Antioxidants as an Epidermal Stem Cell Activator


1
Department of Dermatology, Kyung Hee University Hospital at Gang-dong, Kyung Hee University School of Medicine, Seoul 05278, Korea
2
Department of Dermatology, Seoul National University College of Medicine, Seoul National University Bundang Hospital, Gyeonggi 13620, Korea
*
Author to whom correspondence should be addressed.
Antioxidants 2020, 9(10), 958; https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9100958
Submission received: 1 September 2020
/
Revised: 5 October 2020
/
Accepted: 5 October 2020
/
Published: 7 October 2020
(This article belongs to the Special Issue Antioxidants and Skin Protection II)
Abstract
:Antioxidants may modulate the microenvironment of epidermal stem cells by reducing the production of reactive oxygen species or by regulating the expression of extracellular matrix protein. The extracellular membrane is an important component of the stem cell niche, and microRNAs regulate extracellular membrane-mediated basal keratinocyte proliferation. In this narrative review, we will discuss several antioxidants such as ascorbic acid, plant extracts, peptides and hyaluronic acid, and their effect on the epidermal stem cell niche and the proliferative potential of interfollicular epidermal stem cells in 3D skin equivalent models.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
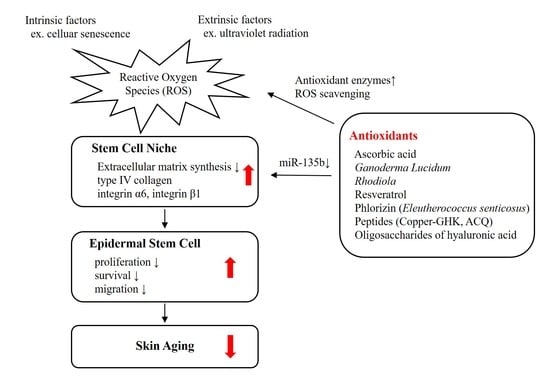
Epidermis, a multilayered tissue, is mainly composed of renewable keratinocytes. Basal keratinocytes proliferate and migrate into the upper layer of the epidermis. They undergo the process of differentiation thereafter in which their cuboidal shape becomes flatten. At the end of their fate, keratinocytes finally lose their nuclei and are eliminated from the skin. Epidermal stem cells or progenitor cells with high proliferative ability contribute to the maintenance of skin through their self-renewal, proliferation, and differentiation capacities [1]. The proliferation and survival of epidermal stem cells is influenced by numerous intrinsic and extrinsic factors [2]. A reduction in the function or number of stem cells may cause impaired self-renewal of the skin, thereby resulting in skin aging [3]. Senescence of keratinocytes by chronological aging or repetitive subculture causes a reduction in stem cell population in vitro [4]. The epidermal stem cells are located in a ‘niche’—specific sites that determine stem cell activity and homeostasis [5]. Stem cell niche provides a stem cell-specific microenvironment. As a result, it regulates the number and function of stem cells, and thus, it is important for the fate of stem cells [6].
Our team has studied the effect of antioxidants on the interfollicular epidermal stem cells [7,8,9,10,11,12]. Well-known antioxidants such as ascorbic acid, resveratrol and copper-GHK, as well as plant extracts that have been used for anti-fatigue or anti-stress treatment in traditional medicines have been evaluated for their role to activate the proliferative potential of epidermal stem cells. In this narrative review, we will discuss the importance of redox balance in the regulation of epidermal stem cells. In vitro and in vivo study results that demonstrate the antioxidant activities of the molecules such as ascorbic acid, plant extracts (Ganoderma lucidum, Rhodiola, resveratrol, Eleutherococcus senticosus), tripeptides, and hyaluronic acid will be overviewed. Experimental data supporting the role of the antioxidants on modulating stem cell microenvironment and stimulating the proliferative potential of epidermal stem cells will be reviewed. Finally, we will discuss the applicability of the antioxidants for anti-aging treatment of skin.
2. Epidermal Stem Cells and Extracellular Matrix
2.1. Interfollicular Epidermal Stem Cells
Three distinct epidermal stem cell niches have been demonstrated—bulge of hair follicle, base of sebaceous gland, and basal layer of interfollicular epidermis [13]. Contrary to hair follicular stem cells, little is known about markers for the interfollicular epidermal stem cells. p63 is a putative marker for the interfollicular epidermal stem cells that is an essential transcription factor for epidermal development and homeostasis [14,15,16]. It determines fate of the keratinocytes by regulating the balance between stemness, differentiation, and senescence [15]. However, it is not a specific marker, as differentiated keratinocytes in the upper epidermis also expresses p63. Thus, other markers, such as proliferating cell nuclear antigen (PCNA) that is involved in the cell cycle in proliferating cells, have been used to identify interfollicular epidermal stem cells [17]. Recently, we reported a new method to identify interfollicular epidermal cells by combining the expressions of p63 and histone deacetylase (HDAC) 1 [13].
2.2. Extracellular Matrix and Stem Cell Niche
Extracellular matrix (ECM) is an important component of the stem cell niche [18]. High levels of integrin have been proposed to be a marker for epidermal stem cells [19]. By expressing high levels of integrins, epidermal stem cells can interact with the ECM. Integrin α6 is a marker of extracellular adhesion receptors that exist along the dermo-epidermal junction [20]. Integrin β1 is expressed on the membrane of basal keratinocytes and mediates the cell-to-matrix and cell-to-cell interactions [21]. Both of these integrins are basement membrane proteins related to the stemness of the epidermis [22].
We previously reported that insulin-like growth factor-binding protein (IGFBP)-2 from hair follicle dermal sheath cells plays a critical role in regulating regenerative capacity of human epidermal keratinocytes [23]. Skin equivalents treated with IGFBP-2 showed thicker and more matured epidermis than control. The expressions of integrin α6 and β1 were upregulated when compared with control. IGFBPs form biologically active complexes with insulin growth factor (IGF)-1 and vitronectin [24,25]. Through the activation of both the IGF receptor and vitronectin-binding integrins, IGF-1 could protect keratinocytes against UVB damage [24]. Furthermore, activation of integrin β1 is involved in the inhibition of differentiation signals via extracellular signal-regulated kinases (ERK) signaling pathway [26]. These findings suggest integrins are important in regulating stem cell physiology and determining the fate of epidermal stem cells in skin [11].
Recently, we reported the relationship between ECM production and epidermal stem cell function via an epigenetic mechanism [27]. MicroRNA (miR135b), which regulates the synthesis of type IV collagen, the major component of cutaneous basement membrane, is involved in the maintenance of epidermal stem cell niche and proliferative potential of epidermal basal cells. Inhibition of miR135b increased the number of p63- and PCNA-positive cells in the epidermis in vitro via enhanced expression of type IV collagen.
2.3. Reactive Oxygen Species Regulates Stem Cell Niche
Since skin constitutes the outer surface of human body that is constantly exposed to environmental stimuli, the epidermal stem cells are regulated by not only the interaction with intrinsic transcriptional regulations but also extrinsic environmental signals [11]. These extrinsic factors could affect stem cell microenvironment, altering the fate of epidermal stem cells [28]. For example, hypoxia is required for the maintenance of undifferentiated stem cell and for controlling the proliferation and survival of stem cells [29]. Reactive oxygen species (ROS) are important in the stem cell development, function, and survival [30]. ROS enhance differentiation of human embryonic stem cells into mesendodermal lineage via ROS-involved signaling pathways [31]. Coenzyme Q10, a well-known antioxidant, has been shown to have protective effect against hypoxia-reperfusion induced damage in neural stem cells [32]. These findings indicate that redox status is critical in regulating stem cells.
ROS modulate the microenvironment surrounding epidermal stem cells, affecting their proliferative potency [7]. It has been shown that epidermal keratinocyte subpopulations with stem-cell like characteristics have lower levels of antioxidant proteins (superoxide dismutase (SOD), catalase, and glutathione peroxidase-1), and decreased activity of SOD and catalase compared with more differentiated keratinocytes [33]. They also exhibit less mitochondrial area, fewer peroxisomes, and lower levels of ROS production than differentiated keratinocytes. Furthermore, aging causes a reduction in the antioxidant capacity of epidermal keratinocytes as shown by decreased concentrations of α-tocopherol, ascorbic acid, and glutathione [34].
2.4. UV Radiation Affects Epidermal Stem Cells Via ROS Production
Chronic ultraviolet (UV) radiation is the main cause of extrinsic skin aging or photoaging. UVB (280–320 nm) and UVA (320–400 nm) radiation causes not only direct injury in molecules such as nucleic acids via cyclobutane pyrimidine dimers and pyrimidine (6–4) pyrimidine photoproducts formation, but also oxidative stress and the production of intracellular ROS [35,36,37,38]. ROS are primarily generated within the inner membrane of mitochondria [38]. Electrons leaked from the mitochondrial electron transport process react with oxygen to form free radicals [38]. UV-induced ROS react with lipids, proteins, and DNA, which leads to cellular apoptosis and gene mutations [39].
To protect against cellular damage from the UV-induced ROS, epidermal keratinocytes express several antioxidant enzymes, such as SOD, catalase, and glutathione peroxidase-1, that regulate the generation of intracellular ROS [33]. The endogenous antioxidant machinery in the epidermis has higher activity or concentration percentages compared with that in dermis: SOD (126%), glutathione peroxidase (61%), glutathione reductase (215%), glucose-6-phosphate dehydrogenase (111%), isocitrate dehydrogenase (313%), α-tocopherol (90%), ubiquinol 10 (900%), ascorbic acid (425%), uric acid (488%), reduced glutathione (513%), and total glutathione (471%) [40]. UVB, which is the most important environmental factor causing skin photoaging, down-regulates the antioxidant enzyme activity, induces the production intracellular ROS in the epidermis, and thus disrupts the cellular defense system [41,42]. As a result, elevated levels of intracellular oxidized proteins are observed in aged skin [43].
3. Antioxidants and Interfollicular Epidermal Stem Cells
3.1. Skin Equivalents
The effect of the antioxidants on the production of ECM and epidermal stem cells could be evaluated using a 3D skin equivalent model rather than two-dimensional keratinocyte cultures to investigative the interaction between interfollicular epidermal stem cell and ECM. The process of constructing skin equivalents is described in our previous study [44]. Dermal substitutes were produced by mixing type I collagen from the tendons of rattails, 10× concentrated DMEM, and neutralization buffer (0.05 N NaOH, 0.26 mM NaHCO3, and 200 mM HEPES), and adding 5 × 105 human fibroblasts. After gelling in a 30 mm polycarbonate filter chamber, 1 × 106 human keratinocytes were seeded onto the dermal substitute. After 24h of submerged culture, they were cultured at the air–liquid interface for twelve days. The growth medium consisted of DMEM and Ham’s nutrient mixture F12 at a ratio of 3:1, 5% FBS, 0.4 μg/mL hydrocortisone, 1 μM isoproterenol, 25 μg/mL ascorbic acid, and 5 μg/mL insulin. EGF with a concentration of 1 ng/mL was added during the submerged culture, and EGF with 10 ng/mL was added during the air–liquid interface culture.
3.2. Ascorbic Acid
3.2.1. Antioxidant Activity
Ascorbic acid is an essential human dietary requirement for the prevention of scurvy. Scurvy is characterized by hemorrhages, wound dehiscence, poor wound healing, and loosening of teeth [45]. Symptoms of scurvy indicate that ascorbic acid is involved in the synthesis of connective tissue. In fact, ascorbic acid catalyzes posttranslational modification of procollagen to produce mature collagen [46]. Hydrolases that convert prolyl and lysyl residues of procollagen to produce stable triple helical structure of collagen require ascorbic acid as a cofactor [46].
Ascorbic acid is a well-known antioxidant that reduces intracellular ROS production. It acts as an electron donor to reduce the production of ROS that is generated during metabolic respiration or mitochondrial oxidative phosphorylation [45]. As a result, ascorbic acid protects cells from oxidative DNA damage, lipid peroxidation, and the oxidation of amino acid residues [47]. Several studies have reported that ascorbic acid decreases UVB-induced ROS production, and prevents ROS-medicated DNA damage and apoptosis in epidermal keratinocytes [48,49]. Pre- and post-treatment of reconstituted human epidermis with ascorbic acid reduced UVB-induced cell death, apoptosis, DNA damage, ROS production, and the inflammatory response by suppressing tumor necrosis factor-α (TNF-α) production [50].
3.2.2. Epigenetic Regulation
In addition to its role as an antioxidant, ascorbic acid also serves as an epigenetic regulator of histone and DNA methylation. Ascorbic acid acts as a cofactor for Fe2+ and α-ketoglutarate-dependent dioxygenases (Fe2+/ α-KGDDs) by transferring electron to reduce ferric iron (Fe3+) to ferrous iron (Fe2+) [51]. Diverse families of Fe2+/ α-KGDDs include collagen prolyl hydroxylases, and epigenetic regulators such as the Jumonji C domain-containing histone demethylases (JHDMs), DNA and RNA demethylases of the AlkB homolog family, and the ten-eleven translocation (TET) family of DNA hydroxylases [52,53,54,55,56,57]. By upregulating the activity of JHDMs or TET DNA hydroxylases, ascorbic acid regulates the embryonic stem cell function and promotes reprogramming of fibroblasts to induced pluripotent stem cells (iPSC) [51]. Ascorbic acid was shown to maintain the proliferation of embryonic stem cell and human mesenchymal stem cells in vitro [58,59]. It enhances the activity of JHDM1a/1b to promote demethylation of H3K36me2/3 in mouse embryonic fibroblasts during reprogramming [53]. Ascorbic acid can also induce a specific loss of H3K9me2, which is a barrier during somatic cell reprogramming into iPSCs, in the embryonic stem cells, and drive the transition of pre-iPSC to iPSC by activating JHDMs [58]. Ascorbic acid enhances iPSC reprogramming and DNA demethylation in mouse and human fibroblasts via TET-dependent production of 5-hydroxymethylcystosine [60,61].
3.2.3. Epidermal Stem Cell Activation
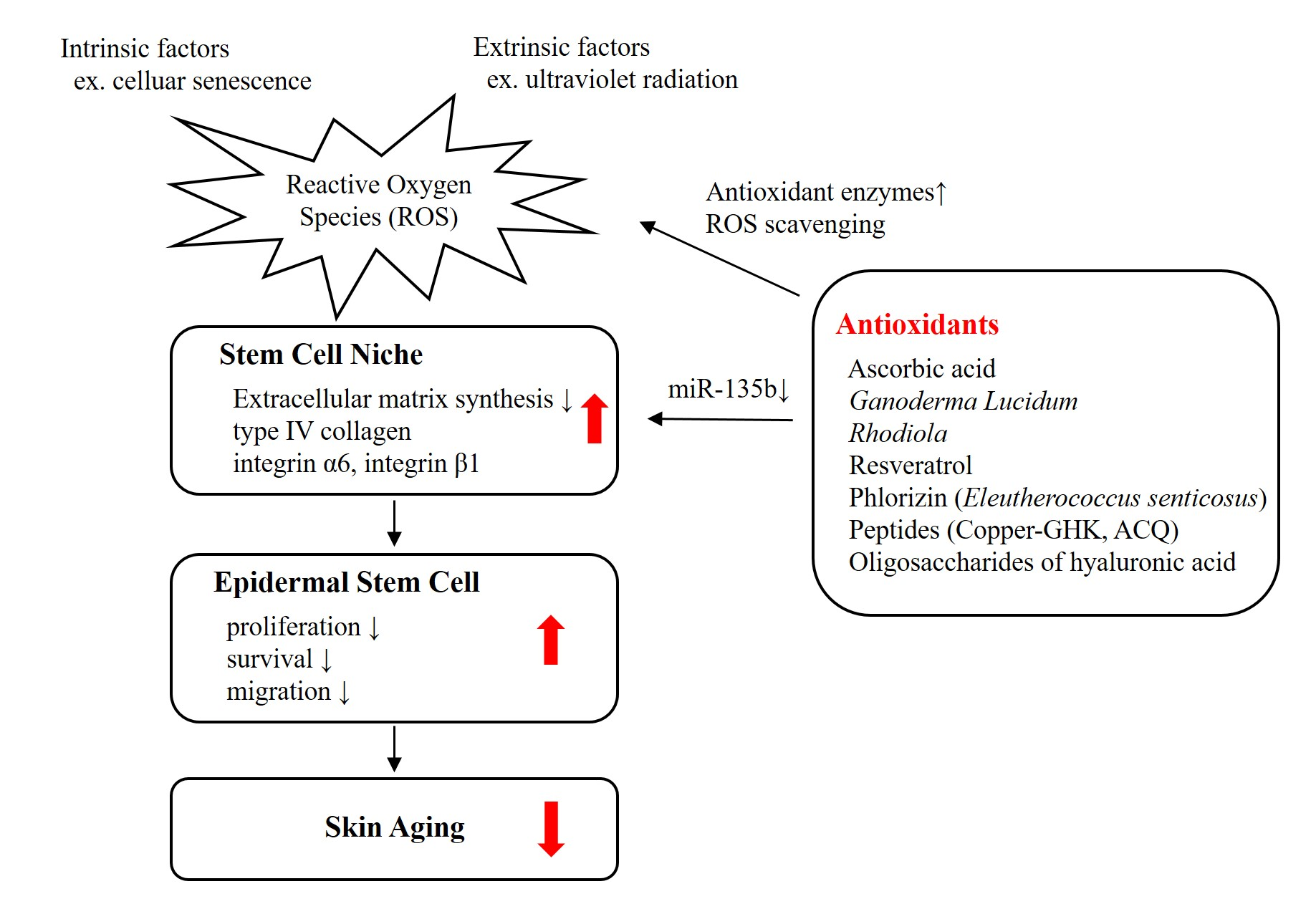
Ascorbic acid enhances the stemness of corneal epithelial stem cells of mouse by promoting ECM production, and thus accelerates epithelial wound healing in the cornea [62]. An in vitro study revealed that ascorbic acid promotes the proliferation of keratinocytes and fibroblasts from skin [63]. Due to its antioxidant activity and role as a stem cell regulator, we investigated the effect of ascorbic acid on epidermal stem cells [7]. Ascorbic acid showed good antioxidant activity at both low (10 μg/mL) and high concentrations (100 μg/mL) in vitro. More cuboid-shaped basal cells that have high proliferative potential were observed in the skin equivalents treated with ascorbic acid compared with control. The expression of integrins α6 and β1 along the basement membrane was upregulated in the ascorbic acid-treated skin equivalents. (Figure 1) The number of p63- and PCNA-positive cells was increased in skin equivalents treated with ascorbic acid. Thus, by altering the integrin expression, ascorbic acid provides favorable stem cell microenvironment in the skin, increasing the stemness and proliferative potential of epidermal basal cells.
3.3. Ganodermal Lucidum
3.3.1. Antioxidant Activity
Ganoderma lucidum is a highly nutritional mushroom that has been used in traditional medicines. It is also known as Lingzhi mushroom in China and Reish in Japan. G. lucidum has drawn attention as a natural compound with diverse bioactivities, including antioxidant, immunomodulatory, anti-allergic, anti-inflammatory, anti-bacterial, anti-fungal, anti-viral, anti-tumor, and anti-melanogenic activities [64]. Effective components of G. lucidum are polyscaccharides glycoproteins, phenols, and protein substances, which provide its antioxidant activity [65].
G. lucidum polysaccharides, a major component of G. lucidum, has a wide spectrum of pharmacologic properties, including antioxidant, anti-aging, anti-tumor, anti-inflammatory, anti-depressant, and immunomodulatory activities [66,67]. G. lucidum polysaccharides exhibits antioxidant activity by scavenging ROS and enhancing enzymatic activity of glutathione peroxidase [68,69]. Injection of G. lucidum polysaccharides into mice resulted in the decreased low-density lipoprotein (LDL) oxidation products, reduced level of malondialdehyde, and increased glutathione peroxidase activity in serum and heart [68]. G. lucidum polysaccharides also scavenged tert-butyl hydroperoxide-induced free radicals in macrophages in vitro and in vivo [69]. By increasing antioxidant enzyme activities and lowering malondialdehyde levels, G. lucidum polysaccharides protects murine skeletal muscles from oxidative stress induced by exhaustive exercise [70]. In addition, G. lucidum polysaccharides protects brain and renal tissue against ischemia-reperfusion by suppressing oxidative stress and cell apoptosis [71,72,73]. Another Ganoderma protein, Ganoderma glycopeptides, also exhibits antioxidant property. Ganoderma glycoprotein increased the activity of SOD and glutathione peroxidase in animal serum, hippocampus, and myocardium, resulting in the reduced level of malondialdehyde [65].
G. lucidum polysaccharides have been widely used in wound healing and anti-aging skin care products [67]. G. lucidum polysaccharides attenuated skin flap ischemia-reperfusion injury via thioredoxin-1-dependent antioxidant and anti-apoptotic pathway [74]. The administration of G. lucidum polysaccharide into mice skin flap undergoing ischemia-reperfusion injury exhibited increased SOD activity, reduced level of malondialdehyde, and subsequent reduction of the apoptotic keratinocytes and fibroblasts. While the depleted level of thiorexin-1, an endogenous redox signaling regulator, was observed in ischemia-reperfusion flaps, administration of G. lucidum polysaccharides restored the level of thiorexin-1 and inhibited subsequent apoptosis signal-regulating kinase 1 (ASK-1)–mitogen-activated protein kinase (MAPK) signaling pathway. G. lucidum polysaccharides also protect fibroblasts against UVB-induced photoaging by reducing the ROS levels [75]. UVB-irradiated fibroblasts treated with G. lucidum polysaccharides showed increased cell viability, decreased number of β-galactosidase-positive fibroblasts, increased expression of c-telopeptides of type I collagen, decreased matrix metalloproteinase (MMP)-1 expression, and reduced level of intracellular ROS compared with the control.
3.3.2. Epidermal Stem Cell Activation
We investigated the antioxidant activity of G. lucidum and its effect on epidermal stem cells [7]. At high concentration (100 μg/mL), G. lucidum showed a good antioxidant activity, comparable to ascorbic acid without keratinocyte toxicity. Skin equivalents treated with G. lucidum had cuboidal epidermal basal cells with higher proliferative potential compared with control. The expression of integrin α6 and β1 along the basement membrane was upregulated in the G. lucidum-treated skin equivalents. (Figure 1) The number of p63- and PCNA-positive cells was increased in skin equivalents treated with G. lucidum. Thus, by altering the integrin expression, the antioxidant G. lucidum provides favorable stem cell niche in the skin, increasing the stemness and proliferative potential of epidermal basal cells.
3.4. Rhodiola
3.4.1. Antioxidant Activity
The genus Rhodiola consists of more than 200 species. Of them, approximately 20 species, including R. rosea, R. sachalinensis, R. crenulata, R. sacra, R. alterna, R. brevipetiolata, R. kirilowi, and R. quadrifida have been used in traditional medicines for anti-fatigue, anti-depressant, and anti-inflammatory drugs in Asia [76]. R. rosea, the most extensively studied species among the Rhodiola plants, has pharmacological effects such as elongation of lifespan, stimulation of central nervous system, elevation of work performance, protection on cardiovascular system, nervous system and liver, and increased resistance against bacterial and virus [76]. Rhodiola contains a high percentage—up to 41.4%—of polyphenols, such as flavonoids, proanthocyanidines, tyrosol, and cinnamyl alcohol, which might provide its antioxidant property [77,78]. Major components of Rhodiola plants are rosavin (rosarian, rhodionin, rhodiosin, and rosin), cinnamyl alcohol, salidroside, and tyrosol [76]. Of them, salidroside and its aglycone, tyrosol, are the most important compounds of Rhodiola, and the quality of crude drugs of Rhodiola depends on the amount of these two compounds [76].
R. imbricate extract exhibited antioxidant activity and reduced lipid peroxide levels in the rat excision wound model [79]. R. imbricate extract accelerated the process of wound healing as shown by increased rate of wound contraction, decreased time taken for epithelialization, and increased cellular proliferation and collagen synthesis compared with control. R. rosea extract was shown to have a high potential for scavenging singlet oxygen and hydrogen peroxide, reducing ferric iron (Fe3+), chelating ferrous iron (Fe2+) and protecting protein thiol [78]. In cultured human keratinocytes exposed to oxidative stress, R. rosea root extract upregulated the cellular antioxidant processes, such as SOD and catalase activities, while it downregulated the glutathione levels, glyceraldehyde-3-phosphate dehydrogenase activity, and thiobarbituric acid reactive substances levels [80]. Oligomeric proanthocyanin from R. rosea enhanced the activities of SOD and glutathione peroxidase, and lowered malondialdehyde level in serum, heart, liver and brain tissues in mice [81]. R. crenulata root extract protected human skin and human keratinocytes cell line (HaCaT) cells from ionizing radiation by attenuating oxidative stress, cell apoptosis, and MMP levels [82].
3.4.2. Epidermal Stem Cell Activation
We investigated the antioxidant activity of R. sachalinensis and its effect on epidermal stem cells [7]. At a high concentration (100 μg/mL), R. sachalinensis showed an antioxidant activity comparable to ascorbic acid without keratinocyte toxicity. Skin equivalents treated with R. sachalinensis showed increased stemness and proliferative potential, and upregulated expression of integrin α6 and β1 along the basement membrane. (Figure 1) The number of p63- and PCNA-positive cells was increased in skin equivalents treated with R. sachalinensis. Therefore, R. sachalinensis upregulates the expression of integrins and provides the favorable stem cell niche for epidermal basal cells proliferation.
3.5. Resveratrol
3.5.1. Antioxidant Activity
Resveratrol (3,5,4′-trihydroxy-trans-stilbene) is a natural polyphenol found in grapes, red wine, and berries [8]. It has been extensively explored in the last decades for a potential therapeutic role against aging as well as in various diseases, including cancers and cardiovascular diseases [83]. Resveratrol has various mechanisms of action including (1) reduction of intracellular ROS, (2) activation of sirtuin 1 (SIRT1), a histone deacetylase that increases DNA stability, prolongs survival in mammals, and provides cellular protection against UV radiation through p53 and c-Jun n-terminal kinase (JNK) pathway modulation, (3) significant cancer chemopreventive potential. Resveratrol could also exert its effect via [3H]-resveratrol specific binding sites in HaCaT cells [84]. Extensive researches on resveratrol have been carried out in the fields of cosmetology and dermatology to demonstrate its anti-melanogenic and antioxidant properties [8].
It has been shown to directly scavenge hydroxyl radical, nitric oxide, superoxide anion radial, and peroxynitrite [85]. Previously, we have also reported ROS scavenging activity of resveratrol by DPPH assay and normal human melanocytes treated with hydrogen peroxide [86]. Resveratrol lowered intracellular level of ROS in human skin fibroblast exposed to hydrogen peroxide in a dose-dependent manner [87]. Human keratinocytes exposed to particulate matter also showed reduced production of ROS in a dose-dependent manner when treated with resveratrol [88]. Beyond its direct antioxidant properties, resveratrol also activates the endogenous antioxidant pathways via the upregulation of the nuclear factor erythroid 2-related factor (Nrf2) [89]. Resveratrol induces the phosphorylation of Nrf2 via the phosphoinositide 3-kinases/Akt (protein kinase B) pathway and releases Nrf2 from the Kelch-like ECH-associated protein (Keap1) [85]. The nuclear translocation of Nrf2 activates the antioxidant response element (ARE), inducing endogenous antioxidant enzymes, such as NADPH quinone dehydrogenase-1 and glutathione peroxidase-2 in normal human keratinocytes [89].
Resveratrol decreased apoptosis of HaCaT cells exposed to sodium nitroprusside that served as a donor of nitric oxide free radical [84]. Resveratrol suppresses the overproduction of nitric oxide, which is relevant to premature skin aging occurring upon chronic UV exposure [90]. Resveratrol protects keratinocytes against the UV- or cigarette smoke-induced oxidative damages [91,92]. In cultured keratinocytes, activation of SIRT1 by resveratrol suppresses UVB- and hydrogen peroxide-induced cell death via inhibition of ROS-mediated JNK activation and p53 acetylation [93]. SIRT1 is also involved in the activation of AMP-activated protein kinase (AMPK) [93]. Activation of the AMPK-forkhead box O (FOXO) 3a cascade by resveratrol prevents keratinocytes from oxidative stress-induced senescence and proliferative dysfunction [94].
3.5.2. Epidermal Stem Cell Activation
We demonstrated that the antioxidant activity of resveratrol provides favorable stem cell niche in the skin and increases the stemness and the proliferative potential of interfollicular epidermal stem cells [8]. In skin equivalents treated with resveratrol, the expression of integrin α6 was upregulated and the number of p63-positive cells was increased.
3.6. Phlorizin (Active Ingredient of Eleutherococcus Senticosus)
3.6.1. Antioxidant Activity
Eleutherococcus senticosus (Acanthopanax senticosus, also known as Siberian ginseng) is a widely used traditional Chinese herb has been used as an anti-fatigue agent [95]. It grows in Eastern region of Russia and East Asia, including China, Korea, and Japan. Several pharmacological studies on E. senticosus has demonstrated its anti-stress, anti-ulcer, anti-irradiation, anti-cancer, anti-inflammatory, and hepatoprotective activities [95]. The leaves of E. senticosus inhibit glycosidase to exhibit anti-bacterial properties [96]. Several chemical compounds consisting E. senticosus have been reported, including triterpenoid saponins, lignans, coumarins, and flavonoids, among which phenolic compounds are considered to be the most active components [95]. Elutheroside B and E are major active lignands of E. senticosus, which have immunomodulatory, antioxidant, and anti-inflammatory activities [95]. Other phenolic compounds such as caffeic acid and chlorogenic acid also showed strong antioxidant activity in vitro and in vivo [97,98]. Water-soluble polysaccharide fractions from E. senticosus had a protective effect against oxidative damage exhibiting scavenging activities for hydroxyl radical, superoxide radicals, and DPPH radicals [99]. Extracts of E. senticosus showed protective activity against tert-butyl hydroperoxide-induced oxidative stress in rat hepatocytes [100]. E. senticosus extracts induced the expression of antioxidant enzymes through the activation of Nrf2, a critical transcription factor that binds to the ARE encoding antioxidant enzymes.
Phlorizin (phloretin-2′-O-glucoside, various named phloridzin, phlorhizin, phlorrhizin), the main ingredient of E. senticosus, belongs to the chemical class of dihydrochalcones [101]. Chalcones (1,3-diaryl-2-propen-1-ones) are flavonoids which display a broad spectrum of bioactivities such as anti-cancer, anti-fungal, anti-bacterial, anti-viral, and anti-inflammatory properties [102]. Dihydrochalcones without α-β double bond comprise phlorectin and its glucoside, phlorizin [103]. Phlorizin is also abundantly present in apples. It has drawn attentions for its pharmacological action to induce renal glycosuria and inhibit intestinal glucose absorption through sodium-linked glucose transporters (SLGTs) [104].
Beyond its anti-diabetic activity, the antioxidant activity of phlorizin and its derivatives has been also investigated in vitro [101,105,106]. The potent antioxidant activity of phloretin was shown by peroxynitrite scavenging and inhibition of lipid peroxidation [103]. Phlorizin was shown to prevent the skin from UVB-induced damage in vitro and in vivo by suppressing ROS overproduction, cyclooxygenase-2 (COX-2) expression, and subsequent inflammatory reactions [107]. Inhibition of p38 and JNK-MAPK signaling pathways are involved in the protective effect of phlorizin [107].
3.6.2. Epidermal Stem Cell Activation
Recently, we reported the antioxidant property of E. senticosus extracts and phlorizin and its effect on skin equivalents [9]. Skin equivalents treated with 0.002% of E. senticosus extracts showed slightly thickener epidermis than control. The expression of p63, PCNA, and integrin α6 was upregulated in skin equivalents treated with E. senticosus extracts. Extraction and chromatography indicated that phlorizin is an active ingredient of E. senticosus extracts. Compared with ascorbic acid, phlorizin had a relatively weak antioxidant activity at low concentrations (1 mM), but equivalent antioxidant activity at high concentrations (5–10 mM). Skin equivalents treated with phlorizin showed increased thickness of the epidermis, an increased staining intensity for p63, PCNA, integrin α6, and integrin β1. (Figure 2) Linear staining of type IV collagen, a major component of basement membrane, increased in a dose-dependent manner in the phlorizin-treated skin equivalents. We also revealed that phlorizin downregulated the level of miR135b, which is involved in the proliferative potential of epidermal stem cells by regulating the type IV collagen synthesis [27]. In conclusion, E. senticosus extracts and their active ingredient, phlorizin, produce a favorable stem cell niche for the epidermal stem cell survival and proliferation via regulating ECM proteins or transmembrane proteins.
3.7. Human Tripeptide “Copper-GHK”
3.7.1. Antioxidant Activity
Peptides are amino acids chains that are involved in various cellular processes [108]. The human tripeptide glycyl-L-histidyl-L-lysine (GHK) is an extracellular matrix-derived peptide with a high affinity for copper that forms the copper-GHK complex [109]. In the copper-GHK complex, Cu2+ ion binds to the nitrogen from the imidazole side chain of histidine, another nitrogen from the α-amino group of glycine, the deprotonated amide nitrogen of the glycine-histidine peptide bond, and the oxygen from the carboxyl group of the lysine from the neighboring complex [109,110]. The concentration of GHK is associated with age, which is present at 200 mg/L in age of 20–25, and at 80 mg/L by the age of 60–80 [109]. Isolated from human plasma in 1973, it was shown to promote the proliferation of hepatoma cells and the survival of normal hepatocytes by providing copper that is necessary for the cellular functions [111]. Subsequent research has found its beneficial actions in skin, nervous tissue, intestine, bone, and blood vessels [109]. It regulates the wound healing and tissue remodeling process via its antioxidant, anti-inflammatory, and regenerative properties [112].
Since the activity of endogenous antioxidants such as SOD1 depends on copper ion, copper deficiency could result in reduced antioxidant enzyme activity and increased oxidative tissue damage [113,114]. Copper-GHK upregulates endogenous antioxidants, supposedly by supplying copper necessary for its function [115]. In rats treated with copper-GHK, the level of SOD and catalase was upregulated during the skin wound healing process [115]. The level of hydroxyl and peroxyl radicals was diminished in the cells treated with copper-GHK [116]. Copper-GHK also reduces oxidative tissue damage by inhibiting iron release and lipid peroxidation in damaged tissues [117]. The presence of iron complexes in damaged tissues increases lipid peroxidation, and thus delays wound healing. Copper-GHK inhibits the release of ferritin iron, probably by binding to the ferritin channels involved in the iron release [117].
3.7.2. Regulation of Gene Expressions
In addition to its antioxidant activity, copper-GHK could regulate a large number of human genes. Copper-GHK reduced the transcription of RNA of 54 genes overexpressed in patients with aggressive colorectal cancer. [118]. It also regulates human genes such as SIGMAR1, EPM2A, NAIP, FGFR2, and SLIT2 that are involved in the development and maintenance of nervous system [109].
3.7.3. Regulation of ECM Production
Several studies have suggested copper-GHK as a potential activator of wound repair. Copper-GHK upregulated the synthesis of type I collagen in fibroblast cultures [119]. In an in vivo study using a rat experimental wound, the injection of copper-GHK stimulated collagen synthesis and the expression of type I and III collagen mRNAs [120]. Glycosaminoglycans (dermatan sulfate, chondroitin sulfate) and proteoglycan (decorin), which interact with collagen to organize and strengthen the fibrillary network, were also increased in vitro and in vivo after treated with copper-GHK [120,121,122]. Glycosaminoglycans and proteoglycans are also involved in several wound healing processes, including cell adhesion, migration, and proliferation [123]. Furthermore, copper-GHK alters the expression of MMPs, extracellular proteinases involved in the degradation of ECM required to eliminate damaged and provisional tissue, and to promote new vessel formation and cell migration, during the wound healing process [124]. Several types of MMPs are selectively expressed when wound healing occurs [124]. In dermal fibroblast cultures incubated with copper-GHK, the expression of MMP-2 and tissue inhibitor of metalloproteinase (TIMP)-1 and 2 was increased [125]. Copper-GHK injection induced the prolonged expression of MMP-9 and the higher level of MMP-2 than control, which resulted in the enhanced wound repair in vivo [124]. Taken together, copper-GHK not only stimulates the production of connective tissue but also regulates the remodeling of ECM by modulating MMPs and TIMPs during the wound healing process that requires a rapid ECM turnover.
3.7.4. Epidermal Stem Cell Activation
We further investigated the effect of copper-GHK on basal keratinocytes [10]. In the monolayer keratinocyte culture, copper-GHK increased the keratinocytes proliferation in a dose-dependent manner in the skin equivalent model. Skin equivalents showed cuboidal-shaped epidermal basal cells with the addition of copper-GHK. Immunohistochemical analysis revealed increased number of PCNA- and p63-positive basal cells. The expression of integrin α6 and β1 was upregulated to a greater degree in the copper-GHK-treated skin equivalents than in the control. Therefore, copper-GHK may increase the survival and proliferation of interfollicular epidermal stem cells by regulating the expression of ECM proteins such as integrin α6 and β1.
3.8. Tripeptide “ACQ: Alanine-Cysteine-Glutamine”
Glutathione is a major endogenous antioxidant within the cells, not only by directly neutralizing free radicals and ROS, but also by inducing the active forms of exogenous antioxidants such as vitamins C and E [126]. Glutathione is a tripeptide with a gamma peptide linkage between the carboxyl group of the glutamate sidechain and the amine group of cysteine, of which cysteine moiety provides the antioxidant property of glutathione [126].
In an effort to find a novel short bioactive antioxidant peptide that allows transcutaneous delivery, we screened peptide library that contains cysteine. We found a novel tripeptide alanine-cysteine-glutamine (ACQ) that has more significant antioxidant property compared with that of glutathione [11]. ACQ protected fibroblasts and keratinocytes from the hydrogen peroxide-induced oxidative stress. The skin protective effect of ACQ against UV irradiation was demonstrated in BALB/c nude mice fed with 0.1% ACQ solution for 2 or 4 weeks, which had thick epidermis and rare apoptotic cells after UV irradiation. DCF staining revealed lower free radical status and higher number of p63-positive cells in the skin of ACQ fed mice than in the skin of control mice. Skin equivalents treated with ACQ showed increase in the epidermal thickness in dose-dependent manner. Confocal microscopic examination revealed increased number of p63-positive cells and upregulated expression of integrin α6 in the ACQ-treated skin equivalents. (Figure 3) Therefore, ACQ is a good antioxidant tripeptide that increases proliferative potential of the epidermal stem cells via upregulation of integrin α6.
3.9. Hyaluronic Acid
3.9.1. Antioxidant Activity
Hyaluronic acid is a natural polysaccharide that is consisted of glucuronic acid and n-acetylglucosamine repeats via β-1, 4 linkage. Repetitive disaccharide units of n-acetyl-d-glucosamine and d-glucuronic acid linked by β (1,4) and β (1,3) glycosidic linkages, respectively, constitute the linear structure of hyaluronic acid [127]. Hyaluronic acid is an essential constituent of ECMs in most mature tissues of vertebrates. The largest quantity of hyaluronic acid appears in skin tissue (7–8 g per average adult human) [127]. Due to its biophysical characteristics including biodegradability, biocompatibility, non-toxicity, and non-immunogenicity, hyaluronic acid has been used in biomedical fields such as osteoarthritis surgery, plastic surgery, ophthalmologic surgery, and tissue engineering [127].
Several reports have demonstrated the antioxidant properties of hyaluronic acid. In a rat model of carbon tetrachloride-induced liver injury, injection of hyaluronic acid reduced serum alanine aminotransferase (ALT) and aspartate aminotransferase (AST) rise and lipid peroxidation, and increased SOD and glutathione peroxidase activities [128]. High molecular weight hyaluronic acid (HMWHA) with average molecular weight of 5400 kDa and 2000 kDa attenuated DNA damage in leukocytes during oxidative burst via reducing intracellular level of reactive oxidants [129]. The mechanisms for ROS reduction of HMWHA are not completely understood. Hydroxyl functional groups in HMWHA could possibly absorb ROS [130]. HMWHA could interact with the CD44 receptor to activate pathways that regulates cellular redox status and intracellular ROS generation [131]. Endocytosed polyanionic hyaluronic acid molecules through the binding to the CD44 receptor of monocytes and granulocytes chelate Fe2+ and Cu2+ anions to suppress the generation of hydroxyl radical [129].
It was also reported that low molecular weight hyaluronic acid (LMWHA, 200–230 kDa) that is able to penetrate the skin prevents ROS damage in granulation tissue and promotes incisional wound healing in a rat experimental model [132,133]. Irradiation of gamma rays to the native hyaluronic acid increased its antioxidant activity by reducing the molecular weight [134]. Two LMWHAs, LMWHA-1 (145 kDa) and LMWHA-2 (45.2 kDa), inhibit lipid peroxidation and scavenge hydroxyl radical 1,1-diphenyl-2-picryldydrazyl radical superoxide anion in vitro [135]. Its antioxidant and free radical scavenging properties were superior to that of the native hyaluronic acid of 1050 kDA. Administration of LMWHA increased the activity of SOD, catalase, glutathione peroxidase, and total antioxidant capacity in cyclophosphamide-induced immunosuppressed mice [135]. However, the mechanism of antioxidant effect of LMWHA needs further study.
3.9.2. Epidermal Stem Cell Activation
We investigated the effect of oligosaccharides of hyaluronic acid (400–2000 Da) on epidermal stem cells [12]. Skin equivalents showed increased epidermal thickness in addition of oligosaccharides of hyaluronic acid. Contrary to the control, oligosaccharides of hyaluronic acid-treated skin equivalents showed positive staining for filaggrin in the granular layer. Filaggrin, which is expressed in the granular layer of epidermis during terminal differentiation, is a late marker of keratinocyte differentiation [136]. Thus, the result indicates that oligosaccharides of hyaluronic acid promotes differentiation as well as proliferation of epidermal keratinocytes. The number of p63-positive cells was greater in the basal layer of oligosaccharides of hyaluronic acid-treated skin equivalents than control, indicating the increased stemness of epidermal stem cells. The staining intensity of integrin α6 and β1 along the dermo-epidermal junction was significantly greater than that of control. Our findings indicate that oligosaccharides of hyaluronic acid affect the survival and differentiation of epidermal stem cells probably by altering the expression of ECM proteins such as integrins α6 and β1.
4. Application in Skin Rejuvenation
For the application of the antioxidants in the field of cosmetology and dermatology, further studies may be necessary to determine the proper concentration of the antioxidants in cosmetic products without safety problem. Though it is widely accepted that antioxidants play a role in preventing skin cancer, the risk of carcinogenesis must be strictly evaluated before the application of the antioxidants [137,138]. Searching for optimized formulation or delivery system is also important to enhance the permeation of the ingredients into skin. For example, ascorbic acid has limited application in the cosmetic field due to the unstable structure and low skin penetration [139]. The chemical modification of the hydroxyl group at the C-2 position of ascorbic acid produced stable derivatives such as 2-O-α-D glucopyranosyl-L-ascorbic acid (ascorbic acid 2-glucoside) [140,141]. Conjugated form of palmitoyl-KVK and ascorbic acid (palmitoyl-KVK-L-ascorbic acid) to enhance stability and skin penetration of ascorbic acid not only increased type I collagen synthesis in dermal fibroblasts but also improved facial wrinkles and pigmentation in vivo [142,143]. Ascorbic acid encapsulated into negatively charged liposomes exhibited increased stability, skin permeation [139]. Type I collagen production by fibroblasts and regeneration of UVA-induced keratinocyte damage were demonstrated in vitro.
Similarly, the natural form of resveratrol is chemically unstable to be used in cosmetic formulations [144]. As an approach to enhance stability of resveratrol in cosmetic formulations, resveratrol was acetylated to resveratryl triacetate as a prodrug form [145]. Resveratryl triacetate, which is converted into resveratrol through the action of endogenous esterases, has higher stability in solutions, lower cytotoxicity, and comparable anti-melanogenic effect in cultured melanocytes when compared with resveratrol [145]. The efficacy and safety of 0.4% resveratryl triacetate cream in treating hyperpigmented spots was also demonstrated in a clinical trial [146]. The 0.4% resveratrol triglycolate cream, another derivative of resveratrol esterified with glycolic acid, also showed efficacy in reducing hyperpigmentation in human skin without adverse skin reactions [144].
We previously reported clinical application of the antioxidant solution containing copper-GHK, oligosaccharides of hyaluronic acid, and Rhodiola extract (GHR formulation) using hydroporation technique in the treatment of periorbital fine wrinkles and melasma [147,148,149]. Hydroporation technique utilizes subsonic flow of air and the microdroplets of the solution into the skin to exfoliate the epidermis gently and to deliver the cosmetic solution into the skin. Weekly treatments for 8 to 12 weeks improved features of aged skin, such as fine wrinkles, pigmentation, and increased blood vessels. Immunhistochemical study revealed the increased staining intensity for type IV collagen and integrin α6 in the dermo-epidermal junction and p63-positive epidermal stem cells, which was consistent with the result of the in vitro studies using skin equivalents [147].
5. Conclusions
Identification of new biological mechanisms associated with skin aging enables the discovery of new active ingredients for cosmetics or cosmeceuticals that may recover biological functions affected by aging [150]. One point of view on skin aging is the dysfunction of epidermal stem cells that regulate self-renewal, proliferation, and differentiation of skin. Stem cell-specific microenvironment, which is regulated by intrinsic and extrinsic signals, could determine the function and survival of the stem cells. Redox status is a critical extrinsic signal that affect the microenvironment surrounding epidermal stem cells. By regulating the expression of ECM, the key component of the stem cell niche, ROS affects the proliferative potential of epidermal basal cells. Radiation of UVB, which is the most important environmental factor causing skin photoaging, also reduces antioxidant enzyme activity, increases intracellular ROS levels in the epidermis, and may contribute to the alteration in the epidermal stem cell function.
This work addresses the issues of oxidative stress associated with skin aging and provides a list of active ingredients to reverse the dysfunction of epidermal stem cells. We overviewed several antioxidants such as ascorbic acid, plant extracts, peptides, and hyaluronic acid that have ROS scavenging activities and protect cells from oxidative stress-induced damages in vitro or in vivo studies. Using the 3D skin equivalent models, we have demonstrated that antioxidants serve as an activator of epidermal stem cells, which leads to increased survival and proliferation of the stem cells. Antioxidants may modulate the microenvironment of epidermal stem cells by altering the production of ECM. Epigenetic mechanisms such as miR135b are involved in the ECM-mediated regulation of basal keratinocyte proliferation.
Author Contributions
Conceptualization, S.-H.K. and K.-C.P.; investigation, S.-H.K.; data curation, S.-H.K.; writing—original draft preparation, S.-H.K.; writing—review and editing, K.-C.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ALT | serum alanine aminotransferase |
| AMPK | AMP-activated protein kinase |
| ASK-1 | apoptosis signal-regulating kinase 1 |
| AST | aspartate aminotransferase |
| ARE | antioxidant response element |
| COX-2 | cyclooxygenase-2 |
| ECM | extracellular matrix |
| ERK | extracellular signal-regulated kinases |
| Fe2+/α-KGDDs | Fe2+- and α-ketoglutarate-dependent dioxygenases |
| FOXO | forkhead box O |
| HaCaT | human skin and human keratinocytes cell line |
| HDAC | histone deacetylase |
| HMWHA | high molecular weight hyaluronic acid |
| iPSC | induced pluripotent stem cell |
| JHDMs | Jumonji C domain-containing histone demethylases |
| JNK | c-Jun n-terminal kinase |
| Keap1 | Kelch-like ECH-associated protein |
| LMWHa | low molecular weight hyaluronic acid |
| MAPK | mitogen-activated protein kinase |
| MMP | matrix metalloproteinase |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| Nrf2 | nuclear factor erythroid 2-related factor |
| PCNA | proliferating cell nuclear antigen |
| ROS | reactive oxygen species |
| SIRT1 | sirtuin 1 |
| SLGT | sodium-linked glucose transporter |
| SOD | superoxide dismutase |
| TET | ten-eleven translocation |
| TIMP | tissue inhibitor of metalloproteinase |
| TNF-α | tumor necrosis factor-α |
| UV | ultraviolet |
References
- Diaz-Flores, L., Jr.; Madrid, J.F.; Gutierrez, R.; Varela, H.; Valladares, F.; Alvarez-Arguelles, H.; Diaz-Flores, L. Adult stem and transit-amplifying cell location. Histol. Histopathol. 2006, 21, 995–1027. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xie, T. Stem cell niche: Structure and function. Annu. Rev. Cell Dev. Biol. 2005, 21, 605–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Li, Y.; Fan, H.; Liu, Z.; Pestell, R.G. miRNAs regulate stem cell self-renewal and differentiation. Front. Genet. 2012, 3, 191. [Google Scholar] [CrossRef] [Green Version]
- Youn, S.W.; Kim, D.S.; Cho, H.J.; Jeon, S.E.; Bae, I.H.; Yoon, H.J.; Park, K.C. Cellular senescence induced loss of stem cell proportion in the skin in vitro. J. Dermatol. Sci. 2004, 35, 113–123. [Google Scholar] [CrossRef]
- Watt, F.M.; Driskell, R.R. The therapeutic potential of stem cells. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Watt, F.M.; Fujiwara, H. Cell-extracellular matrix interactions in normal and diseased skin. Cold Spring Harb. Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.R.; Kang, Y.A.; Shin, J.W.; Na, J.I.; Huh, C.H.; Park, K.C. Redox status is critical for stemness in skin equivalents. Oxid. Med. Cell Longev. 2012, 2012, 819623. [Google Scholar] [CrossRef] [Green Version]
- Na, J.I.; Shin, J.W.; Choi, H.R.; Kwon, S.H.; Park, K.C. Resveratrol as a multifunctional topical hypopigmenting agent. Int. J. Mol. Sci. 2019, 20, 956. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.R.; Nam, K.M.; Lee, H.S.; Yang, S.H.; Kim, Y.S.; Lee, J.; Date, A.; Toyama, K.; Park, K.C. Phlorizin, an active ingredient of Eleutherococcus senticosus, increases proliferative potential of keratinocytes with inhibition of MiR135b and increased expression of type IV collagen. Oxid. Med. Cell Longev. 2016, 2016, 3859721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Y.A.; Choi, H.R.; Na, J.I.; Huh, C.H.; Kim, M.J.; Youn, S.W.; Kim, K.H.; Park, K.C. Copper-GHK increases integrin expression and p63 positivity by keratinocytes. Arch. Dermatol. Res. 2009, 301, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.R.; Shin, J.W.; Na, J.I.; Nam, K.M.; Lee, H.S.; Park, K.C. Novel antioxidant tripeptide “ACQ” can prevent UV-induced cell death and preserve the number of epidermal stem cells. Oxid. Med. Cell Longev. 2015, 2015, 359740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.R.; Kang, Y.A.; Na, J.I.; Huh, S.Y.; Huh, C.H.; Kim, K.H.; Park, K.C. Oligosaccharides of hyaluronic acid increased epidermal cell stemness by modulation of integrin expression. J. Cosmet. Dermatol. 2012, 11, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.W.; Choi, H.R.; Nam, K.M.; Lee, H.S.; Kim, S.A.; Joe, H.J.; Kazumi, T.; Park, K.C. The Co-expression pattern of p63 and HDAC1: A potential way to disclose stem cells in interfollicular epidermis. Int. J. Mol. Sci. 2017, 18, 1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegrini, G.; Dellambra, E.; Golisano, O.; Martinelli, E.; Fantozzi, I.; Bondanza, S.; Ponzin, D.; McKeon, F.; De Luca, M. p63 identifies keratinocyte stem cells. Proc. Natl. Acad. Sci. USA 2001, 98, 3156–3161. [Google Scholar] [CrossRef] [Green Version]
- Candi, E.; Amelio, I.; Agostini, M.; Melino, G. MicroRNAs and p63 in epithelial stemness. Cell Death Differ. 2015, 22, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Koster, M.I.; Kim, S.; Roop, D.R. P63 deficiency: A failure of lineage commitment or stem cell maintenance? J. Investig. Dermatol. Symp. Proc. 2005, 10, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Bravo, R.; Frank, R.; Blundell, P.A.; Macdonald-Bravo, H. Cyclin/PCNA is the auxiliary protein of DNA polymerase-delta. Nature 1987, 326, 515–517. [Google Scholar] [CrossRef]
- Breitkreutz, D.; Koxholt, I.; Thiemann, K.; Nischt, R. Skin basement membrane: The foundation of epidermal integrity--BM functions and diverse roles of bridging molecules nidogen and perlecan. Biomed. Res. Int. 2013, 2013, 179784. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.R.; Byun, S.Y.; Kwon, S.H.; Park, K.C. Niche interactions in epidermal stem cells. World J. Stem Cells 2015, 7, 495–501. [Google Scholar] [CrossRef]
- Sonnenberg, A.; Calafat, J.; Janssen, H.; Daams, H.; van der Raaij-Helmer, L.M.; Falcioni, R.; Kennel, S.J.; Aplin, J.D.; Baker, J.; Loizidou, M.; et al. Integrin alpha 6/beta 4 complex is located in hemidesmosomes, suggesting a major role in epidermal cell-basement membrane adhesion. J. Cell Biol. 1991, 113, 907–917. [Google Scholar] [CrossRef]
- Grose, R.; Hutter, C.; Bloch, W.; Thorey, I.; Watt, F.M.; Fassler, R.; Brakebusch, C.; Werner, S. A crucial role of beta 1 integrins for keratinocyte migration in vitro and during cutaneous wound repair. Development 2002, 129, 2303–2315. [Google Scholar] [PubMed]
- Kretzschmar, K.; Watt, F.M. Markers of epidermal stem cell subpopulations in adult mammalian skin. Cold Spring Harb. Perspect. Med. 2014, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.J.; Bae, I.H.; Chung, H.J.; Kim, D.S.; Kwon, S.B.; Cho, Y.J.; Youn, S.W.; Park, K.C. Effects of hair follicle dermal sheath cells in the reconstruction of skin equivalents. J. Dermatol. Sci. 2004, 35, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Upton, Z.; Cuttle, L.; Noble, A.; Kempf, M.; Topping, G.; Malda, J.; Xie, Y.; Mill, J.; Harkin, D.G.; Kravchuk, O.; et al. Vitronectin: Growth factor complexes hold potential as a wound therapy approach. J. Investig. Dermatol. 2008, 128, 1535–1544. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, T.L.; Van Lonkhuyzen, D.R.; Dawson, R.A.; Kimlin, M.G.; Upton, Z. Insulin-like growth factor-I and UVB photoprotection in human keratinocytes. Exp. Dermatol. 2015, 24, 235–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, A.J.; Haase, I.; Watt, F.M. Signaling via beta1 integrins and mitogen-activated protein kinase determines human epidermal stem cell fate in vitro. Proc. Natl. Acad. Sci. USA 1999, 96, 6728–6733. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.R.; Nam, K.M.; Park, S.J.; Kim, D.S.; Huh, C.H.; Park, W.Y.; Park, K.C. Suppression of miR135b increases the proliferative potential of normal human keratinocytes. J. Investig. Dermatol. 2014, 134, 1161–1164. [Google Scholar] [CrossRef] [Green Version]
- Peerani, R.; Rao, B.M.; Bauwens, C.; Yin, T.; Wood, G.A.; Nagy, A.; Kumacheva, E.; Zandstra, P.W. Niche-mediated control of human embryonic stem cell self-renewal and differentiation. EMBO J. 2007, 26, 4744–4755. [Google Scholar] [CrossRef] [Green Version]
- Mohyeldin, A.; Garzon-Muvdi, T.; Quinones-Hinojosa, A. Oxygen in stem cell biology: A critical component of the stem cell niche. Cell Stem Cell 2010, 7, 150–161. [Google Scholar] [CrossRef] [Green Version]
- Pervaiz, S.; Taneja, R.; Ghaffari, S. Oxidative stress regulation of stem and progenitor cells. Antioxid. Redox Sign. 2009, 11, 2777–2789. [Google Scholar] [CrossRef]
- Ji, A.R.; Ku, S.Y.; Cho, M.S.; Kim, Y.Y.; Kim, Y.J.; Oh, S.K.; Kim, S.H.; Moon, S.Y.; Choi, Y.M. Reactive oxygen species enhance differentiation of human embryonic stem cells into mesendodermal lineage. Exp. Mol. Med. 2010, 42, 175–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Park, H.H.; Choi, H.; Kim, Y.S.; Yu, H.J.; Lee, K.Y.; Lee, Y.J.; Kim, S.H.; Koh, S.H. Coenzyme Q10 protects neural stem cells against hypoxia by enhancing survival signals. Brain Res. 2012, 1478, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Carr, W.J.; Oberley-Deegan, R.E.; Zhang, Y.; Oberley, C.C.; Oberley, L.W.; Dunnwald, M. Antioxidant proteins and reactive oxygen species are decreased in a murine epidermal side population with stem cell-like characteristics. Histochem. Cell Biol. 2011, 135, 293–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhie, G.; Shin, M.H.; Seo, J.Y.; Choi, W.W.; Cho, K.H.; Kim, K.H.; Park, K.C.; Eun, H.C.; Chung, J.H. Aging-and photoaging-dependent changes of enzymic and nonenzymic antioxidants in the epidermis and dermis of human skin in vivo. J. Investig. Dermatol. 2001, 117, 1212–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV radiation and the skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [Green Version]
- Schuch, A.P.; Moreno, N.C.; Schuch, N.J.; Menck, C.F.M.; Garcia, C.C.M. Sunlight damage to cellular DNA: Focus on oxidatively generated lesions. Free Radic. Biol. Med. 2017, 107, 110–124. [Google Scholar] [CrossRef]
- Cesarini, J.P.; Michel, L.; Maurette, J.M.; Adhoute, H.; Bejot, M. Immediate effects of UV radiation on the skin: Modification by an antioxidant complex containing carotenoids. Photodermatol. Photoimmunol. Photomed. 2003, 19, 182–189. [Google Scholar] [CrossRef] [Green Version]
- Droge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- Halliday, G.M. Inflammation, gene mutation and photoimmunosuppression in response to UVR-induced oxidative damage contributes to photocarcinogenesis. Mutat. Res. 2005, 571, 107–120. [Google Scholar] [CrossRef]
- Shindo, Y.; Witt, E.; Han, D.; Epstein, W.; Packer, L. Enzymic and non-enzymic antioxidants in epidermis and dermis of human skin. J. Investig. Dermatol. 1994, 102, 122–124. [Google Scholar] [CrossRef] [Green Version]
- Pence, B.C.; Naylor, M.F. Effects of single-dose ultraviolet radiation on skin superoxide dismutase, catalase, and xanthine oxidase in hairless mice. J. Investig. Dermatol. 1990, 95, 213–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shindo, Y.; Witt, E.; Packer, L. Antioxidant defense mechanisms in murine epidermis and dermis and their responses to ultraviolet light. J. Investig. Dermatol. 1993, 100, 260–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sander, C.S.; Chang, H.; Salzmann, S.; Muller, C.S.; Ekanayake-Mudiyanselage, S.; Elsner, P.; Thiele, J.J. Photoaging is associated with protein oxidation in human skin in vivo. J. Investig. Dermatol. 2002, 118, 618–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.S.; Cho, H.J.; Choi, H.R.; Kwon, S.B.; Park, K.C. Isolation of human epidermal stem cells by adherence and the reconstruction of skin equivalents. Cell. Mol. Life Sci. 2004, 61, 2774–2781. [Google Scholar] [CrossRef] [PubMed]
- Padayatty, S.J.; Levine, M. Vitamin C: The known and the unknown and Goldilocks. Oral Dis. 2016, 22, 463–493. [Google Scholar] [CrossRef] [Green Version]
- Peterkofsky, B. Ascorbate requirement for hydroxylation and secretion of procollagen: Relationship to inhibition of collagen synthesis in scurvy. Am. J. Clin. Nutr. 1991, 54, 1135S–1140S. [Google Scholar] [CrossRef]
- Sram, R.J.; Binkova, B.; Rossner, P., Jr. Vitamin C for DNA damage prevention. Mutat. Res. 2012, 733, 39–49. [Google Scholar] [CrossRef]
- Savini, I.; D’Angelo, I.; Ranalli, M.; Melino, G.; Avigliano, L. Ascorbic acid maintenance in HaCaT cells prevents radical formation and apoptosis by UV-B. Free Radic. Biol. Med. 1999, 26, 1172–1180. [Google Scholar] [CrossRef]
- Stewart, M.S.; Cameron, G.S.; Pence, B.C. Antioxidant nutrients protect against UVB-induced oxidative damage to DNA of mouse keratinocytes in culture. J. Investig. Dermatol. 1996, 106, 1086–1089. [Google Scholar] [CrossRef] [Green Version]
- Kawashima, S.; Funakoshi, T.; Sato, Y.; Saito, N.; Ohsawa, H.; Kurita, K.; Nagata, K.; Yoshida, M.; Ishigami, A. Protective effect of pre- and post-vitamin C treatments on UVB-irradiation-induced skin damage. Sci. Rep. 2018, 8, 16199. [Google Scholar] [CrossRef]
- Cimmino, L.; Neel, B.G.; Aifantis, I. Vitamin C in Stem Cell Reprogramming and Cancer. Trends Cell. Biol. 2018, 28, 698–708. [Google Scholar] [CrossRef] [PubMed]
- Gorres, K.L.; Raines, R.T. Prolyl 4-hydroxylase. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 106–124. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Chen, K.; Zeng, X.; Yang, J.; Wu, Y.; Shi, X.; Qin, B.; Zeng, L.; Esteban, M.A.; Pan, G.; et al. The histone demethylases Jhdm1a/1b enhance somatic cell reprogramming in a vitamin-C-dependent manner. Cell Stem Cell 2011, 9, 575–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, C.; Jia, G.; Hou, G.; Dai, Q.; Zhang, W.; Zheng, G.; Jian, X.; Yang, C.G.; Cui, Q.; He, C. Iron-catalysed oxidation intermediates captured in a DNA repair dioxygenase. Nature 2010, 468, 330–333. [Google Scholar] [CrossRef] [PubMed]
- Gerken, T.; Girard, C.A.; Tung, Y.C.; Webby, C.J.; Saudek, V.; Hewitson, K.S.; Yeo, G.S.; McDonough, M.A.; Cunliffe, S.; McNeill, L.A.; et al. The obesity-associated FTO gene encodes a 2-oxoglutarate-dependent nucleic acid demethylase. Science 2007, 318, 1469–1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaschke, K.; Ebata, K.T.; Karimi, M.M.; Zepeda-Martinez, J.A.; Goyal, P.; Mahapatra, S.; Tam, A.; Laird, D.J.; Hirst, M.; Rao, A.; et al. Vitamin C induces Tet-dependent DNA demethylation and a blastocyst-like state in ES cells. Nature 2013, 500, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.; Mao, S.Q.; Zhao, B.; Chong, Z.; Yang, Y.; Zhao, C.; Zhang, D.; Huang, H.; Gao, J.; Li, Z.; et al. Ascorbic acid enhances Tet-mediated 5-methylcytosine oxidation and promotes DNA demethylation in mammals. J. Am. Chem. Soc. 2013, 135, 10396–10403. [Google Scholar] [CrossRef]
- Chung, T.L.; Brena, R.M.; Kolle, G.; Grimmond, S.M.; Berman, B.P.; Laird, P.W.; Pera, M.F.; Wolvetang, E.J. Vitamin C promotes widespread yet specific DNA demethylation of the epigenome in human embryonic stem cells. Stem Cells 2010, 28, 1848–1855. [Google Scholar] [CrossRef]
- Fujisawa, K.; Hara, K.; Takami, T.; Okada, S.; Matsumoto, T.; Yamamoto, N.; Sakaida, I. Evaluation of the effects of ascorbic acid on metabolism of human mesenchymal stem cells. Stem Cell Res. Ther. 2018, 9, 93. [Google Scholar] [CrossRef]
- Minor, E.A.; Court, B.L.; Young, J.I.; Wang, G. Ascorbate induces ten-eleven translocation (Tet) methylcytosine dioxygenase-mediated generation of 5-hydroxymethylcytosine. J. Biol. Chem. 2013, 288, 13669–13674. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Chen, Y.; Bian, C.; Fujiki, R.; Yu, X. TET2 promotes histone O-GlcNAcylation during gene transcription. Nature 2013, 493, 561–564. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lan, J.; Liu, D.; Backman, L.J.; Zhang, W.; Zhou, Q.; Danielson, P. Ascorbic acid promotes the stemness of corneal epithelial stem/progenitor cells and accelerates epithelial wound healing in the cornea. Stem Cells Transl. Med. 2017, 6, 1356–1365. [Google Scholar] [CrossRef] [PubMed]
- Rembe, J.D.; Fromm-Dornieden, C.; Stuermer, E.K. Effects of vitamin B complex and vitamin C on human skin cells: Is the perceived effect measurable? Adv. Skin Wound Care 2018, 31, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Lin, J.; Yin, Y.; Zhao, J.; Sun, X.; Tang, K. Ganodermataceae: Natural products and their related pharmacological functions. Am. J. Chin. Med. 2007, 35, 559–574. [Google Scholar] [CrossRef] [PubMed]
- Li, L.D.; Mao, P.W.; Shao, K.D.; Bai, X.H.; Zhou, X.W. Ganoderma proteins and their potential applications in cosmetics. Appl. Microbiol. Biotechnol. 2019, 103, 9239–9250. [Google Scholar] [CrossRef]
- Smina, T.P.; Mathew, J.; Janardhanan, K.K.; Devasagayam, T.P. Antioxidant activity and toxicity profile of total triterpenes isolated from Ganoderma lucidum (Fr.) P. Karst occurring in South India. Environ. Toxicol. Pharmacol. 2011, 32, 438–446. [Google Scholar] [CrossRef]
- Yin, Z.; Yang, B.; Ren, H. Preventive and therapeutic effect of Ganoderma (Lingzhi) on skin diseases and care. Adv. Exp. Med. Biol. 2019, 1182, 311–321. [Google Scholar] [CrossRef]
- You, Y.H.; Lin, Z.B. Antioxidant effect of Ganoderma polysaccharide peptide. Yao Xue Xue Bao 2003, 38, 85–88. [Google Scholar]
- You, Y.H.; Lin, Z.B. Protective effects of Ganoderma lucidum polysaccharides peptide on injury of macrophages induced by reactive oxygen species. Acta Pharmacol. Sin. 2002, 23, 787–791. [Google Scholar]
- Zhonghui, Z.; Xiaowei, Z.; Fang, F. Ganoderma lucidum polysaccharides supplementation attenuates exercise-induced oxidative stress in skeletal muscle of mice. Saudi J. Biol. Sci. 2014, 21, 119–123. [Google Scholar] [CrossRef] [Green Version]
- Zhong, D.; Wang, H.; Liu, M.; Li, X.; Huang, M.; Zhou, H.; Lin, S.; Lin, Z.; Yang, B. Ganoderma lucidum polysaccharide peptide prevents renal ischemia reperfusion injury via counteracting oxidative stress. Sci. Rep. 2015, 5, 16910. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.Y.; Tang, Y.P.; Xiang, J.; Wua, P.; Jin, H.M.; Wang, Z.; Mori, M.; Cai, D.F. Neuroprotective effects of water-soluble Ganoderma lucidum polysaccharides on cerebral ischemic injury in rats. J. Ethnopharmacol. 2010, 131, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, Q.; Deng, W.; Li, Y.; Xing, G.; Shi, X.; Du, Y. Neuroprotective effect of pretreatment with Ganoderma lucidum in cerebral ischemia/reperfusion injury in rat hippocampus. Neural Regen. Res. 2014, 9, 1446–1452. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Meng, X.; Yin, J.; Sun, J.; Huang, Q.; Yin, Z. Ganoderma lucidum polysaccharide peptide attenuates skin flap ischemia-reperfusion injury in a thioredoxin-dependent manner. Plast Reconstr. Surg. 2018, 142, 23e–33e. [Google Scholar] [CrossRef]
- Zeng, Q.; Zhou, F.; Lei, L.; Chen, J.; Lu, J.; Zhou, J.; Cao, K.; Gao, L.; Xia, F.; Ding, S.; et al. Ganoderma lucidum polysaccharides protect fibroblasts against UVB-induced photoaging. Mol. Med. Rep. 2017, 15, 111–116. [Google Scholar] [CrossRef] [Green Version]
- Chiang, H.M.; Chen, H.C.; Wu, C.S.; Wu, P.Y.; Wen, K.C. Rhodiola plants: Chemistry and biological activity. J. Food Drug Anal. 2015, 23, 359–369. [Google Scholar] [CrossRef] [Green Version]
- Panossian, A.; Wikman, G.; Sarris, J. Rosenroot (Rhodiola rosea): Traditional use, chemical composition, pharmacology and clinical efficacy. Phytomedicine 2010, 17, 481–493. [Google Scholar] [CrossRef]
- Chen, T.S.; Liou, S.Y.; Chang, Y.L. Antioxidant evaluation of three adaptogen extracts. Am. J. Chin. Med. 2008, 36, 1209–1217. [Google Scholar] [CrossRef]
- Gupta, A.; Kumar, R.; Upadhyay, N.K.; Pal, K.; Kumar, R.; Sawhney, R.C. Effects of Rhodiola imbricata on dermal wound healing. Planta Med. 2007, 73, 774–777. [Google Scholar] [CrossRef]
- Calcabrini, C.; De Bellis, R.; Mancini, U.; Cucchiarini, L.; Potenza, L.; De Sanctis, R.; Patrone, V.; Scesa, C.; Dacha, M. Rhodiola rosea ability to enrich cellular antioxidant defences of cultured human keratinocytes. Arch. Dermatol. Res. 2010, 302, 191–200. [Google Scholar] [CrossRef]
- Zhou, Q.; Yin, Z.P.; Ma, L.; Zhao, W.; Hao, H.W.; Li, H.L. Free radical-scavenging activities of oligomeric proanthocyanidin from Rhodiola rosea L. and its antioxidant effects in vivo. Nat. Prod. Res. 2014, 28, 2301–2303. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.T.; Chang, T.C.; Lai, F.Y.; Lin, C.S.; Chao, H.L.; Lee, S.Y. Rhodiola crenulata Attenuates gamma-Ray Induced Cellular Injury via Modulation of Oxidative Stress in Human Skin Cells. Am. J. Chin. Med. 2018, 46, 175–190. [Google Scholar] [CrossRef]
- Rauf, A.; Imran, M.; Suleria, H.A.R.; Ahmad, B.; Peters, D.G.; Mubarak, M.S. A comprehensive review of the health perspectives of resveratrol. Food Funct. 2017, 8, 4284–4305. [Google Scholar] [CrossRef] [PubMed]
- Bastianetto, S.; Dumont, Y.; Duranton, A.; Vercauteren, F.; Breton, L.; Quirion, R. Protective action of resveratrol in human skin: Possible involvement of specific receptor binding sites. PLoS ONE 2010, 5, e12935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boo, Y.C. Human skin lightening efficacy of resveratrol and its analogs: From in vitro studies to cosmetic applications. Antioxidants (Basel) 2019, 8, 332. [Google Scholar] [CrossRef] [Green Version]
- Kwon, S.H.; Choi, H.R.; Kang, Y.A.; Park, K.C. Depigmenting effect of resveratrol is dependent on FOXO3a activation without SIRT1 activation. Int. J. Mol. Sci. 2017, 18, 1213. [Google Scholar] [CrossRef] [Green Version]
- Jagdeo, J.; Adams, L.; Lev-Tov, H.; Sieminska, J.; Michl, J.; Brody, N. Dose-dependent antioxidant function of resveratrol demonstrated via modulation of reactive oxygen species in normal human skin fibroblasts in vitro. J. Drugs Dermatol. 2010, 9, 1523–1526. [Google Scholar]
- Shin, J.W.; Lee, H.S.; Na, J.I.; Huh, C.H.; Park, K.C.; Choi, H.R. Resveratrol inhibits particulate matter-induced inflammatory responses in human keratinocytes. Int. J. Mol. Sci. 2020, 21, 3446. [Google Scholar] [CrossRef]
- Soeur, J.; Eilstein, J.; Lereaux, G.; Jones, C.; Marrot, L. Skin resistance to oxidative stress induced by resveratrol: From Nrf2 activation to GSH biosynthesis. Free Radic. Biol. Med. 2015, 78, 213–223. [Google Scholar] [CrossRef]
- Weller, R. Nitric oxide: A key mediator in cutaneous physiology. Clin. Exp. Dermatol. 2003, 28, 511–514. [Google Scholar] [CrossRef]
- Vitale, N.; Kisslinger, A.; Paladino, S.; Procaccini, C.; Matarese, G.; Pierantoni, G.M.; Mancini, F.P.; Tramontano, D. Resveratrol couples apoptosis with autophagy in UVB-irradiated HaCaT cells. PLoS ONE 2013, 8, e80728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sticozzi, C.; Cervellati, F.; Muresan, X.M.; Cervellati, C.; Valacchi, G. Resveratrol prevents cigarette smoke-induced keratinocytes damage. Food Funct. 2014, 5, 2348–2356. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Lu, S.; Kivlin, R.; Wallin, B.; Card, E.; Bagdasarian, A.; Tamakloe, T.; Chu, W.M.; Guan, K.L.; Wan, Y. AMP-activated protein kinase contributes to UV-and H2O2-induced apoptosis in human skin keratinocytes. J. Biol. Chem. 2010, 285, 14842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ido, Y.; Duranton, A.; Lan, F.; Weikel, K.A.; Breton, L.; Ruderman, N.B. Resveratrol prevents oxidative stress-induced senescence and proliferative dysfunction by activating the AMPK-FOXO3 cascade in cultured primary human keratinocytes. PLoS ONE 2015, 10, e0115341. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Zhao, H.; Huang, B.; Zheng, C.; Peng, W.; Qin, L. Acanthopanax senticosus: Review of botany, chemistry and pharmacology. Pharmazie 2011, 66, 83–97. [Google Scholar]
- Ge, Y.W.; Zhu, S.; Yoshimatsu, K.; Komatsu, K. MS/MS similarity networking accelerated target profiling of triterpene saponins in Eleutherococcus senticosus leaves. Food Chem. 2017, 227, 444–452. [Google Scholar] [CrossRef]
- Olthof, M.R.; Hollman, P.C.; Katan, M.B. Chlorogenic acid and caffeic acid are absorbed in humans. J. Nutr. 2001, 131, 66–71. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Itagaki, S.; Kurokawa, T.; Ogura, J.; Kobayashi, M.; Hirano, T.; Sugawara, M.; Iseki, K. In vitro and in vivo antioxidant properties of chlorogenic acid and caffeic acid. Int. J. Pharm. 2011, 403, 136–138. [Google Scholar] [CrossRef]
- Chen, R.; Liu, Z.; Zhao, J.; Chen, R.; Meng, F.; Zhang, M.; Ge, W. Antioxidant and immunobiological activity of water-soluble polysaccharide fractions purified from Acanthopanax senticosu. Food Chem. 2011, 127, 434–440. [Google Scholar] [CrossRef]
- Wang, X.; Hai, C.X.; Liang, X.; Yu, S.X.; Zhang, W.; Li, Y.L. The protective effects of Acanthopanax senticosus Harms aqueous extracts against oxidative stress: Role of Nrf2 and antioxidant enzymes. J. Ethnopharmacol. 2010, 127, 424–432. [Google Scholar] [CrossRef]
- Baldisserotto, A.; Malisardi, G.; Scalambra, E.; Andreotti, E.; Romagnoli, C.; Vicentini, C.B.; Manfredini, S.; Vertuani, S. Synthesis, antioxidant and antimicrobial activity of a new phloridzin derivative for dermo-cosmetic applications. Molecules 2012, 17, 13275–13289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calliste, C.A.; Le Bail, J.C.; Trouillas, P.; Pouget, C.; Habrioux, G.; Chulia, A.J.; Duroux, J.L. Chalcones: Structural requirements for antioxidant, estrogenic and antiproliferative activities. Anticancer Res. 2001, 21, 3949–3956. [Google Scholar] [PubMed]
- Rezk, B.M.; Haenen, G.R.; van der Vijgh, W.J.; Bast, A. The antioxidant activity of phloretin: The disclosure of a new antioxidant pharmacophore in flavonoids. Biochem. Biophys. Res. Commun. 2002, 295, 9–13. [Google Scholar] [CrossRef]
- Ehrenkranz, J.R.; Lewis, N.G.; Kahn, C.R.; Roth, J. Phlorizin: A review. Diabet. Metab. Res. Rev. 2005, 21, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Ridgway, T.; O’Reilly, J.; West, G.; Tucker, G.; Wiseman, H. Potent antioxidant properties of novel apple-derived flavonoids with commercial potential as food additives. Biochem. Soc. Trans. 1996, 24, 391S. [Google Scholar] [CrossRef] [Green Version]
- Ridgway, T.; O’Reilly, J.; West, G.; Tucker, G.; Wiseman, H. Antioxidant action of novel derivatives of the apple-derived flavonoid phloridzin compared to oestrogen: Relevance to potential cardioprotective action. Biochem. Soc. Trans. 1997, 25, 106S. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Y.; Dang, Y.; Gao, W.; Zhang, Y.; Xu, P.; Gu, J.; Ye, X. P38 and JNK signal pathways are involved in the regulation of phlorizin against UVB-induced skin damage. Exp. Dermatol. 2015, 24, 275–279. [Google Scholar] [CrossRef]
- Wang, Y.F.; Xu, X.; Fan, X.; Zhang, C.; Wei, Q.; Wang, X.; Guo, W.; Xing, W.; Yu, J.; Yan, J.L.; et al. A cell-penetrating peptide suppresses inflammation by inhibiting NF-kappaB signaling. Mol. Ther. 2011, 19, 1849–1857. [Google Scholar] [CrossRef] [Green Version]
- Pickart, L.; Vasquez-Soltero, J.M.; Margolina, A. The human tripeptide GHK-Cu in prevention of oxidative stress and degenerative conditions of aging: Implications for cognitive health. Oxid. Med. Cell Longev. 2012, 2012, 324832. [Google Scholar] [CrossRef] [Green Version]
- Hureau, C.; Eury, H.; Guillot, R.; Bijani, C.; Sayen, S.; Solari, P.L.; Guillon, E.; Faller, P.; Dorlet, P. X-ray and solution structures of Cu (II) GHK and Cu (II) DAHK complexes: Influence on their redox properties. Chemistry 2011, 17, 10151–10160. [Google Scholar] [CrossRef]
- Pickart, L.; Freedman, J.H.; Loker, W.J.; Peisach, J.; Perkins, C.M.; Stenkamp, R.E.; Weinstein, B. Growth-modulating plasma tripeptide may function by facilitating copper uptake into cells. Nature 1980, 288, 715–717. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Huo, Q.; Kele, P.; Andreopoulos, F.M.; Pham, S.M.; Leblanc, R.M. A new fluorescent chemosensor for copper ions based on tripeptide glycyl-histidyl-lysine (GHK). Org. Lett. 2001, 3, 3277–3280. [Google Scholar] [CrossRef] [PubMed]
- Lutsenko, S.; Bhattacharjee, A.; Hubbard, A.L. Copper handling machinery of the brain. Metallomics 2010, 2, 596–608. [Google Scholar] [CrossRef] [PubMed]
- Hawk, S.N.; Lanoue, L.; Keen, C.L.; Kwik-Uribe, C.L.; Rucker, R.B.; Uriu-Adams, J.Y. Copper-deficient rat embryos are characterized by low superoxide dismutase activity and elevated superoxide anions. Biol. Reprod. 2003, 68, 896–903. [Google Scholar] [CrossRef] [Green Version]
- Arul, V.; Gopinath, D.; Gomathi, K.; Jayakumar, R. Biotinylated GHK peptide incorporated collagenous matrix: A novel biomaterial for dermal wound healing in rats. J. Biomed. Mater. Res. B Appl. Biomater. 2005, 73, 383–391. [Google Scholar] [CrossRef]
- Sakuma, S.; Ishimura, M.; Yuba, Y.; Itoh, Y.; Fujimoto, Y. The peptide glycyl-L-histidyl-L-lysine is an endogenous antioxidant in living organisms, possibly by diminishing hydroxyl and peroxyl radicals. Int. J. Physiol. Pathophysiol. Pharmacol. 2018, 10, 132–138. [Google Scholar]
- Miller, D.M.; DeSilva, D.; Pickart, L.; Aust, S.D. Effects of glycyl-histidyl-lysyl chelated Cu (II) on ferritin dependent lipid peroxidation. Adv. Exp. Med. Biol. 1990, 264, 79–84. [Google Scholar] [CrossRef]
- Hong, Y.; Downey, T.; Eu, K.W.; Koh, P.K.; Cheah, P.Y. A ‛metastasis-prone’ signature for early-stage mismatch-repair proficient sporadic colorectal cancer patients and its implications for possible therapeutics. Clin. Exp. Metastasis 2010, 27, 83–90. [Google Scholar] [CrossRef]
- Maquart, F.X.; Pickart, L.; Laurent, M.; Gillery, P.; Monboisse, J.C.; Borel, J.P. Stimulation of collagen synthesis in fibroblast cultures by the tripeptide-copper complex glycyl-L-histidyl-L-lysine-Cu2+. FEBS Lett. 1988, 238, 343–346. [Google Scholar] [CrossRef] [Green Version]
- Maquart, F.X.; Bellon, G.; Chaqour, B.; Wegrowski, J.; Patt, L.M.; Trachy, R.E.; Monboisse, J.C.; Chastang, F.; Birembaut, P.; Gillery, P.; et al. In vivo stimulation of connective tissue accumulation by the tripeptide-copper complex glycyl-L-histidyl-L-lysine-Cu2+ in rat experimental wounds. J. Clin. Investig. 1993, 92, 2368–2376. [Google Scholar] [CrossRef]
- Wegrowski, Y.; Maquart, F.X.; Borel, J.P. Stimulation of sulfated glycosaminoglycan synthesis by the tripeptide-copper complex glycyl-L-histidyl-L-lysine-Cu2+. Life Sci. 1992, 51, 1049–1056. [Google Scholar] [CrossRef]
- Simeon, A.; Wegrowski, Y.; Bontemps, Y.; Maquart, F.X. Expression of glycosaminoglycans and small proteoglycans in wounds: Modulation by the tripeptide-copper complex glycyl-L-histidyl-L-lysine-Cu (2+). J. Investig. Dermatol. 2000, 115, 962–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hocking, A.M.; Shinomura, T.; McQuillan, D.J. Leucine-rich repeat glycoproteins of the extracellular matrix. Matrix Biol. 1998, 17, 1–19. [Google Scholar] [CrossRef]
- Simeon, A.; Monier, F.; Emonard, H.; Gillery, P.; Birembaut, P.; Hornebeck, W.; Maquart, F.X. Expression and activation of matrix metalloproteinases in wounds: Modulation by the tripeptide-copper complex glycyl-L-histidyl-L-lysine-Cu2+. J. Investig. Dermatol. 1999, 112, 957–964. [Google Scholar] [CrossRef] [PubMed]
- Simeon, A.; Emonard, H.; Hornebeck, W.; Maquart, F.X. The tripeptide-copper complex glycyl-L-histidyl-L-lysine-Cu2+ stimulates matrix metalloproteinase-2 expression by fibroblast cultures. Life Sci. 2000, 67, 2257–2265. [Google Scholar] [CrossRef]
- Forman, H.J.; Zhang, H.; Rinna, A. Glutathione: Overview of its protective roles, measurement, and biosynthesis. Mol. Asp. Med. 2009, 30, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Sudha, P.N.; Rose, M.H. Beneficial effects of hyaluronic acid. Adv. Food Nutr. Res. 2014, 72, 137–176. [Google Scholar] [CrossRef]
- Campo, G.M.; Avenoso, A.; Campo, S.; D’Ascola, A.; Ferlazzo, A.M.; Calatroni, A. The antioxidant and antifibrogenic effects of the glycosaminoglycans hyaluronic acid and chondroitin-4-sulphate in a subchronic rat model of carbon tetrachloride-induced liver fibrogenesis. Chem. Biol. Interact. 2004, 148, 125–138. [Google Scholar] [CrossRef]
- Halicka, H.D.; Mitlitski, V.; Heeter, J.; Balazs, E.A.; Darzynkiewicz, Z. Attenuation of the oxidative burst-induced DNA damage in human leukocytes by hyaluronan. Int. J. Mol. Med. 2009, 23, 695–699. [Google Scholar] [CrossRef] [Green Version]
- Litwiniuk, M.; Krejner, A.; Speyrer, M.S.; Gauto, A.R.; Grzela, T. Hyaluronic acid in inflammation and tissue regeneration. Wounds 2016, 28, 78–88. [Google Scholar]
- Ye, J.; Zhang, H.; Wu, H.; Wang, C.; Shi, X.; Xie, J.; He, J.; Yang, J. Cytoprotective effect of hyaluronic acid and hydroxypropyl methylcellulose against DNA damage induced by thimerosal in Chang conjunctival cells. Graefes Arch. Clin. Exp. Ophthalmol. 2012, 250, 1459–1466. [Google Scholar] [CrossRef] [PubMed]
- Foschi, D.; Castoldi, L.; Radaelli, E.; Abelli, P.; Calderini, G.; Rastrelli, A.; Mariscotti, C.; Marazzi, M.; Trabucchi, E. Hyaluronic acid prevents oxygen free-radical damage to granulation tissue: A study in rats. Int. J. Tissue React. 1990, 12, 333–339. [Google Scholar] [PubMed]
- Trabucchi, E.; Pallotta, S.; Morini, M.; Corsi, F.; Franceschini, R.; Casiraghi, A.; Pravettoni, A.; Foschi, D.; Minghetti, P. Low molecular weight hyaluronic acid prevents oxygen free radical damage to granulation tissue during wound healing. Int. J. Tissue React. 2002, 24, 65–71. [Google Scholar] [PubMed]
- Kim, J.K.; Srinivasan, P.; Kim, J.H.; Choi, J.I.; Park, H.J.; Byun, M.W.; Lee, J.W. Structural and antioxidant properties of gamma irradiated hyaluronic acid. Food Chem. 2008, 109, 763–770. [Google Scholar] [CrossRef]
- Ke, C.; Sun, L.; Qiao, D.; Wang, D.; Zeng, X. Antioxidant acitivity of low molecular weight hyaluronic acid. Food Chem. Toxicol. 2011, 49, 2670–2675. [Google Scholar] [CrossRef]
- Kanitakis, J.; Ramirez-Bosca, A.; Reano, A.; Viac, J.; Roche, P.; Thivolet, J. Filaggrin expression in normal and pathological skin. A marker of keratinocyte differentiation. Virchows Arch. A Pathol. Anat. Histopathol. 1988, 412, 375–382. [Google Scholar] [CrossRef]
- Godic, A.; Poljsak, B.; Adamic, M.; Dahmane, R. The role of antioxidants in skin cancer prevention and treatment. Oxid. Med. Cell Longev. 2014, 2014, 860479. [Google Scholar] [CrossRef]
- Piskounova, E.; Agathocleous, M.; Murphy, M.M.; Hu, Z.; Huddlestun, S.E.; Zhao, Z.; Leitch, A.M.; Johnson, T.M.; DeBerardinis, R.J.; Morrison, S.J. Oxidative stress inhibits distant metastasis by human melanoma cells. Nature 2015, 527, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Maione-Silva, L.; de Castro, E.G.; Nascimento, T.L.; Cintra, E.R.; Moreira, L.C.; Cintra, B.A.S.; Valadares, M.C.; Lima, E.M. Ascorbic acid encapsulated into negatively charged liposomes exhibits increased skin permeation, retention and enhances collagen synthesis by fibroblasts. Sci. Rep. 2019, 9, 522. [Google Scholar] [CrossRef] [Green Version]
- Kumano, Y.; Sakamoto, T.; Egawa, M.; Tanaka, M.; Yamamoto, I. Enhancing effect of 2-O-alpha-D-glucopyranosyl-L-ascorbic acid, a stable ascorbic acid derivative, on collagen synthesis. Biol. Pharm. Bull. 1998, 21, 662–666. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, M.; Arai, N.; Kohno, K.; Ushio, S.; Fukuda, S. Anti-oxidative and anti-aging activities of 2-O-alpha-glucopyranosyl-L-ascorbic acid on human dermal fibroblasts. Eur. J. Pharmacol. 2012, 674, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.I.; Park, J.I.; Kim, H.J.; Kim, D.W.; Kim, S.S. A novel L-ascorbic acid and peptide conjugate with increased stability and collagen biosynthesis. BMB Rep. 2009, 42, 743–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.M.; An, H.S.; Bae, J.S.; Kim, J.Y.; Choi, C.H.; Kim, J.Y.; Lim, J.H.; Choi, J.H.; Song, H.; Moon, S.H.; et al. Effects of palmitoyl-KVK-L-ascorbic acid on skin wrinkles and pigmentation. Arch. Dermatol. Res. 2017, 309, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Jo, D.J.; Seok, J.K.; Kim, S.Y.; Park, W.; Baek, J.H.; Kim, Y.M.; Boo, Y.C. Human skin-depigmenting effects of resveratryl triglycolate, a hybrid compound of resveratrol and glycolic acid. Int. J. Cosmet. Sci. 2018. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Park, J.H.; Suh, H.J.; Lee, I.C.; Koh, J.; Boo, Y.C. Effects of resveratrol, oxyresveratrol, and their acetylated derivatives on cellular melanogenesis. Arch. Dermatol. Res. 2014, 306, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.H.; Seok, J.K.; An, S.M.; Baek, J.H.; Koh, J.S.; Boo, Y.C. A study of the human skin-whitening effects of resveratryl triacetate. Arch. Dermatol. Res. 2015, 307, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Chae, J.B.; Yang, S.H.; Byun, S.Y.; Choi, H.R.; Shin, J.W.; Park, K.C. The effects of hydroporation on melasma with anti-aging cocktail. J. Cosmet. Dermatol. 2017, 16, e15–e20. [Google Scholar] [CrossRef]
- Byun, S.Y.; Chae, J.B.; Na, J.I.; Park, K.C. Significant improvement in crow’s feet after treatment with Jet-M and a mixed solution of copper-GHK, oligo-hyaluronic acid, rhodiolar extract, tranexamic acid, and beta-glucan (GHR formulation). J. Cosmet. Laser Ther. 2016, 18, 293–295. [Google Scholar] [CrossRef]
- Shin, J.W.; Choi, H.R.; Yang, S.H.; Choi, J.Y.; Na, J.I.; Huh, C.H.; Park, K.C. The increase of interfollicular epidermal stem cells and regulation of aryl hydrocarbon receptor and its repressors in the skin through hydroporation with anti-aging cocktail. J. Cosmet. Dermatol. 2019, 18, 1133–1139. [Google Scholar] [CrossRef]
- Lorencini, M.; Brohem, C.A.; Dieamant, G.C.; Zanchin, N.I.; Maibach, H.I. Active ingredients against human epidermal aging. Ageing Res. Rev. 2014, 15, 100–115. [Google Scholar] [CrossRef]
Figure 1.
Immunihistochemical staining for integrins, p63, and PCNA. The skin equivalents were constructed and incubated in the presence of vitamin C, G. lucidum, or R. sachalinensis at 1 or 10 μg/mL. Sections of skin equivalents were stained for integrin α6, integrin β1, p63, and PCNA. Staining intensities of both integrins, p63, and PCNA were significantly increased by addition of vitamin C or plant extracts (magnification x200). (* p < 0.05) (modified from the work by Choi et al. [7]).
Figure 1.
Immunihistochemical staining for integrins, p63, and PCNA. The skin equivalents were constructed and incubated in the presence of vitamin C, G. lucidum, or R. sachalinensis at 1 or 10 μg/mL. Sections of skin equivalents were stained for integrin α6, integrin β1, p63, and PCNA. Staining intensities of both integrins, p63, and PCNA were significantly increased by addition of vitamin C or plant extracts (magnification x200). (* p < 0.05) (modified from the work by Choi et al. [7]).

Figure 2.
Histological and immunohistochemical findings for phlorizin-treated skin equivalents. Skin equivalents were constructed and incubated in the presence of phlorizin (0, 50, 200 μM or 1 mM). Sections of skin equivalents were stained with hematoxylin and eosin (H&E), and analyzed by immunohistochemical staining for integrin α6, integrin β1, type IV collagen, p63, and PCNA (magnification ×200). (modified from the work by Choi et al. [9]).
Figure 2.
Histological and immunohistochemical findings for phlorizin-treated skin equivalents. Skin equivalents were constructed and incubated in the presence of phlorizin (0, 50, 200 μM or 1 mM). Sections of skin equivalents were stained with hematoxylin and eosin (H&E), and analyzed by immunohistochemical staining for integrin α6, integrin β1, type IV collagen, p63, and PCNA (magnification ×200). (modified from the work by Choi et al. [9]).

Figure 3.
Immunohistochemical staining for integrin α6 and p63 in alanine-cysteine-glutamine (ACQ)-treated skin equivalents. The expression of integrin α6 and the number of p63-positive cells were increased significantly in skin equivalents incubated in the presence of ACQ (magnification ×200). (in the courtesy of Choi et al. [11]).
Figure 3.
Immunohistochemical staining for integrin α6 and p63 in alanine-cysteine-glutamine (ACQ)-treated skin equivalents. The expression of integrin α6 and the number of p63-positive cells were increased significantly in skin equivalents incubated in the presence of ACQ (magnification ×200). (in the courtesy of Choi et al. [11]).

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kwon, S.-H.; Park, K.-C. Antioxidants as an Epidermal Stem Cell Activator. Antioxidants 2020, 9, 958. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9100958
AMA Style
Kwon S-H, Park K-C. Antioxidants as an Epidermal Stem Cell Activator. Antioxidants. 2020; 9(10):958. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9100958
Chicago/Turabian StyleKwon, Soon-Hyo, and Kyoung-Chan Park. 2020. "Antioxidants as an Epidermal Stem Cell Activator" Antioxidants 9, no. 10: 958. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9100958
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.