Rhus coriaria L. Fruit Extract Prevents UV-A-Induced Genotoxicity and Oxidative Injury in Human Microvascular Endothelial Cells

 , , , and
, , , and 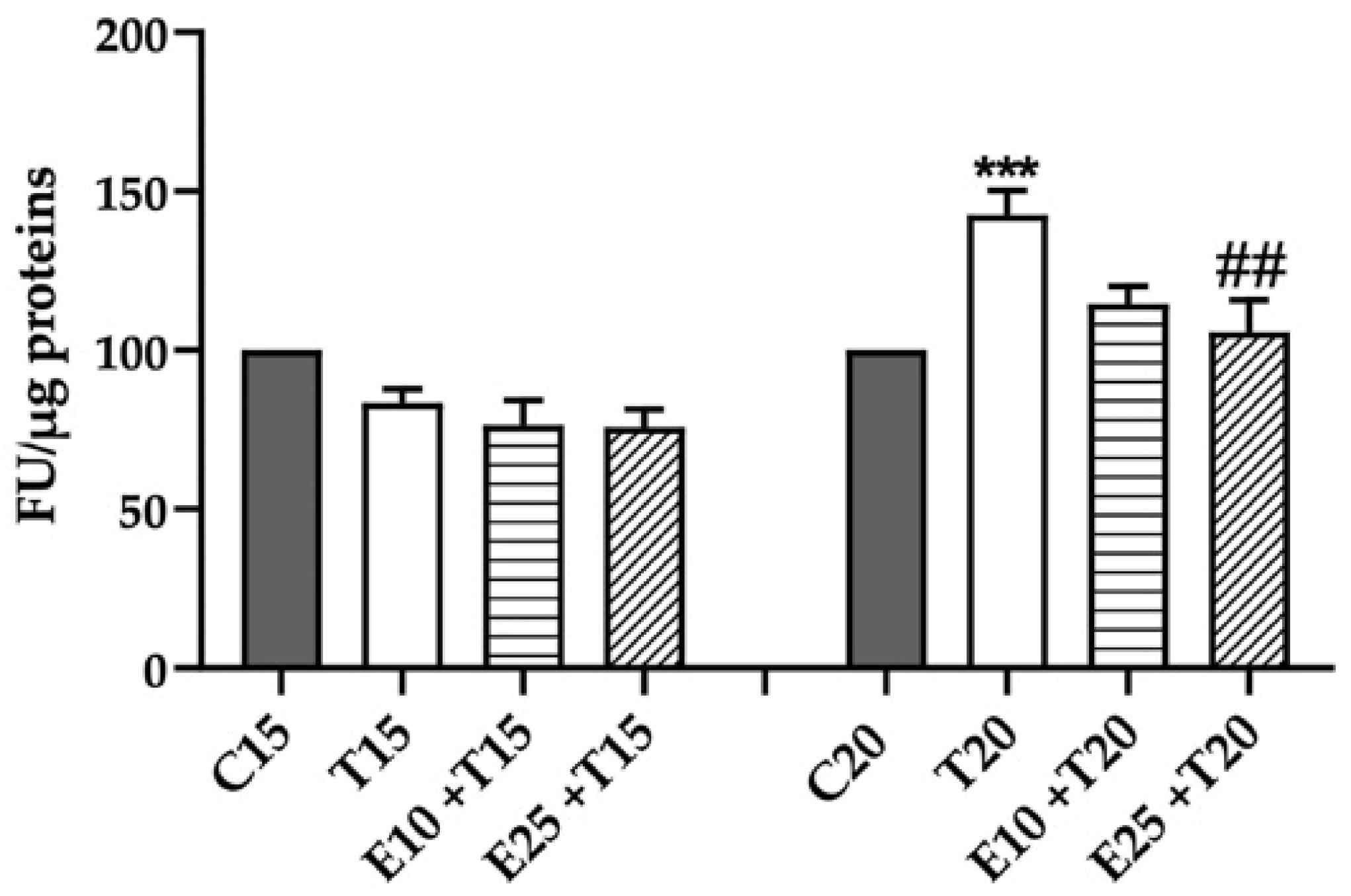{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rhus coriaria L. Extract Preparation and Characterization
2.2. Cell Culture
2.3. Rhus coriaria L. Extract Treatment
2.4. UV-A Radiation Treatment
2.5. ROS Quantification and Lowry Protein Assay
2.6. Quantification of the Total Antioxidant Activity
2.7. Alkaline Comet Assay
2.8. Modified Comet Assay
2.9. Quantification of γ-H2AX and Micronuclei Percentage
2.10. Evaluation of Apoptosis
2.11. Cell Cycle Analysis
2.12. Statistical Analysis
3. Results
3.1. Rhus coriaria L. Extract Decreases UV-A-induced ROS Production
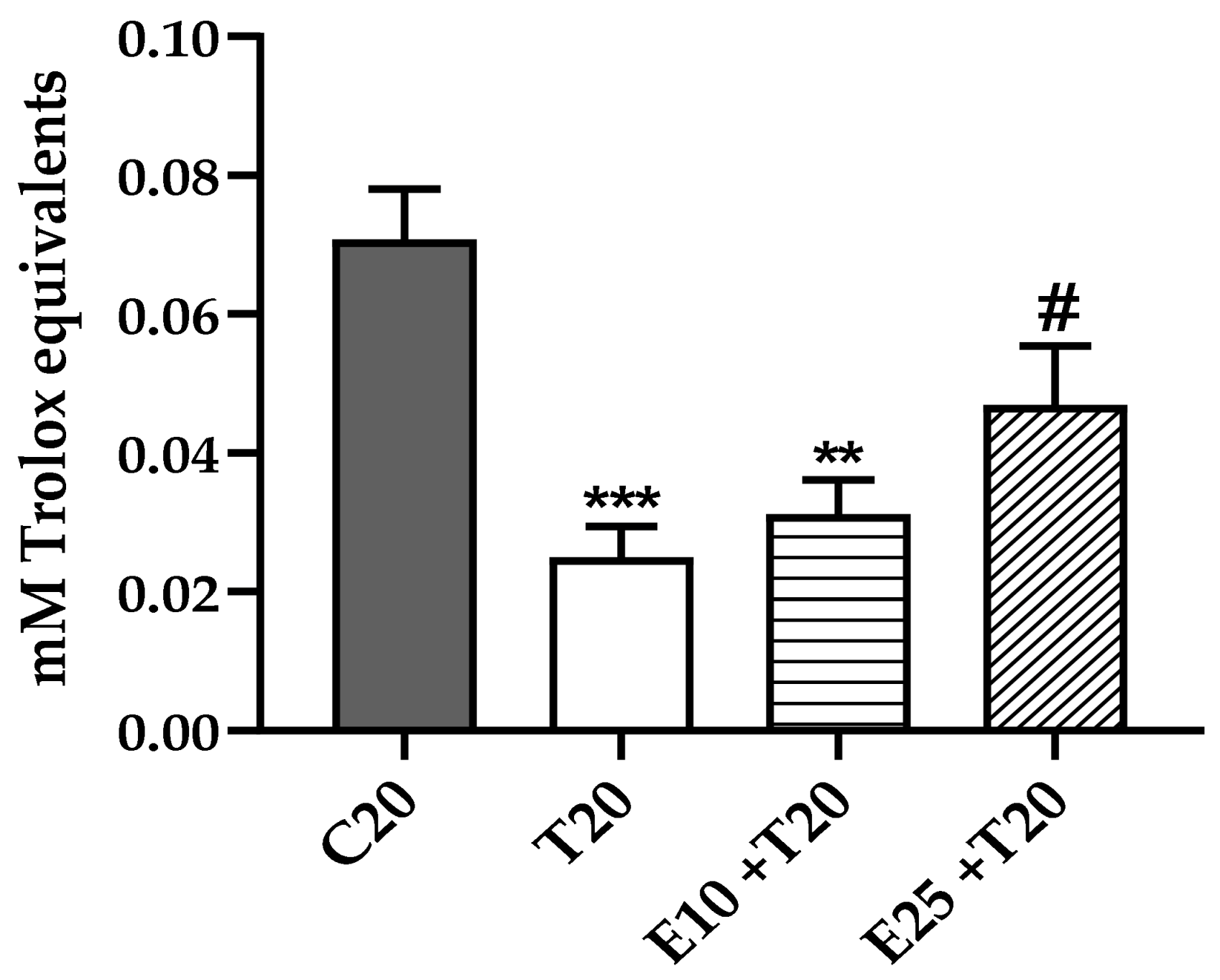
3.2. Total Antioxidant Activity Reduction after UV-A Exposure in the presence of mERC
3.3. Genoprotective Action of mERC Against UV-A Damage
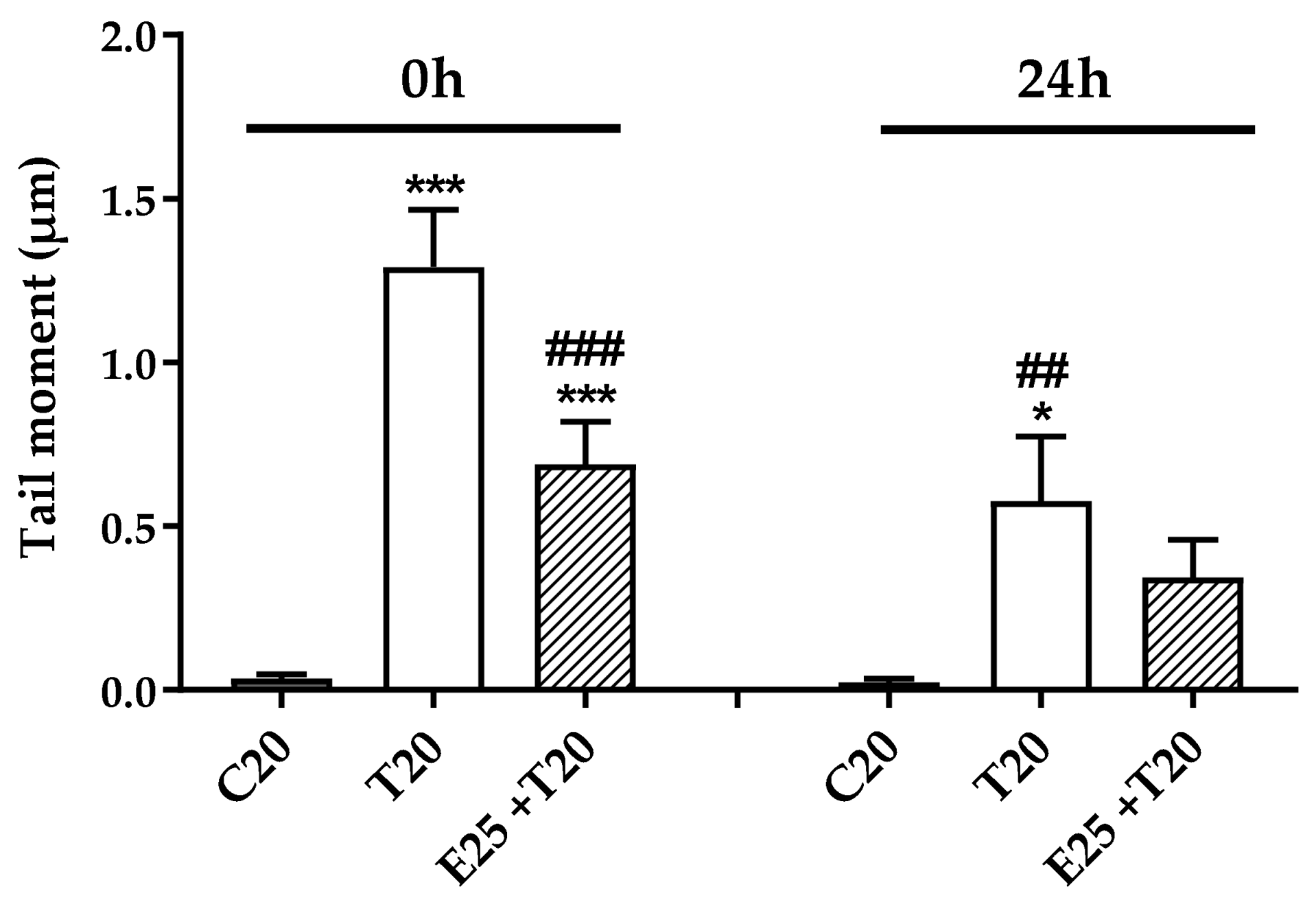
3.3.1. Evaluation of DNA Damage Through Alkaline Comet Assay
3.3.2. Time Course of Genotoxic Damage
3.3.3. DSB Quantification through γ-H2AX Detection
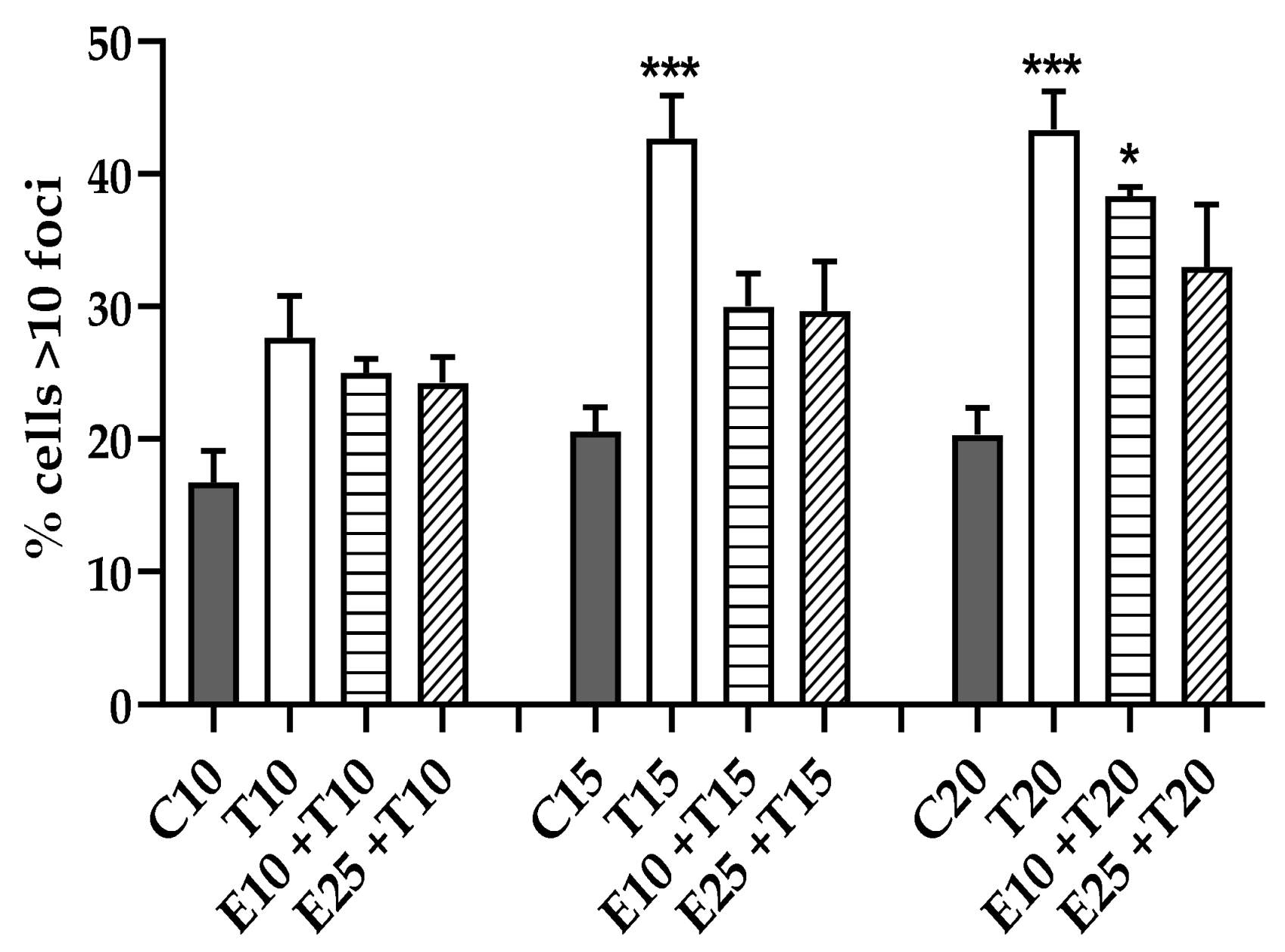
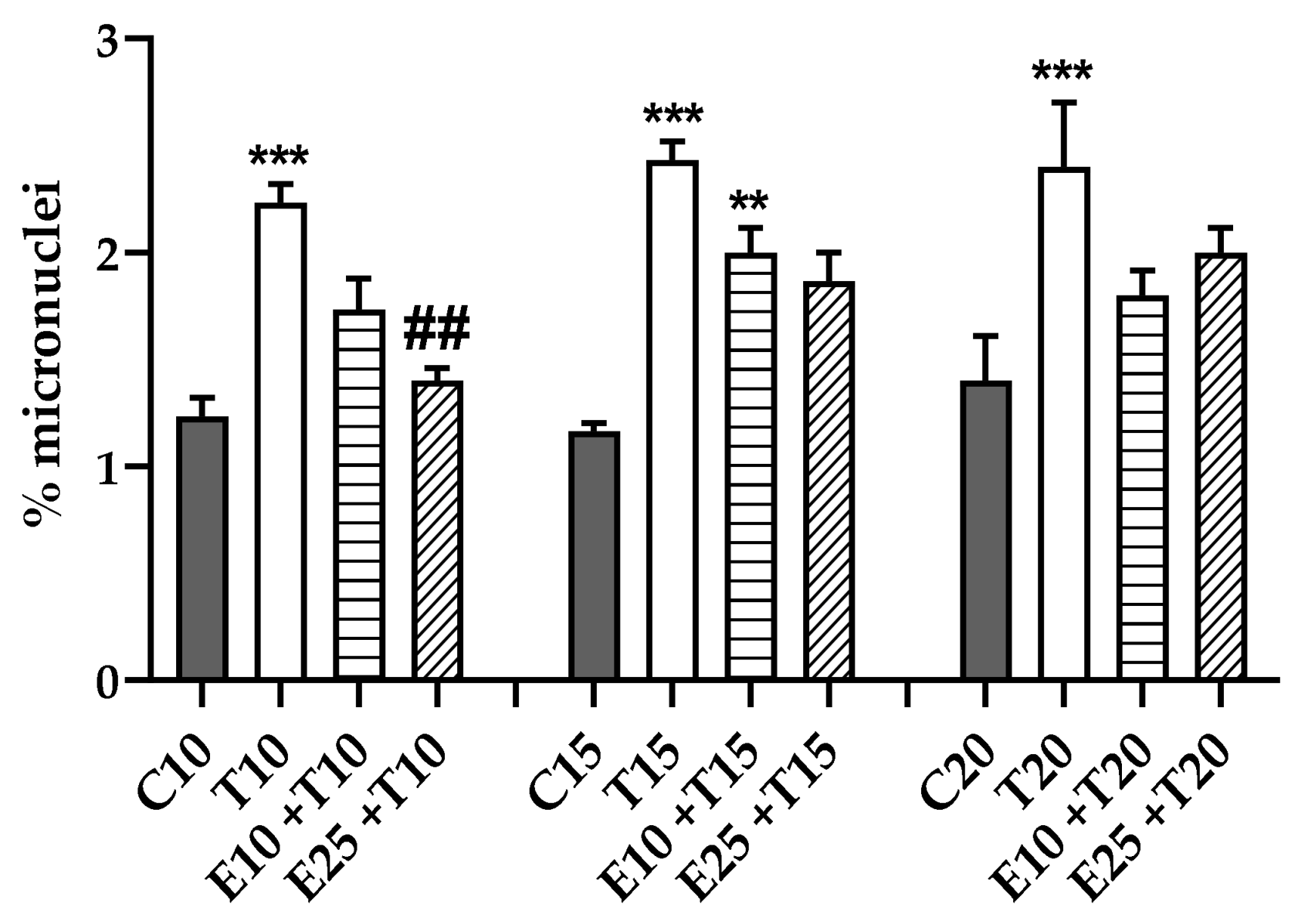
3.3.4. Chromosomal Mis-segregation as Micronuclei Formation
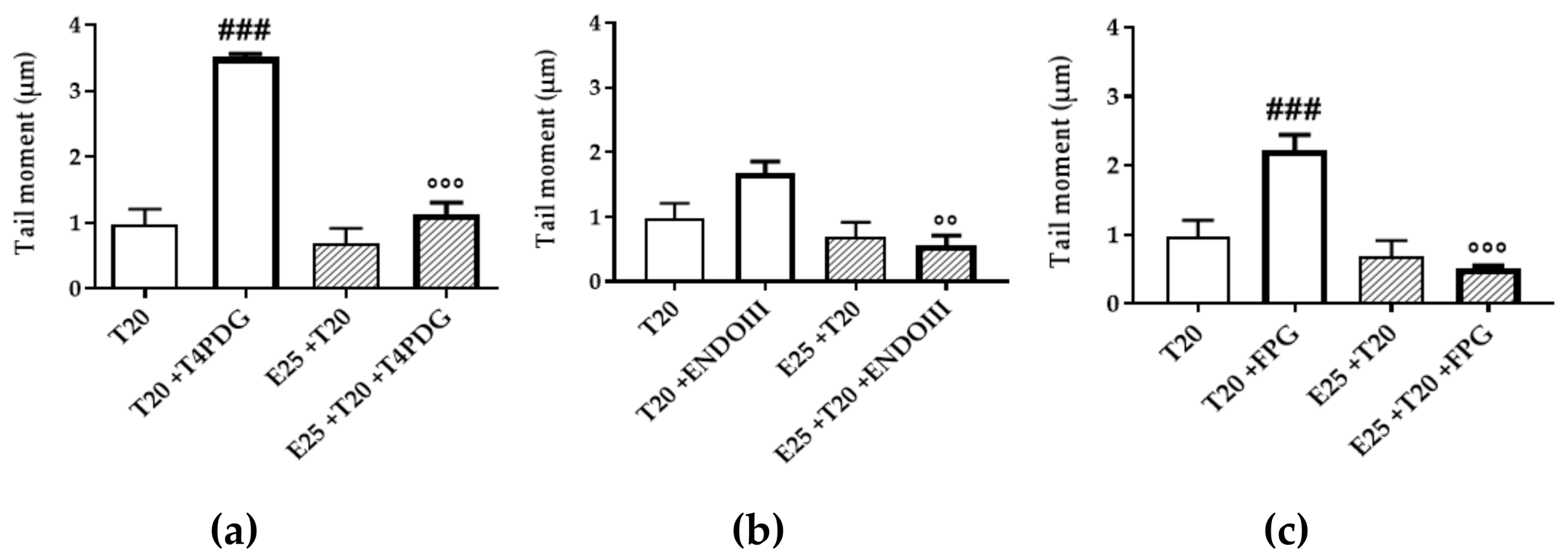
3.4. UV-A Damage Characterization
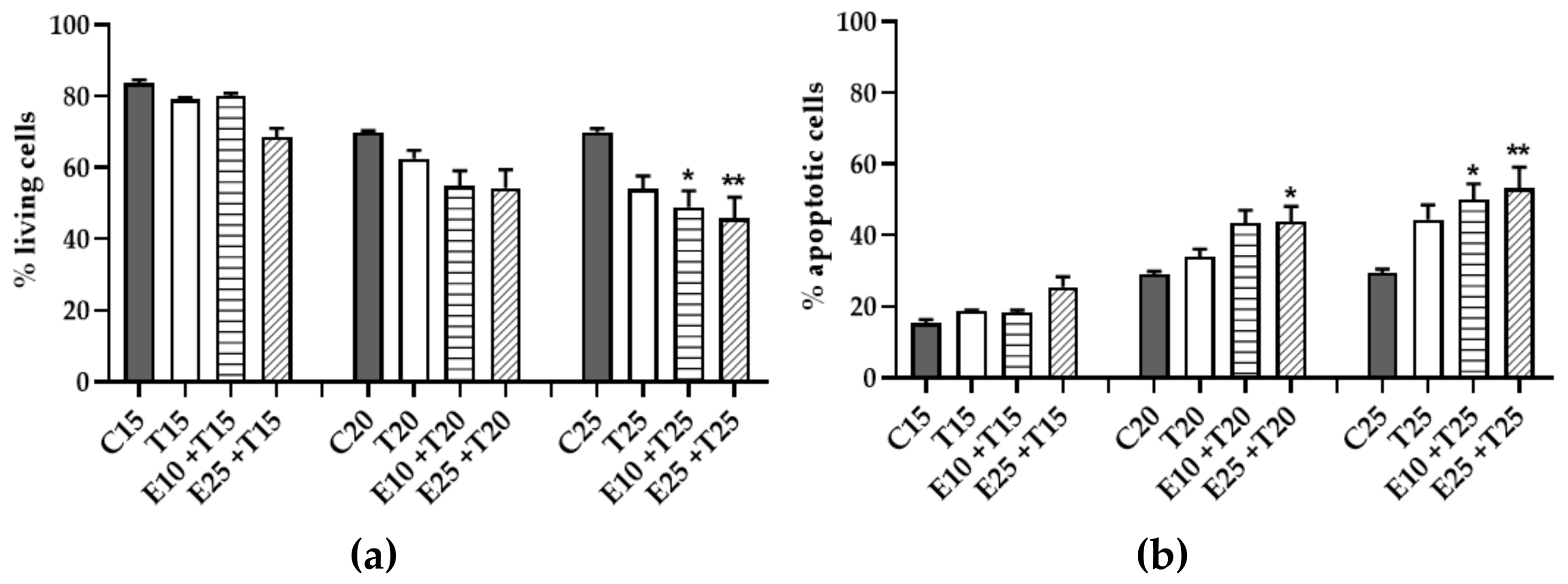
3.5. Assessment of Cytotoxicity
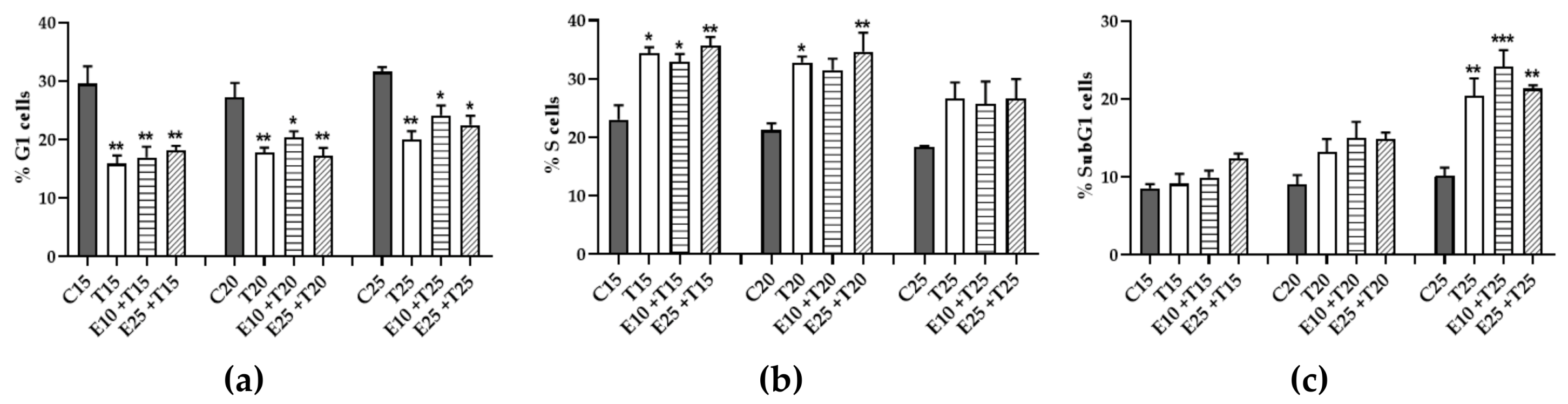
3.6. Role of mERC Pre-treatment and UV-A Exposure on Cell Cycle
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tapiero, H.; Tew, K.D.; Nguyen Ba, G.; Mathé, G. Polyphenols: Do they play a role in the prevention of human pathologies? Biomed. Pharmacother. 2002, 56, 200–207. [Google Scholar] [CrossRef]
- Asgarpanah, J.; Saati, S. An overview of phytochemical and pharmacological properties of Rhus coriaria L. Res. J. Pharmacogn. 2014, 1, 47–54. [Google Scholar]
- Behnammanesh, G.; Khalilpour, S.; Majid, A.S.A.; Majid, A.M.S.A. Pharmacological actions and potential neuroprotective effects of Rhus coriaria L. In addition, Echium amoenum L.: A brief review. WebmedCentral Pharmacol. 2015, 6, WMC005008. [Google Scholar]
- Abu-reidah, I.M.; Jamous, R.M.; Ali-shtayeh, M.S. Phytochemistry, Pharmacological Properties and Industrial Applications of Rhus coriaria L.(Sumac). Jordan J. Biol. Sci. 2014, 7, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Kossah, R.; Nsabimana, C.; Zhang, H.; Chen, W. Optimization of extraction of polyphenols from syrian sumac (Rhus Coriaria L.) and chinese sumac (Rhus Typhina L.) fruit. Res. J. Phytochem. 2010, 4, 146–153. [Google Scholar] [CrossRef] [Green Version]
- Khalilpour, S.; Sangiovanni, E.; Piazza, S.; Fumagalli, M.; Beretta, G.; Dell’Agli, M. In vitro evidence of the traditional use of Rhus coriaria L. fruit against skin inflammatory conditions. J. Ethnopharmacol. 2019, 238, 111829. [Google Scholar] [CrossRef]
- Shabbir, A. Rhus coriaria Linn, a plant of medicinal, nutritional and industrial importance: A review. J. Anim. Plant Sci. 2012, 22, 505–512. [Google Scholar]
- Doğan, A.; Çelik, İ. Healing effects of sumac (Rhus coriaria) in streptozotocin-induced diabetic rats. Pharm. Biol. 2016, 54, 2092–2102. [Google Scholar] [CrossRef] [Green Version]
- Shafiei, M.; Nobakht, M.; Moazzam, A.A. Lipid-lowering effect of Rhus coriaria L. (sumac) fruit extract in hypercholesterolemic rats. Pharmazie 2011, 66, 988–992. [Google Scholar]
- Asgary, S.; Salehizadeh, L.; Keshvari, M.; Taheri, M.; Spence, N.D.; Farvid, M.S.; Rafieian-Kopaei, M.; Sarrafzadegan, N. Potential cardioprotective effects of sumac capsule in patients with hyperlipidemia: A triple-blind randomized, placebo-controlled crossover trial. J. Am. Coll. Nutr. 2018, 37, 286–292. [Google Scholar] [CrossRef]
- Sakhr, K.; El Khatib, S. Physiochemical properties and medicinal, nutritional and industrial applications of Lebanese Sumac (Syrian Sumac-Rhus coriaria): A review. Heliyon 2020, 6, e03207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isik, S.; Tayman, C.; Cakir, U.; Koyuncu, I.; Taskin Turkmenoglu, T.; Cakir, E. Sumac (Rhus coriaria) for the prevention and treatment of necrotizing enterocolitis. J. Food Biochem. 2019, 43, e13068. [Google Scholar] [CrossRef] [PubMed]
- Khalilpour, S.; Behnammanesh, G.; Suede, F.; Ezzat, M.; Muniandy, J.; Tabana, Y.; Ahamed, M.; Tamayol, A.; Majid, A.; Sangiovanni, E.; et al. Neuroprotective and anti-inflammatory effects of rhus coriaria extract in a mouse model of ischemic optic neuropathy. Biomedicines 2018, 6, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalilpour, S.; Latifi, S.; Behnammanesh, G.; Majid, A.M.S.A.; Majid, A.S.A.; Tamayol, A. Ischemic optic neuropathy as a model of neurodegenerative disorder: A review of pathogenic mechanism of axonal degeneration and the role of neuroprotection. J. Neurol. Sci. 2017, 375, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Khalilpour, S.; Behnammanesh, G.; Abdul Majid, A.M.S.; Tamayol, A.; Abdul Majid, A.S. Assessment of neuroprotective properties of Rhus coriaria L. ethanol extract in an in vitro model of retinal degeneration. J. Herb. Med. 2017, 10, 45–52. [Google Scholar] [CrossRef]
- Chakraborty, A.; Ferk, F.; Simić, T.; Brantner, A.; Dušinská, M.; Kundi, M.; Hoelzl, C.; Nersesyan, A.; Knasmüller, S. DNA-protective effects of sumach (Rhus coriaria L.), a common spice: Results of human and animal studies. Mutat. Res.-Fundam. Mol. Mech. Mutagen. 2009, 661, 10–17. [Google Scholar] [CrossRef]
- Heinrich, U.; Moore, C.E.; De Spirt, S.; Tronnier, H.; Stahl, W. Green tea polyphenols provide photoprotection, increase microcirculation, and modulate skin properties of women. J. Nutr. 2011, 141, 1202–1208. [Google Scholar] [CrossRef] [Green Version]
- Nichols, J.A.; Katiyar, S.K. Skin photoprotection by natural polyphenols: Anti-inflammatory, antioxidant and DNA repair mechanisms. Arch. Dermatol. Res. 2010, 302, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Afaq, F.; K Katiyar, S. Polyphenols: Skin photoprotection and inhibition of photocarcinogenesis. Mini-Rev. Med. Chem. 2012, 11, 1200–1215. [Google Scholar]
- Dunaway, S.; Odin, R.; Zhou, L.; Ji, L.; Zhang, Y.; Kadekaro, A.L. Natural antioxidants: Multiple mechanisms to protect skin from solar radiation. Front. Pharmacol. 2018, 9, 392. [Google Scholar] [CrossRef] [Green Version]
- Battie, C.; Jitsukawa, S.; Bernerd, F.; Del Bino, S.; Marionnet, C.; Verschoore, M. New insights in photoaging, UVA induced damage and skin types. Exp. Dermatol. 2014, 23, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Metral, E.; Rachidi, W.; Damour, O.; Demarne, F.; Bechetoille, N. Long-term genoprotection effect of sechium edule fruit extract against UVA irradiation in keratinocytes. Photochem. Photobiol. 2018, 94, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.; Osterwalder, U.; Wang, S.Q.; Burnett, M.; Lim, H.W. Photoprotection: Part II. Sunscreen: Development, efficacy, and controversies. J. Am. Acad. Dermatol. 2013, 69, 867-e1. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, T.; Sasaki, S.; Manabe, Y.; Hirata, T.; Sugawara, T. Preventive effect of dietary astaxanthin on UVA-induced skin photoaging in hairless mice. PLoS ONE 2017, 12, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Cauchard, J.H.; Robinet, A.; Poitevin, S.; Bobichon, H.; Maziere, J.C.; Bellon, G.; Hornebeck, W. UVA-mediated down-regulation of MMP-2 and MT1-MMP coincides with impaired angiogenic phenotype of human dermal endothelial cells. Biochem. Biophys. Res. Commun. 2006, 345, 681–687. [Google Scholar] [CrossRef]
- Seite, S.; Zucchi, H.; Septier, D.; Igondjo-Tchen, S.; Senni, K.; Godeau, G. Elastin changes during chronological and photo-ageing: The important role of lysozyme. J. Eur. Acad. Dermatol. Venereol. 2006, 30, 980–987. [Google Scholar] [CrossRef]
- Hern, S.; Mortimer, P.S. Visualization of dermal blood vessels-capillaroscopy. Clin. Exp. Dermatol. 1999, 24, 473–478. [Google Scholar] [CrossRef]
- Zheng, P.; Kligman, L.H. UVA-induced ultrastructural changes in hairless mouse skin: A comparison to UVB-induced damage. J. Investig. Dermatol. 1993, 100, 194–199. [Google Scholar] [CrossRef] [Green Version]
- D’Alessandro, S.; Magnavacca, A.; Perego, F.; Fumagalli, M.; Sangiovanni, E.; Prato, M.; Dell’Agli, M.; Basilico, N. Effect of hypoxia on gene expression in cell populations involved in wound healing. BioMed Res. Int. 2019, 2019, 2626374. [Google Scholar] [CrossRef] [Green Version]
- Heckmann, M.; Eberlein-Konig, B.; Wollenberg, A.; Przbylla, B.; Plewig, G. Ultraviolet-A radiation induces adhesion molecule expression on human dermal microvascular endothelial cells. Br. J. Dermatol. 1994, 131, 311–318. [Google Scholar] [CrossRef]
- Meroni, E.; Criscuoli, F.; Casiraghi, M.C.; Erba, D.; Papini, N.; Massaccesi, L.; Basilico, N. Metabolic responses in endothelial cells following exposure to ketone bodies. Nutrients 2018, 10, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yohn, J.J.; Lyons, M.B.; Norris, D.A. Cultured human melanocytes from black and white donors have different sunlight and ultraviolet a radiation sensitivities. J. Investig. Dermatol. 1992, 99, 454–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowry, O.; Rosebrough, N.; Farr, L.; Randall, R. Lowry protein assay. J. Biol. Chem. 1951, 193, 365–375. [Google Scholar]
- Muthusamy, G.; Balupillai, A.; Govindasamy, K.; Ramasamy, K.; Ponniresan, V.; Malla, I.; Nagarajan, R. Modified comet assays for the detection of cyclobutane pyrimidine dimers and oxidative base damages. J. Radiat. Cancer Res. 2017, 8, 82–86. [Google Scholar]
- Fenech, M. The in vitro micronucleus technique. Mutat. Res.-Fundam. Mol. Mech. Mutagen. 2000, 455, 81–95. [Google Scholar] [CrossRef]
- Schuch, A.P.; Moreno, N.C.; Schuch, N.J.; Menck, C.F.M.; Garcia, C.C.M. Sunlight damage to cellular DNA: Focus on oxidatively generated lesions. Free Radic. Biol. Med. 2017, 107, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Harrison, D.G. Endothelial dysfunction in cardiovascular diseases: The role of oxidant stress. Circ. Res. 2000, 87, 840–844. [Google Scholar] [CrossRef] [Green Version]
- Truong, V.L.; Jun, M.; Jeong, W.S. Role of resveratrol in regulation of cellular defense systems against oxidative stress. BioFactors 2018, 44, 36–49. [Google Scholar] [CrossRef]
- Kosar, M.; Bozan, B.; Temelli, F.; Baser, K.H.C. Antioxidant activity and phenolic composition of sumac (Rhus coriaria L.) extracts. Food Chem. 2007, 103, 952–959. [Google Scholar] [CrossRef]
- Pourahmad, J.; Eskandari, M.R.; Shakibaei, R.; Kamalinejad, M. A search for hepatoprotective activity of aqueous extract of Rhus coriaria L. against oxidative stress cytotoxicity. Food Chem. Toxicol. 2010, 48, 854–858. [Google Scholar] [CrossRef]
- Segerbäck, D.; Strozyk, M.; Snellman, E.; Hemminki, K. Repair of UV dimers in skin DNA of patients with basal cell carcinoma. Cancer Epidemiol. Biomark. Prev. 2008, 17, 2388–2392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shokrollahi Barough, M.; Hasanzadeh, H.; Barati, M.; Pak, F.; Kokhaei, P.; Rezaei-Tavirani, M. Apoptosis/necrosis induction by ultraviolet, in ER positive and ER negative breast cancer cell lines. Int. J. Cancer Manag. 2015, 8, 41–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wischermann, K.; Popp, S.; Moshir, S.; Scharfetter-Kochanek, K.; Wlaschek, M.; De Gruijl, F.; Hartschuh, W.; Greinert, R.; Volkmer, B.; Faust, A.; et al. UVA radiation causes DNA strand breaks, chromosomal aberrations and tumorigenic transformation in HaCaT skin keratinocytes. Oncogene 2008, 27, 4269–4280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luzhna, L.; Kathiria, P.; Kovalchuk, O. Micronuclei in genotoxicity assessment: From genetics to epigenetics and beyond. Front. Genet. 2013, 4, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadet, J.; Douki, T. Formation of UV-induced DNA damage contributing to skin cancer development. Photochem. Photobiol. Sci. 2018, 17, 1016–1841. [Google Scholar] [CrossRef] [PubMed]
- Mouret, S.; Philippe, C.; Gracia-Chantegrel, J.; Banyasz, A.; Karpati, S.; Markovitsi, D.; Douki, T. UVA-induced cyclobutane pyrimidine dimers in DNA: A direct photochemical mechanism? Org. Biomol. Chem. 2010, 8, 1706–1711. [Google Scholar] [CrossRef] [Green Version]
- Mouret, S.; Baudouin, C.; Charveron, M.; Favier, A.; Cadet, J.; Douki, T. Cyclobutane pyrimidine dimers are predominant DNA lesions in whole human skin exposed to UVA radiation. Proc. Natl. Acad. Sci. USA 2006, 103, 13765–13770. [Google Scholar] [CrossRef] [Green Version]
- Delinasios, G.J.; Karbaschi, M.; Cooke, M.S.; Young, A.R. Vitamin E inhibits the UVAI induction of “light” and “dark” cyclobutane pyrimidine dimers, and oxidatively generated DNA damage, in keratinocytes. Sci. Rep. 2018, 8, 423. [Google Scholar] [CrossRef] [Green Version]
- Gong, C.; Yang, Z.; Zhang, L.; Wang, Y.; Gong, W.; Liu, Y. Quercetin suppresses DNA double-strand break repair and enhances the radiosensitivity of human ovarian cancer cells via p53-dependent endoplasmic reticulum stress pathway. OncoTargets. Ther. 2018, 11, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Lo, H.L.; Nakajima, S.; Ma, L.; Walter, B.; Yasui, A.; Ethell, D.; Owen, L.B. Differential biologic effects of CPD and 6-4PP UV-induced DNA damage on the induction of apoptosis and cell-cycle arrest. BMC Cancer 2005, 5, 135. [Google Scholar] [CrossRef] [Green Version]
- Timocin, T.; Arslan, M.; Basri Ila, H. Evaluation of in vitro and in vivo genotoxic and antigenotoxic effects of Rhus coriaria. Drug Chem. Toxicol. 2019, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Svobodová, A.; Psotová, J.; Walterová, D. Natural phenolics in the prevention of UV-induced skin damage. A review. Biomed. Pap. Med. Fac. Univ. Palacky. Olomouc. Czech Repub. 2003, 147, 137–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masaki, H. Role of antioxidants in the skin: Anti-aging effects. J. Dermatol. Sci. 2010, 58, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Olchowik, E.; Sciepuk, A.; Mavlyanov, S.; Abdullajanova, N.; Zamaraeva, M. Antioxidant capacities of polyphenols from Sumac (Rhus typhina L.) leaves in protection of erythrocytes against oxidative damage. Biomed. Prev. Nutr. 2012, 2, 99–105. [Google Scholar] [CrossRef]
- Pandurangan, A.K.; Mohebali, N.; Norhaizan, M.E.; Looi, C.Y. Gallic acid attenuates dextran sulfate sodium-induced experimental colitis in balb/c mice. Drug Des. Dev. Ther. 2015, 9, 3923–3934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boo, Y.C. Can plant phenolic compounds protect the skin from airborne particulate matter? Antioxidants 2019, 8, 379. [Google Scholar] [CrossRef] [Green Version]
- Choi, D.R.; Jeong, J.H.; Yu, K.S.; Lee, N.S.; Jeong, Y.G.; Kim, D.K.; Na, C.S.; Na, D.S.; Hwang, W.M.; Han, S.Y. Extract of rhus verniciflua stokes protects against renal ischemia-reperfusion injury by enhancing nrf2-mediated induction of antioxidant enzymes. Exp. Ther. Med. 2018, 15, 3827–3835. [Google Scholar] [CrossRef] [Green Version]
- Douki, T.; Sage, E. Dewar valence isomers, the third type of environmentally relevant DNA photoproducts induced by solar radiation. Photochem. Photobiol. Sci. 2016, 15, 24–30. [Google Scholar] [CrossRef]
- Wondrak, G.T.; Jacobson, M.K.; Jacobson, E.L. Endogenous UVA-photosensitizers: Mediators of skin photodamage and novel targets for skin photoprotection. Photochem. Photobiol. Sci. 2006, 5, 215–237. [Google Scholar] [CrossRef]
- Pouget, J.P.; Douki, T.; Richard, M.J.; Cadet, J. DNA damage induced in cells by γ and UVA radiation as measured by HPLC/GC-MS and HPLC-EC and comet assay. Chem. Res. Toxicol. 2000, 13, 541–549. [Google Scholar] [CrossRef]
- Brash, D.E.; Rudolph, J.A.; Simon, J.A.; Lin, A.; Mckenna, G.J.; Baden, H.P.; Halperin, A.J.; Pontén, J. A role for sunlight in skin cancer: UV-induced p53 mutations in squamous cell carcinoma. Proc. Natl. Acad. Sci. USA 1991, 88, 10124–10128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasparrini, M.; Forbes-Hernandez, T.Y.; Afrin, S.; Reboredo-Rodriguez, P.; Cianciosi, D.; Mezzetti, B.; Quiles, J.L.; Bompadre, S.; Battino, M.; Giampieri, F. Strawberry-based cosmetic formulations protect human dermal fibroblasts against UVA-induced damage. Nutrients 2017, 9, 605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista, L.F.Z.; Kaina, B.; Meneghini, R.; Menck, C.F.M. How DNA lesions are turned into powerful killing structures: Insights from UV-induced apoptosis. Mutat. Res.-Rev. Mutat. Res. 2009, 681, 197–208. [Google Scholar] [CrossRef]
- D’Angleo, S.; Martino, E.; Ilisso, C.P.; Bagarolo, M.L.; Porcelli, M.; Cacciapuoti, G. Pro-oxidant and pro-apoptotic activity of polyphenol extract from Annurca apple and its underlying mechanisms in human breast cancer cells. Int. J. Oncol. 2017, 51, 939–948. [Google Scholar]
- Armentano, M.F.; Bisaccia, F.; Miglionico, R.; Russo, D.; Nolfi, N.; Carmosino, M.; Andrade, P.B.; Valentão, P.; Diop, M.S.; Milella, L. Antioxidant and proapoptotic activities of sclerocarya birrea [(A. Rich.) Hochst.] methanolic root extract on the hepatocellular carcinoma cell line HepG2. BioMed Res. Int. 2015, 2015, 561589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, F.J.; Martin, J.R.; Wulff, B.C.; Stoner, G.D.; Tober, K.L.; Oberyszyn, T.M.; Kusewitt, D.F.; Van Buskirk, A.M. Topical treatment with black raspberry extract reduces cutaneous UVB-induced carcinogenesis and inflammation. Cancer Prev. Res. 2009, 2, 665–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvo-Castro, L.; Syed, D.N.; Chamcheu, J.C.; Vilela, F.M.P.; Pérez, A.M.; Vaillant, F.; Rojas, M.; Mukhtar, H. Protective effect of tropical highland blackberry juice (Rubus adenotrichos Schltdl.) against UVB-mediated damage in human epidermal keratinocytes and in a reconstituted skin equivalent model. Photochem. Photobiol. 2013, 89, 1199–1207. [Google Scholar] [CrossRef]
- Marabini, L.; Melzi, G.; Lolli, F.; Dell’Agli, M.; Piazza, S.; Sangiovanni, E.; Marinovich, M. Effects of Vitis vinifera L. leaves extract on UV radiation damage in human keratinocytes (HaCaT). J. Photochem. Photobiol. B Biol. 2020, 204, 111810. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, R.; Liu, J.; Meng, D.; Zhou, Z.; Zhang, Y.; Blanchard, C. Fabrication, structure, and function evaluation of the ferritin-based nano-carrier for food bioactive compounds. Food Chem. 2019, 299, 125097. [Google Scholar] [CrossRef]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nozza, E.; Melzi, G.; Marabini, L.; Marinovich, M.; Piazza, S.; Khalilpour, S.; Dell’Agli, M.; Sangiovanni, E. Rhus coriaria L. Fruit Extract Prevents UV-A-Induced Genotoxicity and Oxidative Injury in Human Microvascular Endothelial Cells. Antioxidants 2020, 9, 292. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9040292
Nozza E, Melzi G, Marabini L, Marinovich M, Piazza S, Khalilpour S, Dell’Agli M, Sangiovanni E. Rhus coriaria L. Fruit Extract Prevents UV-A-Induced Genotoxicity and Oxidative Injury in Human Microvascular Endothelial Cells. Antioxidants. 2020; 9(4):292. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9040292
Chicago/Turabian StyleNozza, Emma, Gloria Melzi, Laura Marabini, Marina Marinovich, Stefano Piazza, Saba Khalilpour, Mario Dell’Agli, and Enrico Sangiovanni. 2020. "Rhus coriaria L. Fruit Extract Prevents UV-A-Induced Genotoxicity and Oxidative Injury in Human Microvascular Endothelial Cells" Antioxidants 9, no. 4: 292. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9040292