
Aeromonas hydrophila OmpW PLGA Nanoparticle Oral Vaccine Shows a Dose-Dependent Protective Immunity in Rohu (Labeo rohita)

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Expression and Purification of A. hydrophila Recombinant OmpW Protein
2.2. Encapsulation of rOmpW in PLGA Nanoparticles
2.3. Characterization of PLGA Nanoparticles and In Vitro Release Test
2.4. Vaccine Preparations for Oral Delivery
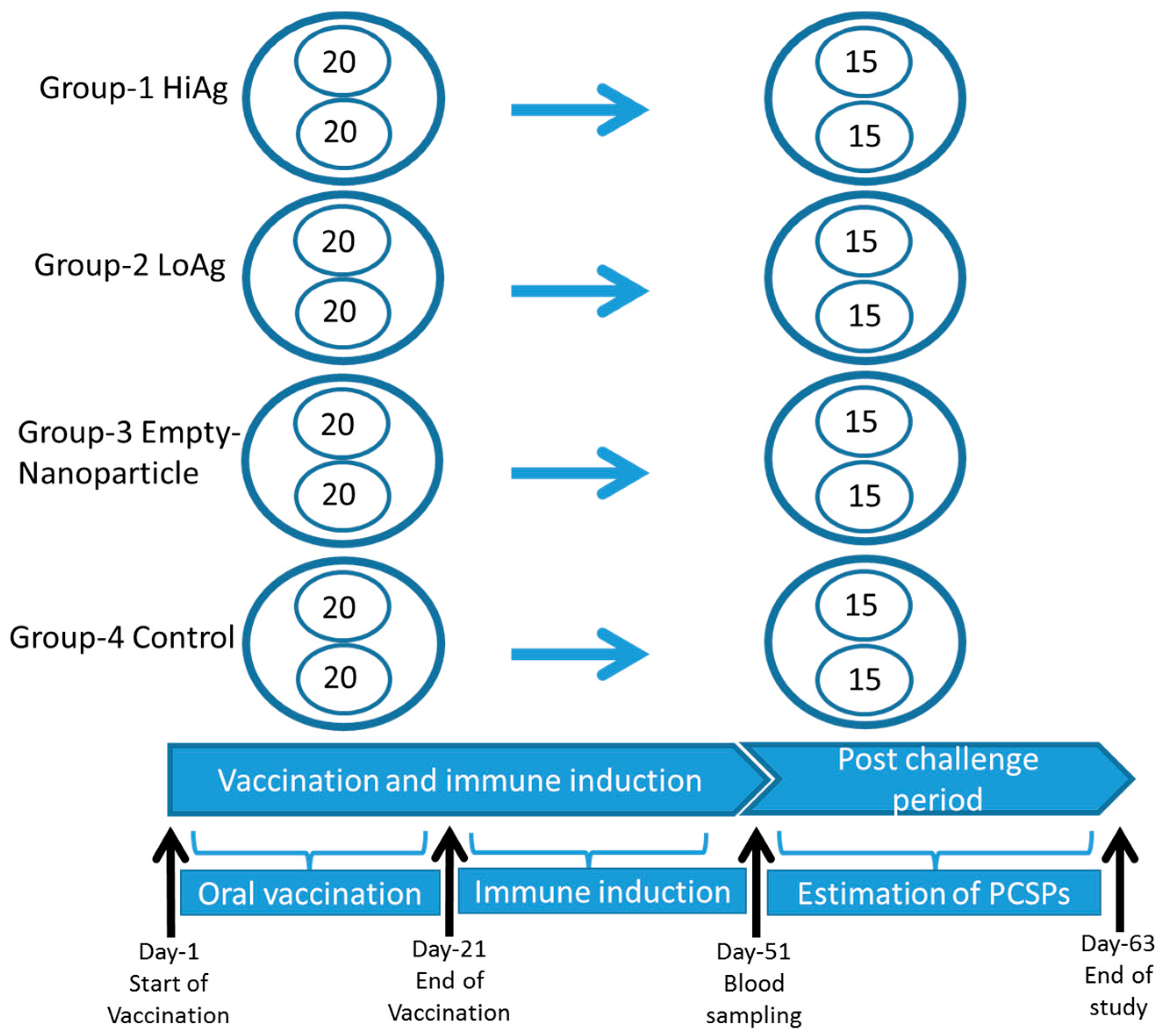
2.5. Vaccination and Challenge
2.6. Serum Inhibitory Assay
2.7. Antibody Response to the rOmpW Protein
3. Results
3.1. Antigen Preparation
3.1.1. Expression, Purification, Concentration, and Encapsulation Efficiency of OmpW
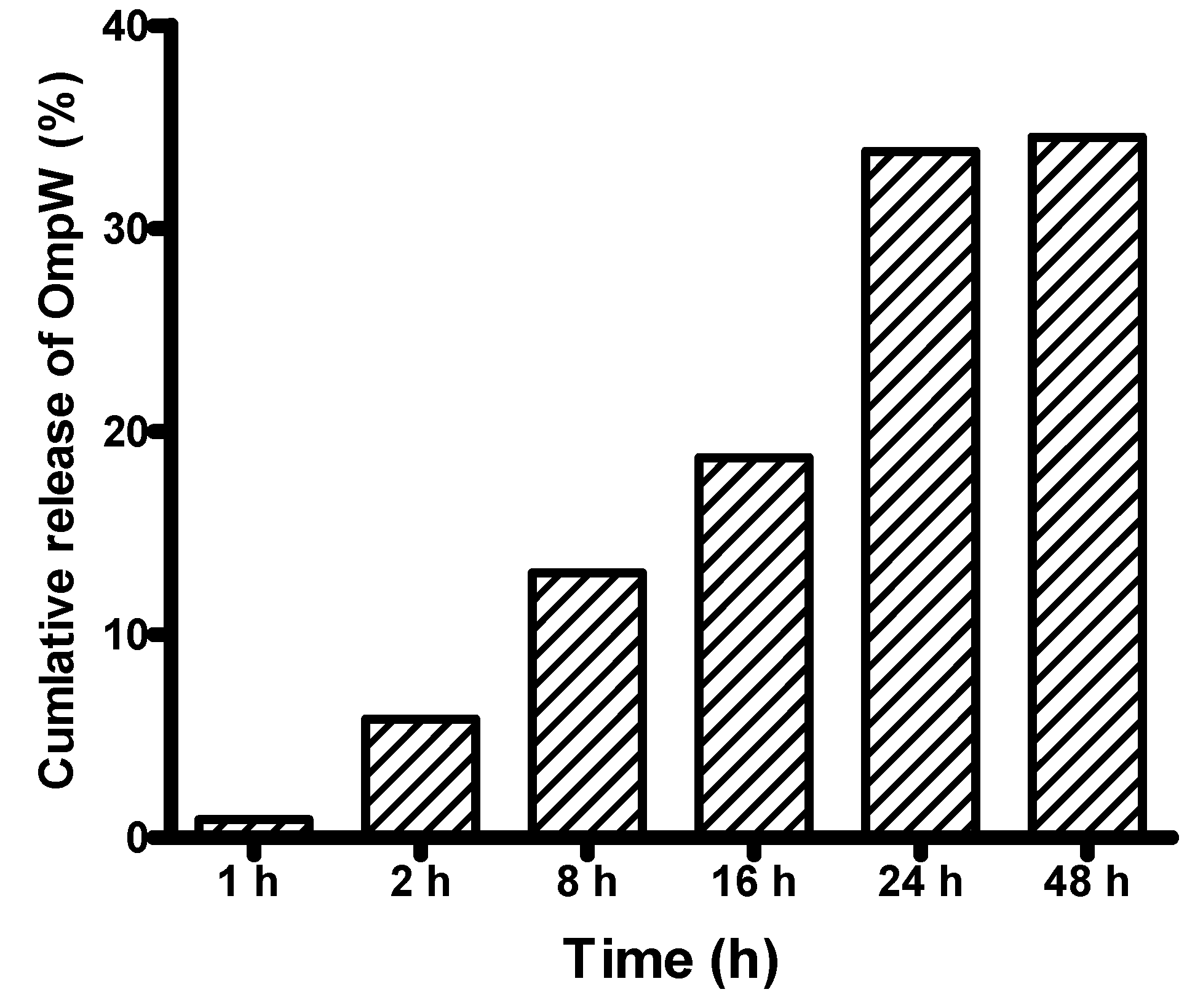
3.1.2. In Vitro Release of the rOmpW Protein from PLGA Nanoparticles
3.2. Vaccination and Challenge of Rohu
3.2.1. Kaplan Meyer’s Survival Analysis
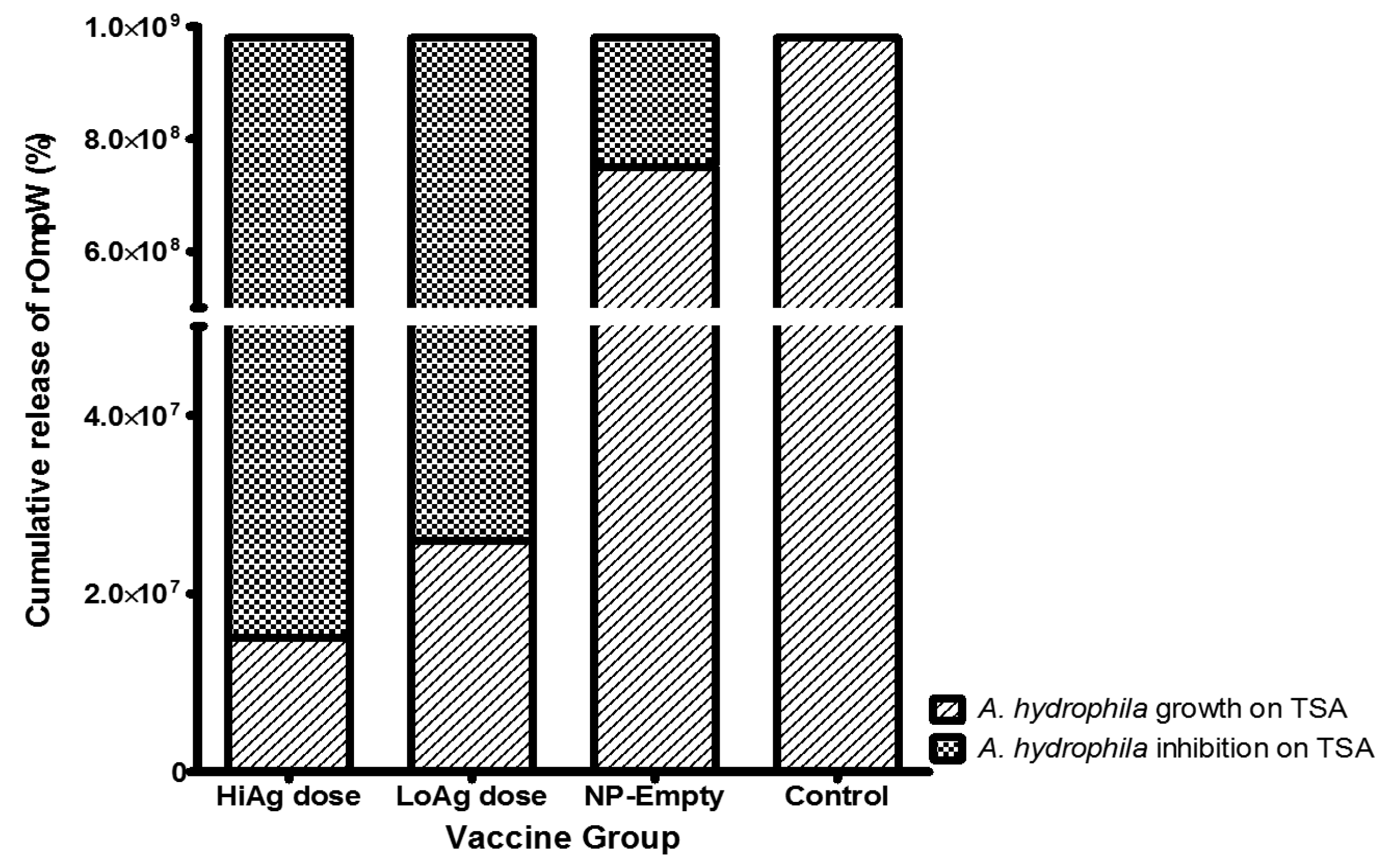
3.2.2. Serum Inhibition of A. hydrophila Growth on Trypticase Soy Agar (TSA) Agar
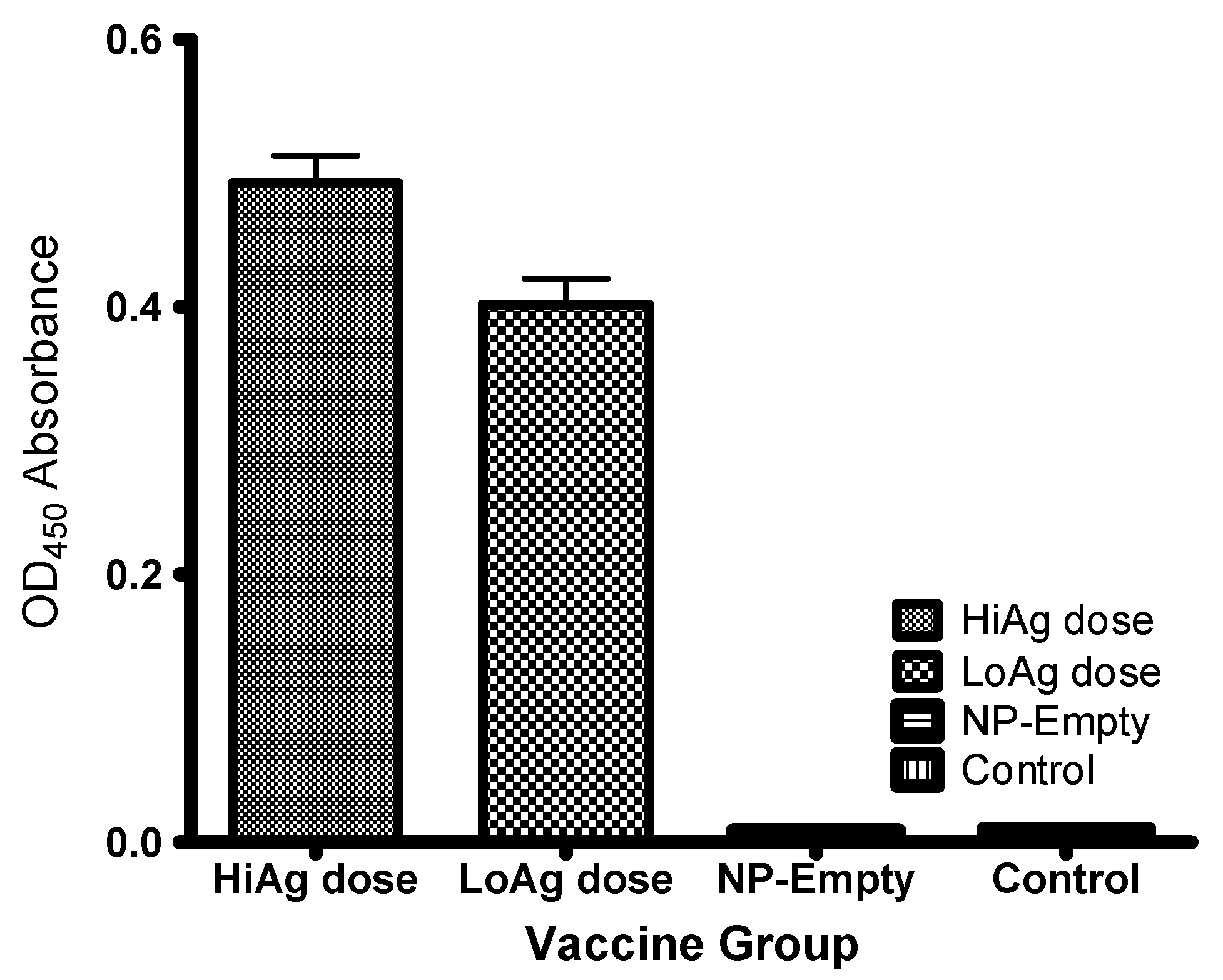
3.2.3. Antibody Responses
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Liu, L.; Gong, Y.-X.; Zhu, B.; Liu, G.-L.; Wang, G.-X.; Ling, F. Effect of a new recombinant Aeromonas hydrophila vaccine on the grass carp intestinal microbiota and correlations with immunological responses. Fish Shellfish Immunol. 2015, 45, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Wu, Y.; Pang, M.; Liu, J.; Lu, C.; Liu, Y. Protective efficacy of recombinant hemolysin co-regulated protein (hcp) of Aeromonas hydrophila in common carp (cyprinus carpio). Fish Shellfish Immunol. 2015, 46, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Nikoskelainen, S.; Verho, S.; Järvinen, S.; Madetoja, J.; Wiklund, T.; Lilius, E.-M. Multiple whole bacterial antigens in polyvalent vaccine may result in inhibition of specific responses in rainbow trout (oncorhynchus mykiss). Fish Shellfish Immunol. 2007, 22, 206–217. [Google Scholar] [CrossRef] [PubMed]
- Gudding, R.; Lillehaug, A.; Evensen, O. Recent developments in fish vaccinology. Vet. Immunol. Immunopathol. 1999, 72, 203–212. [Google Scholar] [CrossRef]
- Chandran, M.R.; Aruna, B.V.; Logambal, S.M.; Michael, R.D. Immunisation of indian major carps against Aeromonas hydrophila by intraperitoneal injection. Fish Shellfish Immunol. 2002, 13, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dash, S.; Das, S.K.; Samal, J.; Ojha, P.K.; Patra, J.K.; Thatoi, H. Dose dependence specific and non-specific immune responses of indian major carp (L. rohita ham) to intraperitoneal injection of formalin killed Aeromonas hydrophila whole cell vaccine. Vet. Res. Commun. 2011, 35, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Munang’andu, H.M.; Mutoloki, S.; Evensen, O. An overview of challenges limiting the design of protective mucosal vaccines for finfish. Front. Immunol. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tafalla, C.; Bøgwald, J.; Dalmo, R.A. Adjuvants and immunostimulants in fish vaccines: Current knowledge and future perspectives. Fish Shellfish Immunol. 2013, 35, 1740–1750. [Google Scholar] [CrossRef] [PubMed]
- Munang’andu, H.M.; Evensen, O. A review of intra- and extra-cellular antigen delivery systems for virus vaccines of finfish. J. Immunol. Res. 2015. [Google Scholar] [CrossRef] [PubMed]
- Tafalla, C.; Bøgwald, J.; Dalmo, R.A.; Munang’andu, H.M.; Evensen, Ø. Adjuvants in Fish Vaccines. In Fish Vaccination; John Wiley & Sons, Inc.: New York, NY, USA, 2014; pp. 68–84. [Google Scholar]
- Munang’andu, H.M.; Mutoloki, S.; Evensen, Ø. Non-replicating vaccines. In Fish Vaccination; John Wiley & Sons, Inc.: New York, NY, USA, 2014; pp. 22–32. [Google Scholar]
- Munang’andu, H.M.; Fredriksen, B.N.; Mutoloki, S.; Brudeseth, B.; Kuo, T.Y.; Marjara, I.S.; Dalmo, R.A.; Evensen, O. Comparison of vaccine efficacy for different antigen delivery systems for infectious pancreatic necrosis virus vaccines in atlantic salmon (Salmo salar L.) in a cohabitation challenge model. Vaccine 2012, 30, 4007–4016. [Google Scholar] [CrossRef] [PubMed]
- Fredriksen, B.N.; Grip, J. PLGA/PLA micro- and nanoparticle formulations serve as antigen depots and induce elevated humoral responses after immunization of atlantic salmon (Salmo salar L.). Vaccine 2012, 30, 656–667. [Google Scholar] [CrossRef] [PubMed]
- Fredriksen, B.N.; Sævareid, K.; McAuley, L.; Lane, M.E.; Bøgwald, J.; Dalmo, R.A. Early immune responses in atlantic salmon (Salmo salar L.) after immunization with PLGA nanoparticles loaded with a model antigen and β-glucan. Vaccine 2011, 29, 8338–8349. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Torrealba, D.; Ruyra, A.; Roher, N. Nanodelivery systems as new tools for immunostimulant or vaccine administration: Targeting the fish immune system. Biology 2015, 4, 664–696. [Google Scholar] [CrossRef] [PubMed]
- Behera, T.; Nanda, P.K.; Mohanty, C.; Mohapatra, D.; Swain, P.; Das, B.K.; Routray, P.; Mishra, B.K.; Sahoo, S.K. Parenteral immunization of fish, Labeo rohita with poly d,l-lactide-co-glycolic acid (PLGA) encapsulated antigen microparticles promotes innate and adaptive immune responses. Fish Shellfish Immunol. 2010, 28, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Hamod, M.A.; Nithin, M.S.; Shukur, Y.N.; Karunasagar, I.; Karunasagar, I. Outer membrane protein k as a subunit vaccine against V. anguillarum. Aquaculture 2012, 354–355, 107–110. [Google Scholar] [CrossRef]
- Maiti, B.; Raghunath, P.; Karunasagar, I.; Karunasagar, I. Cloning and expression of an outer membrane protein OmpW of Aeromonas hydrophila and study of its distribution in Aeromonas spp. J. Appl. Microbiol. 2009, 107, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Anderson, J.M.; Shive, M.S. Biodegradation and biocompatibility of PLA and PLGA microspheres. Adv. Drug Deliv. Rev. 1997, 28, 5–24. [Google Scholar] [CrossRef]
- Coombes, A.G.A.; Yeh, M.-K.; Lavelle, E.C.; Davis, S.S. The control of protein release from poly(d,l-lactide co-glycolide) microparticles by variation of the external aqueous phase surfactant in the water-in oil-in water method. J. Control. Release 1998, 52, 311–320. [Google Scholar] [CrossRef]
- Intra, J.; Salem, A.K. Fabrication, characterization and in vitro evaluation of poly (d,l-lactide-co-glycolide) microparticles loaded with polyamidoamine-plasmid DNA dendriplexes for applications in non-viral gene delivery. J. Pharm. Sci. 2010, 99, 368–384. [Google Scholar] [CrossRef] [PubMed]
- Kofler, N.; Ruedl, C.; Klima, J.; Recheis, H.; Böck, G.; Wick, G.; Wolf, H. Preparation and characterization of poly-(d,l-lactide-co-glycolide) and poly-(l-lactic acid) microspheres with entrapped pneumotropic bacterial antigens. J. Immunol. Methods 1996, 192, 25–35. [Google Scholar] [CrossRef]
- Nayak, B.; Panda, A.K.; Ray, P.; Ray, A.R. Formulation, characterization and evaluation of rotavirus encapsulated PLA and PLGA particles for oral vaccination. J. Microencapsul. 2009, 26, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Rauta, P.R.; Nayak, B. Parenteral immunization of PLA/PLGA nanoparticle encapsulating outer membrane protein (Omp) from Aeromonas hydrophila: Evaluation of immunostimulatory action in Labeo rohita (rohu). Fish Shellfish Immunol. 2015, 44, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Goel, D.; Bhatnagar, R. Recombinant l7/l12 protein entrapping PLGA (poly lactide-co-glycolide) micro particles protect balb/c mice against the virulent b. Abortus 544 infection. Vaccine 2015, 33, 2786–2792. [Google Scholar] [CrossRef] [PubMed]
- Maiti, B.; Shetty, M.; Shekar, M.; Karunasagar, I.; Karunasagar, I. Evaluation of two outer membrane proteins, aha1 and OmpW of Aeromonas hydrophila as vaccine candidate for common carp. Vet. Immunol. Immunopathol. 2012, 149, 298–301. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.H.; Lu, Y.S.; Jian, J.C.; Wang, B.; Huang, Y.C.; Tang, J.F.; Ding, Y.; Wu, Z.H. Protection against Vibrio alginolyticus in crimson snapper Lutjanus erythropterus immunized with a DNA vaccine containing the OmpW gene. Dis. Aquat. Organ. 2013, 106, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Yu, L.; You, Z.; Wei, Y.; Liu, Y. Cloning, expression and immunogenicty analysis of five outer membrane proteins of Vibrio parahaemolyticus zj2003. Fish Shellfish Immunol. 2007, 23, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Qian, R.; Chu, W.; Mao, Z.; Zhang, C.; Wei, Y.; Yu, L. Expression, characterization and immunogenicity of a major outer membrane protein from Vibrio alginolyticus. Acta Biochim. Biophys. Sin. 2007, 39, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Akagi, T.; Wang, X.; Uto, T.; Baba, M.; Akashi, M. Protein direct delivery to dendritic cells using nanoparticles based on amphiphilic poly(amino acid) derivatives. Biomaterials 2007, 28, 3427–3436. [Google Scholar] [CrossRef] [PubMed]
- Uto, T.; Akagi, T.; Hamasaki, T.; Akashi, M.; Baba, M. Modulation of innate and adaptive immunity by biodegradable nanoparticles. Immunol. Lett. 2009, 125, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Uto, T.; Wang, X.; Sato, K.; Haraguchi, M.; Akagi, T.; Akashi, M.; Baba, M. Targeting of antigen to dendritic cells with poly(gamma-glutamic acid) nanoparticles induces antigen-specific humoral and cellular immunity. J. Immunol. 2007, 178, 2979–2986. [Google Scholar] [CrossRef] [PubMed]
- Danhier, F.; Ansorena, E.; Silva, J.M.; Coco, R.; Le Breton, A.; Preat, V. PLGA-based nanoparticles: An overview of biomedical applications. J. Control. Release 2012, 161, 505–522. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Yadav, S.K.; Yadav, S.C. Biodegradable polymeric nanoparticles based drug delivery systems. Colloids Surf. B Biointerfaces 2010, 75, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Kurupati, P.; Teh, B.K.; Kumarasinghe, G.; Poh, C.L. Identification of vaccine candidate antigens of an esbl producing klebsiella pneumoniae clinical strain by immunoproteome analysis. Proteomics 2006, 6, 836–844. [Google Scholar] [CrossRef] [PubMed]
- Nandi, B.; Nandy, R.K.; Mukhopadhyay, S.; Nair, G.B.; Shimada, T.; Ghose, A.C. Rapid method for species-specific identification of vibrio cholerae using primers targeted to the gene of outer membrane protein OmpW. J. Clin. Microbiol. 2000, 38, 4145–4151. [Google Scholar] [PubMed]
- Tahara, K.; Yamamoto, H.; Hirashima, N.; Kawashima, Y. Chitosan-modified poly(d,l-lactide-co-glycolide) nanospheres for improving siRNA delivery and gene-silencing effects. Eur. J. Pharm. Biopharm. 2010, 74, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Munang’andu, H.M.; Fredriksen, B.N.; Mutoloki, S.; Dalmo, R.A.; Evensen, O. Antigen dose and humoral immune response correspond with protection for inactivated infectious pancreatic necrosis virus vaccines in Atlantic salmon (Salmo salar L.). Vet. Res. 2013. [Google Scholar] [CrossRef]
- Munang’andu, H.M.; Mutoloki, S.; Evensen, O. Acquired immunity and vaccination against infectious pancreatic necrosis virus of salmon. Dev. Comp. Immunol. 2014, 43, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Romstad, A.B.; Reitan, L.J.; Midtlyng, P.; Gravningen, K.; Evensen, O. Antibody responses correlate with antigen dose and in vivo protection for oil-adjuvanted, experimental furunculosis (Aeromonas salmonicida subsp. Salmonicida) vaccines in atlantic salmon (Salmo salar L.) and can be used for batch potency testing of vaccines. Vaccine 2013, 31, 791–796. [Google Scholar] [PubMed]
- Munang’andu, H.M.; Mutoloki, S.; Evensen, O. A review of the immunological mechanisms following mucosal vaccination of finfish. Front. Immunol. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutoloki, S.; Alexandersen, S.; Evensen, Ø. Sequential study of antigen persistence and concomitant inflammatory reactions relative to side-effects and growth of atlantic salmon (Salmo salar L.) following intraperitoneal injection with oil-adjuvanted vaccines. Fish Shellfish Immunol. 2004, 16, 633–644. [Google Scholar] [CrossRef] [PubMed]





© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dubey, S.; Avadhani, K.; Mutalik, S.; Sivadasan, S.M.; Maiti, B.; Paul, J.; Girisha, S.K.; Venugopal, M.N.; Mutoloki, S.; Evensen, Ø.; et al. Aeromonas hydrophila OmpW PLGA Nanoparticle Oral Vaccine Shows a Dose-Dependent Protective Immunity in Rohu (Labeo rohita). Vaccines 2016, 4, 21. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines4020021
Dubey S, Avadhani K, Mutalik S, Sivadasan SM, Maiti B, Paul J, Girisha SK, Venugopal MN, Mutoloki S, Evensen Ø, et al. Aeromonas hydrophila OmpW PLGA Nanoparticle Oral Vaccine Shows a Dose-Dependent Protective Immunity in Rohu (Labeo rohita). Vaccines. 2016; 4(2):21. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines4020021
Chicago/Turabian StyleDubey, Saurabh, Kiran Avadhani, Srinivas Mutalik, Sangeetha Madambithara Sivadasan, Biswajit Maiti, Joydeb Paul, Shivani Kallappa Girisha, Moleyur Nagarajappa Venugopal, Stephen Mutoloki, Øystein Evensen, and et al. 2016. "Aeromonas hydrophila OmpW PLGA Nanoparticle Oral Vaccine Shows a Dose-Dependent Protective Immunity in Rohu (Labeo rohita)" Vaccines 4, no. 2: 21. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines4020021