
Physiological and Clinical Aspects of Bioactive Peptides from Marine Animals


1
Division of Energy and Bioengineering, Dongseo University, Busan 47011, Korea
2
Institute of Fluid Mechanics, Friedrich-Alexander-University Erlangen-Nürnberg, 91058 Erlangen, Germany
3
German Engineering Research and Development Center, LSTME-Busan Branch, Busan 46742, Korea
*
Author to whom correspondence should be addressed.
Antioxidants 2022, 11(5), 1021; https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11051021
Submission received: 28 April 2022
/
Revised: 19 May 2022
/
Accepted: 19 May 2022
/
Published: 22 May 2022
(This article belongs to the Section Health Outcomes of Antioxidants and Oxidative Stress)
Abstract
:Biological molecules in nutraceuticals and functional foods have proven physiological properties to treat human chronic diseases. These molecules contribute to applications in the food and pharmaceutical industries by preventing food spoilage and cellular injury. Technological advancement in the screening and characterization of bioactive peptides has enabled scientists to understand the associated molecules. Consistent collaboration among nutritionists, pharmacists, food scientists, and bioengineers to find new bioactive compounds with higher therapeutic potential against nutrition-related diseases highlights the potential of the bioactive peptides for food and pharmaceutic industries. Among the popular dietary supplements, marine animals have always been considered imperative due to their rich nutritional values and byproduct use in the food and pharmaceutical industries. The bioactive peptides isolated from marine animals are well-known for their higher bioactivities against human diseases. The physiological properties of fish-based hydrolyzed proteins and peptides have been claimed through in vitro, in vivo, and clinical trials. However, systematic study on the physiological and clinical significance of these bioactive peptides is scarce. In this review, we not only discuss the physiological and clinical significance of antioxidant and anticancer peptides derived from marine animals, but we also compare their biological activities through existing in vitro and in vivo studies.
Keywords:
anticancer; antioxidant; biopharmaceuticals; clinical; fish; functional foods; in vitro; in vivo; protein hydrolysate1. Introduction
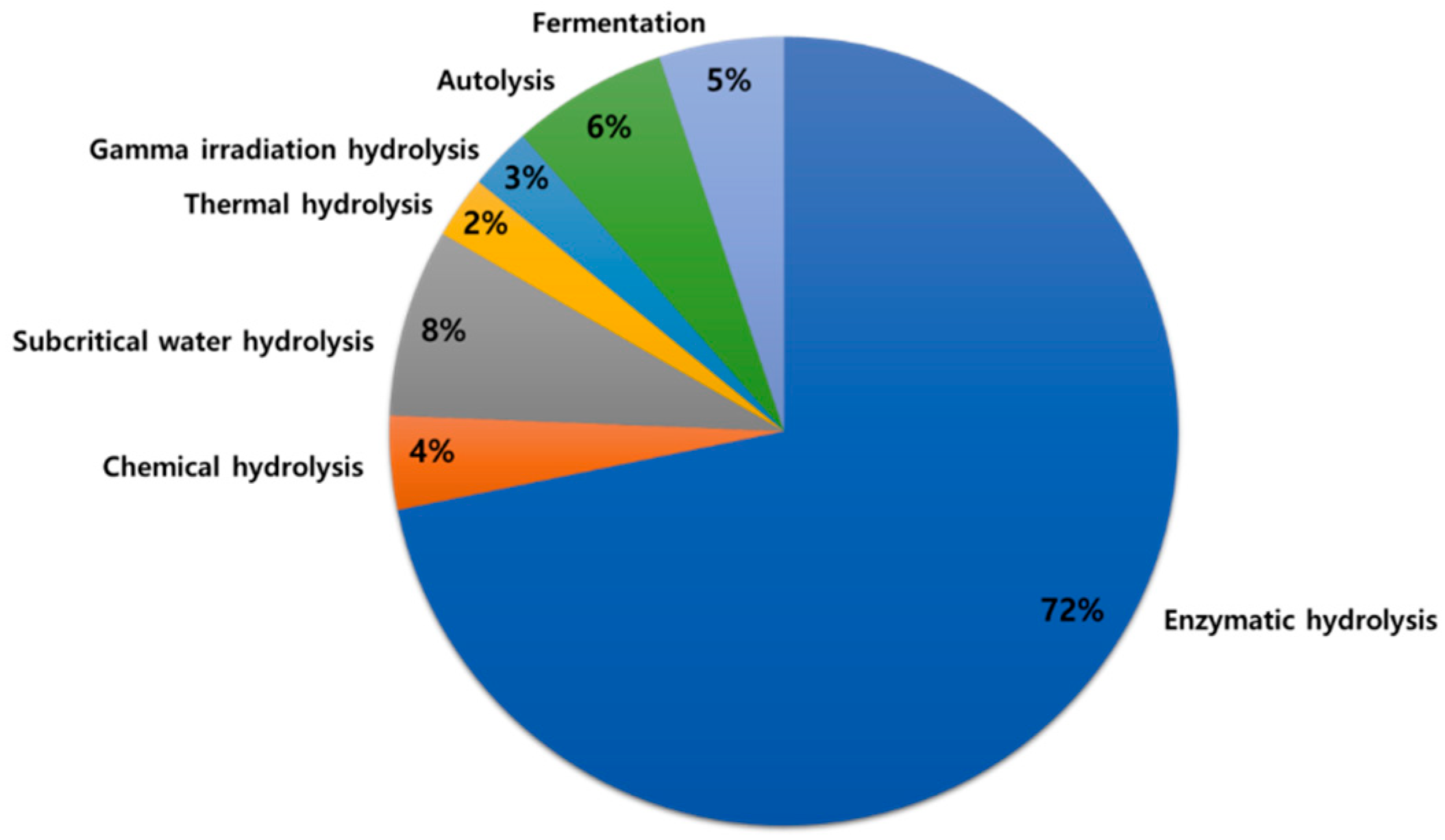
Proteins are important nutrients for sustaining life and supporting normal body functions. Proteins and peptides from marine sources provide a rich source of essential amino acids required for human daily needs [1,2]. Inactive peptides within native proteins require proteolytic release (in vivo digestion) or hydrolysis to reveal biological activity [3,4]. Protein hydrolysis results in the conversion of intact proteins into peptides, usually containing no more than 20 amino acids [4]. Current methods for hydrolysis of marine animal muscle and processing waste byproducts include chemical (acid, alkali, or catalytic) hydrolysis [5], enzymatic hydrolysis [6], gamma irradiation hydrolysis [7], subcritical water hydrolysis [8], thermal hydrolysis [9], autolysis [10], and bacterial fermentation [11] (Figure 1). Among those, enzymatic hydrolysis is the most common method of producing hydrolyzed proteins and peptides. Enzymatic hydrolysis improves solubility, water-binding capacity, and heat stability of myofibrillar proteins, modifies emulsifying and foaming properties of hydrolyzed proteins, as well as improves the nutritional quality of foods [12,13]. Desirable physicochemical properties of a hydrolyzed protein could be achieved by controlling hydrolysis parameters, such as time, pH, temperature, and enzyme concentration [14]. The degree of hydrolysis, on the other hand, affects the size and amino acid profile of peptides, ultimately affecting the taste of the hydrolyzed proteins; for example, bitter taste is associated with a high degree of hydrolysis [15].
Bioactive compounds are either produced naturally from plants, animals, fungi, and microorganisms, such as carotenoids, phenolic compounds, polyphenols, and cordymin [16,17,18,19,20], or generated synthetically, such as propyl gallate, t-butylhydroquinone, butylated hydroxytoluene, and butylated hydroxy anisole [21,22]. Bioactive substances from natural sources have gained much interest in recent years due to consumer’s preferences and health concerns associated with the use of synthetic food additives [23]. The natural protein hydrolysates and peptides isolated from farm animals, such as cows, chickens, and pigs, are relatively cheap and are easily available sources of nutrients [24,25]. However, outbreaks of certain diseases, including avian influenza, mad cow disease, and foot-and-mouth disease, as well as limited use of extracted proteins from pig skin and bones due to religious reasons, have made it obligatory to find alternative sources [26,27]. Recently, the marine-derived protein hydrolysates and bioactive peptides have gained much attention due to their excellent nutritional composition, numerous health benefits and no reported side effects [6,28,29,30,31].
An enormous range of marine-based protein hydrolysates and peptides using different hydrolysis methods have been reported in the literature with significant antioxidant, anticancer, antihypertensive, antimicrobial, and immunomodulatory activities [32,33,34]. Antihypertensive peptides or angiotensin-converting enzyme (ACE) inhibiting peptides have been extensively studied [35]. Due to the side effects, such as taste disturbance and skin rashes, associated with the use of synthetic drugs [36], the interest in finding safer and eco-friendly ACE inhibitory compounds from natural sources has increased [37,38]. The ACE inhibitory properties of the peptides isolated from marine sources, including pacific cod skin [39], sardinella [40], flounder fish [41], and Takifugu bimaculatus skin [42], have been reported.
Antimicrobial peptides from marine sources, including algae, sponges, fungi, oysters, and fish, have been reported [19,43,44,45,46,47,48]. Marine antimicrobial peptides are classified into four families based on their structure [49]. These are (i) linear α-helical peptides, (ii) helical or non-helical peptides that are rich in proline, arginine, histidine, and/or glycine, (iii) hairpin-like β sheet or α helical/β-sheet mixed structure peptides with intra-molecular disulfide bonds, and (iv) cyclic peptides [19,49]. In addition, more than 60 antimicrobial peptide drugs are available on the market, and more than 140 peptides are currently in clinical trials [50]. For example, oyster peptide “CgPep33” shows bioactivities against Gram-positive and Gram-negative bacteria and fungi [48].
Immunomodulatory peptides prevent or treat immune-suppressing diseases by improving cytochrome or antibody synthesis and supplementing mucosal immunity in the gastrointestinal tract [51]. The protein hydrolysates and peptides from marine sources have shown significant immunomodulatory properties [52]. For example, a low-molecular-weight peptide isolated from marine Nibea japonica skin dramatically improved IgG and IgM levels in blood [53]. Furthermore, due to rising cancer cases, the interest in finding a suitable cancer therapy is consistently gaining attention. Traditional cancer treatments, including surgery, chemotherapy, and radiotherapy, could affect healthy cells [54] and suppress the human immune system [55]. The commercially available drugs from natural sources containing bioactive peptides account for 60% of the available anticancer drugs [56]. It could be due to the ability of the anticancer peptides to penetrate and directly bind to the cancer cells via electrostatic interaction [57]. Recent research has claimed that anticancer compounds from marine sources, especially marine natural bioactive products [58] and nanomaterials, as a safer alternative, allowing targeted treatment to cancer cells and not inducing damage to the healthy cells [59,60].
Peptides isolated from marine sources contain essential amino acids [61] and are used in animal and human nutrition. Due to their rich nutritional profile and bioactive properties, marine peptides have several commercial applications in the food, cosmetics, and pharmaceutical industries [62,63]. The in vitro bioactivities of protein hydrolysates have been discussed in the literature; however, a systematic study on the physiological and clinical significance of marine-based purified bioactive peptides is scarce. Hence, there is a need to critically review the physiological and clinical properties of purified bioactive peptides from marine animals. In this review, we discuss the physiological and clinical significance of antioxidant and anticancer properties of peptides purified from marine animals through selected in vitro, in vivo, and clinical studies.
2. Methods
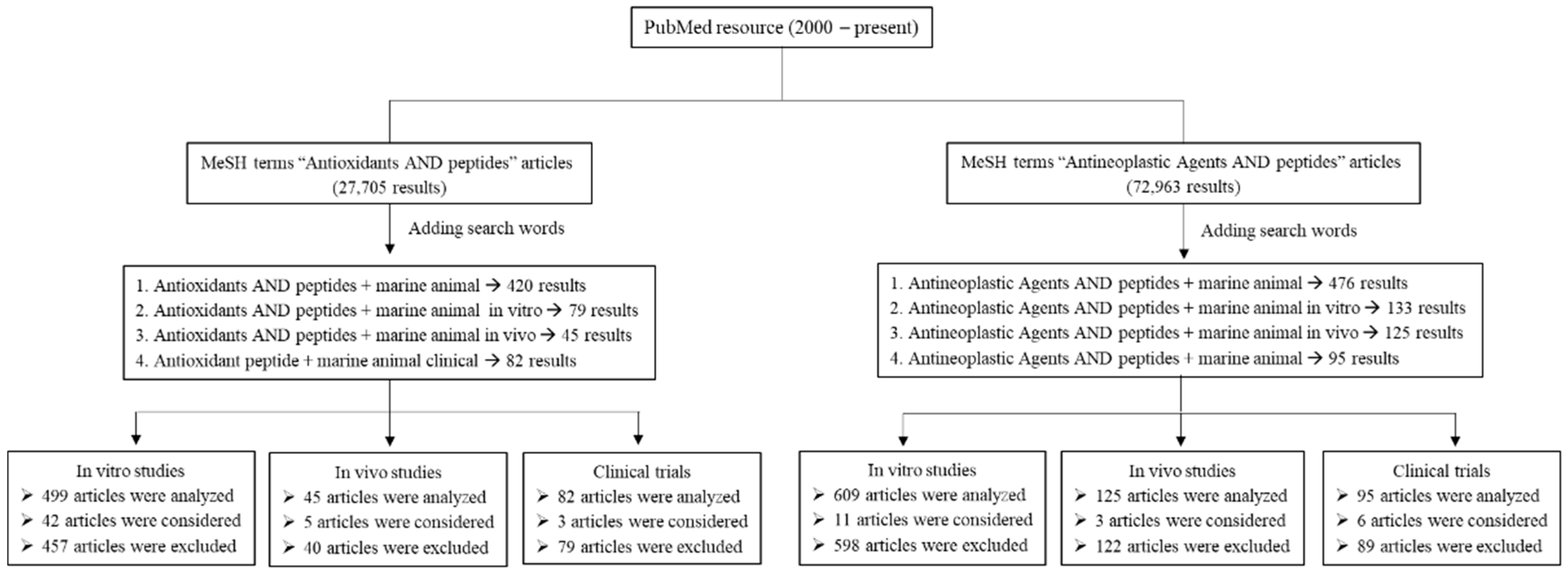
The literature search was conducted using the PubMed database from 2000 to present. Articles were mostly written in English (in vitro, in vivo, and clinical studies). The key search using a comprehensive list of MeSH (Medical Subject Headings) terms followed: Antioxidants AND peptides, Antineoplastic Agents AND peptides. Additional search words included “marine animal”, “marine animal in vitro”, “marine animal in vivo”, and “marine animal clinical”. The search strategy and results are presented in a schematic diagram (Figure 2). Articles were selected based on their relevance to the topic of this review. For antioxidant studies, 626 articles were analyzed, 576 articles were excluded, and 50 articles were considered in this review. For anticancer studies, 829 articles were analyzed, 809 articles were excluded, and 20 articles were considered. In addition, references in each article were further reviewed to find any relevant study using ScienceDirect, Wiley Online Library, SpringerLink, and Google Scholar databases. Articles were extracted and sorted using EndNote to prevent duplicate citations.
3. Antioxidant Properties
An unbalanced electron in the outermost orbital of reactive oxygen species (ROS) makes these molecules unstable and enables them to react with biological macromolecules, such as proteins, lipids, and DNA [64]. The ROS, including superoxide anion radical (O2•−), hydrogen peroxide radical (H2O2), hydroxyl radical (•OH), peroxyl radical (ROO•), and hydroperoxyl radical (HO2•), are byproducts produced by mitochondrial aerobic metabolism [65]. An unbalance in the oxidation reaction chain and ROS ratio causes oxidative stress. This leads to the development of chronic diseases, such as diabetes, cardiovascular diseases, Alzheimer’s, neurodegenerative diseases, and particularly cancer [66], which is one of the leading causes of mortality worldwide [67]. The oxidative stress generated by ROS is suppressed by antioxidant defense mechanisms, including endogenous and exogenous defense systems [68]. Endogenous antioxidants, such as catalase (CAT), superoxide dismutase (SOD), glutathione (GSH), and glutathione peroxidase (GPx), and exogenous antioxidants, such as carotenoids, vitamin E, vitamin C, and polyphenols, act synergistically to inhibit oxidation reaction, maintain redox homeostasis and reduce cell deterioration via suppressing cell damage [68,69,70]. Therefore, maintaining a balance in antioxidants/ROS ratio is vital to avoid oxidative stress [71].
The antioxidant effects of peptides isolated from fish and other marine animals have been studied extensively in vitro and in vivo (including animal-based studies), while human clinical trials have rarely been reported. The antioxidant properties of peptides isolated from marine animals are listed in Table 1, Table 2 and Table 3 and are described below.
3.1. Antioxidant Effects of Bioactive Peptides from Marine Animals—In Vitro Studies
Proteolytic enzymes generally cleave proteins at a specific cleavage bond resulting in peptides and amino acids of varying sizes [117]. The results of enzyme proteolysis depend on the type of enzyme, temperature, pH, time, and enzyme/substrate ratio [118]. Bashir et al. reported high DPPH• scavenging activity (36.34%) of mackerel (Scomber japonicus) peptide, ALSTWTLQLGSTSFSASPM (1049.6 Da), isolated by protamex hydrolysis at 50 °C, pH 8, and in a short time of < 2 h [4]. Zhao and colleagues hydrolyzed Spanish mackerel (Scomberomorous niphonius) at 40 °C and pH 5–9 [92]. The resulted peptide, FGYDWW, showed high DPPH•, •OH, and O2•− scavenging activities of 86.52%, 84%, and 86%, respectively [92]. Similarly, Jang and co-workers observed high DPPH• scavenging activity of 90.66% at 1.0 mg/mL of sandfish peptide, ATSHH (551.25 Da) [104].
The structural features of a peptide, such as size, molecular weight, sequence, and amino acid composition, could directly affect the antioxidant activity of a peptide. The low molecular weight of a peptide (< 1.5 kDa) has been linked with higher ROS radical scavenging activities [4,74,87,95,98]. A low-molecular-weight peptide, TCGGQGR (678 Da) from mackerel byproducts, showed DPPH• scavenging activity of 96% and ABTS• scavenging activity of 100% [95]. The tuna backbone peptide, VKAGFAWTANQQLS (1.5 kDa), showed up to 90% •OH scavenging activity at a concentration of 0.05 mg/mL, and no toxicity to human fetal lung fibroblast cells was reported [73]. Exceptionally, some high molecular weight peptides, such as yellowfin sole peptide, RPNFDLEPPY (13 kDa), have also shown high antioxidant activities [72].
The presence of a hydrophobic amino acid, such as glycine (G), leucine (L), isoleucine (I), and alanine (A) in a peptide sequence could enhance the antioxidant activity of a peptide [4]. These hydrophobic amino acids allow access to the hydrophobic targets, such as cell membrane, and enhance bioavailability [4,119]. Asaduzzaman et al. reported high DPPH• (50%) and ABTS• (40%) scavenging activity of mackerel peptide having glycine [94]. A low-molecular-weight (747 Da) peptide from giant squid (Dosidicus gigas), NGLEGLK having glycine, showed reduced cytotoxicity against human embryonic lung fibroblasts at IC50 of 304–578 µM [86]. In another study, hoki (Johnius belengerii) peptide (1.8 kDa) showed reduced cytotoxicity to human fetal fibroblasts cells at IC50 of 17–172 µM and induced •OH scavenging activity [91]. The peptide, CAAP (360 Da) from flounder having alanine, showed potent DPPH• scavenging activity at IC50 of 26.89 µM [80]. The sturgeon peptide, GDRGESGPA (845.37 Da), having glycine and alanine, showed DPPH• scavenging activity of 40% [101]. Hairtail surimi peptide, DLYANTVLSGGTTMYPGIADR (2.2 kDa), having leucine, glycine, isoleucine, and alanine, showed DPPH• and •OH scavenging activity of 67.07% and 62.08%, respectively [100]. Furthermore, oyster peptide, LANAK (515.29 Da), showing DPPH• scavenging activity of 83.79%, also contained leucine and alanine [87]. The antioxidant peptides from the skin of horse mackerel, NHREDR (856 Da), and croaker, GNRGFACRHA (1101.5 Da), showed DPPH• scavenging activity of 72.3% and 52.7%, respectively, and •OH scavenging activity of 51.2% and 40.3%, respectively [83]. In another study, peptide—ACFL (518.5 Da), isolated from horse mackerel (Magalaspis cordyla) viscera, showed DPPH• and •OH scavenging activity of 57.8% and 45.2%, respectively [93]. These peptides also contained at least one hydrophobic amino acid in their sequences.
The presence of a polar amino acid such as tyrosine (Y) at the C-terminus of a peptide sequence has also been linked with higher antioxidant activities [76,120,121]. The tilapia (Oreochromis niloticus) peptides, DCGY (456.12 Da) and NYDEY (702.26 Da), having tyrosine, showed H2O2 scavenging activity at IC50 of 27.6 µg/mL and 38.4 µg/mL, respectively [75]. Similarly, tilapia skin peptide, YGDEY (645.21 Da), showed •OH scavenging activity at IC50 of 4.61 µg/mL [76], and tilapia scale peptide, GYDGY (646.23 Da), showing potent DPPH• scavenging activity at IC50 of 1.6 µg/mL, contained tyrosine [78]. The Alaska pollock (Theragra chalcogramma) peptide, LPHSGY (672 Da), showed high •OH scavenging activity of 35% at a concentration of 53.6 µM and also contained tyrosine [88].
The marine animal-based antioxidant peptides were explored from different marine fish species, where mackerel and tilapia contributed the most. The antioxidant peptides prepared using enzymatic hydrolysis at temperatures of < 50 °C and pH < 9 resulted in higher antioxidant activities. Irrespective of the source of isolation, smaller molecular weight peptides showed higher radical scavenging potential. Interestingly, the presence of a hydrophobic amino acid and/or a polar amino acid, such as tyrosine, in the peptide sequence showed higher scavenging activities. Despite the promising in vitro activities, most of the above-mentioned peptides have not been evaluated under in vivo or clinical conditions.
3.2. Antioxidant Effects of Bioactive Peptides from Marine Animals—In Vivo Studies
Nazeer et al. purified the peptide KTFCGRH with a molecular weight of 861 Da from a croaker and studied the antioxidant effects in Wistar rats. A total of 100 µg/kg of purified peptide and 20% ethanol were orally administered to experimental rats for 15 days. The rats fed with purified peptides showed improvement in cellular antioxidant enzyme systems and displayed serum CAT, SOD, and glutathione s-transferase (GST) levels of 283.6, 28.42, and 4.3 U/mg protein, respectively. In contrast, the ethanol-administered rates showed CAT, SOD, and GST levels of 196.4, 15.1, and 1.3 U/mg protein, respectively [110]. The purified peptide from horse mackerel viscera, ACFL with a molecular weight of 518 Da, was evaluated in Wistar rats for 15 days. The results showed that 100 µg/kg of peptide and 20% of ethanol-administered rats showed serum CAT, SOD, and GST levels of 290.8, 29.45, and 3.93 U/mg protein, respectively. In contrast, the ethanol-administered rats showed CAT, SOD, and GST levels of 196.4, 15.1, and 1.3 U/mg protein, respectively [111]. Antioxidant properties of the peptide WHKNCFRCAKCGKSL purified from snakehead murrel (Channa striatus) were investigated in wild-type zebrafish. This peptide showed no cytotoxicity to zebrafish larvae at a concentration of 50 µM, and a decrease in malondialdehyde (MDA) levels to 14 µmol/min/mg protein was observed. In contrast, SOD and CAT expressions increased to 22 and 17 U/mg protein, respectively [106].
Hu et al. studied the protective effects of tilapia scales-purified collagen oligopeptides in specific pathogen-free (SPF) Sprague Dawley rats for 30 days [113]. The oral administration of collagen peptides significantly reduced gastric and duodenal ulcer index and MDA content. In comparison, increases in gastric juice pH and expressions of CAT, SOD, and glutathione peroxidase (GPx) were observed [113]. Furthermore, the oligopeptides purified from frigate tuna (Auxis thazard) showed hypouricemic properties. Seven days of treatment of SPF hyperuricemic Kunming mice with tuna oligopeptides resulted in increased SOD and CAT levels of 125 and 0.75 U/mg protein, respectively [112].
The purified marine antioxidant peptides were evaluated using different in vivo models, including albino Wistar rats, SPF-SD mice, and wild zebrafish. The study parameters included changes in body weights, body weight gains, levels of intracellular ROS, levels of lipid peroxidation, and expressions of antioxidant enzyme systems, such as SOD, CAT, GST, and GPx. The reported antioxidant peptides showed enhanced expressions of antioxidant enzymes, reduced MDA levels, and helped to recover oxidative stress.
3.3. Antioxidant Effects of Bioactive Peptides from Marine Animals—Clinical Trials
Human clinical studies of purified antioxidant peptides from marine animals have rarely been reported. Kim et al. reported skin improving properties of low-molecular-weight collagen peptide (LMWCP) from Sutchi catfish skin (Pangasius hypophthalmus) in 40–60-year-old healthy women [114]. Skin hydration was significantly improved (6-fold of placebo), and crow’s feet decreased to 2.81 AU when compared to placebo after 6 weeks of treatment. Average roughness of skin wrinkling—0.17 AU (lower than placebo group) and average roughness of skin elasticity—0.71 AU (higher than placebo group) was noted after 12 weeks. In another study, skin improving properties of CELERGEN® were investigated in 41 adults, including five females. CELERGEN® is a commercial oral supplement that includes marine collagen peptide from the skin of pollock (Pollachius virens), Atlantic halibut (Hippoglossus hippoglossus), European plaice (Pleuronectes platessa), and plant-derived antioxidants [116]. After 4 months of treatment, the thickness and acoustic density of the dermis layer improved to 4133 µm and 6.3, respectively. However, GPx/GST activity and thickness and acoustic density of the epidermis layer were not changed. Another commercial product, GOLD COLLAGEN® ACTIVE, produced from fish collagen, showed skin improving properties when evaluated in 122 subjects [115]. This supplement consists of hydrolyzed fish collagen, vitamins, and other active ingredients. It improved skin elasticity by 40% in the younger age group and reduced joint pain by 43% in the elder age group when compared to the placebo group.
The reported clinical studies on purified antioxidant peptides from marine animals have focused on skin properties, such as skin elasticity, skin hydration, and wrinkling. Two of the above-mentioned products have already been approved and available commercially. Nevertheless, some peptides from marine animals have shown prominent in vitro and in vivo antioxidant activities, and clinical trials are lacking, which need to be performed to better understand their bioactivities.
4. Anticancer Properties
Anticancer peptides have been isolated from various marine animals, such as fish, oysters, mussels, and snails. Anticancer peptides are selective to cancer cells and kill cancerous cells by inhibiting angiogenesis, disrupting tubulin-microtubule balance, and apoptosis [122]. The anticancer properties of several peptides from marine animals have been investigated in vitro and in vivo, including in animal-based studies, and some of these peptides have even been used in human clinical trials. The anticancer properties of peptides isolated from marine animals are listed in Table 4, Table 5 and Table 6 and are described below.
4.1. Anticancer Effects of Bioactive Peptides from Marine Animals—In Vitro Studies
Anticancer peptides from marine animals, especially the phylum Mollusca, have been extensively reported. The peptide, LKEENRRRRD, isolated from sepia ink (Sepia esculenta), inhibited the proliferation of prostate cancer (PC-3) cells and showed a 3.3-fold increase in early apoptosis as compared to the control [127]. In another study, the peptide, AFNIHNRNLL, from mussel (Mytilus coruscus) inhibited various cancer cell lines, including prostate cancer (PC-3), lung cancer (A-549), and breast cancer (MDA-MB-231) cell lines at LC50 of 0.94, 1.41, and 1.22 mg/mL, respectively, and showed no toxicity to the normal cells [125]. The peptide, LANAK (515 Da), isolated from oyster (Saccostrea cucullata), not only showed strong antioxidant activity [87], but it also showed significant anticancer activity against human colon cancer (HT-29) cells at IC50 of 90 µg/mL and showed no cytotoxicity to normal kidney epithelial cells of the African Green Monkey [87]. Kahalalide F (KF), derived from marine mollusk (Elysia rufescens), showed anticancer activity by reducing DNA synthesis in prostate cancer cell lines (DU145, PC3, and LNCaP at IC50 of 0.18, 0.07, and 0.26 µM, respectively), breast cancer cell lines (MCF-7, SKBR-3, MDA-MB-231, and BT474 at IC50 of 0.28, 0.23, 0.39, and 0.26 µM, respectively), colon cancer cell line (LoVo at IC50 of 0.16 µM), and normal cell lines (IMR90, MCF10A, HMEC-1, and HUVEC at IC50 of 3.13, 2.44, 1.88 and 1.62 µM, respectively) [131].
Besides mollusks, the anticancer activity of fish has also been claimed. Finless sole peptide, GFFALIPKIISSPLFKTLLSAVGSALSSSGGQE, known as pardaxin, inhibited colony formation in MN-11 cells (<90% inhibition) [126]. The Japanese flounder (Paralichthys olivaceus) peptides, RKQCIRKCIRRREPHGKMMIRIRRK and KKYRSQRKIRRMRR-KRKYPSFMQ, induced dose-dependent necrosis in HT-29 cells at a concentration of 500 µM [128]. Spanish mackerel peptides showed a more than 80% survival rate in mouse melanoma cancer (B16F10) cells [129]. The tilapia peptide, GIKCRFCCGCCTPGICGVCCRF-NH2, showed a concentration-dependent (50 and 100 µg/mL) inhibition of HeLa and HT1080 cells after 24–72 h treatment. The tilapia peptide inhibited colony formation in HeLa and HT1080 cells by 60% and in HepG2 cells by 50% [130]. Low-molecular-weight antioxidant peptide, FIMGPY (726.9 Da), from skate (Raja porosa) also showed significant anticancer activity against HeLa cells at IC50 of 4.81 mg/mL and no toxicity to normal NIH3T3 cells [120].
Recent studies have claimed significant anticancer properties of marine-derived peptides, even at low concentrations, such as anticancer peptides isolated from tuna (Thunnus tonggol) [123]. The tuna peptides, LPHVLTPEAGAT and PTAEGGVYMVT, were active against breast cancer (MCF-7) cells at IC50 of 8.1–0.7 [123]. The isolated peptides had a molecular weight of 390 Da to 1.4 kDa. In another study, a low-molecular-weight peptide (440 Da) isolated from anchovy sauce inhibited human lymphoma (U937) cells by 50% at 31 µg/mL [124]. Interestingly, these peptides also contained hydrophobic amino acids.
The peptides purified from marine animals, including marine mollusks, tilapia, mackerel, and oysters, showed significant in vitro anticancer activities against breast, leukemia, colon, fibrosarcoma, prostate, and cervical cancers. Interestingly, low-molecular-weight peptides containing hydrophobic amino acids showed high anticancer activities. A similar observation was noted in the case of antioxidant peptides. Irrespective of significant in vitro anticancer activities, in vivo activities of these peptides have hardly been reported.
4.2. Anticancer Effects of Bioactive Peptides from Marine Animals—In Vivo Studies
Marine animals from different phyla, especially Mollusca and Porifera, have revealed significant anticancer activities, which have been tested through in vitro and in vivo studies. Venoms of marine cone snails, genus Conus, containing more than 100,000 small bioactive peptides, have been used to develop potential therapeutic agents for cardiovascular and nervous systems [133]. The anticancer activity of marine vermivorous cone snail (Conus vexillum) venom was investigated by Abdel-Rahman and colleagues using Swiss albino mice injected with Ehrlich’s ascites carcinoma (EAC) cells [133]. Conus venom significantly increased various oxidative stress biomarkers (protein carbonyl content, lipid production, and reactive nitrogen intermediates) of EAC cells after 3–12 h of venom injection. In addition, it significantly reduced CAT and SOD expression levels of EAC cells.
Pardaxin peptide was discovered as a natural agent to inhibit oral squamous cancer [132]. It is derived from finless sole (Pardachirus marmoratus) and have been tested on hamster using an oral squamous cell carcinoma model [132]. Pardaxin at a dose of 75 mg/kg reduced tumor volume to more than half. Furthermore, a 500% decrease in serum prostaglandin E2 (a cancer inducer) levels was observed after taking pardaxin with 5-fluorouracil (an antimetabolite used in clinical therapy). In another study, 14 days of treatment with a high dose of synthetic pardaxin (13 µg/mL) showed a 3-fold reduction in tumor volume of MN-11 cells in C57BL/6 mice, while no side effects were observed in the normal cells [126].
4.3. Anticancer Effects of Bioactive Peptides from Marine Animals—Clinical Trials
The anticancer activity of marine animal-derived individual peptides has not been claimed through clinical trials; however, two decades ago, some anticancer compounds containing marine animal-derived peptides were reported, which are currently known as natural chemotherapeutic agents such as Kahalalide F (KF) [140]. KF is a depsipeptide with a composition ranging from tripeptide to tridecapeptide [141]. KF was first isolated from marine mollusk (Elysia rufescens), and it is currently in clinical phase II. KF has shown significant antitumor activities in prostate cancer patients at a recommended dose of 560 µg/m2 [135]. Another clinical study on 38 cancer patients (colorectal cancer, melanoma cancer, breast cancer, and others) recommended a KF dose of 650 µg/m2 [134]. However, dose-limiting toxicities were observed after 1 h intravenous infusion [134]. Martín-Algarra and co-workers confirmed 650 µg/m2 as the safe recommended dose of KF and suggested KF peptide as a safe chemotherapeutic agent [136]. The KF-treated cancer patients showed an average overall survival of 10.8 months, non-cumulative toxicity, and no side effects such as leukopenia or thrombocytopenia when used with other compounds. However, the use of KF alone for cancer treatment has not yet been documented for advanced malignant patients.
Another marine mollusk (Dolabella auricularia) peptide, DOLA-10, currently in clinical trial phase II [138], showed no changes in tumor volume in clinical phase I trials on 22 cancer patients at a maximum dose of 300 µg/m2 [137]. Pitot et al. recommended a dose of 400 µg/m2 for patients requiring less than two chemotherapies and 325 µg/m2 for patients requiring more than two chemotherapies [142]. Therefore, DOLA-10 at a dose of 400 µg/m2 was used in clinical phase II [138]. Krug et al. studied 400 µg/m2 of DOLA-10 in 10 NSCLC patients [138], where only 2 patients had a stable disease, 3 patients had grade-4 neutropenia, and 2 patients had grade-3 hyperglycemia. Later, Margolin and colleagues again studied DOLA-10 at 400 µg/m2 [139] and observed the pharmacokinetic profile at the total body clearance and volume of distribution in the body at a steady state of 2.61 ± 1.9 L/h/m2 and 28.4 ± 13 L/m2. The above-mentioned studies highlight the potential anticancer properties of marine animal-derived peptides; however, further clinical studies are needed to better understand their mechanism of action.
5. Conclusions and Future Perspectives
The isolation, purification, and characterization of bioactive peptides from marine sources have significantly increased in the last few years. The marine processing byproducts and the underutilized marine organisms, especially fish, have been highlighted as potential sources of bioactive peptides. Different methods, such as proteolytic hydrolysis, chemical hydrolysis, subcritical water hydrolysis, and autolysis, have been used to isolate peptides from marine animals, with enzymatic hydrolysis being the most used. However, the emerging techniques have not been well used. Hence, there is a need to explore the emerging innovative technologies, such as pulsed electric field, ohmic heating, and high hydrostatic pressure processing, to produce peptides with significant bioactivities against human diseases. Even though several studies have reported the bioactivities of marine-based purified peptides, more research is needed to explore their full potential for food, pharmaceutical, and cosmetic industries. While interesting studies on the use of marine animal-based antioxidant and anticancer peptides have been reported in vitro, their bioavailability, functionality, shelf life, and long-term stability need to be addressed through in vivo studies for further use and large-scale production. Furthermore, to use these peptides for human consumption as nutraceuticals, bio-functional foods, or natural additives in food and healthcare products and in biopharmaceuticals, their mechanism of action should be determined through clinical studies to better understand their physiological properties.
Author Contributions
Conceptualization, K.M.I.B.; methodology, S.C. and K.M.I.B.; investigation, S.C. and K.M.I.B.; resources, A.W. and M.-G.C.; data curation, S.C. and K.M.I.B.; writing—original draft preparation, S.C. and K.M.I.B.; writing—review and editing, A.W., M.-G.C. and K.M.I.B.; supervision, M.-G.C. and K.M.I.B.; funding acquisition, M.-G.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the BB21plus (grant number: 2021-0103) funded by Busan Metropolitan City and Busan Institute for Talent & Lifelong Education (BIT); the authors are thankful for their support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Villamil, O.; Váquiro, H.; Solanilla, J.F. Fish viscera protein hydrolysates: Production, potential applications and functional and bioactive properties. Food Chem. 2017, 224, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Halim, N.R.A.; Yusof, H.M.; Sarbon, N.M. Functional and bioactive properties of fish protein hydolysates and peptides: A comprehensive review. Trends Food Sci. Technol. 2016, 51, 24–33. [Google Scholar] [CrossRef]
- Bashir, K.M.I.; Sohn, J.H.; Kim, J.-S.; Choi, J.-S. Identification and characterization of novel antioxidant peptides from mackerel (Scomber japonicus) muscle protein hydrolysates. Food Chem. 2020, 323, 126809. [Google Scholar] [CrossRef]
- Gao, M.-T.; Hirata, M.; Toorisaka, E.; Hano, T. Acid-hydrolysis of fish wastes for lactic acid fermentation. Bioresour. Technol. 2006, 97, 2414–2420. [Google Scholar] [CrossRef]
- Bashir, K.M.I.; Park, Y.-J.; An, J.H.; Choi, S.-J.; Kim, J.-H.; Baek, M.-K.; Kim, A.; Sohn, J.H.; Choi, J.-S. Antioxidant properties of Scomber japonicus hydrolysates prepared by enzymatic hydrolysis. J. Aquat. Food Prod. Technol. 2018, 27, 107–121. [Google Scholar] [CrossRef]
- Choi, J.-I.; Kim, J.-H.; Lee, J.-W. Physiological properties of tuna cooking drip hydrolysate prepared with gamma irradiation. Process Biochem. Process Biochem. 2011, 46, 1875–1878. [Google Scholar] [CrossRef]
- Ahmed, R.; Chun, B.-S. Subcritical water hydrolysis for the production of bioactive peptides from tuna skin collagen. J. Supercrit. Fluids 2018, 141, 88–96. [Google Scholar] [CrossRef]
- Wang, W.; Li, Z.; Liu, J.; Wang, Y.; Liu, S.; Sun, M. Comparison between thermal hydrolysis and enzymatic proteolysis processes for the preparation of tilapia skin. Czech J. Food Sci. 2013, 31, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Prabha, J.; Narikimelli, A.; Sajini, M.I.; Vincent, S. Optimization for autolysis assisted production of fish protein hydrolysate from underutilized fish Pellona ditchela. Int. J. Sci. Eng. Res. 2014, 4, 1863. [Google Scholar] [CrossRef] [Green Version]
- Jemil, I.; Jridi, M.; Nasri, R.; Ktari, N.; Ben Slama-Ben Salem, R.; Mehiri, M.; Hajji, M.; Nasri, M. Functional, antioxidant and antibacterial properties of protein hydrolysates prepared from fish meat fermented by Bacillus subtilis A26. Process Biochem. 2014, 49, 963–972. [Google Scholar] [CrossRef]
- Ozuna, C.; León-Galván, M.F. Cucurbitaceae seed protein hydrolysates as a potential source of bioactive peptides with functional properties. BioMed Res. Int. 2017, 2017, 2121878. [Google Scholar] [CrossRef]
- Lee, J.K.; Byun, H.-G. Characterization of antioxidative peptide purified from black eelpout (Lycodes diapterus) hydrolysate. Fish. Aquat. Sci. 2019, 22, 22. [Google Scholar] [CrossRef]
- Spellman, D.; McEvoy, E.; O’Cuinn, G.; FitzGerald, R.J. Proteinase and exopeptidase hydrolysis of whey protein: Comparison of the TNBS, OPA and pH stat methods for quantification of degree of hydrolysis. Int. Dairy J. 2003, 13, 447–453. [Google Scholar] [CrossRef]
- Kristinsson, H.G.; Rasco, B.A. Biochemical and functional properties of Atlantic salmon (Salmo salar) muscle proteins hydrolyzed with various alkaline proteases. J. Agric. Food Chem. 2000, 48, 657–666. [Google Scholar] [CrossRef]
- Fraga, C.G.; Croft, K.D.; Kennedy, D.O.; Tomás-Barberán, F.A. The effects of polyphenols and other bioactives on human health. Food Funct. 2019, 10, 514–528. [Google Scholar] [CrossRef] [Green Version]
- Rydlewski, A.A.; de Morais, D.R.; Rotta, E.M.; Claus, T.; Vagula, J.M.; da Silva, M.C.; Santos Junior, O.O.; Visentainer, J.V. Bioactive compounds, antioxidant capacity, and fatty acids in different parts of four unexplored fruits. J. Food Qual. 2017, 2017, 8401074. [Google Scholar] [CrossRef] [Green Version]
- Wong, J.H.; Ng, T.B.; Wang, H.; Sze, S.C.; Zhang, K.Y.; Li, Q.; Lu, X. Cordymin, an antifungal peptide from the medicinal fungus Cordyceps militaris. Phytomedicine Int. J. Phytother. Phytopharm. 2011, 18, 387–392. [Google Scholar] [CrossRef]
- Semreen, M.H.; El-Gamal, M.I.; Abdin, S.; Alkhazraji, H.; Kamal, L.; Hammad, S.; El-Awady, F.; Waleed, D.; Kourbaj, L. Recent updates of marine antimicrobial peptides. Saudi Pharm. J. 2018, 26, 396–409. [Google Scholar] [CrossRef]
- Landi, N.; Clemente, A.; Pedone, P.V.; Ragucci, S.; Di Maro, A. An updated review of bioactive peptides from mushrooms in a well-defined molecular weight range. Toxins 2022, 14, 84. [Google Scholar] [CrossRef]
- Gates, K.W. Bioactive compounds from marine foods: Plant and animal sources, by Blanca Hernández-Ledesma and Miguel Herrero (Editors). J. Aquat. Food Prod. Technol. 2014, 23, 313–317. [Google Scholar] [CrossRef]
- Winata, A.; Lorenz, K. Antioxidant potential of 5-N-pentadecylresorcinol. J. Food Processing Preserv. 2007, 20, 417–429. [Google Scholar] [CrossRef]
- Thiansilakul, Y.; Benjakul, S.; Shahidi, F. Compositions, functional properties and antioxidative activity of protein hydrolysates prepared from round scad (Decapterus maruadsi). Food Chem. 2007, 103, 1385–1394. [Google Scholar] [CrossRef]
- Saiga, A.; Okumura, T.; Makihara, T.; Katsuta, S.; Shimizu, T.; Yamada, R.; Nishimura, T. Angiotensin I-converting enzyme inhibitory peptides in a hydrolyzed chicken breast muscle extract. J. Agric. Food Chem. 2003, 51, 1741–1745. [Google Scholar] [CrossRef]
- Li, B. Isolation and identification of antioxidative peptides from porcine collagen hydrolysate by consecutive chromatography and electrospray ionization-mass spectrometry. Food Chem. 2007, 102, 1135–1143. [Google Scholar] [CrossRef]
- Gómez-Guillén, M.C.; Giménez, B.; López-Caballero, M.E.; Montero, M.P. Functional and bioactive properties of collagen and gelatin from alternative sources: A review. Food Hydrocoll. 2011, 25, 1813–1827. [Google Scholar] [CrossRef] [Green Version]
- Trampuz, A.; Prabhu, R.M.; Smith, T.F.; Baddour, L.M. Avian Influenza: A new pandemic threat? Mayo Clin. Proc. 2004, 79, 523–530. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.-L.; Jao, C.-L.; Ho, K.-P.; Hsu, K.-C. Dipeptidyl-peptidase IV inhibitory activity of peptides derived from tuna cooking juice hydrolysates. Peptides 2012, 35, 114–121. [Google Scholar] [CrossRef]
- Harnedy, P.A.; FitzGerald, R.J. Bioactive peptides from marine processing waste and shellfish: A review. J. Funct. Foods 2012, 4, 6–24. [Google Scholar] [CrossRef]
- Bashir, K.M.I.; Mohibbullah, M.; An, J.H.; Choi, J.-Y.; Hong, Y.-K.; Sohn, J.H.; Kim, J.-S.; Choi, J.-S. In vivo antioxidant activity of mackerel (Scomber japonicus) muscle protein hydrolysate. PeerJ 2018, 6, e6181. [Google Scholar] [CrossRef] [Green Version]
- Bkhairia, I.; Kolsi, R.B.A.; Ghorbel, S.; Azzabou, S.; Ktari, N.; Nasri. Anti-inflammatory, antioxidant activities and fatty acid profile of three hydrolysates from Liza aurata by-product influenced by hydrolysis degree. Adv. Tech. Biol. Med. 2019, 7, 2379-1764. [Google Scholar]
- Ishak, N.H.; Sarbon, N.M. A review of protein hydrolysates and bioactive peptides deriving from wastes generated by fish processing. Food Bioprocess Technol. 2018, 11, 2–16. [Google Scholar] [CrossRef]
- Wang, S.-H.; Yu, J. Structure-based design for binding peptides in anti-cancer therapy. Biomaterials 2018, 156, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lungu, I.I.; Grumezescu, A.M.; Volceanov, A.; Andronescu, E. Nanobiomaterials used in cancer therapy: An up-to-date overview. Molecules 2019, 24, 3547. [Google Scholar] [CrossRef] [Green Version]
- Nardo, A.E.; Suárez, S.; Quiroga, A.V.; Añón, M.C. Amaranth as a source of antihypertensive peptides. Front. Plant Sci. 2020, 11, 578631. [Google Scholar] [CrossRef]
- Wijesekara, I.; Kim, S.-K. Angiotensin-I-converting enzyme (ACE) inhibitors from marine resources: Prospects in the pharmaceutical industry. Mar. Drugs 2010, 8, 1080–1093. [Google Scholar] [CrossRef] [Green Version]
- Li, G.H.; Qu, M.R.; Wan, J.Z.; You, J.M. Antihypertensive effect of rice protein hydrolysate with in vitro angiotensin I-converting enzyme inhibitory activity in spontaneously hypertensive rats. Asia Pac. J. Clin. Nutr. 2007, 16 (Suppl. S1), 275–280. [Google Scholar]
- Jäkälä, P.; Vapaatalo, H. Antihypertensive peptides from milk proteins. Pharmaceuticals 2010, 3, 251–272. [Google Scholar] [CrossRef] [Green Version]
- Ngo, D.-H.; Vo, T.-S.; Ryu, B.; Kim, S.-K. Angiotensin- I- converting enzyme (ACE) inhibitory peptides from Pacific cod skin gelatin using ultrafiltration membranes. Process Biochem. 2016, 51, 1622–1628. [Google Scholar] [CrossRef] [Green Version]
- Yathisha, U.G.; Bhat, I.; Karunasagar, I.; Mamatha, B.S. Antihypertensive activity of fish protein hydrolysates and its peptides. Crit. Rev. Food Sci. Nutr. 2019, 59, 2363–2374. [Google Scholar] [CrossRef]
- Rahmdel, M.; Cho, S.M.; Jeon, Y.J.; Lee, D.H. A flounder fish peptide shows anti-hypertensive effects by suppressing the renin-angiotensin-aldosterone system and Endothelin-1. Protein Pept. Lett. 2021, 28, 831–840. [Google Scholar] [CrossRef]
- Cai, S.; Pan, N.; Xu, M.; Su, Y.; Qiao, K.; Chen, B.; Zheng, B.; Xiao, M.; Liu, Z. ACE inhibitory peptide from skin collagen hydrolysate of Takifugu bimaculatus as potential for protecting HUVECs injury. Mar. Drugs 2021, 19, 655. [Google Scholar] [CrossRef]
- Bartlett, T.C.; Cuthbertson, B.J.; Shepard, E.F.; Chapman, R.W.; Gross, P.S.; Warr, G.W. Crustins, homologues of an 11.5-kDa antibacterial peptide, from two species of penaeid shrimp, Litopenaeus vannamei and Litopenaeus setiferus. Mar. Biotechnol. 2002, 4, 278–293. [Google Scholar] [CrossRef]
- Battison, A.L.; Summerfield, R.; Patrzykat, A. Isolation and characterisation of two antimicrobial peptides from haemocytes of the American lobster Homarus americanus. Fish Shellfish. Immunol. 2008, 25, 181–187. [Google Scholar] [CrossRef]
- Dat, T.T.; Cuc, N.T.; Cuong, P.V.; Smidt, H.; Sipkema, D. Diversity and antimicrobial activity of vietnamese sponge-associated bacteria. Mar. Drugs 2021, 19, 353. [Google Scholar] [CrossRef]
- Li, C.; Haug, T.; Styrvold, O.B.; Jørgensen, T.; Stensvåg, K. Strongylocins, novel antimicrobial peptides from the green sea urchin, Strongylocentrotus droebachiensis. Dev. Comp. Immunol. 2008, 32, 1430–1440. [Google Scholar] [CrossRef]
- Erdem Büyükkiraz, M.; Kesmen, Z. Antimicrobial peptides (AMPs): A promising class of antimicrobial compounds. J. Appl. Microbiol. 2022, 132, 1573–1596. [Google Scholar] [CrossRef]
- Liu, Z.; Dong, S.; Xu, J.; Zeng, M.; Song, H.; Zhao, Y. Production of cysteine-rich antimicrobial peptide by digestion of oyster (Crassostrea gigas) with alcalase and bromelin. Food Control 2008, 19, 231–235. [Google Scholar] [CrossRef]
- Falanga, A.; Lombardi, L.; Franci, G.; Vitiello, M.; Iovene, M.R.; Morelli, G.; Galdiero, M.; Galdiero, S. Marine Antimicrobial peptides: Nature provides templates for the design of novel compounds against pathogenic bacteria. Int. J. Mol. Sci. 2016, 17, 785. [Google Scholar] [CrossRef] [Green Version]
- Fosgerau, K.; Hoffmann, T. Peptide therapeutics: Current status and future directions. Drug Discov. Today 2015, 20, 122–128. [Google Scholar] [CrossRef] [Green Version]
- El-Salam, M.H.A.; El-Shibiny, S. Bioactive peptides of buffalo, camel, goat, sheep, mare, and yak milks and milk products. Food Rev. Int. 2013, 29, 1–23. [Google Scholar] [CrossRef]
- Kang, H.K.; Lee, H.H.; Seo, C.H.; Park, Y. Antimicrobial and immunomodulatory properties and applications of marine-derived proteins and peptides. Mar. Drugs 2019, 17, 350. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; He, K.; Dong, X.; Zhang, Z.; Wang, F.; Tang, Y.; Chen, Y.; Ding, G. Immunomodulatory activity of low molecular-weight peptides from Nibea japonica skin in cyclophosphamide-induced immunosuppressed mice. J. Funct. Foods 2020, 68, 103888. [Google Scholar] [CrossRef]
- Parkin, D.M. Global cancer statistics in the year 2000. Lancet Oncol. 2001, 2, 533–543. [Google Scholar] [CrossRef]
- Schirrmacher, V. From chemotherapy to biological therapy: A review of novel concepts to reduce the side effects of systemic cancer treatment. Int. J. Oncol. 2019, 54, 407–419. [Google Scholar] [CrossRef]
- Alves, C.; Silva, J.; Pinteus, S.; Gaspar, H.; Alpoim, M.C.; Botana, L.M.; Pedrosa, R. From marine origin to therapeutics: The antitumor potential of marine algae-derived compounds. Front. Pharmacol. Ethnopharmacol. 2018, 9, 777. [Google Scholar] [CrossRef] [Green Version]
- Felício, M.R.; Silva, O.N.; Gonçalves, S.; Santos, N.C.; Franco, O.L. Peptides with dual antimicrobial and anticancer activities. Front. Chem. 2017, 5, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalifa, S.A.M.; Elias, N.; Farag, M.A.; Chen, L.; Saeed, A.; Hegazy, M.-E.F.; Moustafa, M.S.; Abd El-Wahed, A.; Al-Mousawi, S.M.; Musharraf, S.G.; et al. Marine natural products: A source of novel anticancer drugs. Mar. Drugs 2019, 17, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Z.; Li, M.; Dey, R.; Chen, Y. Nanomaterials for cancer therapy: Current progress and perspectives. J. Hematol. Oncol. 2021, 14, 85. [Google Scholar] [CrossRef] [PubMed]
- Negi, B.; Kumar, D.; Rawat, D.S. Marine peptides as anticancer agents: A remedy to mankind by nature. Curr. Protein Peptide Sci. 2017, 18, 885–904. [Google Scholar] [CrossRef] [PubMed]
- Ettelaie, R.; Zengin, A.; Lishchuk, S.V. Novel food grade dispersants: Review of recent progress. Curr. Opin. Colloid Interface Sci. 2017, 28, 46–55. [Google Scholar] [CrossRef] [Green Version]
- Lafarga, T.; Hayes, M. Bioactive protein hydrolysates in the functional food ingredient industry: Overcoming current challenges. Food Rev. Int. 2017, 33, 217–246. [Google Scholar] [CrossRef]
- Le Gouic, A.V.; Harnedy, P.A.; FitzGerald, R.J. Bioactive Peptides from Fish Protein By-Products. In Bioactive Molecules in Food; Mérillon, J.-M., Ramawat, K.G., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–35. [Google Scholar]
- Nicco, C.; Batteux, F. ROS modulator molecules with therapeutic potential in cancers treatments. Molecules 2017, 23, 84. [Google Scholar] [CrossRef] [Green Version]
- Barrow, C.; Shahidi, F. Marine Nutraceuticals and Functional Foods, 1st ed.; CRC Press: Boca Raton, FL, USA, 2007; p. 512. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative stress, and antioxidants: Back and forth in the pathophysiology of chronic diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN Estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Pihlanto-Leppälä, A. Bioactive peptides derived from bovine whey proteins: Opioid and ace-inhibitory peptides. Trends Food Sci. Technol. 2000, 11, 347–356. [Google Scholar] [CrossRef]
- Ratnam, D.V.; Ankola, D.D.; Bhardwaj, V.; Sahana, D.K.; Kumar, M.N. Role of antioxidants in prophylaxis and therapy: A pharmaceutical perspective. J. Control. Release Off. J. Control. Release Soc. 2006, 113, 189–207. [Google Scholar] [CrossRef]
- Bouayed, J.; Bohn, T. Exogenous antioxidants—Double-edged swords in cellular redox state: Health beneficial effects at physiologic doses versus deleterious effects at high doses. Oxid. Med. Cell. Longev. 2010, 3, 228–237. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Jun, S.-Y.; Park, P.-J.; Jung, W.-K.; Kim, S.-K. Purification and characterization of an antioxidative peptide from enzymatic hydrolysate of yellowfin sole (Limanda aspera). Eur. Food Res. Technol. 2004, 219, 20–26. [Google Scholar] [CrossRef]
- Je, J.-Y.; Zhong-Ji, Q.; Byun, H.-G.; Kim, S.-K. Purification and characterization of an antioxidant peptide obtained from tuna backbone protein by enzymatic hydrolysis. Process Biochem. 2007, 42, 840–846. [Google Scholar] [CrossRef]
- Tao, J.; Zhao, Y.Q.; Chi, C.F.; Wang, B. Bioactive peptides from cartilage protein hydrolysate of spotless smoothhound and their antioxidant activity in vitro. Mar. Drugs 2018, 16, 100. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; He, J.; Zhuang, Y.; Sun, L. Purification and identification of antioxidant peptides from enzymatic hydrolysates of Tilapia (Oreochromis niloticus) frame protein. Molecules 2012, 17, 12836–12850. [Google Scholar] [CrossRef]
- Zhang, Y.; Duan, X.; Zhuang, Y. Purification and characterization of novel antioxidant peptides from enzymatic hydrolysates of tilapia (Oreochromis niloticus) skin gelatin. Peptides 2012, 38, 13–21. [Google Scholar] [CrossRef]
- Ren, Y.; Wu, H.; Chi, Y.; Deng, R.; He, Q. Structural characterization, erythrocyte protection, and antifatigue effect of antioxidant collagen peptides from tilapia (Oreochromis nilotica L.) skin. Food Funct. 2020, 11, 10149–10160. [Google Scholar] [CrossRef]
- Shiao, W.-C.; Wu, T.-C.; Kuo, C.-H.; Tsai, Y.-H.; Tsai, M.-L.; Hong, Y.-H.; Huang, C.-Y. Physicochemical and antioxidant properties of gelatin and gelatin hydrolysates obtained from extrusion-pretreated fish (Oreochromis sp.) scales. Mar. Drugs 2021, 19, 275. [Google Scholar] [CrossRef]
- Ahn, C.-B.; Kim, J.-G.; Je, J.-Y. Purification and antioxidant properties of octapeptide from salmon byproduct protein hydrolysate by gastrointestinal digestion. Food Chem. 2014, 147, 78–83. [Google Scholar] [CrossRef]
- Ko, J.Y.; Lee, J.H.; Samarakoon, K.; Kim, J.S.; Jeon, Y.J. Purification and determination of two novel antioxidant peptides from flounder fish (Paralichthys olivaceus) using digestive proteases. Food Chem. Toxicol. 2013, 52, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Cheng, Q.; Peng, Q.; Yu, X.; Yin, X.; Liang, M.; Ma, C.W.; Huang, Z.; Jia, W. Antioxidant peptides derived from the hydrolyzate of purple sea urchin (Strongylocentrotus nudus) gonad alleviate oxidative stress in Caenorhabditis elegans. J. Funct. Foods 2018, 48, 594–604. [Google Scholar] [CrossRef]
- Pan, X.; Zhao, Y.-Q.; Hu, X.-Y.; Wang, B. Preparation and identification of antioxidant peptides from protein hydrolysate of skate (Raja porosa) cartilage. J. Funct. Foods 2016, 25, 220–230. [Google Scholar] [CrossRef]
- Sampath Kumar, N.S.; Nazeer, R.A.; Jaiganesh, R. Purification and identification of antioxidant peptides from the skin protein hydrolysate of two marine fishes, horse mackerel (Magalaspis cordyla) and croaker (Otolithes ruber). Amino Acids 2012, 42, 1641–1649. [Google Scholar] [CrossRef]
- Wang, W.Y.; Zhao, Y.Q.; Zhao, G.X.; Chi, C.F.; Wang, B. Antioxidant peptides from collagen hydrolysate of redlip croaker (Pseudosciaena polyactis) scales: Preparation, characterization, and cytoprotective effects on H2O2-damaged HepG2 cells. Mar. Drugs 2020, 18, 156. [Google Scholar] [CrossRef] [Green Version]
- Li, X.R.; Chi, C.F.; Li, L.; Wang, B. Purification and identification of antioxidant peptides from protein hydrolysate of scalloped hammerhead (Sphyrna lewini) cartilage. Mar. Drugs 2017, 15, 61. [Google Scholar] [CrossRef]
- Rajapakse, N.; Mendis, E.; Byun, H.-G.; Kim, S.-K. Purification and in vitro antioxidative effects of giant squid muscle peptides on free radical-mediated oxidative systems. J. Nutr. Biochem. 2005, 16, 562–569. [Google Scholar] [CrossRef]
- Umayaparvathi, S.; Meenakshi, S.; Vimalraj, V.; Arumugam, M.; Sivagami, G.; Balasubramanian, T. Antioxidant activity and anticancer effect of bioactive peptide from enzymatic hydrolysate of oyster (Saccostrea cucullata). Biomed. Prev. Nutr. 2014, 4, 343–353. [Google Scholar] [CrossRef]
- Je, J.-Y.; Park, P.-J.; Kim, S.-K. Antioxidant activity of a peptide isolated from Alaska pollack (Theragra chalcogramma) frame protein hydrolysate. Food Res. Int. 2005, 38, 45–50. [Google Scholar] [CrossRef]
- Kim, S.-K.; Kim, Y.-T.; Byun, H.-G.; Nam, K.-S.; Joo, D.-S.; Shahidi, F. Isolation and characterization of antioxidative peptides from gelatin hydrolysate of Alaska pollack skin. J. Agric. Food Chem. 2001, 49, 1984–1989. [Google Scholar] [CrossRef]
- Mendis, E.; Rajapakse, N.; Kim, S.-K. Antioxidant properties of a radical-scavenging peptide purified from enzymatically prepared fish skin gelatin hydrolysate. J. Agric. Food Chem. 2005, 53, 581–587. [Google Scholar] [CrossRef]
- Kim, S.Y.; Je, J.Y.; Kim, S.K. Purification and characterization of antioxidant peptide from hoki (Johnius belengerii) frame protein by gastrointestinal digestion. J. Nutr. Biochem. 2007, 18, 31–38. [Google Scholar] [CrossRef]
- Zhao, G.X.; Yang, X.R.; Wang, Y.M.; Zhao, Y.Q.; Chi, C.F.; Wang, B. Antioxidant peptides from the protein hydrolysate of spanish mackerel (Scomberomorous niphonius) muscle by in vitro gastrointestinal digestion and their in vitro activities. Mar. Drugs 2019, 17, 531. [Google Scholar] [CrossRef] [Green Version]
- Sampath Kumar, N.S.; Nazeer, R.A.; Jaiganesh, R. Purification and biochemical characterization of antioxidant peptide from horse mackerel (Magalaspis cordyla) viscera protein. Peptides 2011, 32, 1496–1501. [Google Scholar] [CrossRef]
- Asaduzzaman, A.K.M.; Getachew, A.T.; Cho, Y.-J.; Park, J.-S.; Haq, M.; Chun, B.-S. Characterization of pepsin-solubilised collagen recovered from mackerel (Scomber japonicus) bone and skin using subcritical water hydrolysis. Int. J. Biol. Macromol. 2020, 148, 1290–1297. [Google Scholar] [CrossRef]
- Kim, N.Y.; Jung, H.Y.; Kim, J.K. Identification and characterisation of a novel heptapeptide mackerel by-product hydrolysate, and its potential as a functional fertiliser component. J. Chromatogr. B 2021, 1180, 122881. [Google Scholar] [CrossRef]
- Wang, Y.-Z.; Wang, Y.-M.; Pan, X.; Chi, C.-F.; Wang, B. Antioxidant mechanisms of the oligopeptides (FWKVV and FMPLH) from muscle hydrolysate of miiuy croaker against oxidative damage of HUVECs. Oxid. Med. Cell. Longev. 2021, 2021, 9987844. [Google Scholar] [CrossRef]
- Naqash, S.Y.; Nazeer, R.A. Antioxidant activity of hydrolysates and peptide fractions of Nemipterus japonicus and Exocoetus volitans muscle. J. Aquat. Food Prod. Technol. 2010, 19, 180–192. [Google Scholar] [CrossRef]
- Wang, K.; Han, L.; Hong, H.; Pan, J.; Liu, H.; Luo, Y. Purification and identification of novel antioxidant peptides from silver carp muscle hydrolysate after simulated gastrointestinal digestion and transepithelial transport. Food Chem. 2021, 342, 128275. [Google Scholar] [CrossRef]
- Chi, C.-F.; Wang, B.; Hu, F.-Y.; Wang, Y.-M.; Zhang, B.; Deng, S.-G.; Wu, C.-W. Purification and identification of three novel antioxidant peptides from protein hydrolysate of bluefin leatherjacket (Navodon septentrionalis) skin. Food Res. Int. 2015, 73, 124–129. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, J.; Tang, Y.; Zhang, Z.; Zhang, Y.; Hu, J. Purification and characterization of antioxidant peptides from hairtail surimi hydrolysates and their effects on beef color stability. J. Food Sci. 2021, 86, 2898–2909. [Google Scholar] [CrossRef]
- Gui, M.; Gao, L.; Rao, L.; Li, P.; Zhang, Y.; Han, J.W.; Li, J. Bioactive peptides identified from enzymatic hydrolysates of sturgeon skin. J. Sci. Food Agric. 2022, 102, 1948–1957. [Google Scholar] [CrossRef]
- Hu, X.; Yang, X.; Wang, T.; Li, L.; Wu, Y.; Zhou, Y.; You, L. Purification and identification of antioxidant peptides from round scad (Decapterus maruadsi) hydrolysates by consecutive chromatography and electrospray ionization-mass spectrometry. Food Chem. Toxicol. 2020, 135, 110882. [Google Scholar] [CrossRef]
- Hu, X.-M.; Wang, Y.-M.; Zhao, Y.-Q.; Chi, C.-F.; Wang, B. Antioxidant peptides from the protein hydrolysate of monkfish (Lophius litulon) muscle: Purification, identification, and cytoprotective function on HepG2 cells damage by H2O2. Mar. Drugs 2020, 18, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, H.L.; Shin, S.R.; Yoon, K.Y. Isolation of antioxidant peptide from sandfish (Arctoscopus japonicus) roe hydrolysate. Korean J. Food Preserv. 2017, 24, 542–549. [Google Scholar] [CrossRef]
- Kula, E.; Kocadag Kocazorbaz, E.; Moulahoum, H.; Alpat, S.; Zihnioglu, F. Extraction and characterization of novel multifunctional peptides from Trachinus draco (greater weever) myofibrillar proteins with ACE/DPP4 inhibitory, antioxidant, and metal chelating activities. J. Food Biochem. 2020, 44, e13179. [Google Scholar] [CrossRef] [PubMed]
- Guru, A.; Lite, C.; Freddy, A.J.; Issac, P.K.; Pasupuleti, M.; Saraswathi, N.T.; Arasu, M.V.; Al-Dhabi, N.A.; Arshad, A.; Arockiaraj, J. Intracellular ROS scavenging and antioxidant regulation of WL15 from cysteine and glycine-rich protein 2 demonstrated in zebrafish in vivo model. Dev. Comp. Immunol. 2021, 114, 103863. [Google Scholar] [CrossRef]
- Guru, A.; Issac, P.K.; Saraswathi, N.T.; Seshadri, V.D.; Gabr, G.A.; Arockiaraj, J. Deteriorating insulin resistance due to WL15 peptide from cysteine and glycine-rich protein 2 in high glucose-induced rat skeletal muscle L6 cells. Cell Biol. Int. 2021, 45, 1698–1709. [Google Scholar] [CrossRef]
- Himaya, S.W.; Ryu, B.; Ngo, D.H.; Kim, S.K. Peptide isolated from Japanese flounder skin gelatin protects against cellular oxidative damage. J. Agric. Food Chem. 2012, 60, 9112–9119. [Google Scholar] [CrossRef]
- Hu, Z.; Sha, X.; Zhang, L.; Huang, S.; Tu, Z. Effect of grass carp scale collagen peptide FTGML on cAMP-PI3K/Akt and MAPK signaling pathways in B16F10 melanoma cells and correlation between anti-melanin and antioxidant properties. Foods 2022, 11, 391. [Google Scholar] [CrossRef]
- Nazeer, R.A.; Kumar, N.S.; Jai Ganesh, R. In vitro and in vivo studies on the antioxidant activity of fish peptide isolated from the croaker (Otolithes ruber) muscle protein hydrolysate. Peptides 2012, 35, 261–268. [Google Scholar] [CrossRef]
- Kumar, N.; Rasool Abdul, N.; Jaiganesh, R. In vivo antioxidant activity of peptide purified from viscera protein hydrolysate of horse mackerel (Magalaspis cordyla). Int. J. Food Sci. Technol. 2012, 47, 1558–1562. [Google Scholar] [CrossRef]
- Wei, L.; Ji, H.; Song, W.; Peng, S.; Zhan, S.; Qu, Y.; Chen, M.; Zhang, D.; Liu, S. Hypouricemic, hepatoprotective and nephroprotective roles of oligopeptides derived from Auxis thazard protein in hyperuricemic mice. Food Funct. 2021, 12, 11838–11848. [Google Scholar] [CrossRef]
- Hu, J.; Liu, R.; Yu, X.; Li, Z.; Liu, X.; Hao, Y.; Zhu, N.; Kang, J.; Li, Y. Protective effects of small-molecule oligopeptides isolated from tilapia fish scale on ethanol-induced gastroduodenal injury in rats. Nutrients 2021, 13, 2078. [Google Scholar] [CrossRef]
- Kim, D.U.; Chung, H.C.; Choi, J.; Sakai, Y.; Lee, B.Y. Oral intake of low-molecular-weight collagen peptide improves hydration, elasticity, and wrinkling in human skin: A randomized, double-blind, placebo-controlled study. Nutrients 2018, 10, 826. [Google Scholar] [CrossRef] [Green Version]
- Czajka, A.; Kania, E.M.; Genovese, L.; Corbo, A.; Merone, G.; Luci, C.; Sibilla, S. Daily oral supplementation with collagen peptides combined with vitamins and other bioactive compounds improves skin elasticity and has a beneficial effect on joint and general wellbeing. Nutr. Res. 2018, 57, 97–108. [Google Scholar] [CrossRef]
- De Luca, C.; Mikhal’chik, E.V.; Suprun, M.V.; Papacharalambous, M.; Truhanov, A.I.; Korkina, L.G. Skin antiageing and systemic redox effects of supplementation with marine collagen peptides and plant-derived antioxidants: A single-blind case-control clinical study. Oxid. Med. Cell. Longev. 2016, 2016, 4389410. [Google Scholar] [CrossRef] [Green Version]
- Tapal, A.; Tiku, P.K. Chapter 27—Nutritional and Nutraceutical Improvement by Enzymatic Modification of Food Proteins. In Enzymes in Food Biotechnology; Kuddus, M., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 471–481. [Google Scholar]
- Laroque, D.; Chabeaud, A.; Guérard, F. Antioxidant capacity of marine protein hydrolysates. In Added value to Fisheries Waste; Bergé, J.P., Ed.; Transworld Research Network: Kerala, India, 2008; pp. 147–161. [Google Scholar]
- Karami, Z.; Akbari-Adergani, B. Bioactive food derived peptides: A review on correlation between structure of bioactive peptides and their functional properties. J. Food Sci. Technol. 2019, 56, 535–547. [Google Scholar] [CrossRef]
- Pan, X.; Zhao, Y.Q.; Hu, F.Y.; Chi, C.F.; Wang, B. Anticancer activity of a hexapeptide from skate (Raja porosa) Cartilage protein hydrolysate in HeLa cells. Mar. Drugs 2016, 14, 153. [Google Scholar] [CrossRef] [Green Version]
- Kawashima, K.; Itoh, H.; Miyoshi, M.; Chibata, I. Antioxidant properties of branched-chain amino acid derivatives. Chem. Pharm. Bull. 1979, 27, 1912–1916. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.K.; Choi, M.-C.; Seo, C.H.; Park, Y. Therapeutic properties and biological benefits of marine-derived anticancer peptides. Int. J. Mol. Sci. 2018, 19, 919. [Google Scholar] [CrossRef] [Green Version]
- Hsu, K.-C.; Li-Chan, E.C.Y.; Jao, C.-L. Antiproliferative activity of peptides prepared from enzymatic hydrolysates of tuna dark muscle on human breast cancer cell line MCF-7. Food Chem. 2011, 126, 617–622. [Google Scholar] [CrossRef]
- Lee, Y.G.; Lee, K.W.; Kim, J.Y.; Kim, K.H.; Lee, H.J. Induction of apoptosis in a human lymphoma cell line by hydrophobic peptide fraction separated from anchovy sauce. Biofactors 2004, 21, 63–67. [Google Scholar] [CrossRef]
- Kim, E.K.; Joung, H.J.; Kim, Y.S.; Hwang, J.W.; Ahn, C.B.; Jeon, Y.J.; Moon, S.H.; Park, P.J. Purification of a novel anticancer peptide from enzymatic hydrolysate of Mytilus coruscus. J. Microbiol. Biotechnol. 2012, 22, 1381–1387. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.P.; Huang, T.C.; Lin, C.C.; Hui, C.F.; Lin, C.H.; Chen, J.Y. Pardaxin, a fish antimicrobial peptide, exhibits antitumor activity toward murine fibrosarcoma in vitro and in vivo. Mar. Drugs 2012, 10, 1852–1872. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.; Jing, Y.; Ding, G.; Yang, Z. Isolation and purification of novel peptides derived from Sepia ink: Effects on apoptosis of prostate cancer cell PC-3. Mol. Med. Rep. 2017, 16, 4222–4228. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Xie, B.; Su, Y.; Gu, Q.; Hao, D.; Liu, H.; Wang, C.; Hu, Y.; Zhang, M. Expression analysis of tissue factor pathway inhibitors TFPI-1 and TFPI-2 in Paralichthys olivaceus and antibacterial and anticancer activity of derived peptides. Vet. Res. 2021, 52, 32. [Google Scholar] [CrossRef]
- Yuan, E.; Nie, S.; Qi, C.; Chang, B.; Ren, J. Effect of Scomberomorus niphonius peptide on the characteristics of resveratrol. Food Funct. 2021, 12, 11449–11459. [Google Scholar] [CrossRef]
- Chang, W.T.; Pan, C.Y.; Rajanbabu, V.; Cheng, C.W.; Chen, J.Y. Tilapia (Oreochromis mossambicus) antimicrobial peptide, hepcidin 1-5, shows antitumor activity in cancer cells. Peptides 2011, 32, 342–352. [Google Scholar] [CrossRef]
- Suárez, Y.; González, L.; Cuadrado, A.; Berciano, M.; Lafarga, M.; Muñoz, A. Kahalalide F, a new marine-derived compound, induces oncosis in human prostate and breast cancer cells. Mol. Cancer Ther. 2003, 2, 863–872. [Google Scholar]
- Han, Y.; Cui, Z.; Li, Y.H.; Hsu, W.H.; Lee, B.H. In vitro and in vivo anticancer activity of Pardaxin against proliferation and growth of oral squamous cell carcinoma. Mar. Drugs 2015, 14, 2. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Rahman, M.A.; Abdel-Nabi, I.M.; El-Naggar, M.S.; Abbas, O.A.; Strong, P.N. Conus vexillum venom induces oxidative stress in Ehrlich’s ascites carcinoma cells: An insight into the mechanism of induction. J. Venom. Anim. Toxins Incl. Trop. Dis. 2013, 19, 10. [Google Scholar] [CrossRef] [Green Version]
- Pardo, B.; Paz-Ares, L.; Tabernero, J.; Ciruelos, E.; García, M.; Salazar, R.; López, A.; Blanco, M.; Nieto, A.; Jimeno, J.; et al. Phase I clinical and pharmacokinetic study of Kahalalide F administered weekly as a 1-hour infusion to patients with advanced solid tumors. Clin. Cancer Res. 2008, 14, 1116–1123. [Google Scholar] [CrossRef] [Green Version]
- Rademaker-Lakhai, J.M.; Horenblas, S.; Meinhardt, W.; Stokvis, E.; de Reijke, T.M.; Jimeno, J.M.; Lopez-Lazaro, L.; Lopez Martin, J.A.; Beijnen, J.H.; Schellens, J.H. Phase I clinical and pharmacokinetic study of Kahalalide F in patients with advanced androgen refractory prostate cancer. Clin. Cancer Res. 2005, 11, 1854–1862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín-Algarra, S.; Espinosa, E.; Rubió, J.; López López, J.J.; Manzano, J.L.; Carrión, L.A.; Plazaola, A.; Tanovic, A.; Paz-Ares, L. Phase II study of weekly Kahalalide F in patients with advanced malignant melanoma. Eur. J. Cancer 2009, 45, 732–735. [Google Scholar] [CrossRef] [PubMed]
- Madden, T.; Tran, H.T.; Beck, D.; Huie, R.; Newman, R.A.; Pusztai, L.; Wright, J.J.; Abbruzzese, J.L. Novel marine-derived anticancer agents: A phase I clinical, pharmacological, and pharmacodynamic study of dolastatin 10 (NSC 376128) in patients with advanced solid tumors. Clin. Cancer Res. 2000, 6, 1293–1301. [Google Scholar] [PubMed]
- Krug, L.M.; Miller, V.A.; Kalemkerian, G.P.; Kraut, M.J.; Ng, K.K.; Heelan, R.T.; Pizzo, B.A.; Perez, W.; McClean, N.; Kris, M.G. Phase II study of dolastatin-10 in patients with advanced non-small-cell lung cancer. Ann. Oncol. 2000, 11, 227–228. [Google Scholar] [CrossRef] [PubMed]
- Margolin, K.; Longmate, J.; Synold, T.W.; Gandara, D.R.; Weber, J.; Gonzalez, R.; Johansen, M.J.; Newman, R.; Baratta, T.; Doroshow, J.H. Dolastatin-10 in metastatic melanoma: A phase II and pharmokinetic trial of the California cancer consortium. Investig. New Drugs 2001, 19, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Hamann, M.T. Technology evaluation: Kahalalide, F. PharmaMar. Curr. Opin. Mol. Ther. 2004, 6, 657–665. [Google Scholar] [PubMed]
- Ciavatta, M.L.; Lefranc, F.; Carbone, M.; Mollo, E.; Gavagnin, M.; Betancourt, T.; Dasari, R.; Kornienko, A.; Kiss, R. Marine mollusk-derived agents with antiproliferative activity as promising anticancer agents to overcome chemotherapy resistance. Med. Res. Rev. 2017, 37, 702–801. [Google Scholar] [CrossRef]
- Pitot, H.C.; McElroy, E.A., Jr.; Reid, J.M.; Windebank, A.J.; Sloan, J.A.; Erlichman, C.; Bagniewski, P.G.; Walker, D.L.; Rubin, J.; Goldberg, R.M.; et al. Phase I trial of dolastatin-10 (NSC 376128) in patients with advanced solid tumors. Clin. Cancer Res. 1999, 5, 525–531. [Google Scholar]
Figure 1.
Current methods of producing marine animal-based protein hydrolysates and peptides.

Figure 2.
Schematic diagram of search strategy.


{kind=link}
{kind=link}
Table 1.
Antioxidant effects of bioactive peptides from marine animals—in vitro studies.
| Peptide Name/Sequence | Size of Peptide (Da) | Source | Cell Line | Analysis | Major Findings | Reference |
|---|---|---|---|---|---|---|
| RPNFDLEPPY | 13 K | Yellowfin sole (Limanda aspera) | - | TBARS assay, HPLC |
| [72] |
| VKAGFAWTANQQLS | 1519 | Tuna—backbone | MRC-5 and ECV304 | DPPH•, O2•−, and •OH scavenging assay, lipid peroxidation inhibition assay, FPLC, MTT assay, Q-TOF MS |
| [73] |
|
| Spotless smooth hound—cartilage(Mustelus griseus) | HepG2 | DPPH•, •OH, O2•−, and ABTS• scavenging activity, lipid peroxidation inhibition assay, MTT assay |
| [74] |
|
| Tilapia—frame (Oreochromis niloticus) | - | DPPH•, •OH, O2•−, and H2O2 scavenging activity |
| [75] |
|
| Tilapia—skin (Oreochromis niloticus) | - | DPPH•, •OH, and O2•− scavenging activity, Q-TOF MS, RP-HPLC |
| [76] |
| - | Tilapia—skin (Oreochromis niloticus) | Erythrocytes | DPPH•, •OH, and O2•− scavenging activity, HPLC, ESI-MS/MS, hemolysis assay |
| [77] |
|
| Tilapia—scales (Oreochromis niloticus) | - | DPPH• and ABTS• scavenging activity, FRAP assay, HPLC, LC/ESI-MS/MS |
| [78] |
| FLNEFLHV | 1018.48 | Salmon—pectoral fin | - | DPPH• and ABTS• scavenging activity, FRAP assay |
| [79] |
|
| Flounder fish (Paralichthys olivaceus) | Vero | DPPH• scavenging activity |
| [80] |
|
| Purple sea urchin—gonad (Strongylocentrotus nudus) | - | DPPH• scavenging activity |
| [81] |
|
| Skate—cartilage (Raja porosa) | - | DPPH•, •OH, O2•−, and ABTS• scavenging activity, lipid peroxidation inhibition assay |
| [82] |
| GNRGFACRHA | 1101.5 | Croaker—skin (Otolithes ruber) | - | DPPH• and •OH scavenging activity, lipid peroxidation inhibition assay, Fe2+ chelating activity, IEC, GPC, ESI-MS/MS |
| [83] |
|
| Redlip Croaker—scales (Pseudosciaena polyactis) | HepG2 | DPPH•, •OH, and O2•− scavenging activity, ROS levels, lipid peroxidation inhibition assay, IEC, gel filtration chromatography, RP-HPLC |
| [84] |
|
| Scalloped Hammer—cartilage (Sphyrna lewini) | - | DPPH•, ABTS•, •OH, and O2•− scavenging activity, lipid peroxidation inhibition assay, IEC, GPC, RP-HPLC, Q-TOF MS |
| [85] |
|
| Giant squid (Dosidicus gigas) | MRC-5 | •OH, and O2•− scavenging activity, carbon-centered radical scavenging assay, Q-TOF MS, lipid peroxidation inhibition assay, MTT assay |
| [86] |
|
| Oyster (Saccostrea cucullata) | Vero | DPPH• scavenging activity, Q-TOF MS, UPLC, GPC |
| [87] |
| LPHSGY | 672 | Alaska pollock —frame (Theragra chalcogramma) | - | •OH scavenging activity, HPLC |
| [88] |
| - | Alaska pollock —skin | Ac2F | TBAR assay, MTT assay |
| [89] |
| HGPLGPL | 797 | Hoki—skin (Johnius belengerii) | Hep3B | DPPH• and O2•− scavenging activity, SOD, GPx, and CAT activity |
| [90] |
| GSTVPERTHPACPNFN | 1801 | Hoki—frame (Johnius belengerii) | MRC-5 | DPPH•, •OH, ROO•, and O2•− scavenging activity, IEC, HPLC, Q-TOF MS, MTT assay |
| [91] |
|
| Chub mackerel (Scomber japonicus) | - | DPPH• scavenging activity, FRAP assay, SOD-like activity, MALDI-TOF MS/MS, RP-HPLC |
| [4] |
|
| Spanish mackerel (Scomberomorus niphonius) | - | DPPH•, •OH, and O2•− scavenging activity, lipid peroxidation inhibition assay, plasmid DNA protective assay |
| [92] |
| NHREDR | 856 | Horse mackerel—skin (Magalaspis cordyla) | - | DPPH• and •OH scavenging activity, lipid peroxidation inhibition assay, Fe2+ chelating activity, IEC, GPC, ESI-MS/MS |
| [83] |
| ACFL | 518.5 | Horse mackerel—viscera (Magalaspis cordyla) | - | Lipid peroxidation inhibition assay, DPPH•, and •OH scavenging activity, FPLC, Q-TOF MS |
| [93] |
|
| Mackerel—skin (Scomber japonicus) | - | MALDI-TOF MS/MS, DPPH•, and ABTS• scavenging activity, FRAP assay, Fe2+ chelating assay, FTIR assay |
| [94] |
|
| Mackerel—bone (Scomber japonicus) | - | MALDI-TOF MS/MS, DPPH•, and ABTS• scavenging activity, FRAP assay, Fe2+ chelating assay, FTIR assay |
| [94] |
|
| Mackerel—byproducts (Scomber japonicus) | - | Protease activity, MALDI-TOF MS/MS, DPPH•, and ABTS• scavenging activity |
| [95] |
|
| Chinese drum (Miichthys miiuy) | HUVECs | Hoechst 33,342 staining assay, SOD and GPx levels, DNA oxidative damage by H2O2 |
| [96] |
| Purified peptide fraction—II | - | Exocoetus volitans | Vero | DPPH•, •OH, and O2•− scavenging activity, lipid peroxidation inhibition assay, MTT assay, IEC, HPLC |
| [97] |
| LVPVAVF | 743.45 | Silver carp (Hypophthalmichthys molitrix) | Caco-2 | DPPH• scavenging activity, lipid peroxidation inhibition assay, intracellular ROS, SE-HPLC, RP-HPLC, ORAC, FRAP |
| [98] |
|
| Bluefin leatherjacket—skin (Navodon septentrionalis) | - | DPPH•, •OH, and O2•− scavenging activity, metal chelating activity, RP-HPLC |
| [99] |
| DLYANTVLSGGTTMYPGIADR | 2214.06 | Hairtail surimi | - | DPPH• and •OH scavenging activity, TAO assay, gel filtration chromatography, RP-HPLC, LC-MS/MS |
| [100] |
| GDRGESGPA | 845.37 | Sturgeon—skin (Acipenser schrencki) | - | Fe2+ chelating activity, DPPH• scavenging activity, SP-RP-HPLC, LC-MS/MS |
| [101] |
| KGFR | 506 | Round scad (Decapterus maruadsi) | - | DPPH• and •OH scavenging activity, FRAP, gel filtration chromatography, RP-HPLC, LC-MS/MS |
| [102] |
|
| Monkfish (Lophius litulon) | HepG2 | DPPH•, •OH, H2O2, and O2•− scavenging activity, ROS levels, lipid peroxidation inhibition assay, Q-TOF MS, MTT assay |
| [103] |
| ATSHH | 551.25 | Sandfish (Arctoscopus japonicus) | - | DPPH•, •OH, and O2•− scavenging activity, RP-HPLC, Q-TOF MS |
| [104] |
| - | Greater weever (Trachinus draco) | - | RP-HPLC, CUPRAC, Cu2+ chelating activity, RP-HPLC, nano-ESI-MS/MS |
| [105] |
| Purified peptide fraction—I | - | Nemipterus japonicus | Vero | DPPH•, •OH, and O2•− scavenging activity, lipid peroxidation inhibition assay, MTT assay, IEC, HPLC |
| [97] |
| WL15 (WHKNCFRCAKCGKSL) | - | Snakehead murrel (Channa striatus) | HDF | DPPH•, ABTS•, O2•−, and H2O2 scavenging activity, cytotoxicity assay |
| [106] |
| WL15 | - | Snakehead murrel (Channa striatus) | L6 myoblast | Intracellular ROS level, lipid peroxidation inhibition assay, SOD and CAT assay, MTT assay |
| [107] |
| GGFDMG | 582 | Japanese flounder—skin (Paralichthys olivaceus) | RAW264.t | FPLC, HPLC, DPPH•, cell viability assay, lipid peroxidation inhibition assay, radical-mediated damage to membrane lipids, DNA and protein |
| [108] |
| FTGML | - | Grass carp—scales (Latrunculia magnifica) | B16F10 | CCK-8 assay, apoptosis rate |
| [109] |
AAPH: 2,2′-Azobis (2-amidinopropane) dihydrochloride; ABTS•: 2,2′-Azino-bis (3-ethylbenzthiazoline-6-sulfonic acid) radical; Ac2F: Donryu rat liver cells; CAT: Catalase; CUPRAC: Cupric reducing antioxidant capacity; DPPH•: 2,2-Diphenyl-1-picrylhydrazyl radical; ECV304: Human endothelial cells; ESI-MS/MS: Electrospray ionization-mass spectrometry; FPLC: Fast protein liquid chromatography; FRAP: Ferric reducing antioxidant power; FTIR: Fourier transform infrared spectroscopy; GPC: Gel permeation chromatography; GPx: Glutathione peroxidase; GSH: Glutathione; IEC: Ion-exchange chromatography; HepG2: Human hepatocellular liver carcinoma cells; Hep3B: Human hepatoma cells; HO•: Hydroxyl peroxide radical; HPLC: High-performance liquid chromatography; H2O2: Hydrogen peroxide; IC50: The half maximal inhibitory concentration; LC-MS/MS: Liquid chromatography-tandem mass spectrometry; LC/ESI-MS/MS: Liquid chromatography-electrospray ionization-mass spectrometry; L6 myoblast: Immortalized rat skeletal (L6) myoblast cells; MALDI-TOF MS/MS: Matrix-assisted laser desorption ionization-time of flight mass spectrometry; MDA: Malonaldehyde; MRC-5: Human embryonic lung fibroblast cells; MTT: 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide; nano-ESI-MS/MS: Nano liquid chromatography-tandem mass spectrometry; ORAC: Oxygen radical absorbance capacity; O2•−: Superoxide radical; Q-TOF MS: Quadrupole time of flight mass spectrometry; RAW264.t: Mouse monocyte cells; ROO•: Peroxyl redical; RP-HPLC: Reversed-phase high-performance liquid chromatography; SEC: Size-exclusion chromatography; SP-RP-HPLC: Semipreparative reversed-phase high-performance liquid chromatography; SOD: Superoxide dismutase; TAO: Total antioxidant capacity; TBARS: Thiobarbituric acid reactive substances; UPLC: Ultra-performance liquid chromatography; Vero: Kidney epithelial cells of the African Green Monkey.
Table 2.
Antioxidant effects of bioactive peptides from marine animals—in vivo studies.
| Peptide Name/Peptide Containing Compound | Size of Peptide (Da) | Source | Organism | Analysis | Major Findings | Reference |
|---|---|---|---|---|---|---|
| WL15 | - | Snakehead murrel (Channa striatus) | Adult zebrafish (wild-type, AB strain; 4 months old) | ROS production assay, cytotoxicity assay, caspase 3, SOD, CAT, GST, GPx, and GCS expressions |
| [106] |
| KTFCGRH | 861.6 | Croaker (Otolithes ruber) | 9 Adult albino Wistar male rats (body weight range: 150–180 g) | CAT, GST, and SOD activity |
| [110] |
| ACFL | 518.5 | Horse mackerel—viscera (Magalaspis cordyla) | 9 adult albino Wistar male rats (body weight range: 150–180 g) | CAT, SOD, and GST activity |
| [111] |
| Oligopeptide | 1000 | Frigate tuna (Auxis thazard) | 60 Kunming SPF-SD male mice 4 weeks old (average body weight: 20 ± 2 g) | Body weight and organ index, XOD, MDA, ADA, SOD, and CAT activity, URAT1, GLUT9, OAT1, and ABCG2 expressions |
| [112] |
| Collagen oligopeptides | <1000 | Tilapia—scales | 72 SPF-SD male rats (6–8 weeks old) | SOD, GPx, CAT, and MDA activity |
| [113] |
ABCG2: ATP-binding cassette subfamily G2; ADA: Adenosine deaminase activity; CAT: Catalase; GCS: γ-glutamyl cysteine synthetase; GLUT9: Glucose transporter; GPx: Glutathione peroxidase; GST: Glutathione S-transferase; MDA: Malondialdehyde; OAT1: Organic anion transporter; ROS: Reactive oxygen species; SPF-SD rats: Specific pathogen-free Sprague Dawley rats; SOD: Superoxide dismutase; URAT1: Urate transporter; XOD: Xanthine oxidase.
Table 3.
Antioxidant effects of bioactive peptides from marine animals—clinical trials.
| Peptide/Peptide Containing Compound | Source | Subject | Analysis | Major Findings | Phase | Reference |
|---|---|---|---|---|---|---|
| Low-molecular-weight collagen peptide (LMWCP) | Sutchi catfish—skin (Pangasius hypophthalmus) | 64 healthy male adults (40–60 years old) | Skin hydration, wrinkling, and elasticity |
| I | [114] |
| GOLD COLLAGEN® ACTIVE Hydrolyzed fish collagen complex (Approved and patent granted in Italy: 0001413152) | Fish collagen | 122 adults (91 male, 29 female) (21–70 years old) | Skin elasticity, histological examination of skin, SPQs, joint health questionnaire |
| Approved | [115] |
| CELERGEN® Marine collagen peptides combined with plant peptides | Skin of Pollachius virens, Hippoglossus hippoglossus, and Pleuronectes platessa, | 41 adults (36 male, 5 female) (37–72 years old) | Plasma levels of nitrites/nitrates, MDA, Cu- and Zn-SOD3, facial skin properties |
| Approved | [116] |
AU: Arbitrary units; MDA: Malondialdehyde; Cu-SOD: Copper superoxide dismutase; SPQs: Self-perception questionnaires; Zn SOD: Zinc superoxide dismutase.
Table 4.
Anticancer effects of bioactive peptides from marine animals—in vitro studies.
| Peptide Name/Sequence | Size of Peptide (Da) | Source | Cell Line | Analysis | Major Findings | Reference |
|---|---|---|---|---|---|---|
|
| Tuna fish (Thunnus tonggol) | MCF-7 | Antiproliferative activity, MTT assay |
| [123] |
| Peptide fraction-AobsII | 440.9 | Anchovy—sauce | U937 | MTT assay, DNA fragmentation analysis, cell cycle analysis, silica gel chromatography |
| [124] |
| LANAK | 515.29 | Oyster (Saccostrea cucullata) | HT-29 | MTT assay, AO/EtBr staining, comet assay |
| [87] |
| AFNIHNRNLL | - | Mussel (Mytilus coruscus) | PC-3, A549, MDA-MB-231 | Apoptosis assay, IEC, HPLC |
| [125] |
| Pardaxin | - | Finless sole (Pardachirus marmoratus) | MN-11 | MTS assay, membrane structure examination, apoptosis assay |
| [126] |
| LKEENRRRRD | 1371.53 | Sepia—ink (Sepia esculenta) | PC-3 | Apoptosis assay, AO/EtBr staining, Western blot |
| [127] |
| - | Japanese flounder (Paralichthys olivaceus) | HT-29 | MTT assay, qRT-PCR |
| [128] |
|
| Spanish mackerel (Scomberomorus niphonius) | B16F10 | MTT assay, UPLC, UPLC-ESI-MS/MS |
| [129] |
| FIMGPY | 726.9 | Skate—cartilage (Raja porosa) | HeLa | MTT assay, Western blot |
| [120] |
| GIKCRFCCGCCTPGICGVCCRF-NH2 (Amidated C-terminus) | - | Tilapia (Oreochromis mossambicus) | HeLa, HepG2, HT1080, COS-7, WS-1 | MTT assay, soft-agar assay, wound-healing assay, LDH release assay, AO/EtBr staining |
| [130] |
| KF | 1476 | Marine mollusk (Elysia rufescens) | PC3, DU145, LNCaP, SKBR-3, BT474, MCF-7, MDA-MB-231, LoVo, MCF10A, HUVEC, HMEC-1, IMR90 | Measurement of DNA synthesis, effect of BCL-2, MDR-1, and HER2/neu, cytotoxicity assay |
| [131] |
AO/EtBr: Acridine orange/ethidium bromide; A549: Type II pulmonary epithelial cells; BCL-2: B-cell lymphoma 2; BT474: Human breast cancer cells; BT549: Breast carcinoma cells; B16F10: murine melanoma cell line from a C57BL/6J mouse; Caco-2: Human colon cancer cells; COS-7: Cercopithecus aethiops kidney cells; DU145: Human prostate cancer cell lines; HCT15: Colon carcinoma cells; HeLa: Human cervix adenocarcinoma cells; HepG2: Human liver cancer cells; HER2/neu: Human epidermal growth factor receptor 2; HMEC-1: Human Mammary epithelial cells; HPLC: High-performance liquid chromatography; HT-29: Human colon cancer cells; HT1080: Human fibrosarcoma cells; HUVEC: Human umbilical vein endothelial cell line; IEC: Ion-exchange chromatography; IMR90: Normal lung fibroblast cells; IC50: The half maximal inhibitory concentration; KF: Kahalalide F; LC50: Lethal concentration that kills 50% of test population; LDH: Lactate dehydrogenase; LNCaP: Human prostate cancer cells; LoVo: Human colon (supraclavicular lymph node metastasis) cells; MCF-7: Human breast cancer cells; MCF10A: Human breast epithelial cell line; MDA-MB-231: Human breast cancer cells; MDR-1: Multidrug resistance mutation; MKN45: Human gastric carcinoma cells; MN-11: Mouse fibrosarcoma cells; MTT: 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide; MTS: 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium; NUGC-4: Signet-ring carcinoma cells; PC3: Prostate cancer cells; qRT-PCR: Real-Time Quantitative Reverse Transcription PCR; SKBR-3: Human breast cancer cell; UPLC-ESI-MS/MS: Ultra-performance liquid chromatography-electrospray ionization-mass spectrometry; U937: Human lymphoma cells; WS-1: Human kidney cells.
Table 5.
Anticancer effects of bioactive peptides from marine animals—in vivo studies.
| Peptide Name/Peptide Containing Compound | Size of Peptide (Da) | Source | Organism | Analysis | Major Findings | Reference |
|---|---|---|---|---|---|---|
| Pardaxin | - | Finless sole (Pardachirus marmoratus) | Syrian golden hamsters (5–6 weeks old) | Tumor volume measurement, PGE2 level assay |
| [132] |
| 20 C57BL/6 female mice (35 days old) | Analysis of GOT, GPT, BUN, Cre, UA, TCHO, TG, TBIL, TP, and ALB levels, cytokine expression levels, and immunohistochemistry |
| [126] | |||
| Conus venom | - | Vexillum cone snail (Conus vexillum) | 54 adult Swiss albino male mice (Average body weight: 20–25 g) | Lipid peroxidation inhibition assay, estimation of nitric oxide, CAT, Cu/Zn SOD, and LDH activity, TAC and GSH content |
| [133] |
ALB: Albumin; BUN: Blood urea nitrogen; CAT: Catalase; Cre: Creatinine; CU/Zn SOD: Copper/zinc superoxide dismutase; GOT: Glutamic oxaloacetic transaminase; GPT: Glutamic pyruvic transaminase; GSH: Glutathione; LDH: Lactate dehydrogenase; MDA: Malondialdehyde; PGE2: Prostaglandin E2; TAC: Total antioxidant capacity; TBIL: Total bilirubin; TCHO: Total cholesterol; TG: Triglyceride; TP: Total protein; UA: Uric acid.
Table 6.
Anticancer effects of bioactive peptides from marine animals—clinical trials.
| Peptide/Peptide Containing Compound | Source | Subject | Analysis | Major Findings | Phase | Reference |
|---|---|---|---|---|---|---|
| KF | Marine mollusk (Elysia rufescens) | 38 cancer patients (23 male and 15 female; 27–73 years old) | Pharmacokinetic analysis |
| I | [134] |
| 32 cancer patients (49–81 years old) | Pharmacokinetic analysis |
| I | [135] | ||
| 24 cancer patients (28–89 years old) | RECIST |
| II | [136] | ||
| DOLA-10 | Marine mollusk (Dolabella Auricularia) | 22 cancer patients (10 male and 12 female; 27–72 years old) | Pharmacokinetic and pharmacodynamic analysis |
| I | [137] |
| 10 NSCLC patients (female; 47–80 years old) | Physical examination, complete blood count, computed timigraphy, and liver function tests |
| II | [138] | ||
| 12 advanced melanoma patients (7 male and 5 female; 26–80 years old) | Pharmacokinetic analysis |
| II | [139] |
DOLA-10: Dolastatin-10; KF: Kahalalide F; NSCLC: Non-small-cell lung carcinoma; RECIST: Response evaluation criteria in solid tumor.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chakniramol, S.; Wierschem, A.; Cho, M.-G.; Bashir, K.M.I. Physiological and Clinical Aspects of Bioactive Peptides from Marine Animals. Antioxidants 2022, 11, 1021. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11051021
AMA Style
Chakniramol S, Wierschem A, Cho M-G, Bashir KMI. Physiological and Clinical Aspects of Bioactive Peptides from Marine Animals. Antioxidants. 2022; 11(5):1021. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11051021
Chicago/Turabian StyleChakniramol, Sukwasa, Andreas Wierschem, Man-Gi Cho, and Khawaja Muhammad Imran Bashir. 2022. "Physiological and Clinical Aspects of Bioactive Peptides from Marine Animals" Antioxidants 11, no. 5: 1021. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11051021
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.