
Influence of a Heterologous (ChAdOx1-nCoV-19/BNT162b2) or Homologous (BNT162b2/BNT162b2) Vaccination Regimen on the Antibody and T Cell Response to a Third Vaccination with BNT162b2

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Antibody Assay
2.3. T Cell Assay
2.4. Statistical Analysis
3. Results
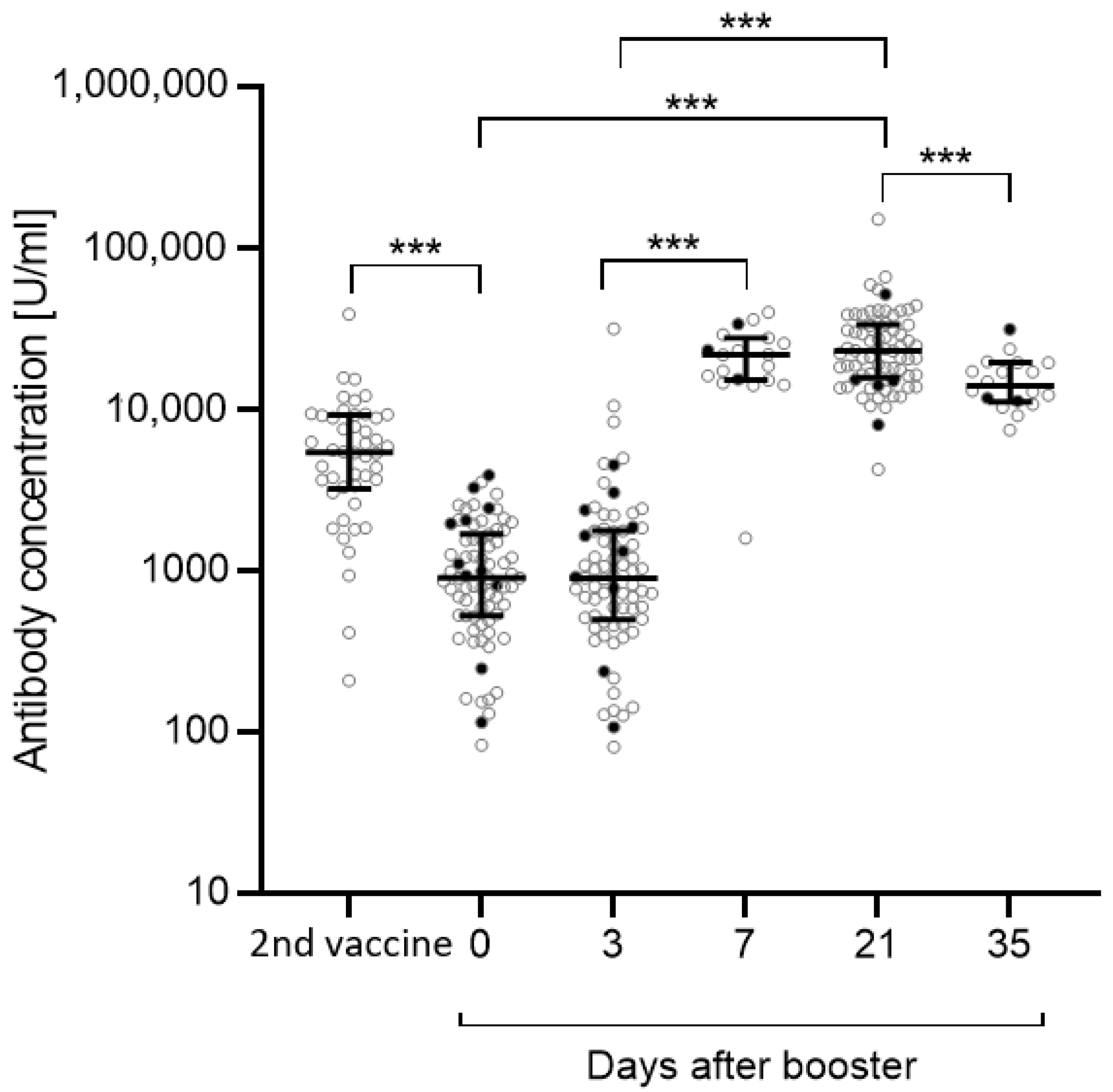
3.1. Antibody Concentrations
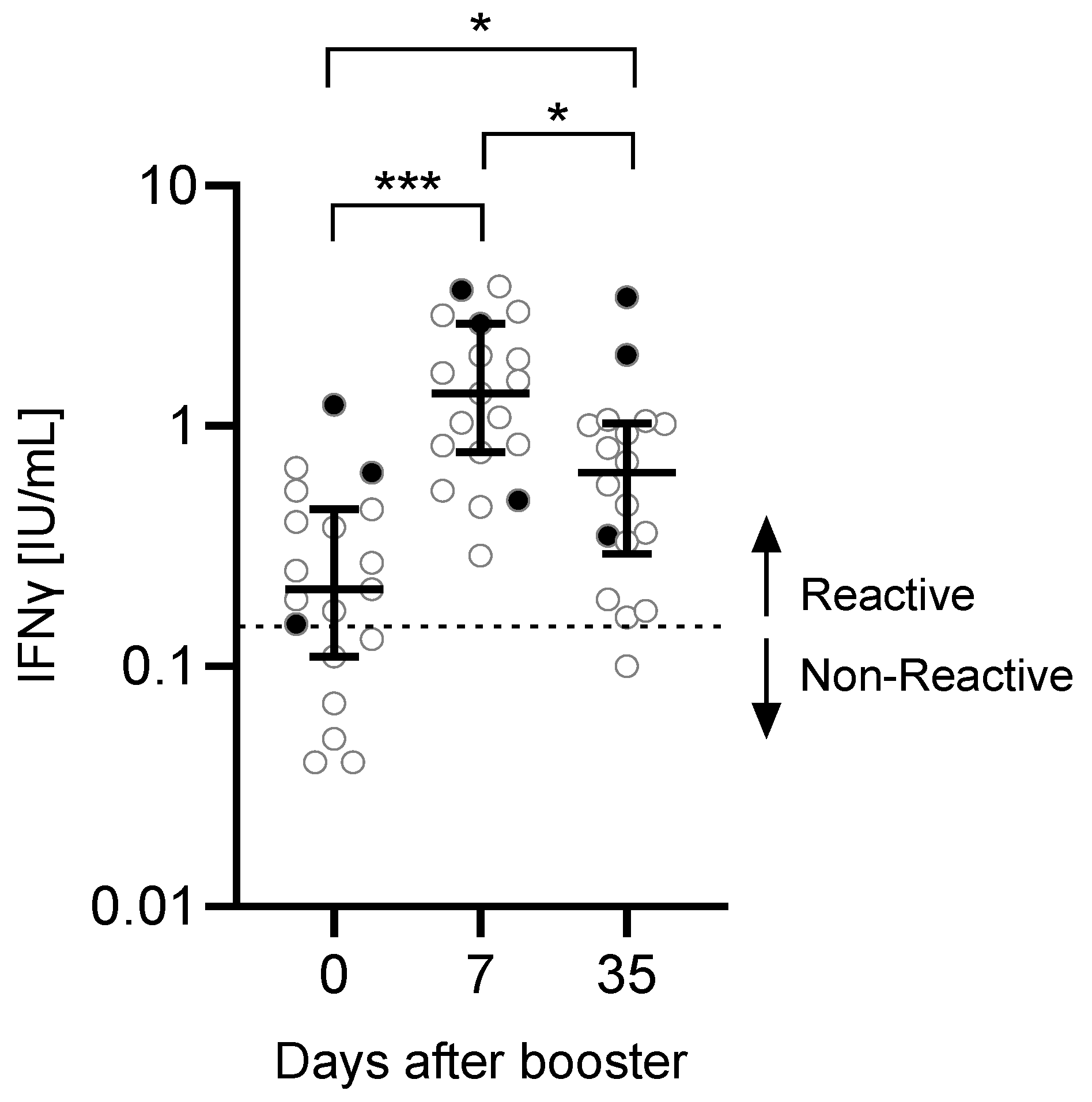
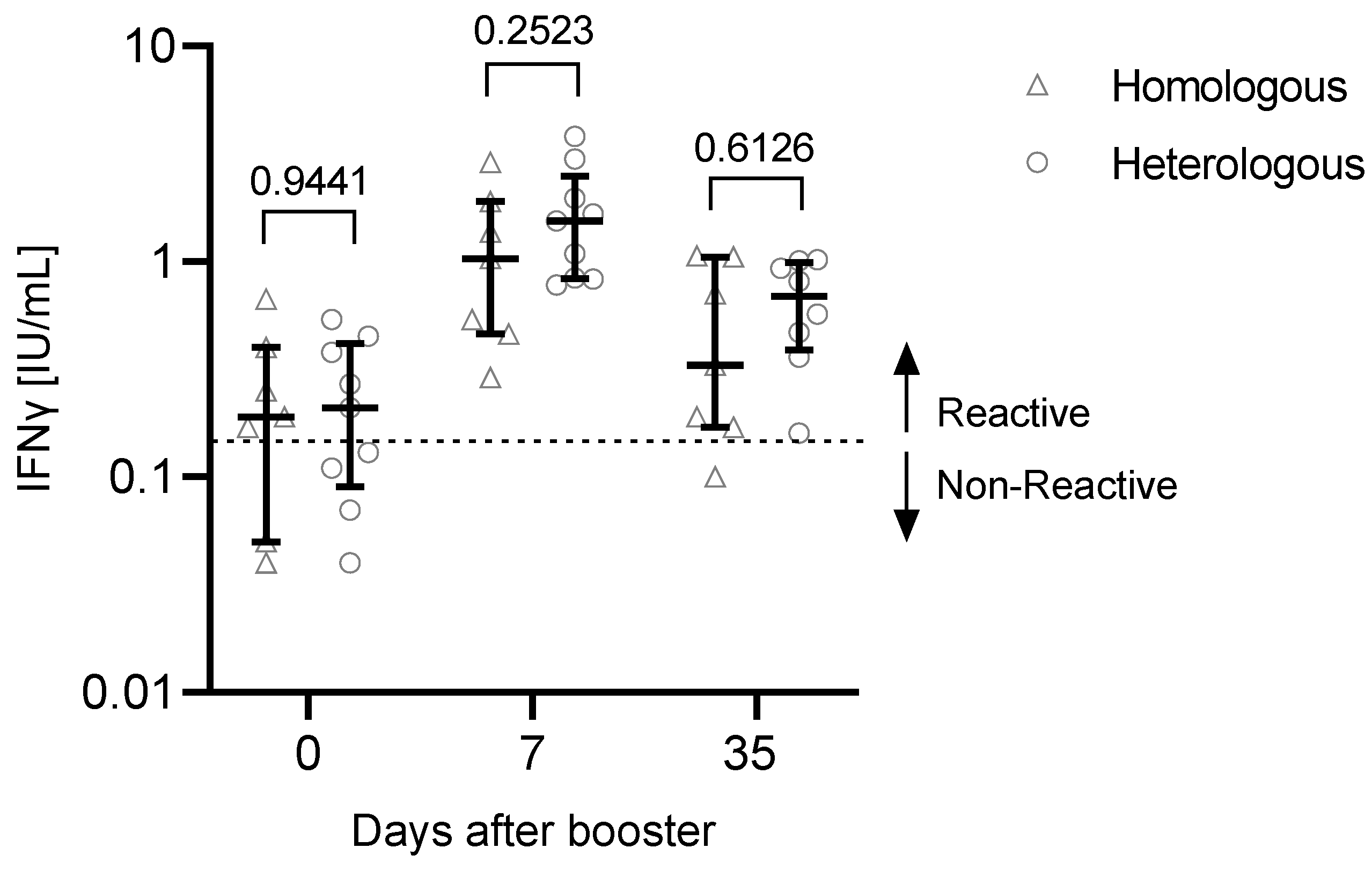
3.2. T Cell Response to the Third Vaccination
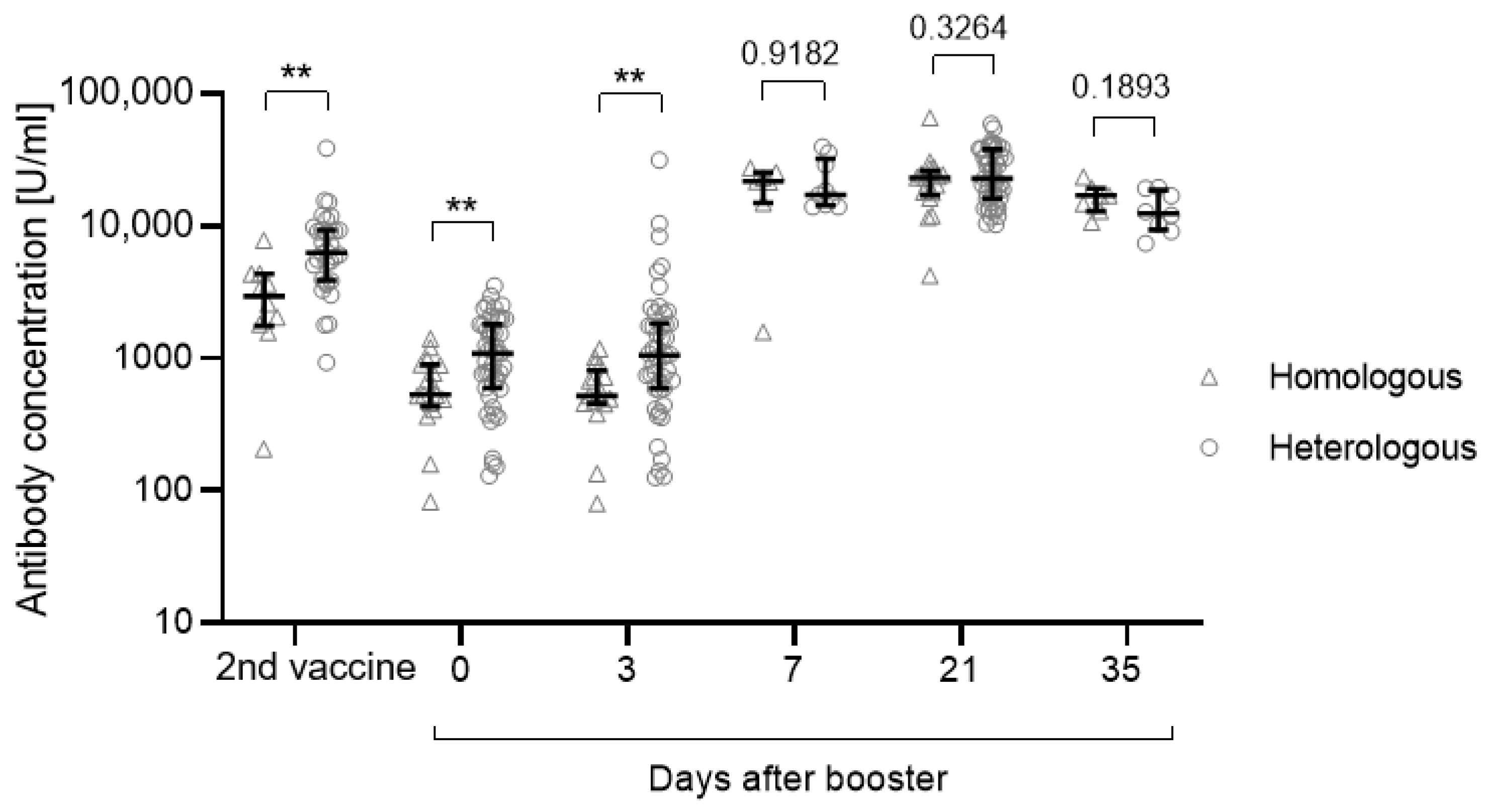
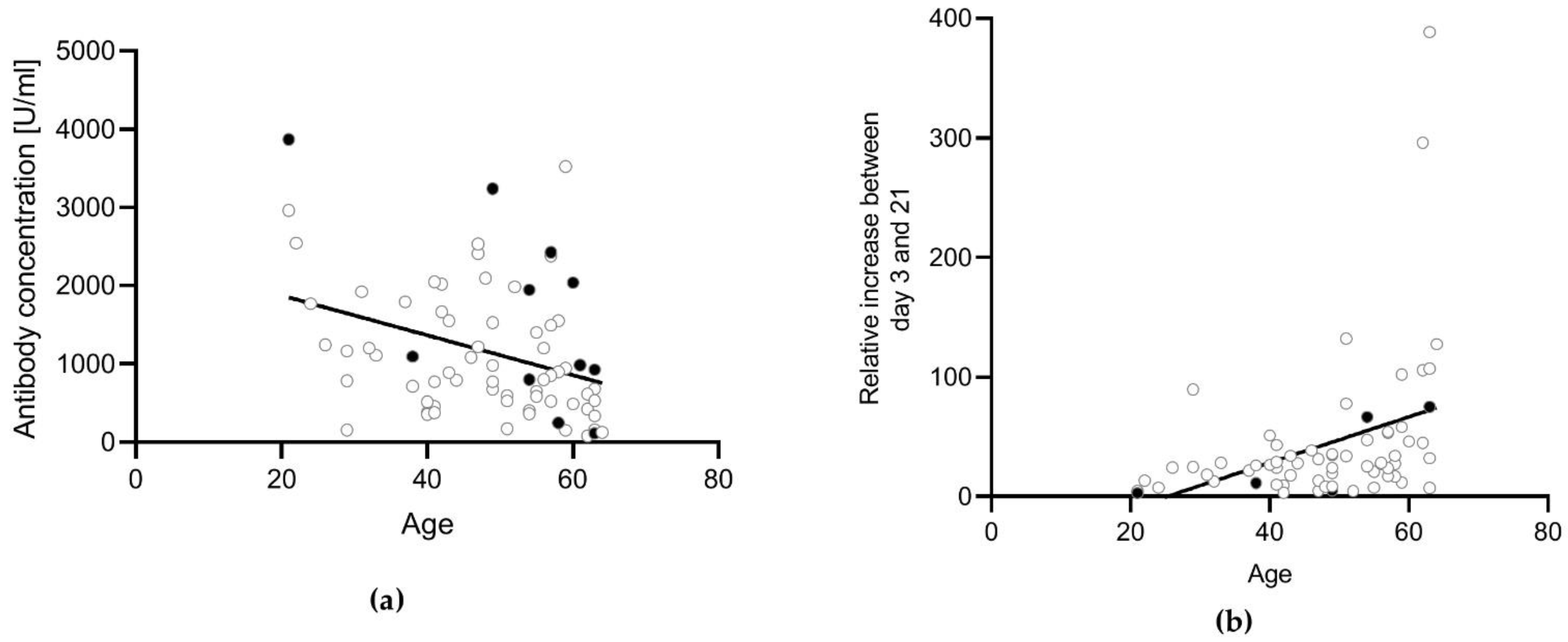
3.3. Influence of the Vaccination Regimen on the Antibody and T Cell Response and Influence of Gender and Age
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kuhlmann, C.; Mayer, C.K.; Claassen, M.; Maponga, T.; Burgers, W.A.; Keeton, R.; Riou, C.; Sutherland, A.D.; Suliman, T.; Shaw, M.L.; et al. Breakthrough Infections with SARS-CoV-2 Omicron Variant Despite Booster Dose of mRNA. Vaccine 2021, 399, 625–626. [Google Scholar] [CrossRef]
- Puhach, O.; Adea, K.; Hulo, N.; Sattonnet, P.; Genecand, C.; Iten, A.; Bausch, F.J.; Kaiser, L.; Vetter, P.; Eckerle, I.; et al. Infectious viral load in unvaccinated and vaccinated patients infected with SARS-CoV-2 WT, Delta and Omicron. MedRxiv 2022. [Google Scholar] [CrossRef]
- Arbel, R.; Hammerman, A.; Sergienko, R.; Friger, M.; Peretz, A.; Netzer, D.; Yaron, S. BNT162b2 Vaccine Booster and Mortality Due to Covid-19. N. Engl. J. Med. 2021, 385, 2413–2420. [Google Scholar] [CrossRef] [PubMed]
- Bar-On, Y.M.; Goldberg, Y.; Mandel, M.; Bodenheimer, O.; Freedman, L.; Alroy-Preis, S.; Ash, N.; Huppert, A.; Milo, R. Protection against Covid-19 by BNT162b2 Booster across Age Groups. N. Engl. J. Med. 2021, 385, 2421–2430. [Google Scholar] [CrossRef] [PubMed]
- Khoury, D.S.; Cromer, D.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Subbarao, K.; Kent, S.J.; Triccas, J.A.; Davenport, M.P. Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nat. Med. 2021, 27, 1205–1211. [Google Scholar] [CrossRef]
- Atmar, R.L.; Lyke, K.E.; Deming, M.E.; Jackson, L.A.; Branche, A.R.; El Sahly, H.M.; Rostad, C.A.; Martin, J.M.; Johnston, C.; Rupp, R.E.; et al. Homologous and Heterologous Covid-19 Booster Vaccinations. N. Engl. J. Med. 2022, 386, 1046–1057. [Google Scholar] [CrossRef] [PubMed]
- Rose, R.; Neumann, F.; Grobe, O.; Lorentz, T.; Fickenscher, H.; Krumbholz, A. Humoral immune response after different SARS-CoV-2 vaccination regimens. BMC Med. 2022, 20, 31. [Google Scholar] [CrossRef]
- He, Q.; Mao, Q.; An, C.; Zhang, J.; Gao, F.; Bian, L.; Li, C.; Liang, Z.; Xu, M.; Wang, J. Heterologous prime-boost: Breaking the protective immune response bottleneck of COVID-19 vaccine candidates. Emerg. Microbes Infect. 2021, 10, 629–637. [Google Scholar] [CrossRef]
- Stuart, A.S.V.; Shaw, R.H.; Liu, X.; Greenland, M.; Aley, P.K.; Andrews, N.J.; Cameron, J.C.; Charlton, S.; Clutterbuck, E.A.; Collins, A.M.; et al. Immunogenicity, safety, and reactogenicity of heterologous COVID-19 primary vaccination incorporating mRNA, viral-vector, and protein-adjuvant vaccines in the UK (Com-COV2): A single-blind, randomised, phase 2, non-inferiority trial. Lancet 2022, 399, 36–49. [Google Scholar] [CrossRef]
- Cheng, S.M.S.; Mok, C.K.P.; Leung, Y.W.Y.; Ng, S.S.; Chan, K.C.K.; Ko, F.W.; Chen, C.; Yiu, K.; Lam, B.H.S.; Lau, E.H.Y.; et al. Neutralizing antibodies against the SARS-CoV-2 Omicron variant following homologous and heterologous CoronaVac or BNT162b2 vaccination. Nat. Med. 2022, 28, 486–489. [Google Scholar] [CrossRef]
- Groß, R.; Zanoni, M.; Seidel, A.; Conzelmann, C.; Gilg, A.; Krnavek, D.; Erdemci-Evin, S.; Mayer, B.; Hoffmann, M.; Pöhlmann, S.; et al. Heterologous ChAdOx1 nCoV-19 and BNT162b2 prime-boost vaccination elicits potent neutralizing antibody responses and T cell reactivity against prevalent SARS-CoV-2 variants. EBioMedicine 2022, 75, 103761. [Google Scholar] [CrossRef] [PubMed]
- Romero-Olmedo, A.J.; Schulz, A.R.; Hochstätter, S.; Das Gupta, D.; Virta, I.; Hirseland, H.; Staudenraus, D.; Camara, B.; Münch, C.; Hefter, V.; et al. Induction of robust cellular and humoral immunity against SARS-CoV-2 after a third dose of BNT162b2 vaccine in previously unresponsive older adults. Nat. Microbiol. 2022, 7, 195–199. [Google Scholar] [CrossRef]
- Schmidt, T.; Klemis, V.; Schub, D.; Schneitler, S.; Reichert, M.C.; Wilkens, H.; Sester, U.; Sester, M.; Mihm, J. Cellular immunity predominates over humoral immunity after homologous and heterologous mRNA and vector-based COVID-19 vaccine regimens in solid organ transplant recipients. Am. J. Transplant. 2021, 21, 3990–4002. [Google Scholar] [CrossRef] [PubMed]
- Ledford, H. ‘Killer’ immune cells still recognize Omicron variant. Nature 2022, 601, 307. [Google Scholar] [CrossRef]
- Jaganathan, S.; Stieber, F.; Rao, S.N.; Nikolayevskyy, V.; Manissero, D.; Allen, N.; Boyle, J.; Howard, J. Preliminary Evaluation of QuantiFERON SARS-CoV-2 and QIAreach Anti-SARS-CoV-2 Total Test in Recently Vaccinated Individuals. Infect. Dis. Ther. 2021, 10, 2765–2776. [Google Scholar] [CrossRef] [PubMed]
- Tychala, A.; Meletis, G.; Katsimpourlia, E.; Gkeka, I.; Dimitriadou, R.; Sidiropoulou, E.; Skoura, L. Evaluation of the QuantiFERON SARS-CoV-2 assay to assess cellular immunogenicity of the BNT162b2 mRNA COVID-19 vaccine in individuals with low and high humoral response. Hum. Vaccines Immunother. 2021, 17, 5148–5149. [Google Scholar] [CrossRef] [PubMed]
- Van Praet, J.T.; Vandecasteele, S.; De Roo, A.; De Vriese, A.S.; Reynders, M. Humoral and Cellular Immunogenicity of the BNT162b2 Messenger RNA Coronavirus Disease 2019 Vaccine in Nursing Home Residents. Clin. Infect. Dis. 2021, 73, 2145–2147. [Google Scholar] [CrossRef] [PubMed]
- Costa Clemens, S.A.; Weckx, L.; Clemens, R.; Almeida Mendes, A.V.; Ramos Souza, A.; Silveira, M.B.V.; da Guarda, S.N.F.; de Nobrega, M.M.; Pinto, M.I.d.; Gonzalez, I.G.S.; et al. Heterologous versus homologous COVID-19 booster vaccination in previous recipients of two doses of CoronaVac COVID-19 vaccine in Brazil (RHH-001): A phase 4, non-inferiority, single blind, randomised study. Lancet 2022, 399, 521–529. [Google Scholar] [CrossRef]
- Pérez-Then, E.; Lucas, C.; Monteiro, V.S.; Miric, M.; Brache, V.; Cochon, L.; Vogels, C.B.F.; Malik, A.A.; de la Cruz, E.; Jorge, A.; et al. Neutralizing antibodies against the SARS-CoV-2 Delta and Omicron variants following heterologous CoronaVac plus BNT162b2 booster vaccination. Nat. Med. 2022, 28, 481–485. [Google Scholar] [CrossRef]
- Nomura, Y.; Sawahata, M.; Nakamura, Y.; Koike, R.; Katsube, O.; Hagiwara, K.; Niho, S.; Masuda, N.; Tanaka, T.; Sugiyama, K. Attenuation of Antibody Titers from 3 to 6 Months after the Second Dose of the BNT162b2 Vaccine Depends on Sex, with Age and Smoking Risk Factors for Lower Antibody Concentrations at 6 Months. Vaccines 2021, 9, 1500. [Google Scholar] [CrossRef]
- Klein, S.L.; Pekosz, A.; Park, H.-S.; Ursin, R.L.; Shapiro, J.R.; Benner, S.E.; Littlefield, K.; Kumar, S.; Naik, H.M.; Betenbaugh, M.J.; et al. Sex, age, and hospitalization drive antibody responses in a COVID-19 convalescent plasma donor population. J. Clin. Investig. 2020, 130, 6141–6150. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Phillips, D.J.; White, T.; Sayal, H.; Aley, P.K.; Bibi, S.; Dold, C.; Fuskova, M.; Gilbert, S.C.; Hirsch, I.; et al. Correlates of protection against symptomatic and asymptomatic SARS-CoV-2 infection. Nat. Med. 2021, 27, 2032–2040. [Google Scholar] [CrossRef] [PubMed]
- Post, N.; Eddy, D.; Huntley, C.v.S.; May, C.I.; Shrotri, M.; Leeman, D.; Rigby, S.; Williams, S.V.; Bermingham, W.H.; Kellam, P.; et al. Antibody response to SARS-CoV-2 infection in humans: A systematic review. PLoS ONE 2020, 15, e0244126. [Google Scholar] [CrossRef]
- Lee, B.; Ko, J.H.; Park, J.; Moon, H.W.; Baek, J.Y.; Jung, S.; Lim, H.Y.; Kim, K.C.; Huh, K.; Cho, S.Y.; et al. Estimating the Neutralizing Effect and Concentration Correlation of Semi-Quantitative Anti-SARS-CoV-2 Antibody Immunoassays. Front. Cell. Infect. Microbiol. 2022, 12, 822599. [Google Scholar] [CrossRef] [PubMed]
- GeurtsvanKessel, C.H.; Geers, D.; Schmitz, K.S.; Mykytyn, A.Z.; Lamers, M.M.; Bogers, S.; Scherbeijn, S.; Gommers, L.; Sablerolles, R.S.G.; Nieuwkoop, N.N.; et al. Divergent SARS CoV-2 Omicron-reactive T- and B cell responses in COVID-19 vaccine recipients. Sci. Immunol. 2022, 7, eabo2202. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | |
|---|---|
| Gender: m/f | 15/60 |
| Vaccination: Homologous/Heterologous/ COVID-19 + vaccination | 20/53/2 |
| Previous infection: yes/no/unknown | 5/64/6 |
| Median (IQR) | |
| Age | 51 (41–58) |
| AB after second vaccination | 5365 (3281–9024) |
| AB day 0 | 895 (524–1609) |
| AB day 3 | 890 (500–1743) |
| AB day 21 | 22,801 (15,990–33,055) |
| Variable | |
|---|---|
| Gender: m/f | 5/14 |
| Vaccination: Homologous/Heterologous/ COVID-19 + vaccination | 7/10/2 |
| Previous infection: yes/no/unknown | 2/16/1 |
| Median (IQR) | |
| Age | 43 (41–56) |
| AB after second vaccination | 4950 (3705–8524) |
| AB day 0 | 1110 (711–1609) |
| T cells day 0 [IFNγ IU/mL] | 0.21 (0.12–0.425) |
| AB day 3 | 1054 (669–1779) |
| AB day 7 | 21,503 (15,156–26,486) |
| T cells day 7 [IFNγ IU/mL] | 1.37 (0.805–2.32) |
| AB day 21 | 18,319 (14,254–26,486) |
| AB day 35 | 13,853 (11,278–18,707) |
| T cells day 35 [IFNγ IU/mL] | 0.64 (0.335–1.018) |
| Variable | Homologous Vaccination | Heterologous Vaccination |
|---|---|---|
| 17 participants | 47 participants | |
| Age, median (IQR) | 55 (49–60) | 47 (40–57) |
| Gender: m/f | 2/15 | 11/36 |
| AB second vacc., median (IQR) | 2948 (1875–4169) | 6306 (3913–9293) |
| AB day 0, median (IQR) | 532 (458–888) | 1087 (623–1780) |
| AB day 3, median (IQR) | 520 (459–723) | 1054 (589–1817) |
| AB day 21, median (IQR) | 23,147 (17,884–24,630) | 22,957 (16,337–38,068) |
| Variable | Homologous Vaccination | Heterologous Vaccination |
|---|---|---|
| 7 participants | 9 participants | |
| Age, median (IQR) | 55 (49–57) | 41 (33–43) |
| Gender: m/f | 1/6 | 3/6 |
| T cells day 0 [IFNγ IU/mL], median (IQR) | 0.19 (0.11–0.325) | 0.21 (0.11–0.38) |
| T cells day 7 [IFNγ IU/mL], median (IQR) | 1.03 (0.5–1.635) | 1.54 (0.84–1.97) |
| T cells day 35 [IFNγ IU/mL], median (IQR) | 0.33 (0.18–0.88) | 0.69 (0.443–0.95) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reiter, R.; Von Blanckenburg, P.; Mutters, R.; Thiemer, J.; Geßner, R.; Seifart, U. Influence of a Heterologous (ChAdOx1-nCoV-19/BNT162b2) or Homologous (BNT162b2/BNT162b2) Vaccination Regimen on the Antibody and T Cell Response to a Third Vaccination with BNT162b2. Vaccines 2022, 10, 788. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines10050788
Reiter R, Von Blanckenburg P, Mutters R, Thiemer J, Geßner R, Seifart U. Influence of a Heterologous (ChAdOx1-nCoV-19/BNT162b2) or Homologous (BNT162b2/BNT162b2) Vaccination Regimen on the Antibody and T Cell Response to a Third Vaccination with BNT162b2. Vaccines. 2022; 10(5):788. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines10050788
Chicago/Turabian StyleReiter, Rieke, Pia Von Blanckenburg, Reinier Mutters, Julia Thiemer, Reinhard Geßner, and Ulf Seifart. 2022. "Influence of a Heterologous (ChAdOx1-nCoV-19/BNT162b2) or Homologous (BNT162b2/BNT162b2) Vaccination Regimen on the Antibody and T Cell Response to a Third Vaccination with BNT162b2" Vaccines 10, no. 5: 788. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines10050788