Chemical Characterization and Bioactivity of Extracts from Thymus mastichina: A Thymus with a Distinct Salvianolic Acid Composition

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Standards and Reagents
2.2. Plant Material
2.3. Preparation of Extracts
2.4. Total Phenolic Compound, Total Flavonoid, and Ortho-Diphenol Contents
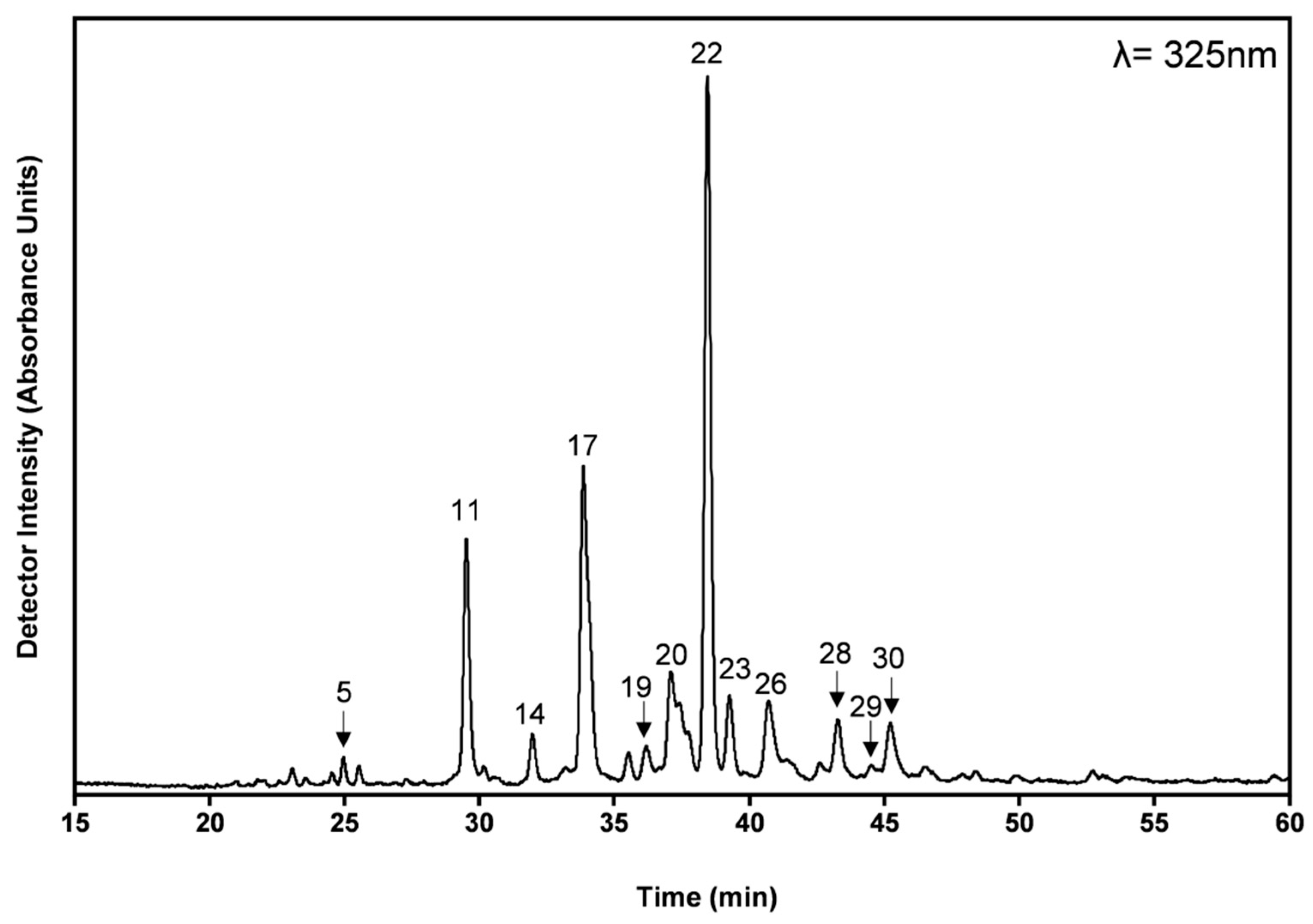
2.5. Profiling and Quantification of Individual Phenolic Compounds by High Performance Liquid Chromatography with Diode Array Detector (HPLC-DAD) and High Performance Liquid Chromatography with Electrospray Ionization and Tandem Mass Spectrometry Detection (HPLC-ESI-MSn)
2.6. Determination of Oleanolic Acid and Ursolic Acid in Hydroethanolic Extracts
2.7. In Vitro Antioxidant Activity Assessment
2.7.1. ABTS Radical Cation Decolorization Assay
2.7.2. Hydroxyl Radicals Scavenging Assay
2.7.3. Nitric Oxide Radical Scavenging Assay
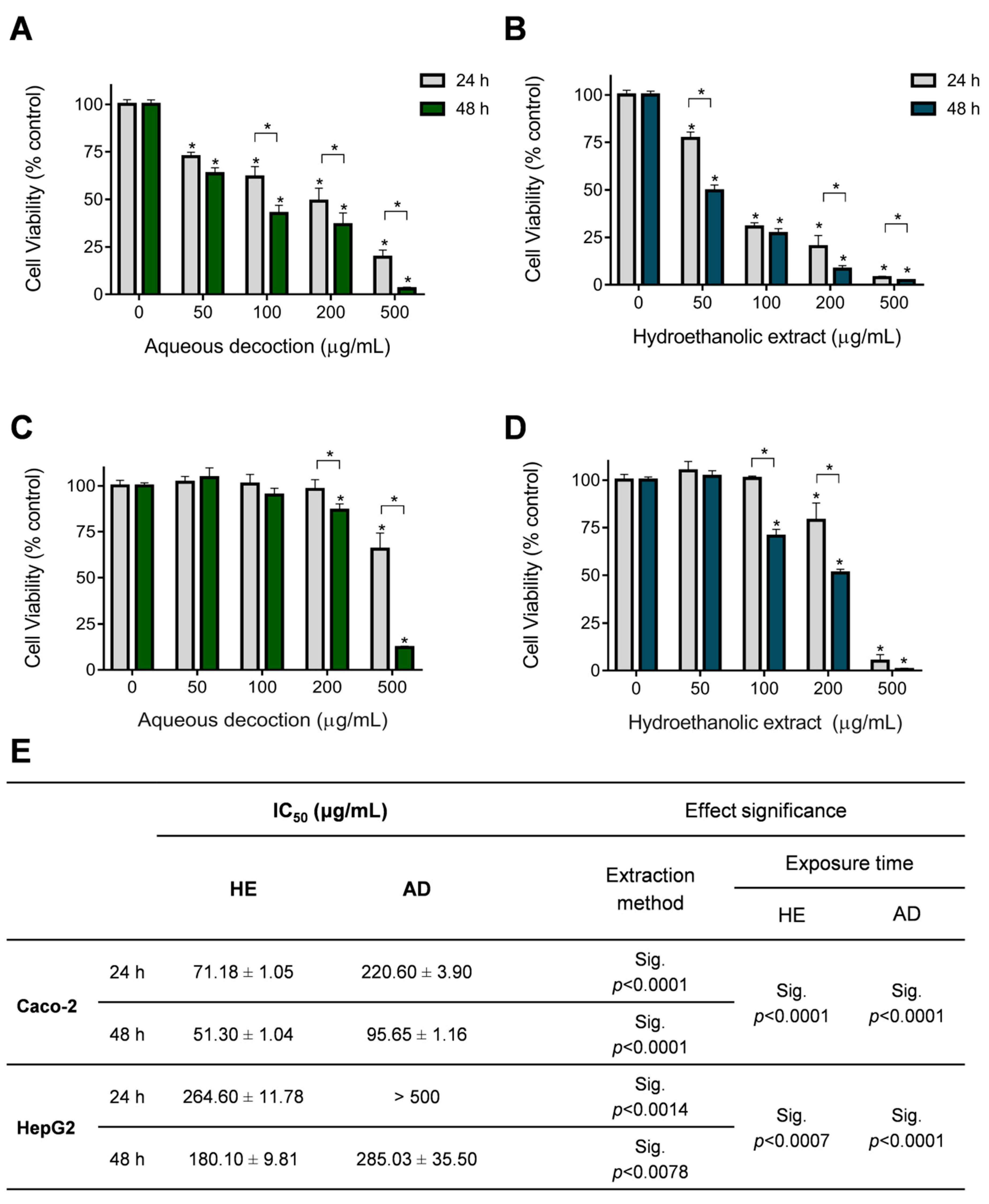
2.8. In Vitro Cell Viability Assay
2.9. Statistical Analysis
3. Results and Discussion
3.1. Extract Yield and Chemical Composition of Extracts
3.2. Phenolic Compound Profiles in Aqueous Decoction and Hydroethanolic Extracts
3.3. Oleanolic Acid and Ursolic acid Contents
3.4. In Vitro Antioxidant Activity
3.5. Anti-Proliferative Effect of T. mastichina Extracts
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Chemical Compounds Studied in this Article
References
- Kumar, A.; Premoli, M.; Aria, F.; Bonini, S.A.; Maccarinelli, G.; Gianoncelli, A.; Memo, M.; Mastinu, A. Cannabimimetic plants: Are they new cannabinoidergic modulators? Planta 2019, 249, 1681–1694. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.M.; Silva, S.C.; Soares, J.P.; Martins-Gomes, C.; Teixeira, J.P.; Leal, F.; Gaivão, I. Ginkgo biloba L. leaf extract protects HepG2 cells against paraquat-induced oxidative DNA damage. Plants 2019, 8, 556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Seedi, H.R.; Burman, R.; Mansour, A.; Turki, Z.; Boulos, L.; Gullbo, J.; Goransson, U. The traditional medical uses and cytotoxic activities of sixty-one Egyptian plants: Discovery of an active cardiac glycoside from Urginea maritima. J. Ethnopharmacol. 2013, 145, 746–757. [Google Scholar] [CrossRef] [PubMed]
- Leal, F.; Taghouti, M.; Nunes, F.M.; Silva, A.M.; Coelho, A.C.; Matos, M. Thymus Plants: A review—Micropropagation, molecular and antifungal activity. In Active Ingredients from Aromatic and Medicinal Plants; El-Shemy, H.A., Ed.; InTech: London, UK, 2017; Chapter 7; pp. 107–126. [Google Scholar] [CrossRef] [Green Version]
- Kindl, M.; Blazekovic, B.; Bucar, F.; Vladimir-Knezevic, S. Antioxidant and Anticholinesterase Potential of Six Thymus Species. Evid. Based Complement. Altern. Med. 2015, 2015, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins-Gomes, C.; Souto, E.B.; Cosme, F.; Nunes, F.M.; Silva, A.M. Thymus carnosus extracts induce anti-proliferative activity in Caco-2 cells through mechanisms that involve cell cycle arrest and apoptosis. J. Funct. Foods 2019, 54, 128–135. [Google Scholar] [CrossRef]
- Mahmoudi, M.; Morteza-Semnani, K.; Mojra, E. Anti-inflammatory and antinociceptive activity of Thymus pubescens extract. Fitoterapia 2008, 79, 361–365. [Google Scholar] [CrossRef]
- Khouya, T.; Ramchoun, M.; Hmidani, A.; Amrani, S.; Harnafi, H.; Benlyas, M.; Filali Zegzouti, Y.; Alem, C. Anti-inflammatory, anticoagulant and antioxidant effects of aqueous extracts from Moroccan thyme varieties. Asian Pac. J. Trop. Biomed. 2015, 5, 636–644. [Google Scholar] [CrossRef] [Green Version]
- Martins-Gomes, C.; Taghouti, M.; Schäfer, J.; Bunzel, M.; Silva, A.M.; Nunes, F.M. Chemical characterization and bioactive properties of decoctions and hydroethanolic extracts of Thymus carnosus Boiss. J. Funct. Foods 2018, 43, 154–164. [Google Scholar] [CrossRef]
- Esmaeili-Mahani, S.; Falahi, F.; Yaghoobi, M.M. Proapoptotic and antiproliferative effects of Thymus caramanicus on human breast cancer cell line (MCF-7) and its interaction with anticancer drug vincristine. Evid. Based Complement. Altern. Med. 2014, 2014, 7. [Google Scholar] [CrossRef] [Green Version]
- Dall’Acqua, S.; Peron, G.; Ferrari, S.; Gandin, V.; Bramucci, M.; Quassinti, L.; Martonfi, P.; Maggi, F. Phytochemical investigations and antiproliferative secondary metabolites from Thymus alternans growing in Slovakia. Pharm. Biol. 2017, 55, 1162–1170. [Google Scholar] [CrossRef] [Green Version]
- Taghouti, M.; Martins-Gomes, C.; Schafer, J.; Felix, L.M.; Santos, J.A.; Bunzel, M.; Nunes, F.M.; Silva, A.M. Thymus pulegioides L. as a rich source of antioxidant, anti-proliferative and neuroprotective phenolic compounds. Food Funct. 2018, 9, 3617–3629. [Google Scholar] [CrossRef] [PubMed]
- Pereira, O.R.; Peres, A.M.; Silva, A.M.S.; Domingues, M.R.M.; Cardoso, S.M. Simultaneous characterization and quantification of phenolic compounds in Thymus x citriodorus using a validated HPLC–UV and ESI–MS combined method. Food Res. Int. 2013, 54, 1773–1780. [Google Scholar] [CrossRef]
- Gordo, J.; Maximo, P.; Cabrita, E.; Lourenco, A.; Oliva, A.; Almeida, J.; Filipe, M.; Cruz, P.; Barcia, R.; Santos, M.; et al. Thymus mastichina: Chemical constituents and their anti-cancer activity. Nat. Prod. Commun. 2012, 7, 1491–1494. [Google Scholar] [CrossRef] [PubMed]
- Pardo-de-Santayana, M.; Tardio, J.; Blanco, E.; Carvalho, A.M.; Lastra, J.J.; San Miguel, E.; Morales, R. Traditional knowledge of wild edible plants used in the northwest of the Iberian Peninsula (Spain and Portugal): A comparative study. J. Ethnobiol. Ethnomed. 2007, 3, 27. [Google Scholar] [CrossRef] [Green Version]
- Barros, L.; Heleno, S.A.; Carvalho, A.M.; Ferreira, I.C.F.R. Lamiaceae often used in Portuguese folk medicine as a source of powerful antioxidants: Vitamins and phenolics. LWT Food Sci. Technol. 2010, 43, 544–550. [Google Scholar] [CrossRef]
- Méndez-Tovar, I.; Sponza, S.; Asensio-S-Manzanera, M.C.; Novak, J. Contribution of the main polyphenols of Thymus mastichina subsp. mastichina to its antioxidant properties. Ind. Crop. Prod. 2015, 66, 291–298. [Google Scholar] [CrossRef]
- Moldao-Martins, M.; Beirao-da-Costa, S.; Neves, C.; Cavaleiro, C.; Salgueiro, L.; Beirao-da-Costa, M.L. Olive oil flavoured by the essential oils of Mentha x piperita and Thymus mastichina L. Food Qual. Prefer. 2004, 15, 447–452. [Google Scholar] [CrossRef]
- Delgado, T.; Marinero, P.; Asensio-S-Manzanera, M.C.; Asensio, C.; Herrero, B.; Pereira, J.A.; Ramalhosa, E. Antioxidant activity of twenty wild Spanish Thymus mastichina L. populations and its relation with their chemical composition. LWT Food Sci. Technol. 2014, 57, 412–418. [Google Scholar] [CrossRef] [Green Version]
- Jia, Z.; Tang, M.C.; Wu, J.M. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Shahidi, F. Nutraceuticals and functional foods: Whole versus processed foods. Trends Food Sci. Technol. 2009, 20, 376–387. [Google Scholar] [CrossRef]
- Daliu, P.; Santini, A.; Novellino, E. From pharmaceuticals to nutraceuticals: Bridging disease prevention and management. Expert Rev. Clin. Pharmacol. 2019, 12, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Santini, A.; Tenore, G.C.; Novellino, E. Nutraceuticals: A paradigm of proactive medicine. Eur. J. Pharm. Sci. 2017, 96, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.; Felizardo, C.; Fernandes-Silva, A.A.; Nunes, F.M.; Barros, A. Polyphenolic compounds, antioxidant activity and l-phenylalanine ammonia-lyase activity during ripening of olive cv. “Cobrancosa” under different irrigation regimes. Food Res. Int. 2013, 51, 412–421. [Google Scholar] [CrossRef]
- Rao, M.N.A. Nitric oxide scavenging by curcuminoids. J. Pharm. Pharmacol. 1997, 49, 105–107. [Google Scholar] [CrossRef]
- Severino, P.; Andreani, T.; Jager, A.; Chaud, M.V.; Santana, M.H.A.; Silva, A.M.; Souto, E.B. Solid lipid nanoparticles for hydrophilic biotech drugs: Optimization and cell viability studies (Caco-2 & HEPG-2 cell lines). Eur. J. Med. Chem. 2014, 81, 28–34. [Google Scholar] [CrossRef]
- Andreani, T.; Kiill, C.P.; de Souza, A.L.R.; Fangueiro, J.F.; Fernandes, L.; Doktorovova, S.; Santos, D.L.; Garcia, M.L.; Gremiao, M.P.D.; Souto, E.B.; et al. Surface engineering of silica nanoparticles for oral insulin delivery: Characterization and cell toxicity studies. Colloids Surf. B 2014, 123, 916–923. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.M.; Martins-Gomes, C.; Coutinho, T.E.; Fangueiro, J.F.; Sanchez-Lopez, E.; Pashirova, T.N.; Andreani, T.; Souto, E.B. Soft cationic nanoparticles for drug delivery: Production and cytotoxicity of solid lipid nanoparticles (SLNs). Appl. Sci. 2019, 9, 4438. [Google Scholar] [CrossRef] [Green Version]
- Stankovic, M.S.; Radic, Z.S.; Blanco-Salas, J.; Vazquez-Pardo, F.M.; Ruiz-Tellez, T. Screening of selected species from Spanish flora as a source of bioactive substances. Ind. Crops Prod. 2017, 95, 493–501. [Google Scholar] [CrossRef]
- Martins-Gomes, C.; Nunes, F.M.; Sampaio, A.; Souto, E.B.; Silva, A.M. Rosmarinic acid: Sources, bioactivities and health benefits. In Phytochemicals: Plant Sources and Potential Health Benefits; Ryan, I., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2019; pp. 109–146. [Google Scholar]
- Afonso, A.F.; Pereira, O.R.; Válega, M.; Silva, A.M.S.; Cardoso, S.M. Metabolites and biological activities of Thymus zygis, Thymus pulegioides, and Thymus fragrantissimus grown under organic cultivation. Molecules 2018, 23, 1514. [Google Scholar] [CrossRef] [Green Version]
- Ziani, B.E.C.; Heleno, S.A.; Bachari, K.; Dias, M.I.; Alves, M.J.; Barros, L.; Ferreira, I.C.F.R. Phenolic compounds characterization by LC-DAD-ESI/MSn and bioactive properties of Thymus algeriensis Boiss. & Reut. and Ephedra alata Decne. Food Res. Int. 2019, 116, 312–319. [Google Scholar] [CrossRef] [Green Version]
- Desta, K.T.; Kim, G.S.; Abd El-Aty, A.M.; Raha, S.; Kim, M.B.; Jeong, J.H.; Warda, M.; Hacimuftuoglu, A.; Shin, H.C.; Shim, J.H.; et al. Flavone polyphenols dominate in Thymus schimperi Ronniger: LC-ESI-MS/MS characterization and study of anti-proliferative effects of plant extract on AGS and HepG2 cancer cells. J. Chromatogr. B 2017, 1053, 1–8. [Google Scholar] [CrossRef]
- Nagy, T.O.; Solar, S.; Sontag, G.; Koenig, J. Identification of phenolic components in dried spices and influence of irradiation. Food Chem. 2011, 128, 530–534. [Google Scholar] [CrossRef]
- Kozics, K.; Klusova, V.; Srancikova, A.; Mucaji, P.; Slamenova, D.; Hunakova, L.; Kusznierewicz, B.; Horvathova, E. Effects of Salvia officinalis and Thymus vulgaris on oxidant-induced DNA damage and antioxidant status in HepG2 cells. Food Chem. 2013, 141, 2198–2206. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Liang, Q.L.; Luo, G.A.; Zhao, Z.Z.; Jiang, Z.H. Multi-component HPLC fingerprinting of Radix Salviae miltiorrhizae and its LC-MS-MS identification. Chem. Pharm. Bull. 2005, 53, 677–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afonso, F.A.; Pereira, R.O.; Fernandes, S.F.Â.; Calhelha, C.R.; Silva, M.S.A.; Ferreira, C.F.R.I.; Cardoso, M.S. The Health-Benefits and Phytochemical Profile of Salvia apiana and Salvia farinacea var. Victoria Blue Decoctions. Antioxidants 2019, 8, 241. [Google Scholar] [CrossRef] [Green Version]
- Ai, C.B.; Li, L.N. Salvianolic acids D and E: Two new depsides from Salvia miltiorrhiza. Planta Med. 1992, 58, 197–199. [Google Scholar] [CrossRef]
- Xie, X.; Sun, W.; Miao, J.; Huang, J.; Xu, J.; Liu, X.; Sun, H.; Tong, L.; Sun, G. Development and validation of a UFLC-MS/MS method for determination of 7′(Z)-(8″S, 8‴S)-epi-salvianolic acid E, (7′R, 8′R, 8″S, 8‴S)-epi-salvianolic acid B and salvianolic acid B in rat plasma and its application to pharmacokinetic studies. J. Chromatogr. B 2016, 1022, 6–12. [Google Scholar] [CrossRef]
- Pereira, O.R.; Cardoso, S.M. Overview on Mentha and Thymus polyphenols. Curr. Anal. Chem. 2013, 9, 382–396. [Google Scholar] [CrossRef] [Green Version]
- Janicsak, G.; Veres, K.; Kakasy, A.Z.; Mathe, I. Study of the oleanolic and ursolic acid contents of some species of the Lamiaceae. Biochem. Syst. Ecol. 2006, 34, 392–396. [Google Scholar] [CrossRef]
- Petrović, S.; Ušjak, L.; Milenković, M.; Arsenijević, J.; Drobac, M.; Drndarević, A.; Niketić, M. Thymus dacicus as a new source of antioxidant and antimicrobial metabolites. J. Funct. Foods 2017, 28, 114–121. [Google Scholar] [CrossRef]
- Wang, Q.-L.; Wu, Q.; Tao, Y.-Y.; Liu, C.-H.; El-Nezami, H. Salvianolic acid B modulates the expression of drug-metabolizing enzymes in HepG2 cells. Hepatobiliary Pancreat. Dis. Int. 2011, 10, 502–508. [Google Scholar] [CrossRef]
- Hao, Y.; Xie, T.; Korotcov, A.; Zhou, Y.; Pang, X.; Shan, L.; Ji, H.; Sridhar, R.; Wang, P.; Califano, J.; et al. Salvianolic acid B inhibits growth of head and neck squamous cell carcinoma in vitro and in vivo via cyclooxygenase-2 and apoptotic pathways. Int. J. Cancer 2009, 124, 2200–2209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, T.; Rasul, A.; Sarfraz, A.; Sarfraz, I.; Hussain, G.; Anwar, H.; Riaz, A.; Liu, S.; Wei, W.; Li, J.; et al. Salvianolic acid A & B: Potential cytotoxic polyphenols in battle against cancer via targeting multiple signaling pathways. Int. J. Biol. Sci. 2019, 15, 2256–2264. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Tang, L.; Yi, Q. Salvianolic acids: Potential source of natural drugs for the treatment of fibrosis disease and cancer. Front. Pharmacol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, J.A.; Costa, R.; Fraga, H. Climate change impacts on thermal growing conditions of main fruit species in Portugal. Clim. Chang. 2017, 140, 273–286. [Google Scholar] [CrossRef]
- Costa, R.; Fraga, H.; Fernandes, P.M.; Santos, J.A. Implications of future bioclimatic shifts on Portuguese forests. Reg. Environ. Chang. 2017, 17, 117–127. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Hydroethanolic Extract | Aqueous Decoction | ||
|---|---|---|---|
| Extraction yield (%, w/w) | 13.78 ± 0.42 | 9.32 ± 1.74 * | |
| Chemical composition | |||
| Total phenols (mg Caffeic acid eq./g) | Ext. | 178.89 ± 8.89 | 134.76 ± 2.64 * |
| D.P. | 24.61 ± 0.67 | 12.51 ± 2.97 * | |
| Total flavonoids (mg Catechin eq./g) | Ext. | 184.45 ± 5.79 | 195.53 ± 48.78 |
| D.P. | 25.44 ± 1.57 | 17.37 ± 1.14 * | |
| Ortho-diphenols (mg Caffeic acid eq./g) | Ext. | 157.69 ± 19.34 | 107.87 ± 12.42 * |
| D.P. | 21.65 ± 2.83 | 10.49 ± 3.06 * | |
| CAntioxidant activity | |||
| ABTS•+ (mmol Trolox eq./g) | Ext. | 1.48 ± 0.06 | 0.96 ± 0.10 * |
| D.P. | 0.20± 0.00 | 0.08±0.01 * | |
| •OH radicals + EDTA (% inhibition) | 43.22 ± 5.28 | 48.52 ± 4.44 | |
| •OH radicals − EDTA (% inhibition) | 27.63 ± 2.56 | 28.23 ± 3.88 | |
| NO• radicals (% inhibition, after 120 min) | 38.87 ± 4.13 | 38.91 ± 3.01 | |
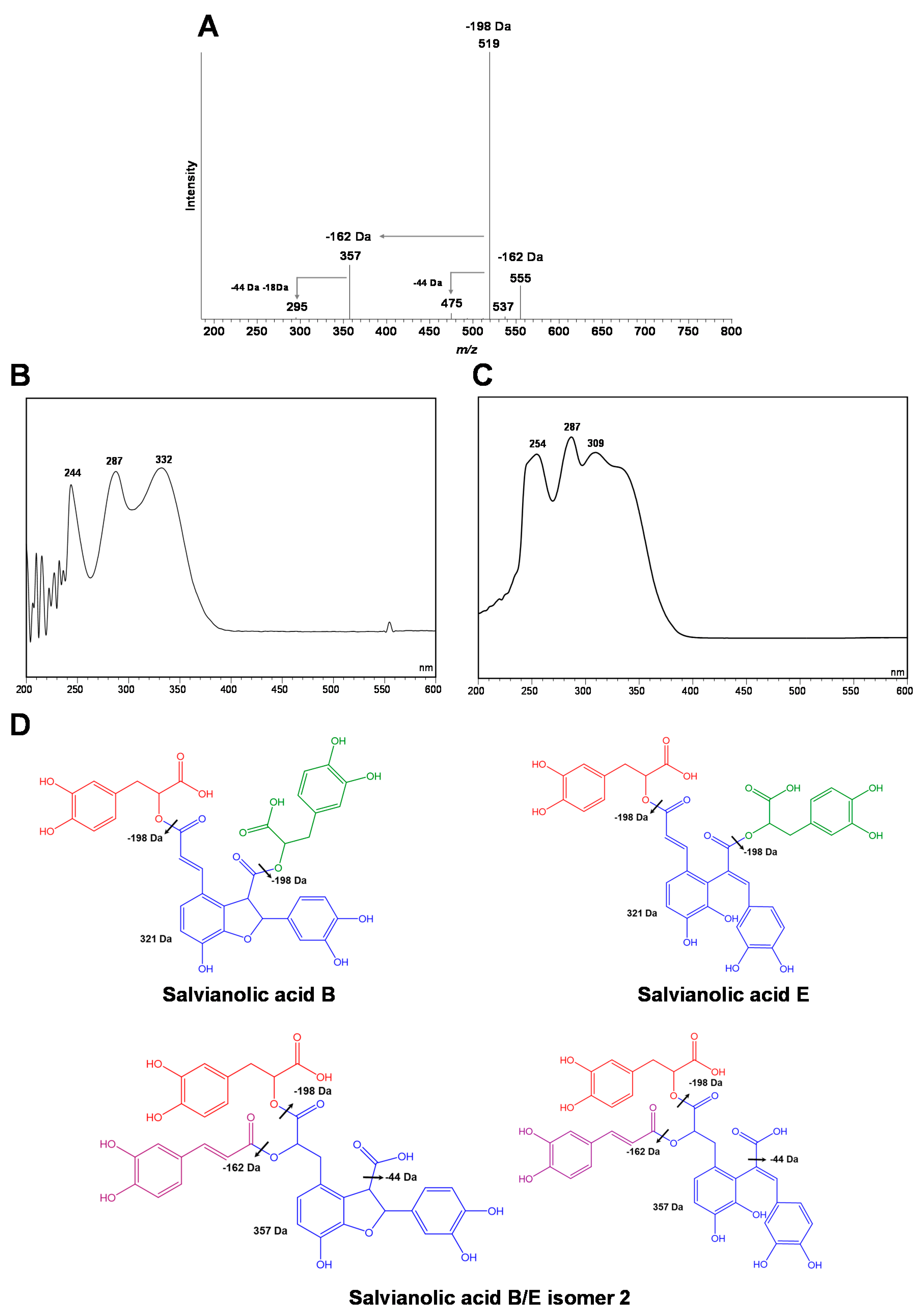
| Peak Number | Compound | R.T. (min) | ESI-MS2 | Quantification | ||||
|---|---|---|---|---|---|---|---|---|
| HE | AD | Extraction Method Sig. | ||||||
| mg/g D.P. | mg/g Extract | mg/g D.P. | mg/g Extract | |||||
| 1 | Eriodictyol-di-O-hexoside | 21.85 ± 0.07 | [611]:449;287 | n.q. | n.q. | n.q. | n.q. | |
| 2 | Naringenin-di-hexoside | 22.12 ± 0.07 | [595]:433;271 | n.q. | n.q. | n.d. | n.d. | |
| 3 | Chlorogenic acid | 23.09 ± 0.13 | [353]:191;179;173;135 | n.q. | n.q. | n.d. | n.d. | |
| 4 | Apigenin-(6,8)-C-diglucoside | 24.63 ± 0.09 | [593]:575;503;473;383 353 | n.q. | n.q. | n.d. | n.d. | |
| 5 | Hydroxyjasmonic acid–hexoside | 24.82 ± 0.06 | [387]:369;225;207;163 | n.q. | n.q. | n.q. | n.q. | |
| 6 | Caffeic acid | 25.08 ± 0.10 | [179]:135 | n.d. | n.d. | 0.21 ± 0.00 | 2.27 ± 0.03 | * |
| 7 | Eriodictyol-O-hexoside | 25.65 ± 0.04 | [449]:287 | n.q. | n.q. | n.q. | n.q. | |
| 8 | Unknown | 25.67 ± 0.04 | [495]:486;451;375;368 | n.d. | n.d. | n.q. | n.q. | |
| 9 | Prolithospermic acid | 28.33 ± 0.04 | [357]:313;269;245;203 | n.d. | n.d. | n.q. | n.q. | |
| 10 | Naringenin-O-hexoside | 29.19 ± 0.17 | [433]:313;271;267;137 | n.d. | n.d. | n.q. | n.q. | |
| 11 | Quercetin-O-hexoside | 29.66 ± 0.11 | [463]:301 | 2.80 ± 0.15 | 20.34 ± 1.11 | 0.77 ± 0.34 | 8.35 ± 3.69 | * |
| 12 | Naringenin-O-hexoside | 29.98 ± 0.62 | [433]:313;271 | n.q. | n.q. | n.q. | n.q. | |
| 13 | Eriodictyol-O-hexuronide | 31.95 ± 0.22 | [463]:287;175 | n.q. | n.q. | n.d. | n.d. | |
| 14 | Luteolin-O-hexoside | 32.11 ± 0.10 | [447]:285 | n.q. | n.q. | n.q. | n.q. | |
| 15 | Salvianolic acid F derivative | 32.72 ± 0.19 | [375]:313;269;179;135 | n.d. | n.d. | n.q. | n.q. | |
| 16 | Quercetin-O-hexuronide | 33.29 ± 0.15 | [477]:301 | n.q. | n.q. | n.d. | n.d. | |
| 17 | Luteolin-O-hexoside | 34.18 ± 0.18 | [447]:285 | 2.87 ± 0.56 | 20.85 ± 4.08 | 0.87 ± 0.29 | 9.34 ± 3.15 | * |
| 18 | Unknown | 35.63 ± 0.20 | [523]:505;477;454;391 | n.q. | n.q. | n.d. | n.d. | |
| 19 | Salvianolic acid B/E isomer 2 | 36.49 ± 0.37 | [717]:555;519;475;357;295 | 2.26 ± 0.09 | 16.40 ± 0.65 | 0.7±0.14 | 7.49 ± 1.55 | * |
| 20 | Salvianolic acid A isomer | 37.33 ± 0.22 | [493]:383;313;295 | 4.20 ± 0.42 | 30.47 ± 3.03 | 1.73 ± 0.54 | 18.57 ± 5.74 | * |
| 21 | Luteolin-O-hexorunide | 37.73 ± 0.29 | [461]:285;175 | n.q. | n.q. | n.q. | n.q. | |
| 22 | Rosmarinic acid | 38.65 ± 0.22 | [359]:223;179;161 | 8.00 ± 0.92 | 58.06 ± 6.74 | 3.15 ± 1.07 | 33.79 ± 11.48 | |
| 23 | Apigenin-O-hexoside | 39.46 ± 0.23 | [431]:269 | n.q. | n.q. | n.q. | n.q. | |
| 24 | Chrysoeriol-O-hexoside | 39.38 ± 0.16 | [461]:299;160 | n.q. | n.q. | n.q. | n.q. | |
| 25 | Salvianolic acid K | 40.81 ± 0.01 | [555]:537;493;359 | 0.57 ± 0.13 | 4.15 ± 0.96 | 0.07 ± 0.01 | 0.75 ± 0.07 | * |
| 26 | Salvianolic acid I | 41.24 ± 0.40 | [537]:493;359 | 2.13 ± 0.14 | 15.44 ± 1.01 | n.d. | n.d. | * |
| 27 | Quercetin-O-hexoside-hexuronide | 42.83 ± 0.33 | [639]:301 | n.q. | n.q. | n.q. | n.q. | |
| 28 | Apigenin-O-hexuronide | 43.59 ± 0.37 | [445]:269;175 | n.q. | n.q. | n.q. | n.q. | |
| 29 | Chrysoeriol-O-hexuronide | 44.79 ± 0.38 | [475]:299 | 1.05 ± 0.16 | 7.51 ± 1.13 | 0.43 ± 0.13 | 4.61 ± 1.35 | n.s |
| 30 | Salvianolic acid K isomer | 45.74 ± 0.64 | [555]:493;359 | n.q. | n.q. | n.q. | n.q. | |
| 31 | Luteolin | 52.99 ± 0.32 | [285]:241;217;199;75;151 | n.q. | n.q. | n.q. | n.q. | |
| Total phenolic compounds | 23.87 ± 1.48 | 173.23 ± 10.74 | 7.93 ± 2.52 | 85.17 ± 27.01 | * | |||
| Total flavonoids | 6.71 ± 0.87 | 48.71 ± 6.32 | 2.07 ± 0.76 | 22.30 ± 8.20 | * | |||
| Total phenolic acids | 17.16 ± 0.61 | 124.52 ± 4.42 | 5.86 ± 1.75 | 62.87 ± 18.81 | * | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taghouti, M.; Martins-Gomes, C.; Schäfer, J.; Santos, J.A.; Bunzel, M.; Nunes, F.M.; Silva, A.M. Chemical Characterization and Bioactivity of Extracts from Thymus mastichina: A Thymus with a Distinct Salvianolic Acid Composition. Antioxidants 2020, 9, 34. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9010034
Taghouti M, Martins-Gomes C, Schäfer J, Santos JA, Bunzel M, Nunes FM, Silva AM. Chemical Characterization and Bioactivity of Extracts from Thymus mastichina: A Thymus with a Distinct Salvianolic Acid Composition. Antioxidants. 2020; 9(1):34. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9010034
Chicago/Turabian StyleTaghouti, Meriem, Carlos Martins-Gomes, Judith Schäfer, João A. Santos, Mirko Bunzel, Fernando M. Nunes, and Amélia M Silva. 2020. "Chemical Characterization and Bioactivity of Extracts from Thymus mastichina: A Thymus with a Distinct Salvianolic Acid Composition" Antioxidants 9, no. 1: 34. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9010034