
SupT1 Cell Infusion as a Possible Cell-Based Therapy for HIV: Results from a Pilot Study in Hu-PBMC BRGS Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Cells
2.3. HIV Virus
2.4. Treatment and Control Groups
2.5. Blood Sampling of the Animals
2.6. Mouse Features
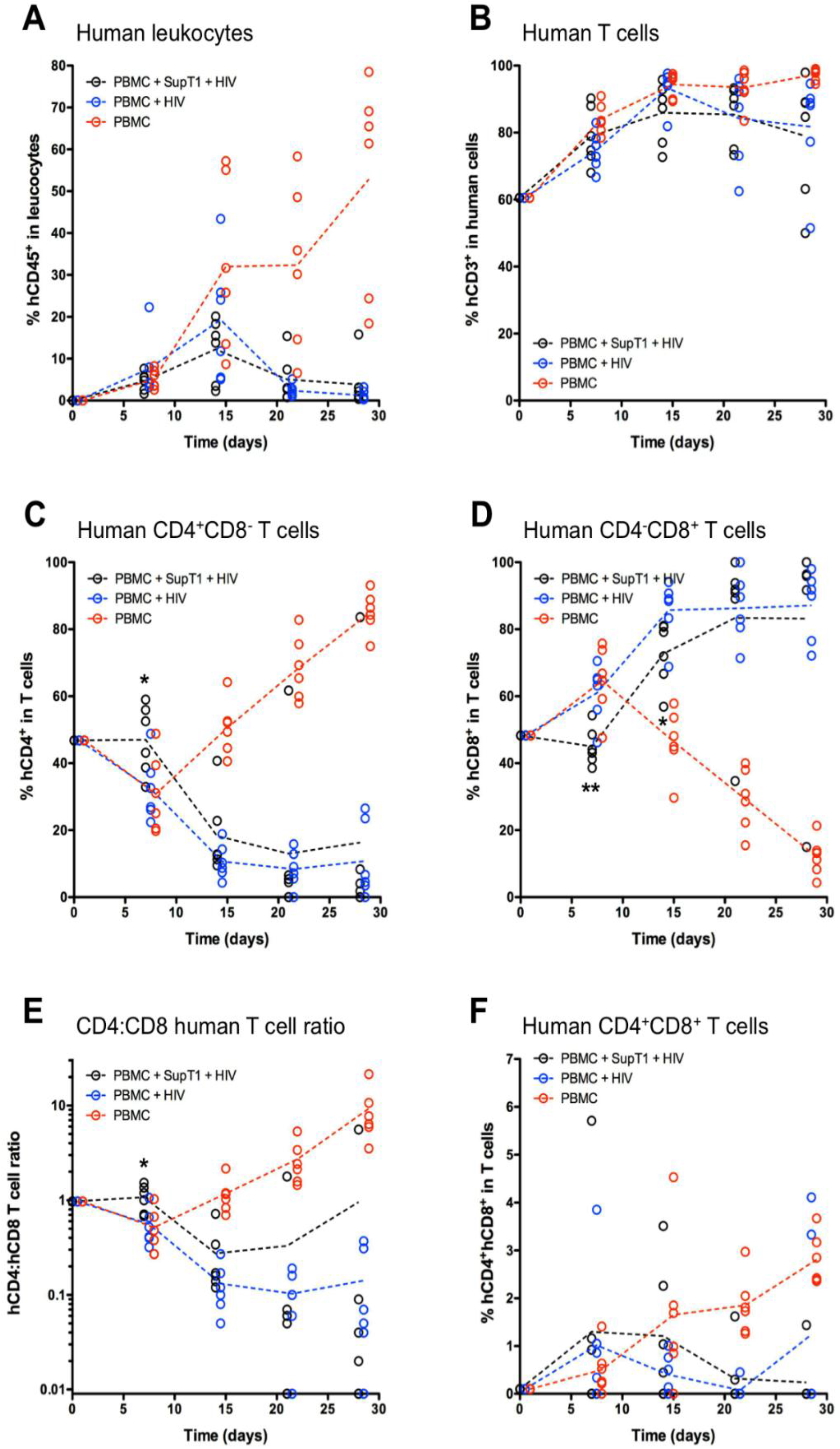
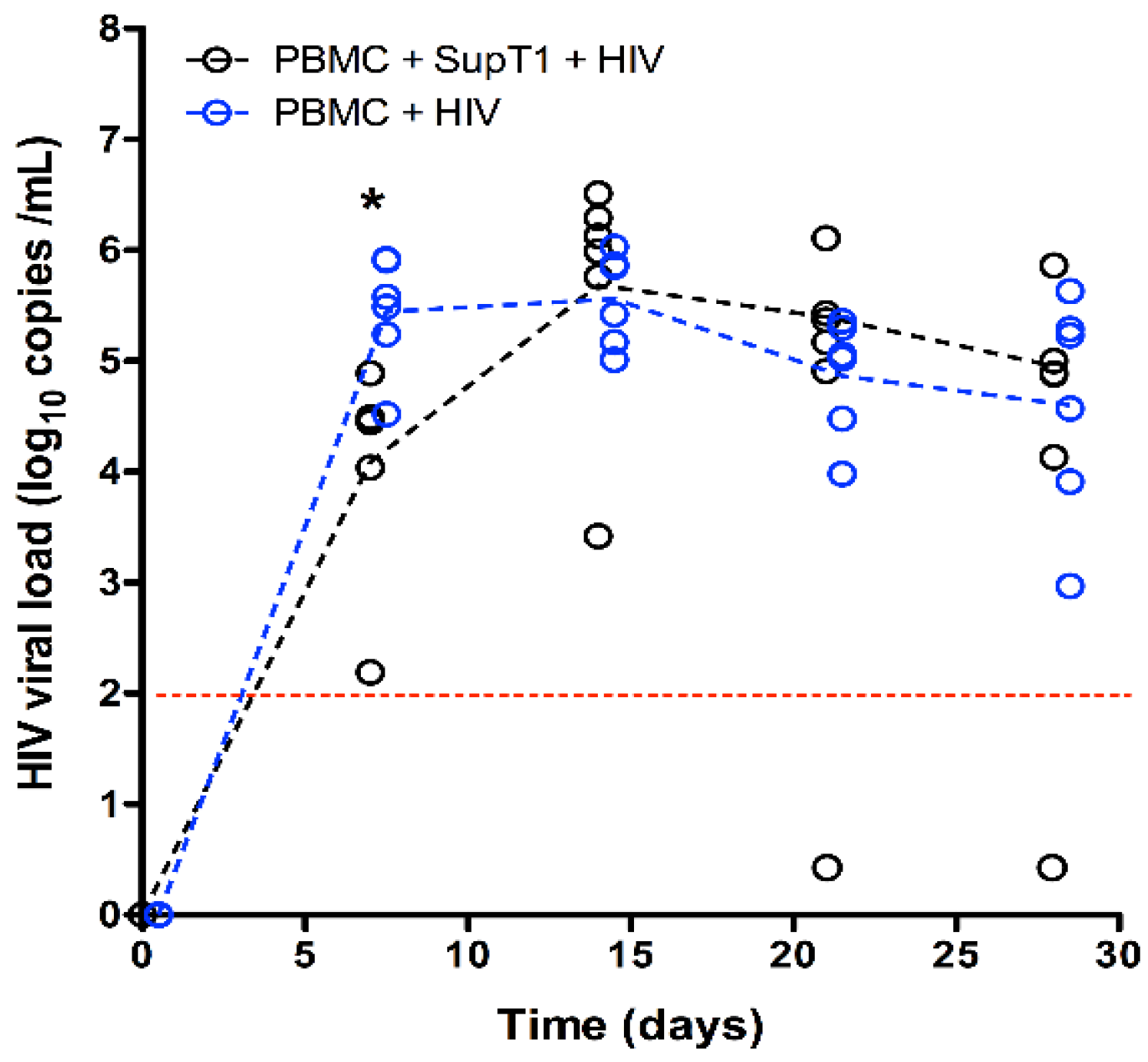
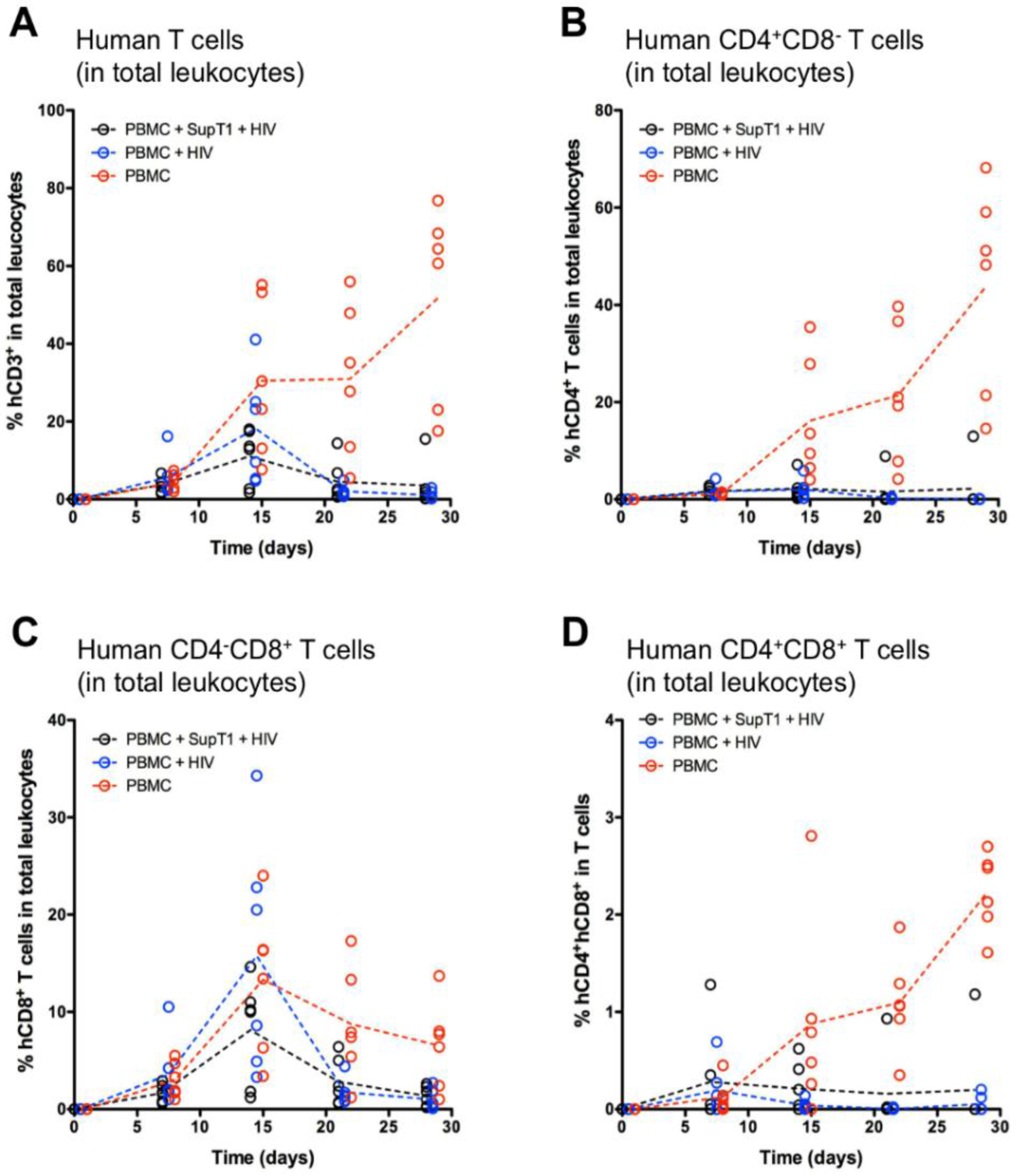
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fior, J. An initial in vitro investigation into the potential therapeutic use of SupT1 cells to prevent AIDS in HIV-seropositive individuals. PLoS ONE 2012, 7, e37511. [Google Scholar] [CrossRef] [PubMed]
- Das, A.T.; Land, A.; Braakman, I.; Klaver, B.; Berkhout, B. HIV-1 evolves into a nonsyncytium-inducing virus upon prolonged culture in vitro. Virology 1999, 263, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.P.; Burkly, L.C.; Connor, R.I.; Cao, Y.; Tizard, R.; Ho, D.D.; Fisher, R.A. Adaptation of two primary human immunodeficiency virus type 1 isolates to growth in transformed T cell lines correlates with alterations in the responses of their envelope glycoproteins to soluble CD4. AIDS Res. Hum. Retroviruses 1993, 9, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Turner, S.; Tizard, R.; DeMarinis, J.; Pepinsky, R.B.; Zullo, J.; Schooley, R.; Fisher, R. Resistance of primary isolates of human immunodeficiency virus type 1 to neutralization by soluble CD4 is not due to lower affinity with the viral envelope glycoprotein gp120. Proc. Natl. Acad. Sci. USA 1992, 89, 1335–1339. [Google Scholar] [CrossRef] [PubMed]
- Gabuzda, D.H.; Lawrence, K.; Langhoff, E.; Terwilliger, E.; Dorfman, T.; Haseltine, W.A.; Sodroski, J. Role of vif in replication of human immunodeficiency virus type 1 in CD4+ T lymphocytes. J. Virol. 1992, 66, 6489–6495. [Google Scholar] [PubMed]
- Wang, Y.J.; Wang, X.; Zhang, H.; Zhou, L.; Liu, S.; Kolson, D.L.; Song, L.; Ye, L.; Ho, W.Z. Expression and regulation of antiviral protein APOBEC3G in human neuronal cells. J. Neuroimmunol. 2009, 206, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Nathans, R.; Cao, H.; Sharova, N.; Ali, A.; Sharkey, M.; Stranska, R.; Stevenson, M.; Rana, T.M. Small-molecule inhibition of HIV-1 Vif. Nat. Biotechnol. 2008, 26, 1187–1192. [Google Scholar] [CrossRef] [PubMed]
- Cen, S.; Peng, Z.G.; Li, X.Y.; Li, Z.R.; Ma, J.; Wang, Y.M.; Fan, B.; You, X.F.; Wang, Y.P.; Liu, F.; et al. Small molecular compounds inhibit HIV-1 replication through specifically stabilizing APOBEC3G. J. Biol. Chem. 2010, 285, 16546–16552. [Google Scholar] [CrossRef] [PubMed]
- Zuo, T.; Liu, D.; Lv, W.; Wang, X.; Wang, J.; Lv, M.; Huang, W.; Wu, J.; Zhang, H.; Jin, H.; et al. Small-molecule inhibition of human immunodeficiency virus type 1 replication by targeting the interaction between Vif and ElonginC. J. Virol. 2012, 86, 5497–5507. [Google Scholar] [CrossRef] [PubMed]
- Jeeninga, R.E.; Jan, B.; van der Linden, B.; van den Berg, H.; Berkhout, B. Construction of a minimal HIV-1 variant that selectively replicates in leukemic derived T-cell lines: towards a new virotherapy approach. Cancer Res. 2005, 65, 3347–3355. [Google Scholar] [PubMed]
- Fior, J. Is a pacific coexistence between virus and host the unexploited path that may lead to an HIV functional cure? Viruses 2013, 5, 753–757. [Google Scholar] [CrossRef] [PubMed]
- Legrand, N.; Huntington, N.D.; Nagasawa, M.; Bakker, A.Q.; Schotte, R.; Strick-Marchand, H.; de Geus, S.J.; Pouw, S.M.; Böhne, M.; Voordouw, A.; et al. Functional CD47/signal regulatory protein alpha (SIRP(alpha)) interaction is required for optimal human T- and natural killer- (NK) cell homeostasis in vivo. Proc. Natl. Acad. Sci. USA 2011, 108, 13224–13229. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.D.; Shatsky, M.; Cohen, P.S.; Warnke, R.; Link, M.P.; Glader, B.E. Monoclonal antibody and enzymatic profiles of human malignant T-lymphoid cells and derived cell lines. Cancer Res. 1984, 44, 5657–5660. [Google Scholar] [PubMed]
- Rizza, P.; Santini, S.M.; Logozzi, M.A.; Lapenta, C.; Sestili, P.; Gherardi, G.; Lande, R.; Spada, M.; Parlato, S.; Belardelli, F.; et al. T-cell dysfunctions in hu-PBL-SCID mice infected with human immunodeficiency virus (HIV) shortly after reconstitution: in vivo effects of HIV on highly activated human immune cells. J. Virol. 1996, 70, 7958–7964. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Group | n | T0 | T + 2 hours | T + 4 hours | T + 1 week | T + 2 weeks | T + 3 weeks |
|---|---|---|---|---|---|---|---|
| A | 6 | 20 × 106 PBMC | 40 × 106 SupT1 | 105 TCID50 HIV LAI | 40 × 106 SupT1 | 40 × 106 SupT1 | 40 × 106 SupT1 |
| B (pos-CT) | 6 | 20 × 106 PBMC | - | 105 TCID50 HIV LAI | - | - | - |
| C (neg-CT) | 6 | 20 × 106 PBMC | - | - | - | - | - |
| Target | Label | Clone | Origin | Dilution |
|---|---|---|---|---|
| hCD45 | PerCP | H130 | BioLegend | 1:50 |
| mCD45 | APC E780 | 30-F11 | eBioscience | 1:200 |
| hCD3 | E450 | UCHT1 | eBioscience | 1:50 |
| hCD4 | PE | MEM-241 | ImmunoTools | 1:25 |
| hCD8 | FITC | MEM-31 | ImmunoTools | 1:12.5 |
| Group | A | B | C |
|---|---|---|---|
| Content | PBMC/SupT1/HIV | PBMC/HIV | PBMC |
| HIV-1 Status | + | + | − |
| # mice | 6 | 6 | 6 |
| Mouse ID (age at onset) | F 40 (17 weeks) | F 43 (19 weeks) | F 46 (19 weeks) |
| F 41 (17 weeks) | F 44 (19 weeks) | F 47 (19 weeks) | |
| F 42 (17 weeks) | F 45 (19 weeks) | F 48 (19 weeks) | |
| M 49 (15 weeks) | M 52 (15 weeks) | M 55 (15 weeks) | |
| M 50 (15 weeks) | M 53 (15 weeks) | M 56 (15 weeks) | |
| M 51 (15 weeks) | M 54 (15 weeks) | M 57 (15 weeks) |
© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fior, J. SupT1 Cell Infusion as a Possible Cell-Based Therapy for HIV: Results from a Pilot Study in Hu-PBMC BRGS Mice. Vaccines 2016, 4, 13. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines4020013
Fior J. SupT1 Cell Infusion as a Possible Cell-Based Therapy for HIV: Results from a Pilot Study in Hu-PBMC BRGS Mice. Vaccines. 2016; 4(2):13. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines4020013
Chicago/Turabian StyleFior, Jonathan. 2016. "SupT1 Cell Infusion as a Possible Cell-Based Therapy for HIV: Results from a Pilot Study in Hu-PBMC BRGS Mice" Vaccines 4, no. 2: 13. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines4020013