
The CD8+ T Cell-Mediated Immunity Induced by HPV-E6 Uploaded in Engineered Exosomes Is Improved by ISCOMATRIXTM Adjuvant

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures and Adjuvant
2.2. Production, Purification, and Quantification of Exosomes
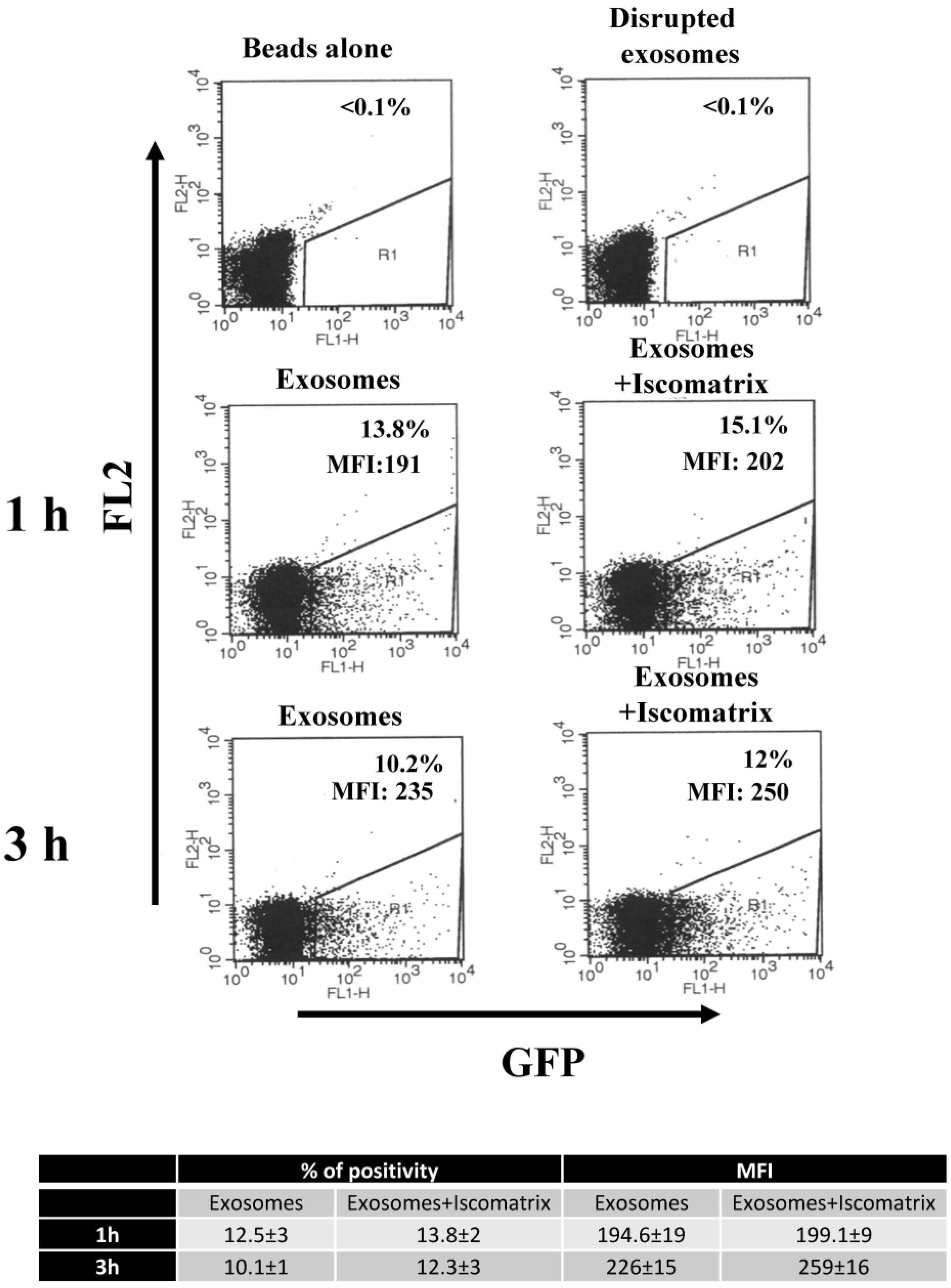
2.3. Fluorescence-Activated Cell Sorting(FACS) Analysis of Bead-Exosome Complexes
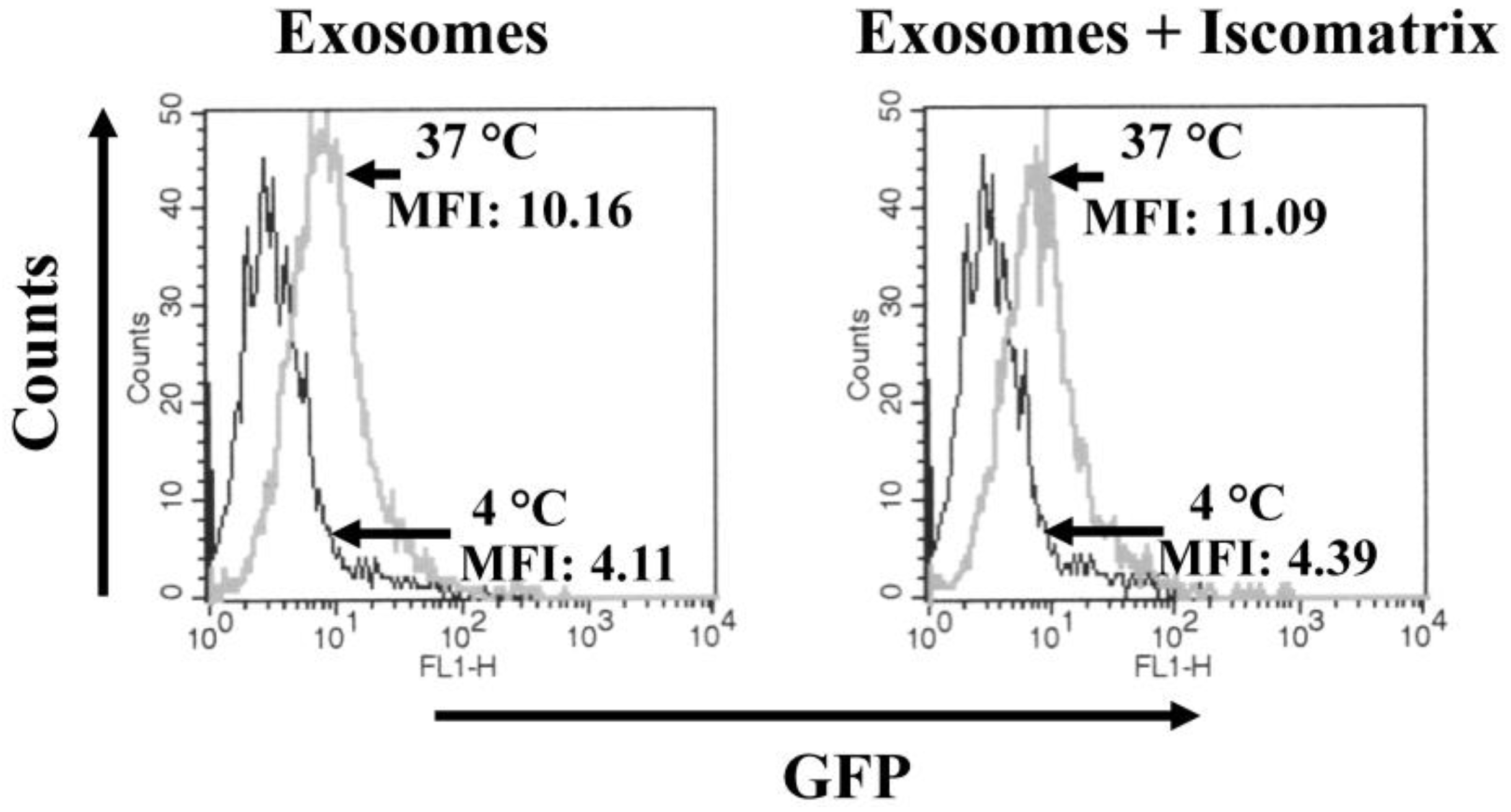
2.4. Detection of Exosome Cell Internalization
2.5. Western Blot Analysis
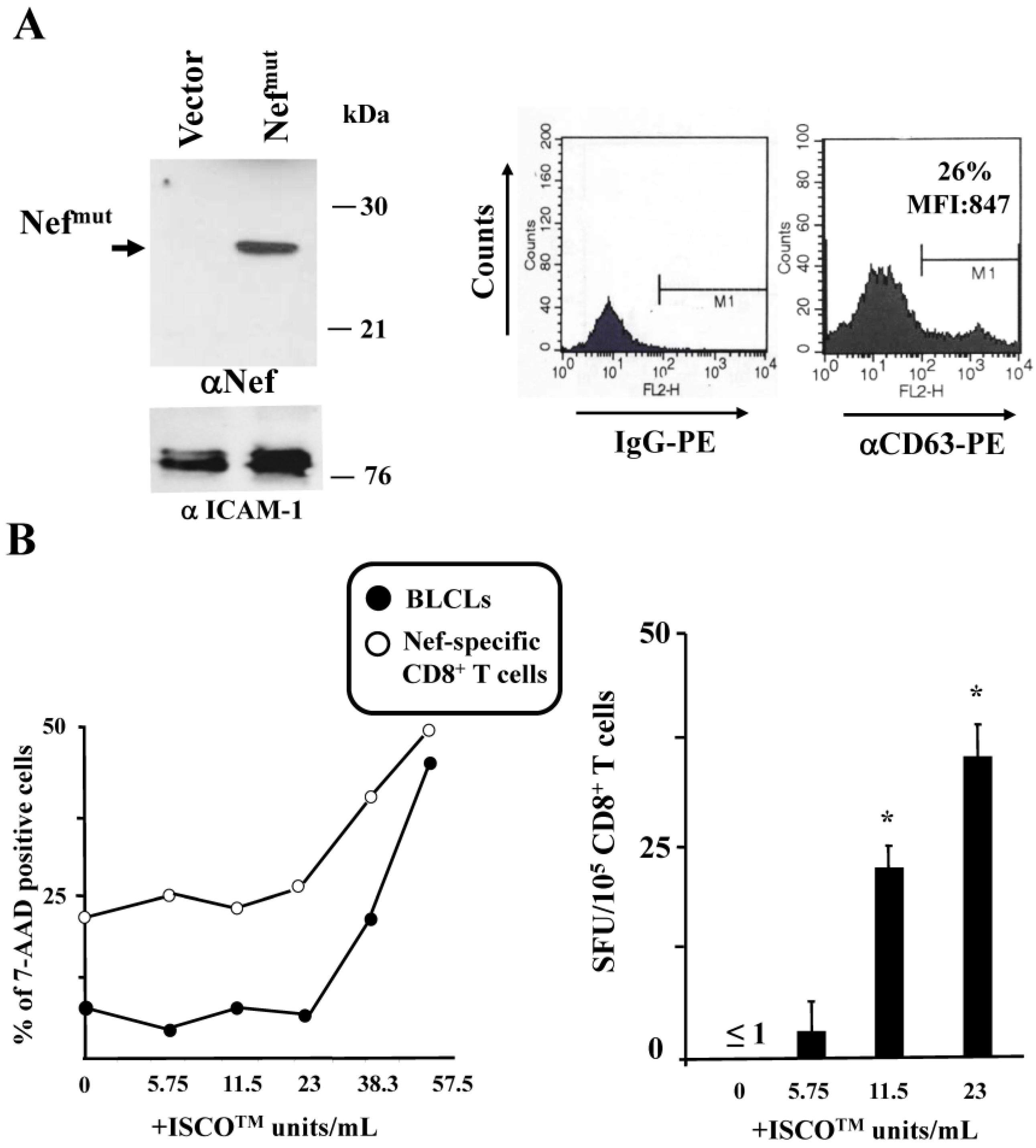
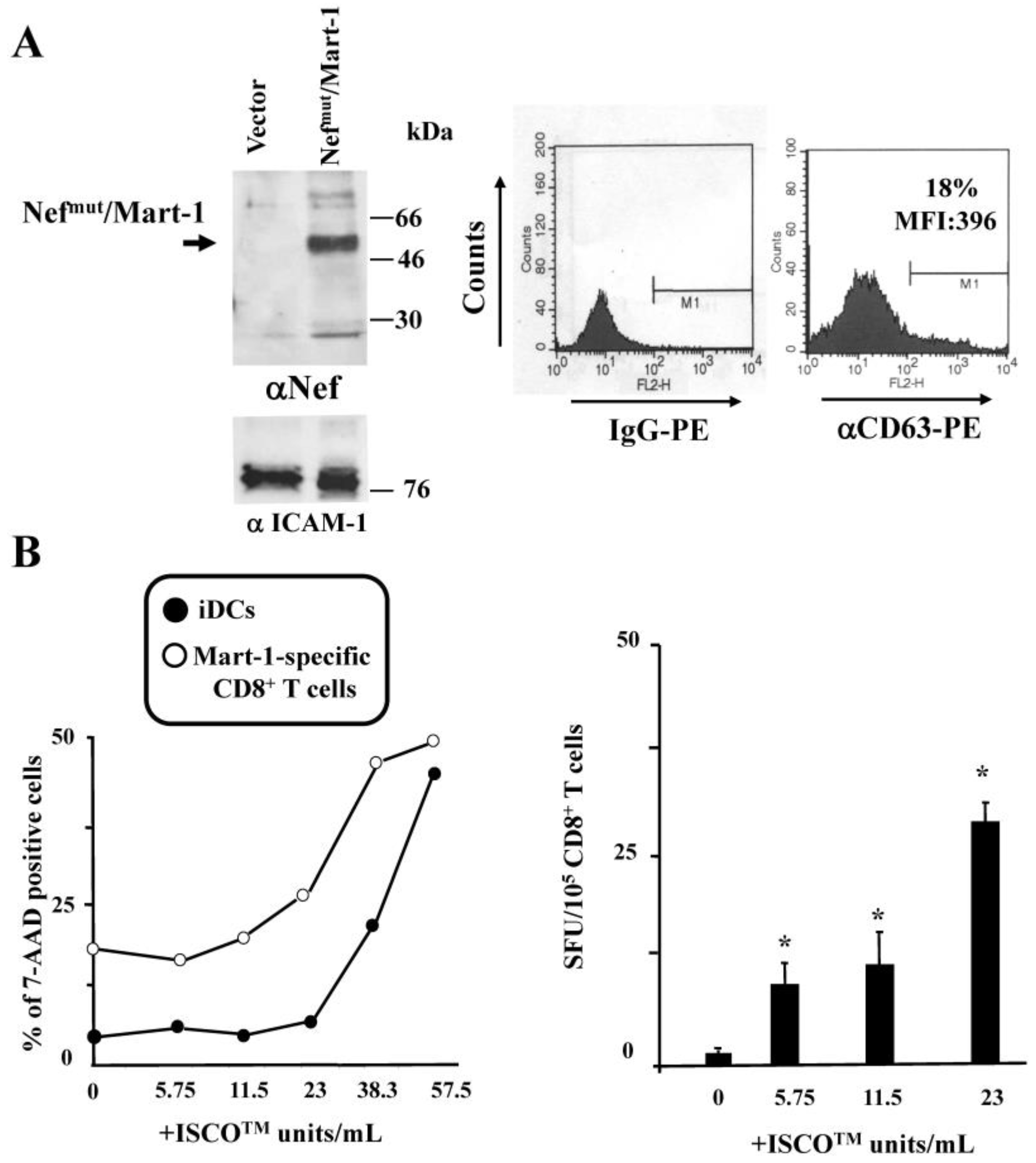
2.6. Cross-Presentation Assay
2.7. Immunogenicity Assay
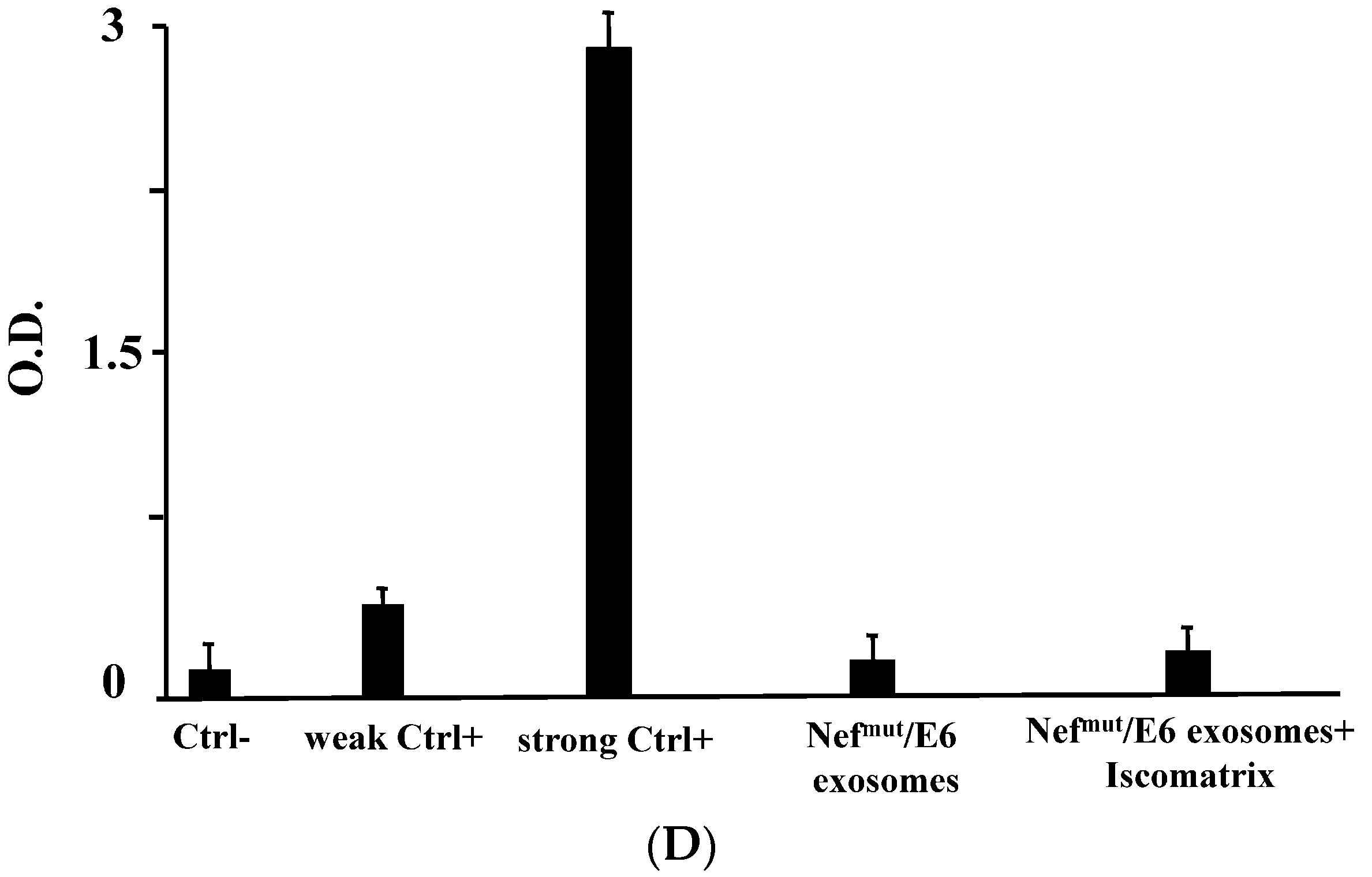
2.8. Detection of Anti-HPV-E6 Abs
2.9. Cytotoxic T Lymphocyte (CTL) Assay
2.10. Statistical Analysis
3. Results
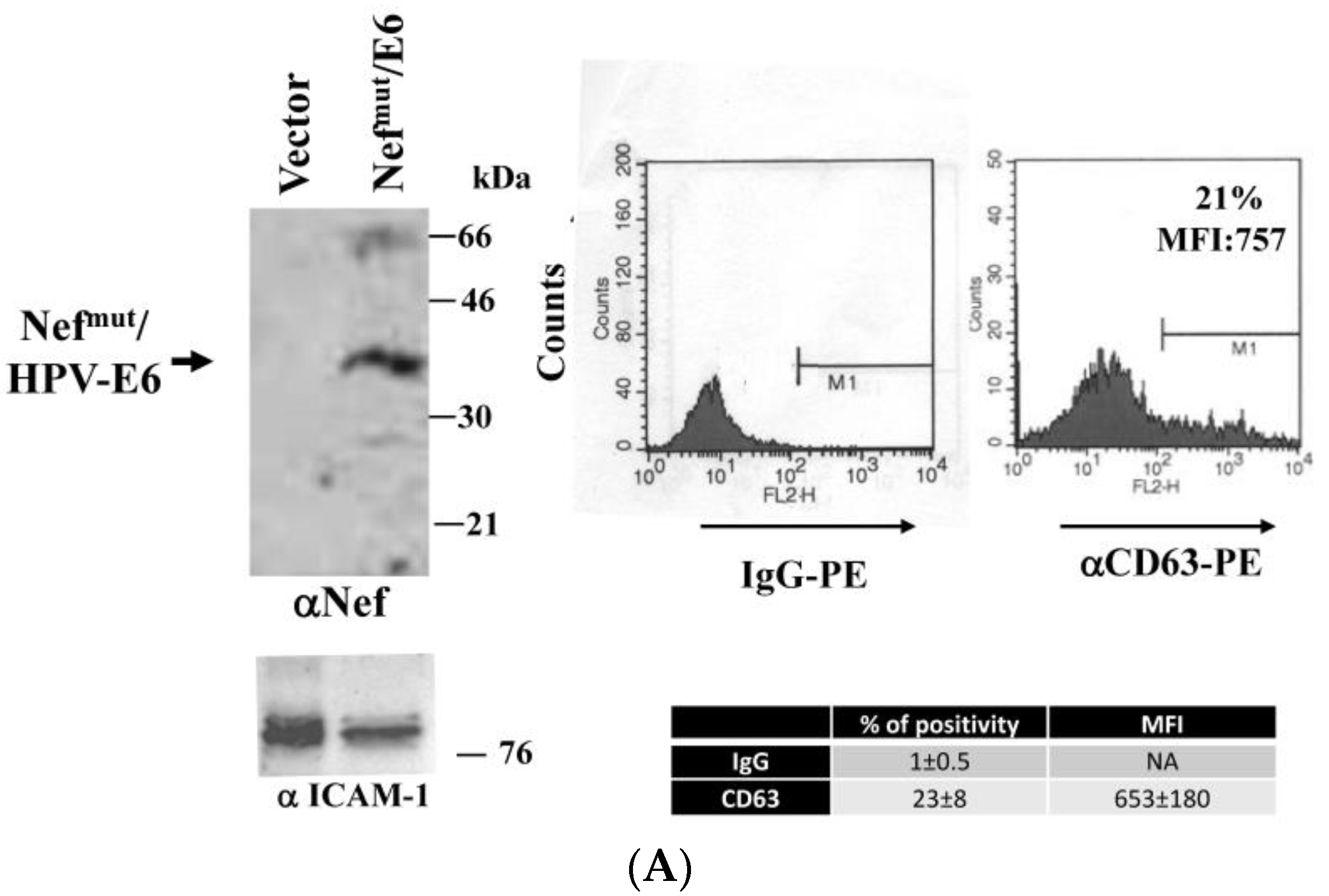
3.1. ISCOMATRIXTM Adjuvant Does Not Affect Both Structure and Cell Entry Efficiency of Exosomes
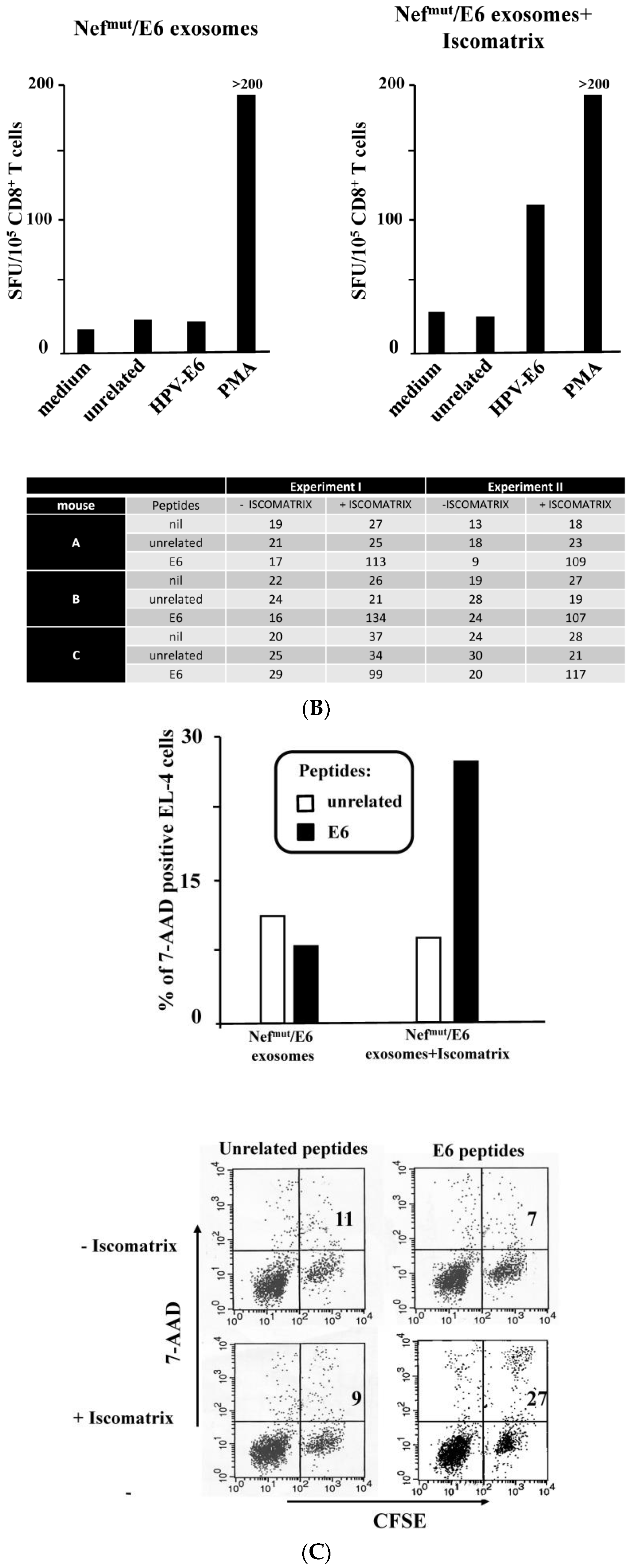
3.2. ISCOMATRIXTM Adjuvant Increases Cross-Presentation of Antigens Uploaded in Engineered Exosomes
3.3. ISCOMATRIXTM Adjuvant and Exosome Co-Administration in Mice Increases the Pool of CD8+ T Lymphocytes Specific for the Antigen Uploaded in Engineered Exosomes
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Milane, L.; Singh, A.; Mattheolabakis, G.; Suresh, M.; Amiji, M.M. Exosome mediated communication within the tumor microenvironment. J. Control Release 2015, 219, 278–294. [Google Scholar] [CrossRef] [PubMed]
- De Toro, J.; Herschlik, L.; Waldner, C.; Mongini, C. Emerging roles of exosomes in normal and pathological conditions: New insights for diagnosis and therapeutic applications. Front. Immunol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Morse, M.A.; Garst, J.; Osada, T.; Khan, S.; Hobeika, A.; Clay, T.M.; Valente, N.; Shreeniwas, R.; Sutton, M.A.; Delcayre, A.; et al. A phase I study of dexosome immunotherapy in patients with advanced non-small cell lung cancer. J. Transl. Med. 2005. [Google Scholar] [CrossRef] [PubMed]
- Escudier, B.; Dorval, T.; Chaput, N.; Andre, F.; Caby, M.P.; Novault, S.; Flament, C.; Leboulaire, C.; Borg, C.; Amigorena, S.; et al. Vaccination of metastatic melanoma patients with autologous dendritic cell (DC) derived-exosomes: Results of the first phase I clinical trial. J. Transl. Med. 2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, S.; Wei, D.; Wu, Z.; Zhou, X.; Wei, X.; Huang, H.; Li, G. Phase I clinical trial of autologous ascites-derived exosomes combined with GM-CSF for colorectal cancer. Mol. Ther. 2008, 16, 782–790. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.; De La Pena, H.; Seifalian, A.M. The application of exosomes as a nanoscale cancer vaccine. Int. J. Nanomed. 2010, 5, 889–900. [Google Scholar]
- Chaput, N.; Thery, C. Exosomes: Immune properties and potential clinical implementations. Semin. Immunopathol. 2011, 33, 419–440. [Google Scholar] [CrossRef] [PubMed]
- Hartman, Z.C.; Wei, J.P.; Glass, O.K.; Guo, H.T.; Lei, G.J.; Yang, X.Y.; Osada, T.; Hobeika, A.; Delcayre, A.; Le Pecq, J.B.; et al. Increasing vaccine potency through exosome antigen targeting. Vaccine 2011, 29, 9361–9367. [Google Scholar] [CrossRef] [PubMed]
- Xiu, F.M.; Cai, Z.J.; Yang, Y.S.; Wang, X.J.; Wang, J.L.; Cao, X.T. Surface anchorage of superantigen SEA promotes induction of specific antitumor immune response by tumor-derived exosomes. J. Mol. Med. 2007, 85, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Usami, Y.; Popov, S.; Popova, E.; Inoue, M.; Weissenhorn, W.; Gottlinger, H.G. The ESCRT pathway and HIV-1 budding. Biochem. Soc. Trans. 2009, 37, 181–184. [Google Scholar] [CrossRef] [PubMed]
- Booth, A.M.; Fang, Y.; Fallon, J.K.; Yang, J.M.; Hildreth, J.E.K.; Gould, S.J. Exosomes and HIV Gag bud from endosome-like domains of the T cell plasma membrane. J. Cell Biol. 2006, 172, 923–935. [Google Scholar] [CrossRef] [PubMed]
- Lenassi, M.; Cagney, G.; Liao, M.F.; Vaupotic, T.; Bartholomeeusen, K.; Cheng, Y.F.; Krogan, N.J.; Plemenitas, A.; Peterlin, B.M. HIV Nef is secreted in exosomes and triggers apoptosis in bystander CD4(+) T cells. Traffic 2010, 11, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Muratori, C.; Cavallin, L.E.; Kratzel, K.; Tinari, A.; De Milito, A.; Fais, S.; D’Aloja, P.; Federico, M.; Vullo, V.; Fomina, A.; et al. Massive secretion by T cells is caused by HIV Nef in infected cells and by Nef transfer to bystander cells. Cell Host Microbe 2009, 6, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Basmaciogullari, S.; Pizzato, M. The activity of Nef on HIV-1 infectivity. Front. Microbiol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Peretti, S.; Schiavoni, I.; Pugliese, K.; Federico, M. Cell death induced by the herpes simplex virus-1 thymidine kinase delivered by human immunodeficiency virus-1-based virus-like particles. Mol. Ther. 2005, 12, 1185–11396. [Google Scholar] [CrossRef] [PubMed]
- Lattanzi, L.; Federico, M. A strategy of antigen incorporation into exosomes: Comparing cross-presentation levels of antigens delivered by engineered exosomes and by lentiviral virus-like particles. Vaccine 2012, 30, 7229–7237. [Google Scholar] [CrossRef] [PubMed]
- Drane, D.; Gittleson, C.; Boyle, J.; Moraskovsky, E. ISCOMATRIX (TM) adjuvant for prophylactic and therapeutic vaccines. Expert Rev. Vaccines 2007, 6, 761–6772. [Google Scholar] [CrossRef] [PubMed]
- Maraskovsky, E.; Schnurr, M.; Wilson, N.S.; Robson, N.C.; Boyle, J.; Drane, D. Development of prophylactic and therapeutic vaccines using the ISCOMATRIX adjuvant. Immunol. Cell Biol. 2009, 87, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Di Bonito, P.; Ridolfi, B.; Columba-Cabezas, S.; Giovannelli, A.; Chiozzini, C.; Manfredi, F.; Anticoli, S.; Arenaccio, C.; Federico, M. HPV-E7 Delivered by engineered exosomes elicits a protective CD8+ T cell-mediated immune response. Viruses 2015, 7, 1079–1099. [Google Scholar] [CrossRef] [PubMed]
- Di Bonito, P.; Grasso, F.; Mochi, S.; Petrone, L.; Fanales-Belasio, E.; Mei, A.; Cesolini, A.; Laconi, G.; Conrad, H.; Bernhard, H.; et al. Anti-tumor CD8(+) T cell immunity elicited by HIV-1-based virus-like particles incorporating HPV-16 E7 protein. Virology 2009, 395, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Rivoltini, L.; Kawakami, Y.; Sakaguchi, K.; Southwood, S.; Sette, A.; Robbins, P.F.; Marincola, F.M.; Salgaller, M.L.; Yannelli, J.R.; Appella, E.; et al. Induction of tumor-reactive CTL from peripheral blood and tumor-infiltrating lymphocytes of melanoma patients by in vitro stimulation with an immunodominant peptide of the human melanoma antigen MART-1. J. Immunol. 1995, 154, 2257–2265. [Google Scholar] [PubMed]
- Arenaccio, C.; Anticoli, S.; Manfredi, F.; Chiozzini, C.; Olivetta, E.; Federico, M. Latent HIV-1 is activated by exosomes from cells infected with either replication-competent or defective HIV-1. Retrovirology 2015. [Google Scholar] [CrossRef] [PubMed]
- Thery, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr. Protoc. Cell Biol. 2006. [Google Scholar] [CrossRef]
- Rieu, S.; Geminard, C.; Rabesandratana, H.; Sainte-Marie, J.; Vidal, M. Exosomes released during reticulocyte maturation bind to fibronectin via integrin alpha 4 beta 1. Eur. J. Biochem. 2000, 267, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Bauer, S.; Heeg, K.; Wagner, H.; Lipford, G.B. Identification of H-2Kb binding and immunogenic peptides from human papilloma virus tumor antigens E6 and E7. Scand. J. Immunol. 1995, 42, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Di Bonito, P.; Grasso, F.; Mochi, S.; Accardi, L.; Dona, M.G.; Branca, M.; Costa, S.; Mariani, L.; Agarossi, A.; Ciotti, M.; et al. Serum antibody response to Human papillomavirus (HPV) infections detected by a novel ELISA technique based on denatured recombinant HPV16 L1, L2, E4, E6 and E7 proteins. Infect. Agent Cancer 2006. [Google Scholar] [CrossRef] [PubMed]
- Busam, K.J.; Jungbluth, A.A. Melan-A, a new melanocytic differentiation marker. Adv. Anat. Pathol. 1999, 6, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Wilson, N.S.; Yang, B.; Baz Morelli, A.; Koernig, S.; Yang, A.; Loeser, S.; Airey, D.; Provan, L.; Hass, P.; Braley, H.; et al. ISCOMATRIX vaccines mediate CD8(+) T-cell cross-priming by a MyD88-dependent signaling pathway. Immunol. Cell Biol. 2012, 90, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Joffre, O.P.; Segura, E.; Savina, A.; Amigorena, S. Cross-presentation by dendritic cells. Nat. Rev. Immunol. 2012, 12, 557–569. [Google Scholar] [CrossRef] [PubMed]
- Davis, I.D.; Chen, W.; Jackson, H.; Parente, P.; Shackleton, M.; Hopkins, W.; Chen, Q.; Dinopoulos, N.; Luke, T.; Murphy, R.; et al. Recombinant NY-ESO-1 protein with ISCOMATRIX adjuvant induces broad integrated Ab and CD4+ and CD8+ T cell responses in humans. Proc. Natl. Acad. Sci. USA 2004, 101, 10697–10702. [Google Scholar] [CrossRef] [PubMed]
- Drane, D.; Maraskovsky, E.; Gibson, R.; Mitchell, S.; Barnden, M.; Moskwa, A.; Shaw, D.; Gervase, B.; Coates, S.; Houghton, M.; et al. Priming of CD4+ and CD8+ T cell responses using a HCV core ISCOMATRIX vaccine: A phase I study in healthy volunteers. Hum. Vaccines 2009, 5, 151–157. [Google Scholar] [CrossRef]
- Duewell, P.; Kisser, U.; Heckelsmiller, K.; Hoves, S.; Stoitzner, P.; Koernig, S.; Morelli, A.B.; Clausen, B.E.; Dauer, M.; Eigler, A.; et al. ISCOMATRIX adjuvant combines immune activation with antigen delivery to dendritic cells in vivo leading to effective cross-priming of CD8+ T cells. J. Immunol. 2011, 187, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Wilson, N.S.; Duewell, P.; Yang, B.; Marsters, S.; Koenig, S.; Latz, E.; Maraskovsky, E.; Baz Morelli, A.; Schurr, M.; Ashkenazi, A. Inflammatory dependent and-independent IL-18 production mediates immunity to the ISCOMATRIX adjuvant. J. Immunol. 2014, 92, 3259–3268. [Google Scholar] [CrossRef] [PubMed]







© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manfredi, F.; Di Bonito, P.; Ridolfi, B.; Anticoli, S.; Arenaccio, C.; Chiozzini, C.; Baz Morelli, A.; Federico, M. The CD8+ T Cell-Mediated Immunity Induced by HPV-E6 Uploaded in Engineered Exosomes Is Improved by ISCOMATRIXTM Adjuvant. Vaccines 2016, 4, 42. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines4040042
Manfredi F, Di Bonito P, Ridolfi B, Anticoli S, Arenaccio C, Chiozzini C, Baz Morelli A, Federico M. The CD8+ T Cell-Mediated Immunity Induced by HPV-E6 Uploaded in Engineered Exosomes Is Improved by ISCOMATRIXTM Adjuvant. Vaccines. 2016; 4(4):42. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines4040042
Chicago/Turabian StyleManfredi, Francesco, Paola Di Bonito, Barbara Ridolfi, Simona Anticoli, Claudia Arenaccio, Chiara Chiozzini, Adriana Baz Morelli, and Maurizio Federico. 2016. "The CD8+ T Cell-Mediated Immunity Induced by HPV-E6 Uploaded in Engineered Exosomes Is Improved by ISCOMATRIXTM Adjuvant" Vaccines 4, no. 4: 42. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines4040042