
Effects of KLK Peptide on Adjuvanticity of Different ODN Sequences

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Adjuvants
2.3. Immunization and Sample Collection
2.4. Adaptive Immune Responses
2.5. Innate Immune Responses
2.6. Statistical Analysis
3. Results
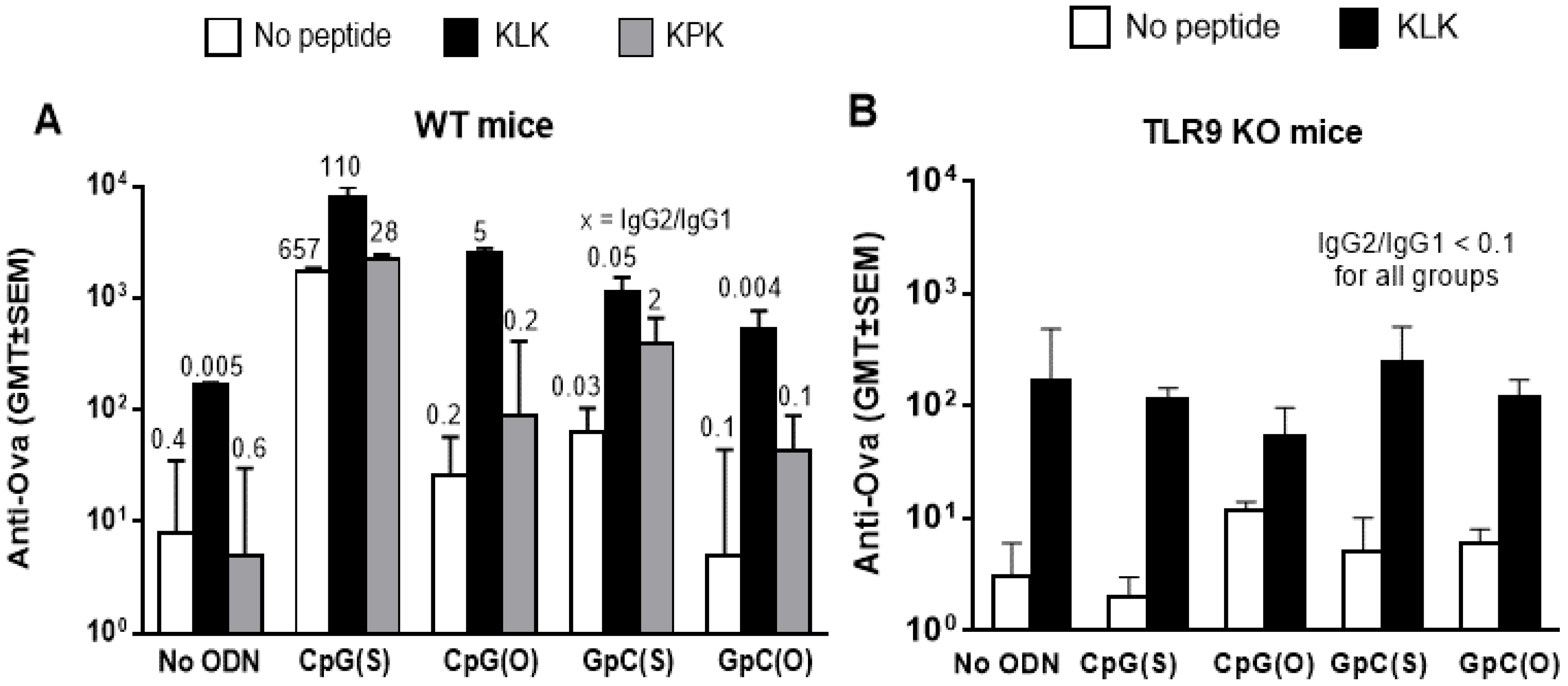
3.1. Humoral Immunity
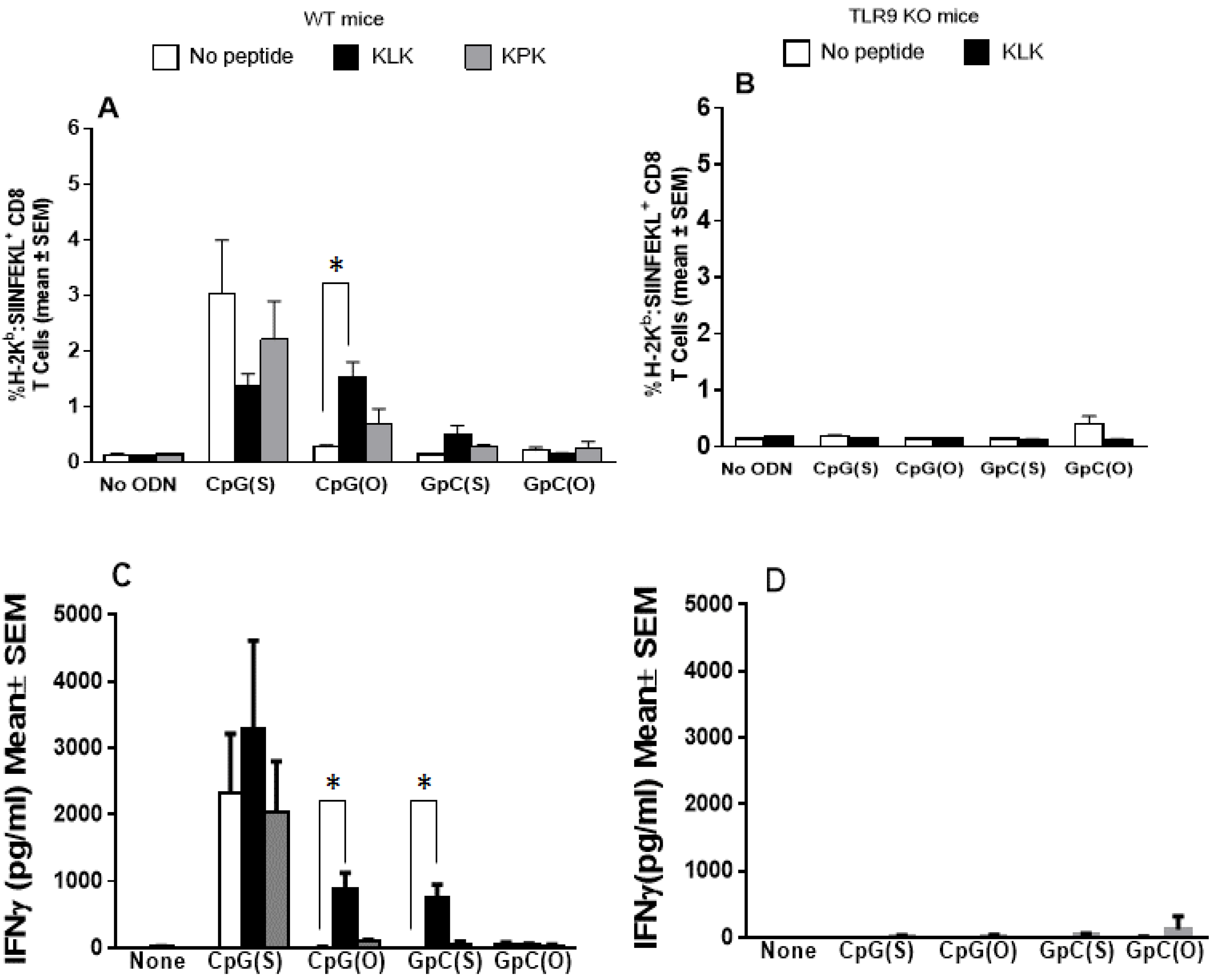
3.2. Cellular Immunity
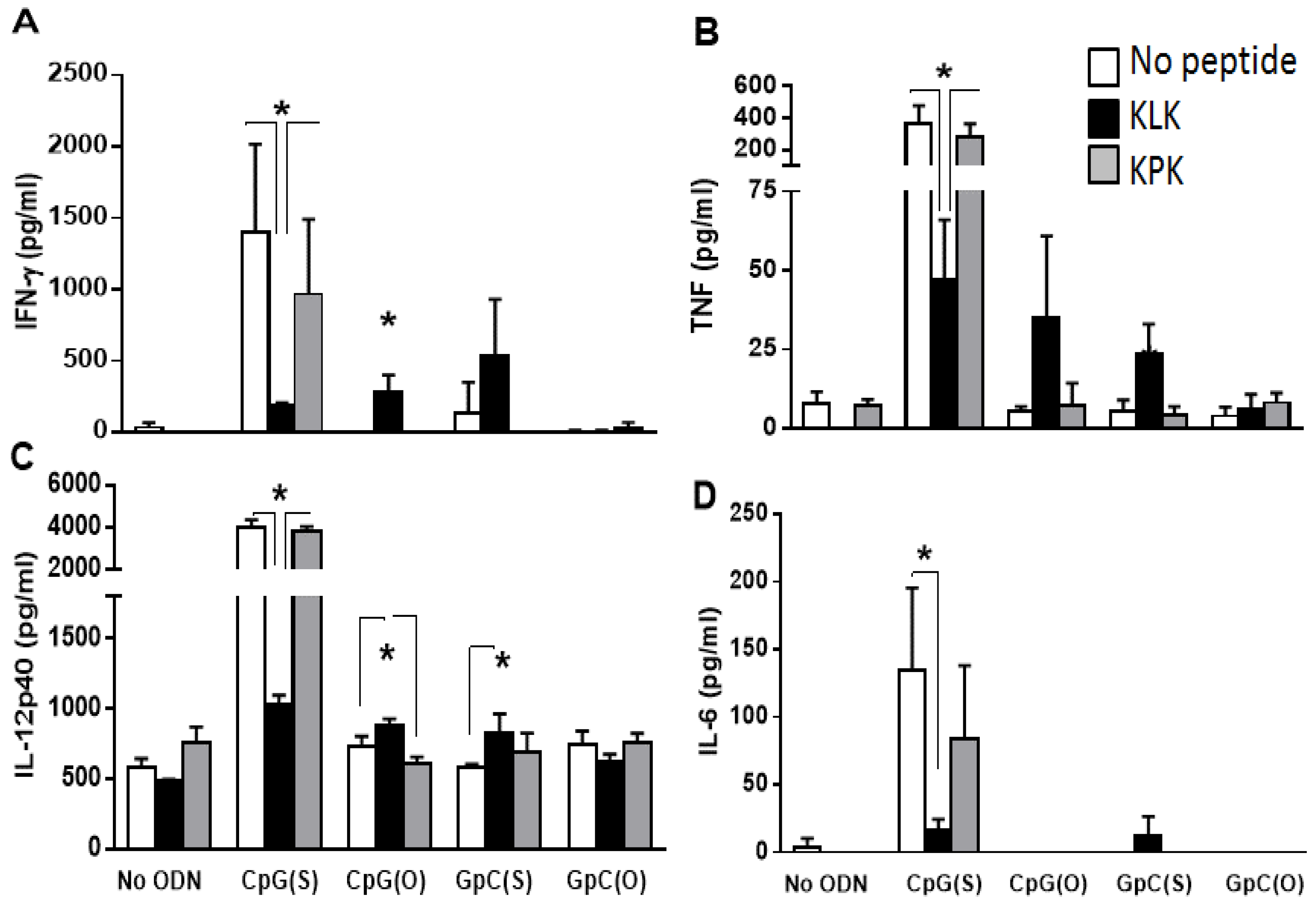
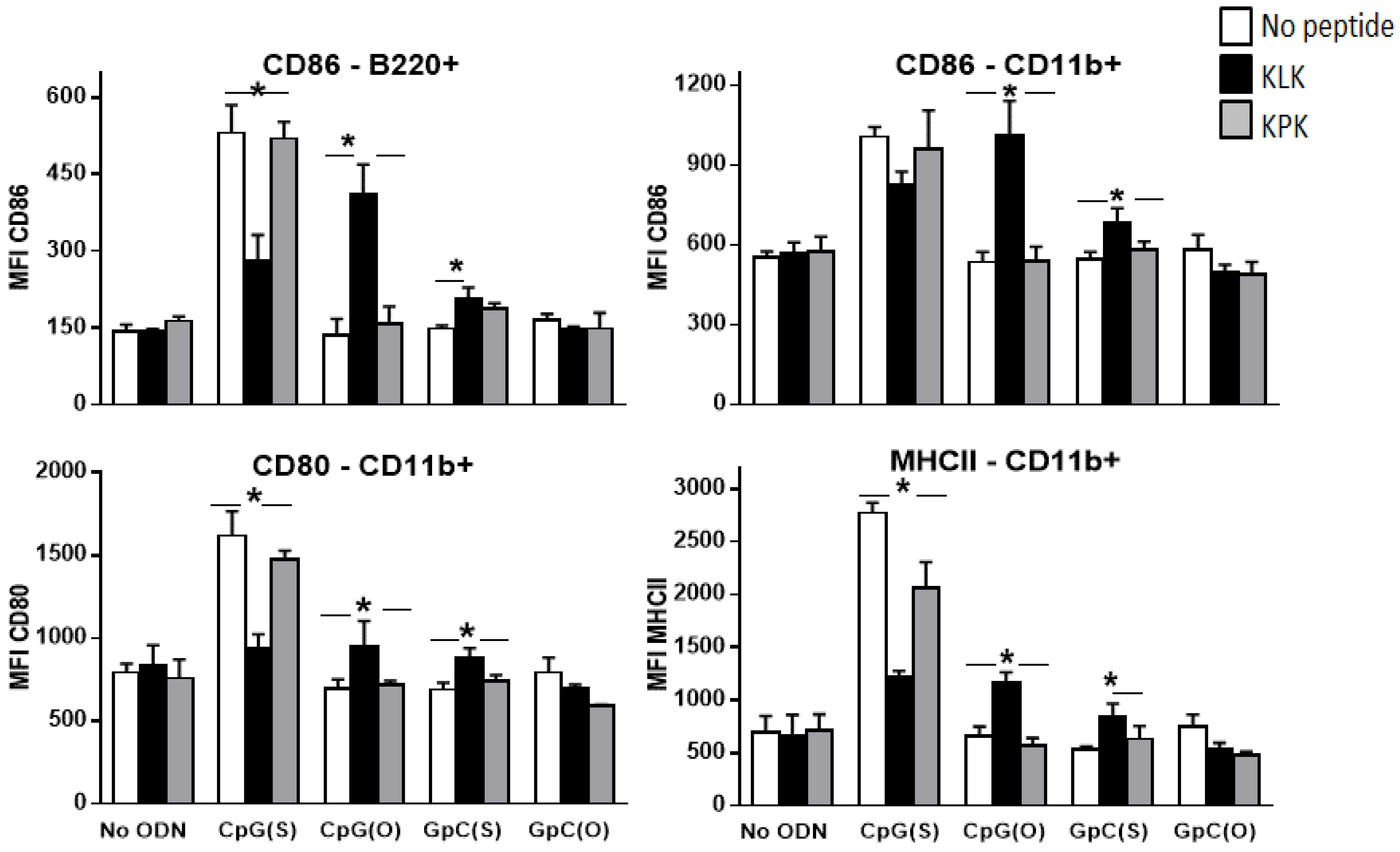
3.3. Impact of KLK on Innate Immune Stimulation
3.4. Impact of KLK on Intracellular Delivery of CpG(S)
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| TLR | Toll Like Receptor |
| APC | Antigen Presenting Cell |
| ODN | Oligideoxynucleotide |
| ELISpot | Enzyme-Linked Immunospot |
| ELISA | Enzyme-Linked Immunosorbent Assay |
| WT | Wild Type |
| FAM | Fluorescein Amidite |
| OVA | Ovalbumin |
| DC | Dendritic Cell |
References
- Dalpke, A.H.; Zimmermann, S.; Albrecht, I.; Heeg, K. Phosphodiester CpG oligonucleotides as adjuvants: polyguanosine runs enhance cellular uptake and improve immunostimulative activity of phosphodiester CpG oligonucleotides in vitro and in vivo. Immunology 2002, 106, 102–112. [Google Scholar]
- Krieg, A.M. The role of CpG motifs in innate immunity. Curr. Opin. Immunol. 2000, 12, 35–43. [Google Scholar] [CrossRef]
- Takeshita, F.; Leifer, C.A.; Gursel, I.; Ishii, K.J.; Takeshita, S.; Gursel, M.; Klinman, D.M. Cutting edge: Role of Toll-like receptor 9 in CpG DNA-induced activation of human cells. J. Immunol. 2001, 167, 3555–3558. [Google Scholar] [CrossRef] [PubMed]
- Splith, K.; Neundorf, I. Antimicrobial peptides with cell-penetrating peptide properties and vice versa. Eur. Biophys. J. 2011, 40, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.H.; Homma, K.; Kanegasaki, S.; Natori, S. Activation of human neutrophils by a synthetic anti-microbial peptide, KLKLLLLLKLK-NH2, via cell surface calreticulin. Eur. J. Biochem. 1999, 266, 878–885. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.H.; Homma, K.J.; Kanegasaki, S.; Natori, S. Activation of human monocyte cell line U937 via cell surface calreticulin. Cell Stress Chaperones 2001, 6, 148–152. [Google Scholar] [CrossRef]
- Fritz, J.H.; Brunner, S.; Birnstiel, M.L.; Buschle, M.; Gabain, A.; Mattner, F.; Zauner, W. The artificial antimicrobial peptide KLKLLLLLKLK induces predominantly a TH2-type immune response to co-injected antigens. Vaccine 2004, 22, 3274–3284. [Google Scholar] [CrossRef] [PubMed]
- Aagaard, C.; Hoang, T.T.; Izzo, A.; Billeskov, R.; Troudt, J.; Arnett, K.; Keyser, A.; Elvang, T.; Andersen, P.; Dietrich, J. Protection and polyfunctional T cells induced by Ag85B-TB10.4/IC31 against Mycobacterium tuberculosis is highly dependent on the antigen dose. PLoS ONE 2009, 4, e5930. [Google Scholar] [CrossRef] [PubMed]
- Riedl, K.; Riedl, R.; von Gabain, A.; Nagy, E.; Lingnau, K. The novel adjuvant IC31 strongly improves influenza vaccine-specific cellular and humoral immune responses in young adult and aged mice. Vaccine 2008, 26, 3461–3468. [Google Scholar] [CrossRef] [PubMed]
- Kamath, A.T.; Valenti, M.P.; Rochat, A.F.; Agger, E.M.; Lingnau, K.; von, G.A.; Andersen, P.; Lambert, P.H.; Siegrist, C.A. Protective anti-mycobacterial T cell responses through exquisite in vivo activation of vaccine-targeted dendritic cells. Eur. J. Immunol. 2008, 38, 1247–1256. [Google Scholar] [CrossRef] [PubMed]
- Kamath, A.T.; Rochat, A.F.; Valenti, M.P.; Agger, E.M.; Lingnau, K.; Andersen, P.; Lambert, P.H.; Siegrist, C.A. Adult-like anti-mycobacterial T cell and in vivo dendritic cell responses following neonatal immunization with Ag85B-ESAT-6 in the IC31 adjuvant. PLoS ONE 2008, 3, e3683. [Google Scholar] [CrossRef] [PubMed]
- Olafsdottir, T.A.; Lingnau, K.; Nagy, E.; Jonsdottir, I. IC31, a two-component novel adjuvant mixed with a conjugate vaccine enhances protective immunity against pneumococcal disease in neonatal mice. Scand. J. Immunol. 2009, 69, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Geldenhuys, H.; Mearns, H.; Miles, D.J.; Tameris, M.; Hokey, D.; Shi, Z.; Bennett, S.; Andersen, P.; Kromann, I.; Hoff, S.T.; et al. The tuberculosis vaccine H4:IC31 is safe and induces a persistent polyfunctional CD4 T cell response in South African adults: A randomized controlled trial. Vaccine 2015, 33, 3592–3599. [Google Scholar] [CrossRef] [PubMed]
- Lenz, N.; Schindler, T.; Kagina, B.M.; Zhang, J.D.; Lukindo, T.; Mpina, M.; Bang, P.; Kromann, I.; Hoff, S.T.; Andersen, P.; et al. Antiviral Innate Immune Activation in HIV-Infected Adults Negatively Affects H1/IC31-Induced Vaccine-Specific Memory CD4+ T Cells. Clin. Vaccine Immunol. 2015, 22, 688–696. [Google Scholar] [CrossRef] [PubMed]
- Luabeya, A.K.; Kagina, B.M.; Tameris, M.D.; Geldenhuys, H.; Hoff, S.T.; Shi, Z.; Kromann, I.; Hatherill, M.; Mahomed, H.; Hanekom, W.A.; et al. First-in-human trial of the post-exposure tuberculosis vaccine H56:IC31 in Mycobacterium tuberculosis infected and non-infected healthy adults. Vaccine 2015, 33, 4130–4140. [Google Scholar] [CrossRef] [PubMed]
- Reither, K.; Katsoulis, L.; Beattie, T.; Gardiner, N.; Lenz, N.; Said, K.; Mfinanga, E.; Pohl, C.; Fielding, K.L.; Jeffery, H.; et al. Safety and immunogenicity of H1/IC31(R), an adjuvanted TB subunit vaccine, in HIV-infected adults with CD4+ lymphocyte counts greater than 350 cells/mm3: a phase II, multi-centre, double-blind, randomized, placebo-controlled trial. PLoS ONE 2014, 9, e114602. [Google Scholar] [CrossRef] [PubMed]
- Van Dissel, J.T.; Arend, S.M.; Prins, C.; Bang, P.; Tingskov, P.N.; Lingnau, K.; Nouta, J.; Klein, M.R.; Rosenkrands, I.; Ottenhoff, T.H.; et al. Ag85B-ESAT-6 adjuvanted with IC31 promotes strong and long-lived Mycobacterium tuberculosis specific T cell responses in naive human volunteers. Vaccine 2010, 28, 3571–3581. [Google Scholar] [CrossRef] [PubMed]
- Weeratna, R.D.; Brazolot Millan, C.L.; McCluskie, M.J.; Davis, H.L. CpG ODN can re-direct the Th bias of established Th2 immune responses in adult and young mice. FEMS Immunol. Med. Microbiol. 2001, 32, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Davis, H.L.; Weeratna, R.; Waldschmidt, T.J.; Tygrett, L.; Schorr, J.; Krieg, A.M.; Weeratna, R. CpG DNA is a potent enhancer of specific immunity in mice immunized with recombinant hepatitis B surface antigen. J. Immunol. 1998, 160, 870–876. [Google Scholar] [PubMed]
- McCluskie, M.J.; Krieg, A.M. Enhancement of infectious disease vaccines through TLR9-dependent recognition of CpG DNA. Curr. Top. Microbiol. Immunol. 2006, 311, 155–178. [Google Scholar] [PubMed]
- Vollmer, J.; Krieg, A.M. Immunotherapeutic applications of CpG oligodeoxynucleotide TLR9 agonists. Adv. Drug Deliv. Rev. 2009, 61, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Klinman, D.M.; Currie, D.; Gursel, I.; Verthelyi, D. Use of CpG oligodeoxynucleotides as immune adjuvants. Immunol. Rev. 2004, 199, 201–216. [Google Scholar] [CrossRef] [PubMed]
- Uhlmann, E.; Ryte, A.; Peyman, A. Studies on the mechanism of stabilization of partially phosphorothioated oligonucleotides against nucleolytic degradation. Antisense Nucleic Acid Drug Dev. 1997, 7, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Matson, S.; Herrera, C.J.; Fisher, E.; Yu, H.; Krieg, A.M. Comparison of cellular binding and uptake of antisense phosphodiester, phosphorothioate, and mixed phosphorothioate and methylphosphonate oligonucleotides. Antisense Res. Dev. 1993, 3, 53–66. [Google Scholar] [PubMed]
- Schellack, C.; Prinz, K.; Egyed, A.; Fritz, J.H.; Wittmann, B.; Ginzler, M.; Swatosch, G.; Zauner, W.; Kast, C.; Akira, S.; et al. IC31, a novel adjuvant signaling via TLR9, induces potent cellular and humoral immune responses. Vaccine 2006, 24, 5461–5472. [Google Scholar] [PubMed]
- Hemmi, H.; Takeuchi, O.; Kawai, T.; Kaisho, T.; Sato, S.; Sanjo, H.; Matsumoto, M.; Hoshino, K.; Wagner, H.; Takeda, K.; et al. A Toll-like receptor recognizes bacterial DNA. Nature 2000, 408, 740–745. [Google Scholar] [PubMed]
- Krieg, A.M. CpG motifs in bacterial DNA and their immune effects. Annu. Rev. Immunol. 2002, 20, 709–760. [Google Scholar] [CrossRef] [PubMed]
- Lalvani, A.; Pathan, A.A.; McShane, H.; Wilkinson, R.J.; Latif, M.; Conlon, C.P.; Pasvol, G.; Hill, A.V. Rapid detection of Mycobacterium tuberculosis infection by enumeration of antigen-specific T cells. Am. J. Respir. Crit. Care Med. 2001, 163, 824–828. [Google Scholar] [CrossRef]
- McCluskie, M.J.; Weeratna, R.D.; Evans, D.M.; Makinen, S.; Drane, D.; Davis, H.L. CpG ODN and ISCOMATRIX adjuvant: A synergistic adjuvant combination inducing strong T-Cell IFN-gamma responses. Biomed. Res. Int. 2013. [Google Scholar] [CrossRef] [PubMed]
- Krieg, A.M. CpG DNA: Trigger of sepsis, mediator of protection, or both? Scand. J. Infect. Dis. 2003, 35, 653–659. [Google Scholar] [PubMed]
- Weiner, G.J. The immunobiology and clinical potential of immunostimulatory CpG oligodeoxynucleotides. J. Leukoc. Biol. 2000, 68, 455–463. [Google Scholar] [PubMed]
- Krieg, A.M.; Yi, A.K.; Matson, S.; Waldschmidt, T.J.; Bishop, G.A.; Teasdale, R.; Koretzky, G.A.; Klinman, D.M. CpG motifs in bacterial DNA trigger direct B-cell activation. Nature 1995, 374, 546–549. [Google Scholar] [CrossRef] [PubMed]
- Haas, T.; Metzger, J.; Schmitz, F.; Heit, A.; Muller, T.; Latz, E.; Wagner, H. The DNA sugar backbone 2′ deoxyribose determines toll-like receptor 9 activation. Immunity 2008, 28, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Chikh, G.; de Jong, S.D.; Sekirov, L.; Raney, S.G.; Kazem, M.; Wilson, K.D.; Cullis, P.R.; Dutz, J.P.; Tam, Y.K. Synthetic methylated CpG ODNs are potent in vivo adjuvants when delivered in liposomal nanoparticles. Int. Immunol. 2009, 21, 757–767. [Google Scholar] [CrossRef] [PubMed]
- Aichinger, M.C.; Ginzler, M.; Weghuber, J.; Zimmermann, L.; Riedl, K.; Schutz, G.; Nagy, E.; von Gabain, A.; Schweyen, R.; Henics, T. Adjuvating the adjuvant: facilitated delivery of an immunomodulatory oligonucleotide to TLR9 by a cationic antimicrobial peptide in dendritic cells. Vaccine 2011, 29, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Nag, S.; Vidi, P.A.; Irudayaraj, J. Single molecule in vivo analysis of toll-like receptor 9 and CpG DNA interaction. PLoS ONE 2011, 6, e17991. [Google Scholar] [CrossRef] [PubMed]
- Lande, R.; Gregorio, J.; Facchinetti, V.; Chatterjee, B.; Wang, Y.H.; Homey, B.; Cao, W.; Wang, Y.H.; Su, B.; Nestle, F.O.; et al. Plasmacytoid dendritic cells sense self-DNA coupled with antimicrobial peptide. Nature 2007, 449, 564–569. [Google Scholar] [PubMed]
- Lahoud, M.H.; Ahmet, F.; Zhang, J.G.; Meuter, S.; Policheni, A.N.; Kitsoulis, S.; Lee, C.N.; O’Keeffe, M.; Sullivan, L.C.; Brooks, A.G.; et al. DEC-205 is a cell surface receptor for CpG oligonucleotides. Proc. Natl. Acad. Sci. USA 2012, 109, 16270–16275. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, K.; Rutz, M.; Schlatter, B.; Metzger, J.; Luppa, P.B.; Schmitz, F.; Haas, T.; Heit, A.; Bauer, S.; Wagner, H. CpG motif-independent activation of TLR9 upon endosomal translocation of “natural” phosphodiester DNA. Eur. J. Immunol. 2006, 36, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Shargh, V.H.; Jaafari, M.R.; Khamesipour, A.; Jaafari, I.; Jalali, S.A.; Abbasi, A.; Badiee, A. Liposomal SLA co-incorporated with PO CpG ODNs or PS CpG ODNs induce the same protection against the murine model of leishmaniasis. Vaccine 2012, 30, 3957–3964. [Google Scholar] [PubMed]
- Roberts, T.L.; Dunn, J.A.; Sweet, M.J.; Hume, D.A.; Stacey, K.J. The immunostimulatory activity of phosphorothioate CpG oligonucleotides is affected by distal sequence changes. Mol. Immunol. 2011, 48, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Sester, D.P.; Naik, S.; Beasley, S.J.; Hume, D.A.; Stacey, K.J. Phosphorothioate backbone modification modulates macrophage activation by CpG DNA. J. Immunol. 2000, 165, 4165–4173. [Google Scholar] [CrossRef]
- Kim, D.; Kwon, S.; Ahn, C.S.; Lee, Y.; Choi, S.Y.; Park, J.; Kwon, H.Y.; Kwon, H.J. Adjuvant effect of liposome-encapsulated natural phosphodiester CpG-DNA. BMB Rep. 2011, 44, 758–763. [Google Scholar] [CrossRef] [PubMed]
- De Jong, S.D.; Basha, G.; Wilson, K.D.; Kazem, M.; Cullis, P.; Jefferies, W.; Tam, Y. The immunostimulatory activity of unmethylated and methylated CpG oligodeoxynucleotide is dependent on their ability to colocalize with TLR9 in late endosomes. J. Immunol. 2010, 184, 6092–6102. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, P.; Peh, C.A. LL-37 promotes rapid sensing of CpG oligonucleotides by B lymphocytes and plasmacytoid dendritic cells. J. Immunol. 2010, 184, 1425–1435. [Google Scholar] [CrossRef] [PubMed]
- Kindrachuk, J.; Jenssen, H.; Elliott, M.; Townsend, R.; Nijnik, A.; Lee, S.F.; Gerdts, V.; Babiuk, L.A.; Halperin, S.A.; Hancock, R.E. A novel vaccine adjuvant comprised of a synthetic innate defence regulator peptide and CpG oligonucleotide links innate and adaptive immunity. Vaccine 2009, 27, 4662–4671. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Li, H.; Jiang, Z.; Xu, C.; Cheng, Q.; Yang, Z.; Cao, G.; Zhang, L. Synthetic innate defence regulator peptide enhances in vivo immunostimulatory effects of CpG-ODN in newborn piglets. Vaccine 2010, 28, 6006–6013. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ODN | Backbone | Sequence |
|---|---|---|
| CpG(S) 1 | Phosphorothioate (PS) | TCCATGACGTTCCTGACGTT |
| CpG(O) | Phosphodiester (PO) | TCCATGACGTTCCTGACGTT |
| GpC(S) | Phosphorothioate (PS) | TCCATGAGCTTCCTGAGCTT |
| GpG(O) | Phosphodiester (PO) | TCCATGAGCTTCCTGAGCTT |
| Compound | Peptides | Oligodeoxynucleotides (ODN) | ||||
|---|---|---|---|---|---|---|
| KLK | KPK | CpG(S) | CpG(O) | GpC(S) | GpC(O) | |
| Single agent | ** vs. OVA | NS vs. OVA | **** vs. OVA | NS vs. OVA | NS vs. OVA | NS vs. OVA |
| KLK combo | **** vs. OVA | **** vs. OVA | **** vs. OVA | ** vs. OVA | ||
| NS vs. CpG(S) | ** vs. KLK | NS vs. KLK | NS vs. KLK | |||
| KPK combo | **** vs. OVA | NS vs. OVA | ** vs. OVA | NS vs. OVA | ||
| NS vs. CpG(S) | NS vs. KPK | ** vs. KPK | NS vs. KPK | |||
| Treatment | CD11c+ APC | CD11b+ APC | ||||
|---|---|---|---|---|---|---|
| 5 h | 24 h | 48 h | 5 h | 24 h | 48 h | |
| Naïve (no ODN) | 121 ± 2 | 121 ± 2 | 121 ± 2 | 99 ± 8 | 99 ± 8 | 99 ± 8 |
| CpG(S) | 409 ± 45 | 584 ± 105 | 565 ± 71 | 384 ± 30 | 405 ± 6 | 324 ± 1 |
| CpG(S) + KLK | 124 ± 6 | 133 ± 4 | 157 ± 8 | 92 ± 9 | 109 ± 10 | 117 ± 1 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chikh, G.; Luu, R.; Patel, S.; Davis, H.L.; Weeratna, R.D. Effects of KLK Peptide on Adjuvanticity of Different ODN Sequences. Vaccines 2016, 4, 14. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines4020014
Chikh G, Luu R, Patel S, Davis HL, Weeratna RD. Effects of KLK Peptide on Adjuvanticity of Different ODN Sequences. Vaccines. 2016; 4(2):14. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines4020014
Chicago/Turabian StyleChikh, Ghania, Rachel Luu, Shobhna Patel, Heather L. Davis, and Risini D. Weeratna. 2016. "Effects of KLK Peptide on Adjuvanticity of Different ODN Sequences" Vaccines 4, no. 2: 14. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines4020014