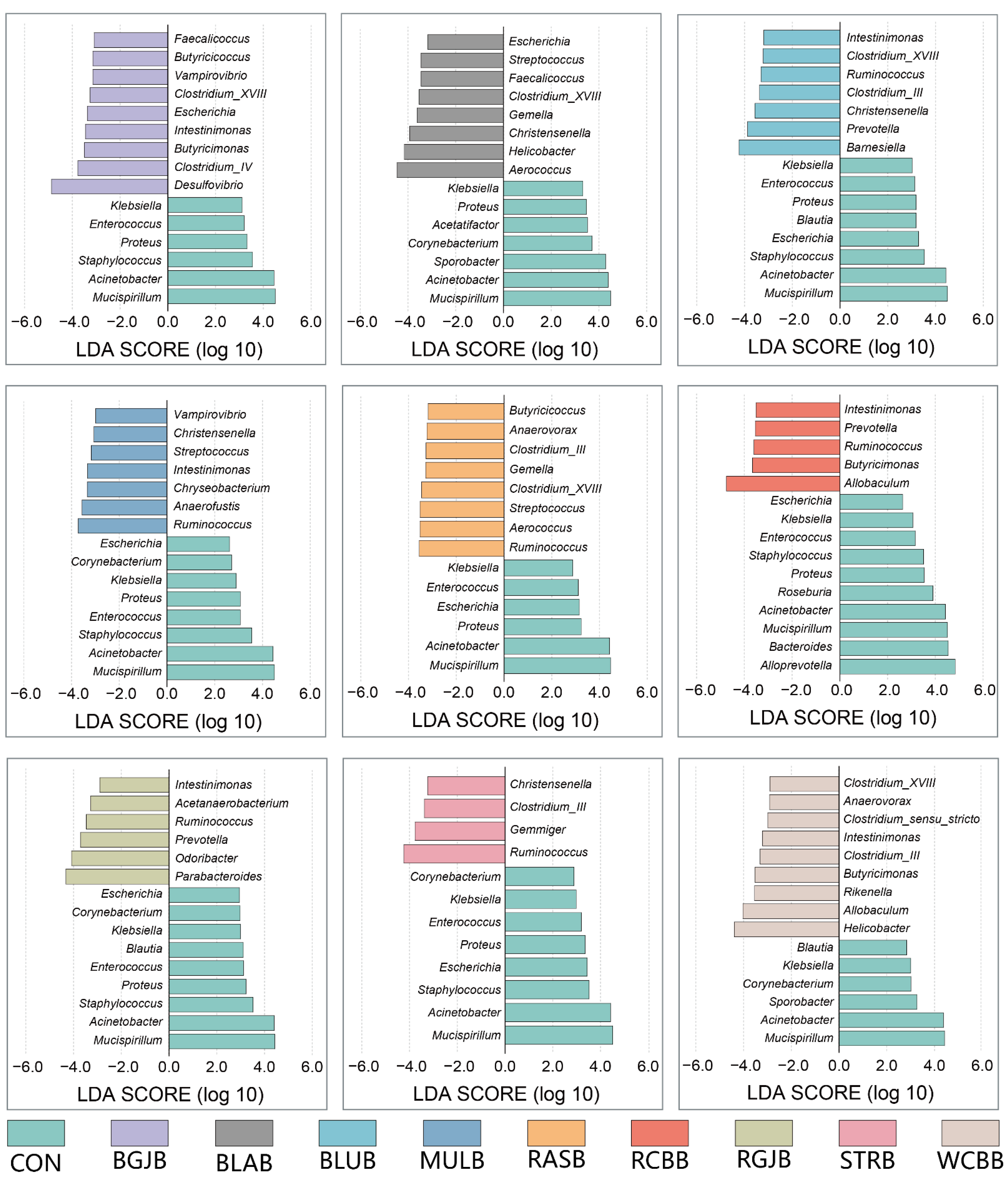
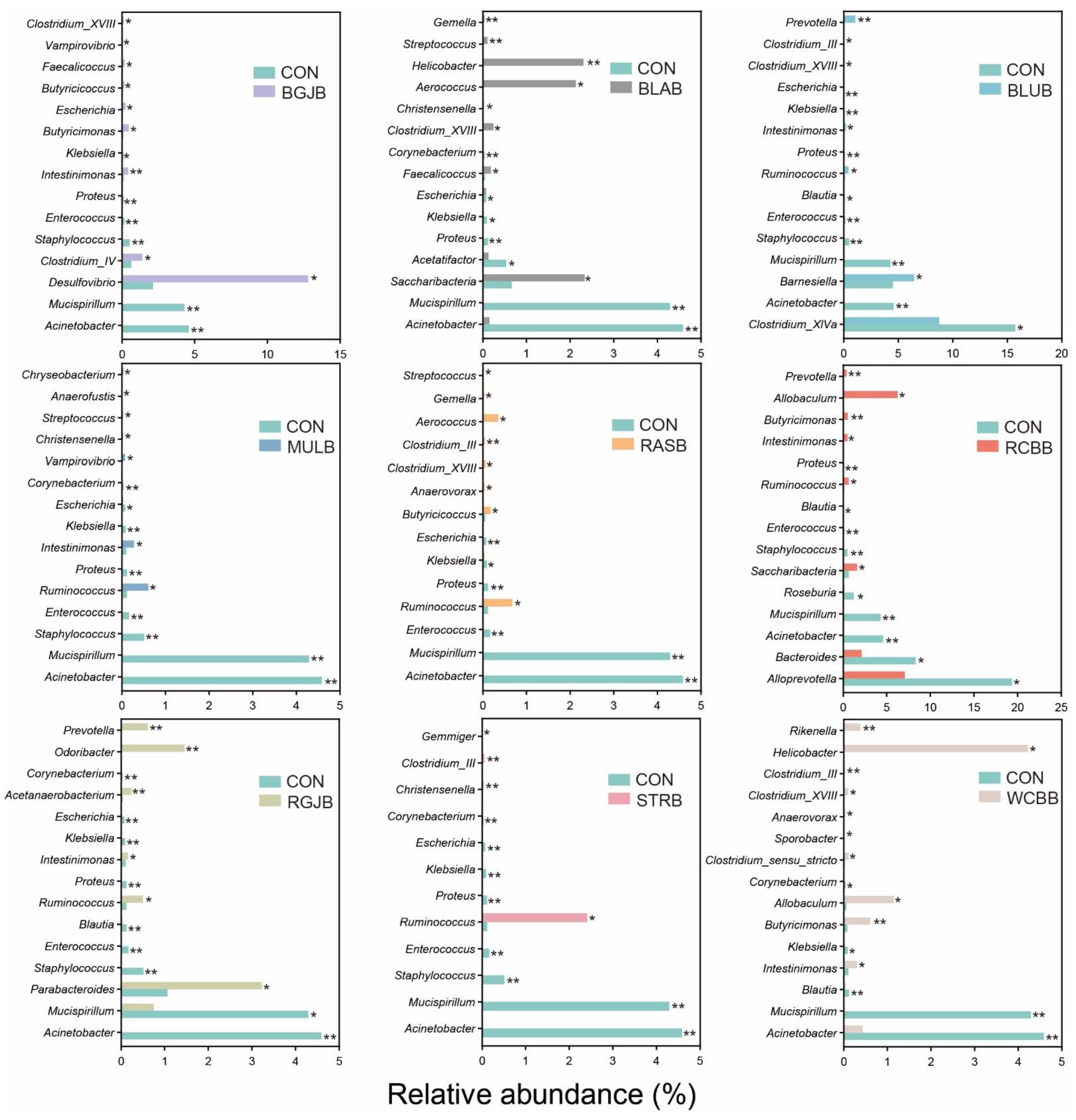
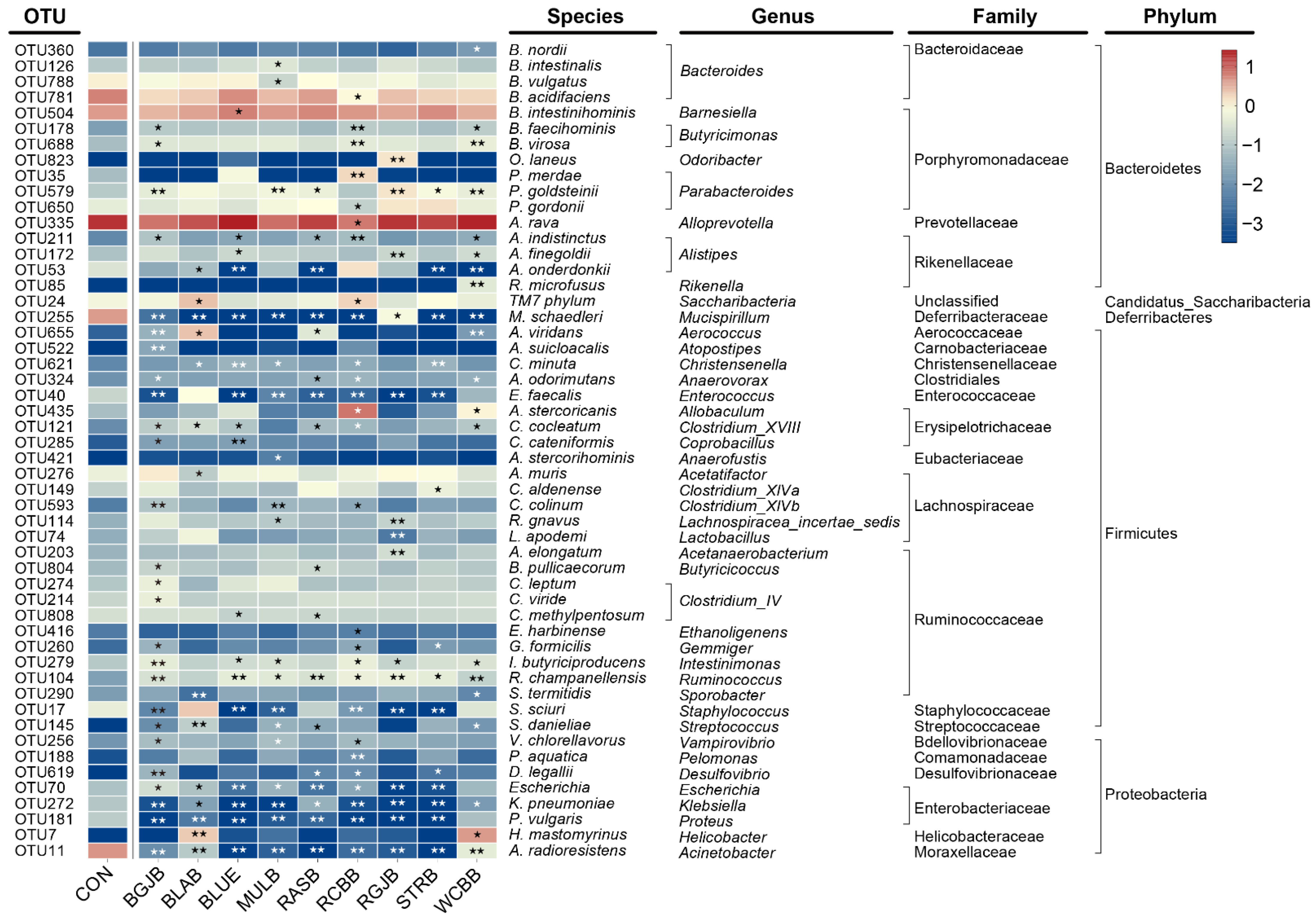
3.4. Influence of Berries Extracts on the Gut Microbiota of Healthy Objects
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yi, L.; Ma, S.; Ren, D. Phytochemistry and bioactivity of Citrus flavonoids: A focus on antioxidant, anti-inflammatory, anticancer and cardiovascular protection activities. Phytochem. Rev. 2017, 16, 479–511. [Google Scholar] [CrossRef]
- Fraga, C.G.; Croft, K.D.; Kennedy, D.O.; Tomas-Barberan, F.A. The effects of polyphenols and other bioactives on human health. Food Funct. 2019, 10, 514–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavefve, L.; Howard, L.R.; Carbonero, F. Berry polyphenols metabolism and impact on human gut microbiota and health. Food Funct. 2020, 11, 45–65. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Lopez, O.; Cervantes-Ceja, M.L.; Vigna-Perez, M.; Hernandez-Perez, T. Berries: Improving human health and healthy aging, and promoting quality life—A review. Plant Foods Hum. Nutr. 2010, 65, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Nile, S.H.; Park, S.W. Edible berries: Bioactive components and their effect on human health. Nutrition 2014, 30, 134–144. [Google Scholar] [CrossRef]
- Tian, L.; Tan, Y.; Chen, G.; Wang, G.; Sun, J.; Ou, S.; Chen, W.; Bai, W. Metabolism of anthocyanins and consequent effects on the gut microbiota. Crit. Rev. Food Sci. Nutr. 2019, 59, 982–991. [Google Scholar] [CrossRef]
- de Mejia, E.G.; Zhang, Q.; Penta, K.; Eroglu, A.; Lila, M.A. The Colors of Health: Chemistry, Bioactivity, and Market Demand for Colorful Foods and Natural Food Sources of Colorants. Annu. Rev. Food Sci. Technol. 2020, 11, 145–182. [Google Scholar] [CrossRef] [Green Version]
- Pap, N.; Fidelis, M.; Azevedo, L.; Vieira do Carmo, M.A.; Wang, D.; Mocan, A.; Rodrigues Pereira, E.P.; Xavier-Santos, D.; Sant’Ana, A.S.; Yang, B.; et al. Berry polyphenols and human health: Evidence of antioxidant, anti-inflammatory, microbiota modulation, and cell-protecting effects. Curr. Opin. Food Sci. 2021, 42, 167–186. [Google Scholar] [CrossRef]
- Skrovankova, S.; Sumczynski, D.; Mlcek, J.; Jurikova, T.; Sochor, J. Bioactive Compounds and Antioxidant Activity in Different Types of Berries. Int. J. Mol. Sci. 2015, 16, 24673–24706. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Xiao, H. Whole Food-Based Approaches to Modulating Gut Microbiota and Associated Diseases. Annu. Rev. Food Sci. Technol. 2020, 11, 119–143. [Google Scholar] [CrossRef] [Green Version]
- Martel, J.; Ojcius, D.M.; Chang, C.-J.; Lin, C.-S.; Lu, C.-C.; Ko, Y.-F.; Tseng, S.-F.; Lai, H.-C.; Young, J.D. Anti-obesogenic and antidiabetic effects of plants and mushrooms. Nat. Rev. Endocrinol. 2017, 13, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulange, C.L.; Neves, A.L.; Chilloux, J.; Nicholson, J.K.; Dumas, M.-E. Impact of the gut microbiota on inflammation, obesity, and metabolic disease. Genome Med. 2016, 8, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonnenburg, J.L.; Backhed, F. Diet-microbiota interactions as moderators of human metabolism. Nature 2016, 535, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Loo, Y.T.; Howell, K.; Chan, M.; Zhang, P.; Ng, K. Modulation of the human gut microbiota by phenolics and phenolic fiber-rich foods. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1268–1298. [Google Scholar] [CrossRef]
- Govers, C.; Kasikci, M.B.; van der Sluis, A.A.; Mes, J.J. Review of the health effects of berries and their phytochemicals on the digestive and immune systems. Nutr. Rev. 2018, 76, 29–46. [Google Scholar] [CrossRef]
- Leng, F.; Cao, J.P.; Ge, Z.W.; Wang, Y.; Zhao, C.N.; Wang, S.P.; Li, X.; Zhang, Y.L.; Sun, C.D. Transcriptomic Analysis of Root Restriction Effects on Phenolic Metabolites during Grape Berry Development and Ripening. J. Agric. Food Chem. 2020, 68, 9090–9099. [Google Scholar] [CrossRef]
- Wang, Y.; Qian, J.; Cao, J.; Wang, D.; Liu, C.; Yang, R.; Li, X.; Sun, C. Antioxidant Capacity, Anticancer Ability and Flavonoids Composition of 35 Citrus (Citrus reticulata Blanco) Varieties. Molecules 2017, 22, 1114. [Google Scholar] [CrossRef]
- Zhou, L.; Xie, M.; Yang, F.; Liu, J. Antioxidant activity of high purity blueberry anthocyanins and the effects on human intestinal microbiota. LWT-Food Sci. Technol. 2020, 117, 108621. [Google Scholar] [CrossRef]
- Reynoso-Camacho, R.; Sotelo-González, A.M.; Patiño-Ortiz, P.; Rocha-Guzmán, N.E.; Pérez-Ramírez, I.F. Berry by-products obtained from a decoction process are a rich source of low- and high-molecular weight extractable and non-extractable polyphenols. Food Bioprod. Process. 2021, 127, 371–387. [Google Scholar] [CrossRef]
- Renai, L.; Scordo, C.V.A.; Chiuminatto, U.; Ulaszewska, M.; Giordani, E.; Petrucci, W.A.; Tozzi, F.; Nin, S.; Del Bubba, M. Liquid Chromatographic Quadrupole Time-of-Flight Mass Spectrometric Untargeted Profiling of (Poly)phenolic Compounds in Rubus idaeus L. and Rubus occidentalis L. Fruits and Their Comparative Evaluation. Antioxidants 2021, 10, 704. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Huang, H.; Zhang, Q.; Fan, F.; Xu, C.; Sun, C.; Li, X.; Chen, K. Phytochemical Characterization of Chinese Bayberry (Myrica rubra Sieb. et Zucc.) of 17 Cultivars and Their Antioxidant Properties. Int. J. Mol. Sci. 2015, 16, 12467–12481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolniak-Ostek, J.; Kucharska, A.Z.; Sokol-Letowska, A.; Fecka, I. Characterization of Phenolic Compounds of Thorny and Thornless Blackberries. J. Agric. Food Chem. 2015, 63, 3012–3021. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Meng, X.; Li, B. Profiling of anthocyanins from blueberries produced in China using HPLC-DAD-MS and exploratory analysis by principal component analysis. J. Food Compost. Anal. 2016, 47, 1–7. [Google Scholar] [CrossRef]
- Nzeuwa, I.B.Y.; Xia, Y.; Qiao, Z.; Feng, F.; Bian, J.; Liu, W.; Qu, W. Comparison of the origin and phenolic contents of Lycium ruthenicum Murr. by high-performance liquid chromatography fingerprinting combined with quadrupole time-of-flight mass spectrometry and chemometrics. J. Sep. Sci. 2017, 40, 1234–1243. [Google Scholar] [CrossRef] [PubMed]
- Tao, W.; Haiyang, L.; Fengzhong, W.; Yi, W. Characterization of polyphenols from Lycium ruthenicum fruit by UPLC-Q-TOF/MSE and their antioxidant activity in Caco-2 cells. J. Agric. Food Chem. 2016, 64, 2280–2288. [Google Scholar] [CrossRef]
- Alissa, E.M.; Ferns, G.A. Dietary fruits and vegetables and cardiovascular diseases risk. Crit. Rev. Food Sci. Nutr. 2017, 57, 1950–1962. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Casado, A. The Health Potential of Fruits and Vegetables Phytochemicals: Notable Examples. Crit. Rev. Food Sci. Nutr. 2016, 56, 1097–1107. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, X.; Zhan, L.; Xu, C.; Sun, L.; Jiang, H.; Sun, C.; Li, X. LC-Q-TOF-MS Characterization of Polyphenols from White Bayberry Fruit and Its Antidiabetic Effect in KK-A(y) Mice. ACS Omega 2020, 5, 17839–17849. [Google Scholar] [CrossRef]
- Sun, Q.; Du, M.; Kang, Y.; Zhu, M.-J. Prebiotic effects of goji berry in protection against inflammatory bowel disease. Crit. Rev. Food Sci. Nutr. 2021, 1–25. [Google Scholar] [CrossRef]
- Huang, D.J.; Ou, B.X.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Diaz de Barboza, G.; Guizzardi, S.; Moine, L.; Tolosa de Talamoni, N. Oxidative stress, antioxidants and intestinal calcium absorption. World J. Gastroenterol. 2017, 23, 2841–2853. [Google Scholar] [CrossRef] [PubMed]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and prebiotics in intestinal health and disease: From biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Catalkaya, G.; Venema, K.; Lucini, L.; Rocchetti, G.; Delmas, D.; Daglia, M.; De Filippis, A.; Xiao, H.; Quiles, J.L.; Xiao, J.; et al. Interaction of dietary polyphenols and gut microbiota: Microbial metabolism of polyphenols, influence on the gut microbiota, and implications on host health. Food Front. 2020, 1, 109–133. [Google Scholar] [CrossRef]
- Huttenhower, C.; Gevers, D.; Knight, R.; Abubucker, S.; Badger, J.H.; Chinwalla, A.T.; Creasy, H.H.; Earl, A.M.; FitzGerald, M.G.; Fulton, R.S.; et al. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Wong, D.; Nielsen, T.B.; Bonomo, R.A.; Pantapalangkoor, P.; Luna, B.; Spellberg, B. Clinical and Pathophysiological Overview of Acinetobacter Infections: A Century of Challenges. Clin. Microbiol. Rev. 2017, 30, 409–447. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wang, X.H.; Chen, Q.B.; Luo, L.Y.; Ma, M.J.; Xiao, B.; Zeng, L. Camellia sinensis and Litsea coreana Ameliorate Intestinal Inflammation and Modulate Gut Microbiota in Dextran Sulfate Sodium-Induced Colitis Mice. Mol. Nutr. Food Res. 2020, 64, 11. [Google Scholar] [CrossRef]
- Herp, S.; Raj, A.C.D.; Silva, M.S.; Woelfel, S.; Stecher, B. The human symbiont Mucispirillum schaedleri: Causality in health and disease. Med. Microbiol. Immunol. 2021, 210, 173–179. [Google Scholar] [CrossRef]
- Zhao, H.A.; Cheng, N.; Zhou, W.Q.; Chen, S.N.; Wang, Q.; Gao, H.; Xue, X.F.; Wu, L.M.; Cao, W. Honey Polyphenols Ameliorate DSS-Induced Ulcerative Colitis via Modulating Gut Microbiota in Rats. Mol. Nutr. Food Res. 2019, 63, 12. [Google Scholar] [CrossRef]
- Fiore, E.; Van Tyne, D.; Gilmore, M.S. Pathogenicity of Enterococci. Microbiol. Spectr. 2019, 7, 23. [Google Scholar] [CrossRef]
- Rashid, T.; Wilson, C.; Ebringer, A. The Link between Ankylosing Spondylitis, Crohn’s Disease, Klebsiella, and Starch Consumption. Clin. Dev. Immunol. 2013, 9, 872632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Reau, A.J.; Suen, G. The Ruminococci: Key symbionts of the gut ecosystem. J. Microbiol. 2018, 56, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Bui, T.P.N.; Shetty, S.A.; Lagkouvardos, I.; Ritari, J.; Chamlagain, B.; Douillard, F.P.; Paulin, L.; Piironen, V.; Clavel, T.; Plugge, C.M.; et al. Comparative genomics and physiology of the butyrate-producing bacterium Intestinimonas butyriciproducens. Environ. Microbiol. Rep. 2016, 8, 1024–1037. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Kang, K.; Li, Y.L.; Sang, L.X.; Chang, B. Tea polyphenols protect mice from acute ethanol-Induced liver injury by modulating the gut microbiota and short-chain fatty acids. J. Funct. Foods 2021, 87, 10. [Google Scholar] [CrossRef]
- Garcia-Mantrana, I.; Selma-Royo, M.; Alcantara, C.; Collado, M.C. Shifts on Gut Microbiota Associated to Mediterranean Diet Adherence and Specific Dietary Intakes on General Adult Population. Front. Microbiol. 2018, 9, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhu, J.Y.; Zhang, X.; Zhao, D.G.; Ma, Y.Y.; Li, D.L.; Ho, C.T.; Huang, Q.R. Aged citrus peel (chenpi) extract causes dynamic alteration of colonic microbiota in high-fat diet induced obese mice. Food Funct. 2020, 11, 2667–2678. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; You, J.M.; Wang, Z.R.; Liu, Y.; Wang, B.; Du, M.; Zou, T.D. Curcumin alleviates high-fat diet-induced hepatic steatosis and obesity in association with modulation of gut microbiota in mice. Food Res. Int. 2021, 143, 12. [Google Scholar] [CrossRef]
- Precup, G.; Vodnar, D.C. Gut Prevotella as a possible biomarker of diet and its eubiotic versus dysbiotic roles: A comprehensive literature review. Br. J. Nutr. 2019, 122, 131–140. [Google Scholar] [CrossRef]
- Gonzalez-Sarrias, A.; Romo-Vaquero, M.; Garcia-Villalba, R.; Cortes-Martin, A.; Selma, M.V.; Espin, J.C. The Endotoxemia Marker Lipopolysaccharide-Binding Protein is Reduced in Overweight-Obese Subjects Consuming Pomegranate Extract by Modulating the Gut Microbiota: A Randomized Clinical Trial. Mol. Nutr. Food Res. 2018, 62, 10. [Google Scholar] [CrossRef]
- van der Hee, B.; Wells, J.M. Microbial Regulation of Host Physiology by Short-chain Fatty Acids. Trends Microbiol. 2021, 29, 700–712. [Google Scholar] [CrossRef]
- Liu, Y.; Lin, Q.; Huang, X.; Jiang, G.T.; Li, C.; Zhang, X.; Liu, S.L.; He, L.Y.; Liu, Y.L.; Dai, Q.Z.; et al. Effects of Dietary Ferulic Acid on the Intestinal Microbiota and the Associated Changes on the Growth Performance, Serum Cytokine Profile, and Intestinal Morphology in Ducks. Front. Microbiol. 2021, 12, 13. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.Y.; Xiao, D.; Liu, W.; Song, Y.F.; Zou, B.R.; Li, L.; Li, P.; Cai, Y.; Liu, D.L.; Liao, Q.F.; et al. Intake of Ganoderma lucidum polysaccharides reverses the disturbed gut microbiota and metabolism in type 2 diabetic rats. Int. J. Biol. Macromol. 2020, 155, 890–902. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.J.; Yan, Y.M.; Wan, P.; Dong, W.; Huang, K.Y.; Ran, L.W.; Mi, J.; Lu, L.; Zeng, X.X.; Cao, Y.L. Effects of long-term intake of anthocyanins from Lycium ruthenicum Murray on the organism health and gut microbiota in vivo. Food Res. Int. 2020, 130, 11. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.J.; Chang, Y.G.; Gao, Y.; Wang, X.; Chen, X.; Wang, Y.M.; Xue, C.H.; Tang, Q.J. Dietary fucoidan of Acaudina molpadioides alters gut microbiota and mitigates intestinal mucosal injury induced by cyclophosphamide. Food Funct. 2017, 8, 3383–3393. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Cao, J.H.; Jin, H.Q.; Shan, Y.K.; Fang, J.; Liu, F. Beneficial impacts of fermented celery (Apium graveolens L.) juice on obesity prevention and gut microbiota modulation in high-fat diet fed mice. Food Funct. 2021, 12, 9151–9164. [Google Scholar] [CrossRef]
- Li, X.; Zeng, F.; Huang, Y.F.; Liu, B. The Positive Effects of Grifola frondosa Heteropolysaccharide on NAFLD and Regulation of the Gut Microbiota. Int. J. Mol. Sci. 2019, 20, 5302. [Google Scholar] [CrossRef] [Green Version]
- Tian, B.M.; Zhao, J.H.; Zhang, M.; Chen, Z.F.; Ma, Q.Y.; Liu, H.C.; Nie, C.X.; Zhang, Z.Q.; An, W.; Li, J.X. Lycium ruthenicum Anthocyanins Attenuate High-Fat Diet-Induced Colonic Barrier Dysfunction and Inflammation in Mice by Modulating the Gut Microbiota. Mol. Nutr. Food Res. 2021, 65, 16. [Google Scholar] [CrossRef]
- Simpson, C.A.; Diaz-Arteche, C.; Eliby, D.; Schwartz, O.S.; Simmons, J.G.; Cowan, C.S.M. The gut microbiota in anxiety and depression—A systematic review. Clin. Psychol. Rev. 2021, 83, 18. [Google Scholar] [CrossRef]
- Lin, Y.S.; Zheng, X.H.; Chen, J.F.; Luo, D.D.; Xie, J.H.; Su, Z.R.; Huang, X.Q.; Yi, X.Q.; Wei, L.; Cai, J.; et al. Protective Effect of Bruguiera gymnorrhiza (L.) Lam. Fruit on Dextran Sulfate Sodium-Induced Ulcerative Colitis in Mice: Role of Keap1/Nrf2 Pathway and Gut Microbiota. Front. Pharmacol. 2020, 10, 17. [Google Scholar] [CrossRef] [Green Version]
- Sorbara, M.T.; Pamer, E.G. Microbiome-based therapeutics. Nat. Rev. Microbiol. 2022, 20, 365–380. [Google Scholar] [CrossRef]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, Y.; Wang, Y.; Xu, H.; Mei, X.; Yu, D.; Wang, Y.; Li, W. Antioxidant Properties of Probiotic Bacteria. Nutrients 2017, 9, 521. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | Retention Time/Min | MS (m/z) | MS2 Ions (Relative Abundance) (m/z) | Molecular Formula | Tentative Identification | References |
|---|---|---|---|---|---|---|
| 1 | 6.0576 | 465.1039; [M]+ | 303.0496 (100) | C21H21O12 | Delphinidin hexoside (Delphinidin-3-O-galactoside or delphinidin-3-O-glucoside) | [19,20] |
| 2 | 6.3155 | 611.162; [M]+ | 287.0556 (100), 611.1637 (13.08) | C27H31O16 | Cyanidin-3-O-sophoroside | [21] |
| 3 | 6.7586 | 757.2215; [M]+ | 287.0552 (100), 757.2215 (38.43) | C33H41O20 | Cyanidin-3-O-2G-glucosylrutinoside | [21] |
| 4 | 6.9068 | 449.109; [M]+ | 287.0558 (100) | C21H20O11 | Cyanidin-3-O-glucoside | [22] |
| 5 | 7.5025 | 595.1671; [M]+ | 287.0557 (100), 595.1650 (11.67) | C27H30O15 | Cyanidin-3-O-rutinoside | [21,23] |
| 6 | 7.5919 | 479.1196; [M]+ | 317.0665 (100) | C22H23O12 | Petunidin hexoside (Petunidin-3-O-galactoside or Petunidin-3-O-glucoside) | [19,20] |
| 7 | 7.9218 | 433.1144; [M]+ | 271.0604 (100) | C21H21O10 | Pelargonidin-3-O-glucoside | [23] |
| 8 | 7.9453 | 419.0982; [M]+ | 287.0551 (100) | C20H19O10 | Cyanidin-3-O-arabinoside | [19] |
| 9 | 8.6335 | 579.1721; [M]+ | 271.0616 (100) | C27H31O14 | Pelargonidin-3-O-rutinoside | [23] |
| 10 | 8.6412 | 449.1088; [M]+ | 317.0662 (100) | C21H21O11 | Petunidin-3-O-arabinoside | [24] |
| 463.1247; [M]+ | 301.0707 (100) | C22H23O11 | Peonidin hexoside (Peonidin-3-O-galactoside or peonidin-3-O-glucoside) | [20,21,25] | ||
| 11 | 8.7697 | 468.2126; [M-H]− | 135.0457 (62.47), 289.1558 (19.2), 306.1827 (30.77), 332.1622 (100), 468.2156 (70.9) | C25H31N3O6 | N,N′-dicaffeoylspermidine | [25] |
| 12 | 9.0895 | 419.0983; [M]+ | 287.0559 (100) | C20H18O10 | Cyanidin-3-O-xyloside | [23] |
| 13 | 9.18 | 493.1349; [M]+ | 331.0823 (100) | C23H25O12 | Malvidin-3-O-galactoside | [19,24] |
| 14 | 9.7379 | 493.1352; [M]+ | 331.0824 (100) | C23H25O12 | Malvidin-3-O-glucoside | [19,24] |
| 15 | 9.9434 | 535.1094; [M]+ | 287.0547 (100) | C24H22O14 | Cyanidin-3-O-(6-O-malonyl-β-d)-glucoside | [23] |
| 16 | 10.2993 | 463.1243; [M]+ | 331.0809 (100) | C22H23O11 | Malvidin-3-O-arabinoside | [24] |
| 17 | 10.7913 | 593.1518; [M]+ | 287.0553 (100) | C25H20O17 | Cyanidin-3-O-dioxalylglucoside | [23] |
| 18 | 10.9592 | 919.2509; [M]+ | 303.0512 (100), 465.105 (12.72), 757.2022 (25.8), 919.2569 (60.23) | C42H47O23 | Delphinidin-3-O-rutinoside(trans-p-coumaroyl)-5-O-glucoside | [26] |
| 19 | 11.1733 | 519.1152; [M]+ | 271.0609 (100) | C24H23O13 | Pelargonidin-3-O-malonylglucoside | [23] |
| 20 | 12.4938 | 933.2703; [M]+ | 317.0668 (100), 479.1206 (33.98), 711.2168 (57.19), 933.2718 (56.51) | C43H48O23 | Petunidin-3-O-rutinoside(cis-p-coumaroyl)-5-O-glucoside or petunidin-3-O-rutinoside(trans-p-coumaroyl)-5-O-glucoside | [25] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Shu, Y.; Chen, Y.; Ge, Z.; Zhang, C.; Cao, J.; Li, X.; Wang, Y.; Sun, C. Evaluation of Antioxidant Capacity and Gut Microbiota Modulatory Effects of Different Kinds of Berries. Antioxidants 2022, 11, 1020. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11051020
Chen J, Shu Y, Chen Y, Ge Z, Zhang C, Cao J, Li X, Wang Y, Sun C. Evaluation of Antioxidant Capacity and Gut Microbiota Modulatory Effects of Different Kinds of Berries. Antioxidants. 2022; 11(5):1020. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11051020
Chicago/Turabian StyleChen, Jiebiao, Yichen Shu, Yanhong Chen, Zhiwei Ge, Changfeng Zhang, Jinping Cao, Xian Li, Yue Wang, and Chongde Sun. 2022. "Evaluation of Antioxidant Capacity and Gut Microbiota Modulatory Effects of Different Kinds of Berries" Antioxidants 11, no. 5: 1020. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11051020