Predominance of a Drifted Influenza A (H3N2) Clade and Its Association with Age-Specific Influenza Vaccine Effectiveness Variations, Influenza Season 2018–2019

Abstract
:1. Introduction
2. Materials and Methods
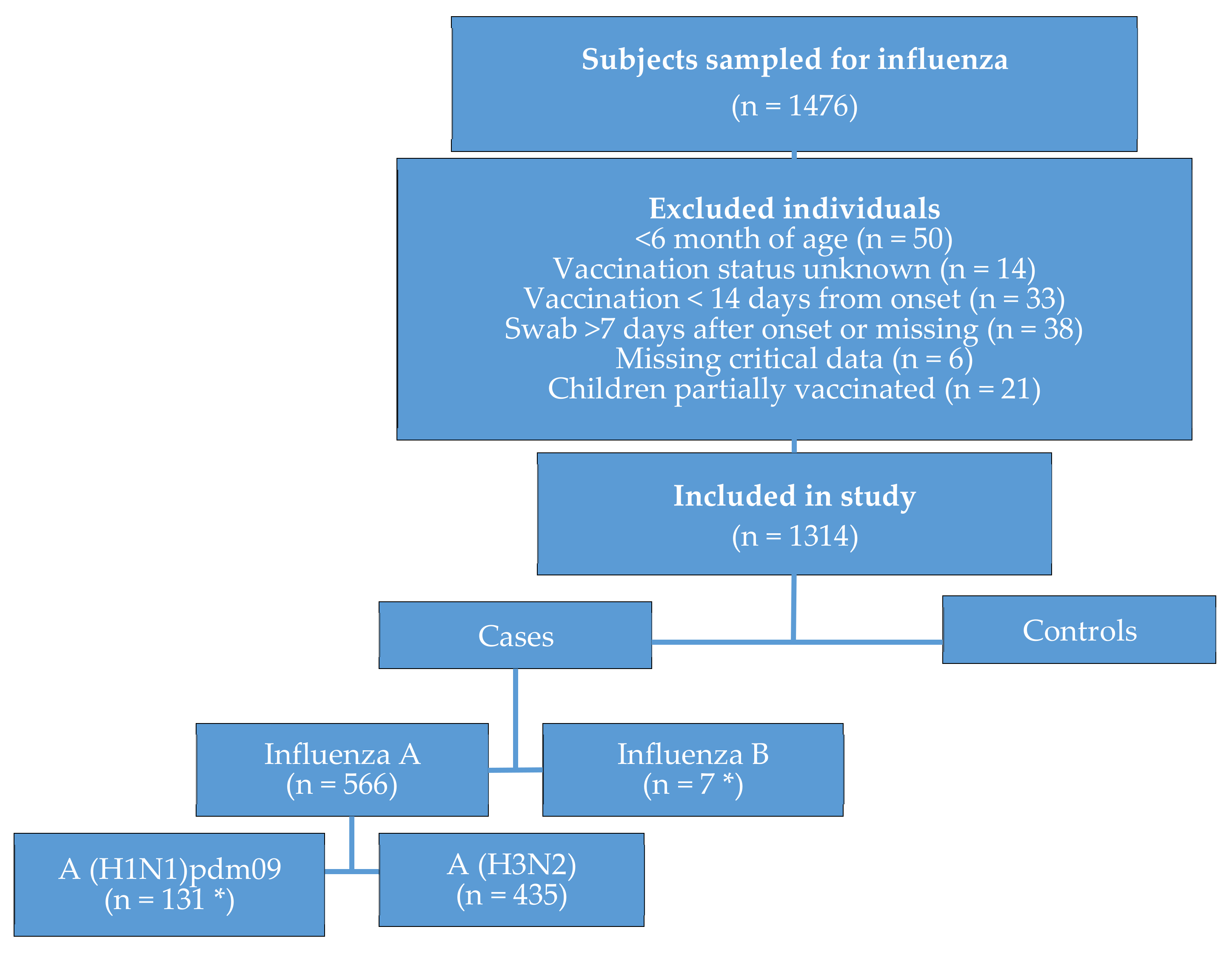
2.1. Study Period and Population
2.2. Laboratory Methods
2.3. Vaccine Effectiveness Study Design
2.4. Statistical Analysis
2.5. Ethical Considerations
3. Results
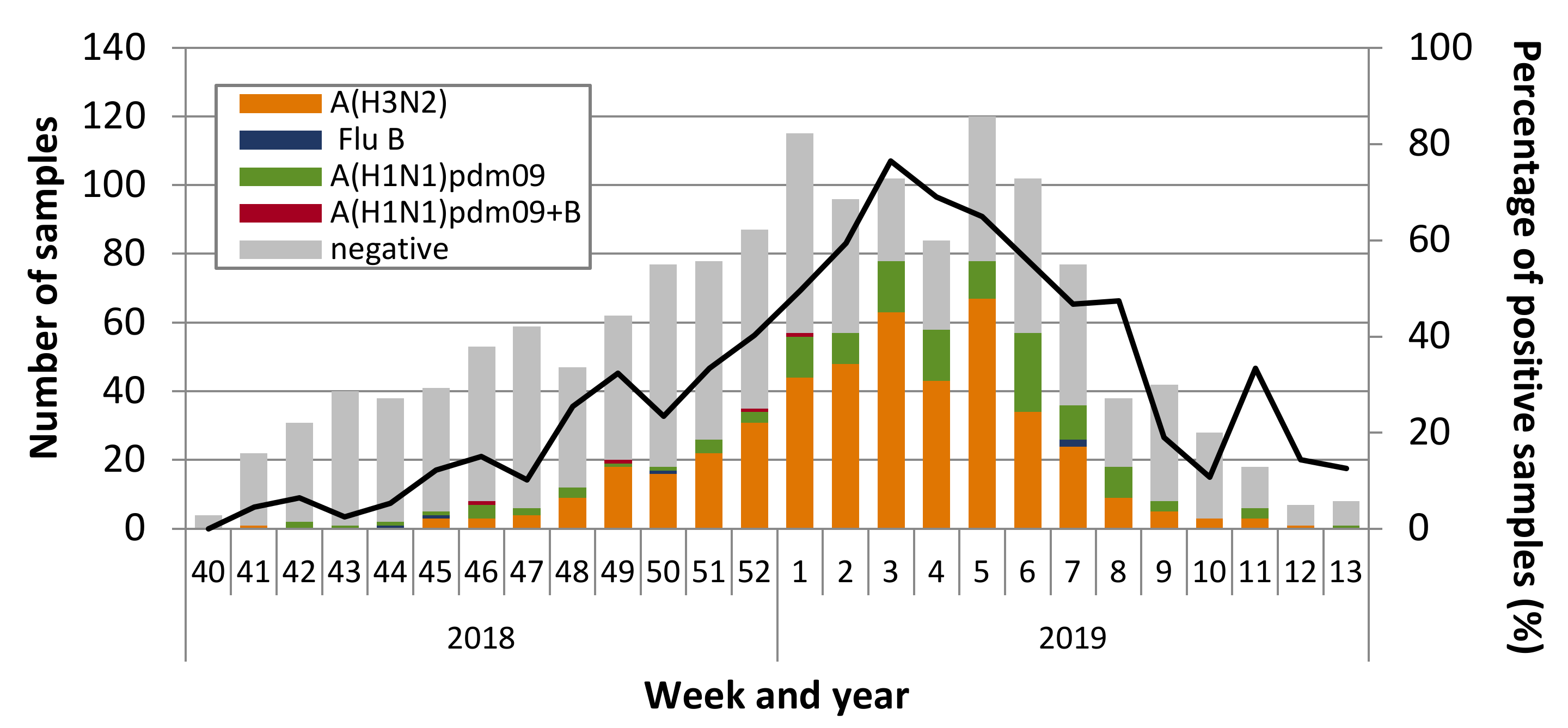
3.1. Influenza Virus Circulation and Characterization
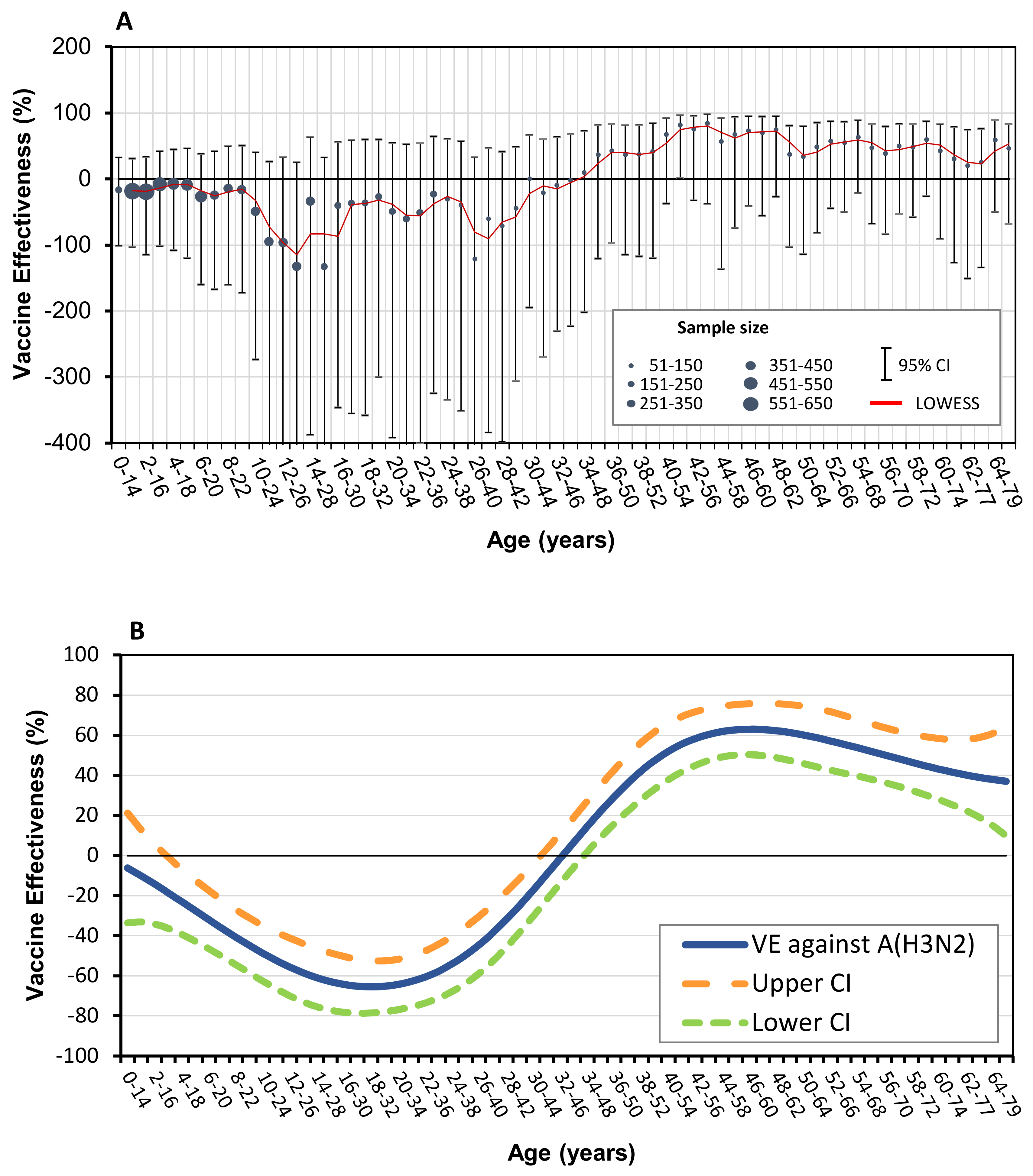
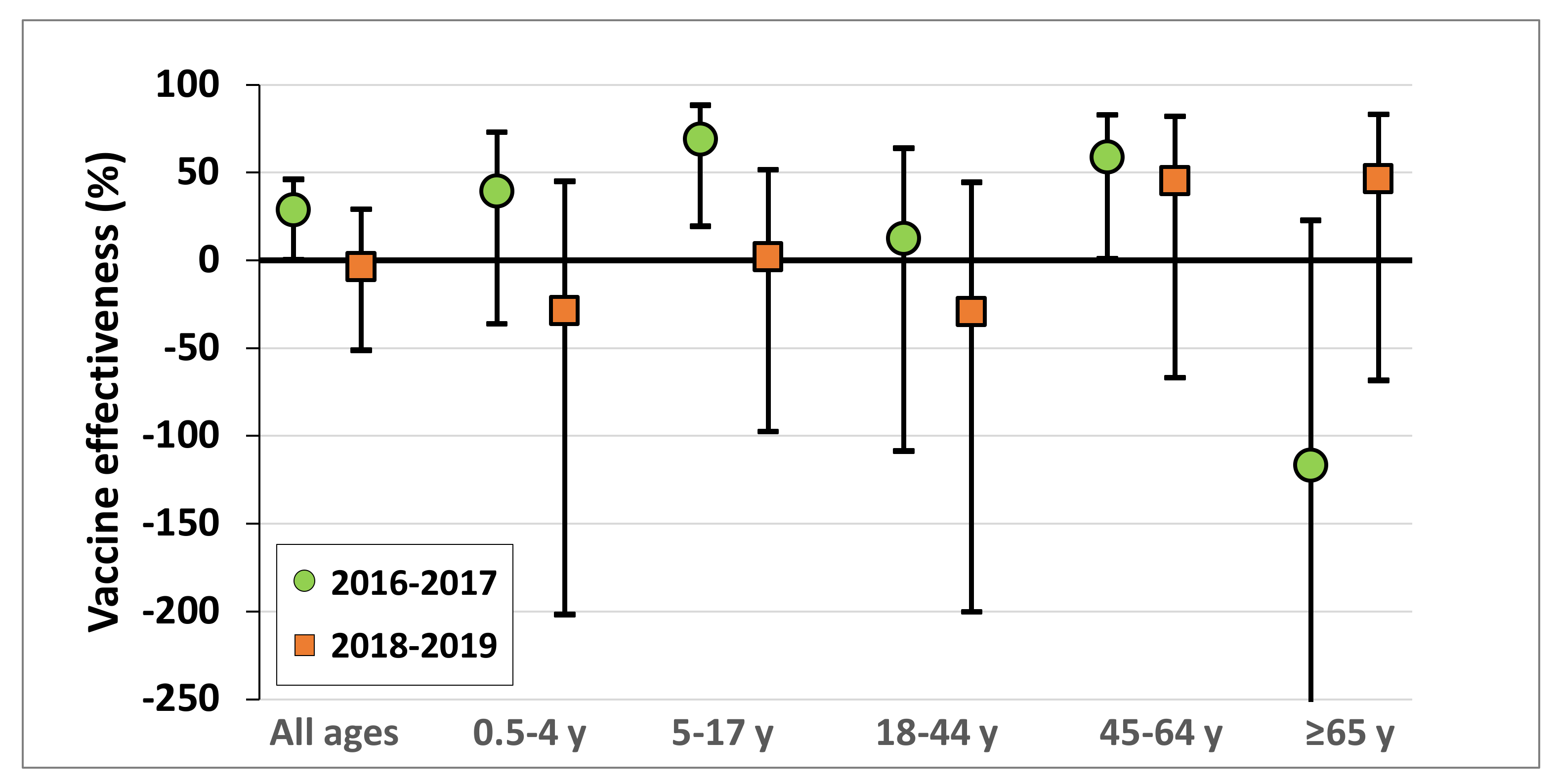
3.2. Vaccine Effectiveness against Influenza A (H3N2)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Addendum to the recommended Composition of Influenza Virus Vaccines for Use in the 2019–2020 Northern Hemisphere Influenza Season. 2019. Available online: https://www.who.int/influenza/vaccines/virus/recommendations/201902_recommendation_addendum.pdf?ua=1 (accessed on 23 May 2019).
- Smith, D.J.; Lapedes, A.S.; De Jong, J.C.; Bestebroer, T.M.; Rimmelzwaan, G.F.; Fouchier, R.A.M.; Osterhaus, A.D.M.E. Mapping the antigenic and genetic evolution of influenza virus. Science 2004, 305, 371–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaron-Yakoby, H.; Sefty, H.; Pando, R.; Dichtiar, R.; Katz, M.; Stein, Y.; Mandelboim, M.; Mendelson, E.; Shohat, T.; Glatman-Freedman, A. Effectiveness of influenza vaccine in preventing medically-attended influenza virus infection in primary care, Israel, influenza seasons 2014/15 and 2015/16. Eurosurveillance 2018, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glatman-Freedman, A.; Drori, Y.; Beni, S.A.; Friedman, N.; Pando, R.; Sefty, H.; Tal, I.; McCauley, J.; Rahav, G.; Keller, N.; et al. Genetic divergence of Influenza A(H3N2) amino acid substitutions mark the beginning of the 2016–2017 winter season in Israel. J. Clin. Virol. 2017, 93, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Israel Center for Disease Control. Surveillance of Influenza-Like Illness in Israel, Weekly Update Report for Week 11, Ending 18 March 2017. Available online: https://www.health.gov.il/PublicationsFiles/flu18032017e.pdf2017 (accessed on 17 October 2017).
- Soebiyanto, R.P.; Gross, D.; Jorgensen, P.; Buda, S.; Bromberg, M.; Kaufman, Z.; Prosenc, K.; Sočan, M.; Alonso, T.V.; Widdowson, M.-A.; et al. Associations between meteorological parameters and influenza activity in Berlin (Germany), Ljubljana (Slovenia), Castile and León (Spain) and Israeli districts. PLoS ONE 2015, 10, e0134701. [Google Scholar] [CrossRef] [Green Version]
- Israel Ministry of Health. Vaccination Guidelines-Influenza (in Hebrew) 2018. Available online: http://www.health.gov.il/UnitsOffice/HD/PH/epidemiology/td/docs/360_Influenza.pdf (accessed on 24 November 2018).
- Stein, Y.; Mandelboim, M.; Sefty, H.; Pando, R.; Mendelson, E.; Shohat, T.; Glatman-Freedman, A.; Muhamed, A.; Arkadi, A.; Yoav, A.; et al. Seasonal influenza vaccine effectiveness in preventing laboratory-confirmed influenza in primary care in Israel, 2016–2017 Season: Insights into novel age-specific analysis. Clin. Infect. Dis. 2017, 66, 1383–1391. [Google Scholar] [CrossRef]
- Meningher, T.; Hindiyeh, M.; Regev, L.; Sherbany, H.; Mendelson, E.; Mandelboim, M. Relationships between A (H1N1) pdm09 influenza infection and infections with other respiratory viruses. Influ. Other Respir. Viruses 2014, 8, 422–430. [Google Scholar] [CrossRef]
- Mandelboim, M.; Glatman-Freedman, A.; Drori, Y.; Sherbany, H.; Pando, R.; Sefty, H.; Zadka, H.; Shohat, T.; Keller, N.; Mendelson, E. Ineffectiveness of the 2014–2015 H3N2 influenza vaccine. Oncotarget 2015, 7, 1185–1192. [Google Scholar] [CrossRef] [Green Version]
- Ndifon, W.; Wingreen, N.S.; Levin, S.A. Differential neutralization efficiency of hemagglutinin epitopes, antibody interference, and the design of influenza vaccines. Proc. Natl. Acad. Sci. USA 2009, 106, 8701–8706. [Google Scholar] [CrossRef] [Green Version]
- Wiley, D.C.; Skehel, J.J. The structure and function of the hemagglutinin membrane glycoprotein of influenza virus. Annu. Rev. Biochem. 1987, 56, 365–394. [Google Scholar] [CrossRef]
- Wedde, M.; Biere, B.; Wolff, T.; Schweiger, B. Evolution of the hemagglutinin expressed by human influenza A(H1N1)pdm09 and A(H3N2) viruses circulating between 2008–2009 and 2013–2014 in Germany. Int. J. Med Microbiol. 2015, 305, 762–775. [Google Scholar] [CrossRef]
- Both, G.W.; Sleigh, M.J.; Cox, N.J.; Kendal, A.P. Antigenic drift in influenza virus H3 hemagglutinin from 1968 to 1980: Multiple evolutionary pathways and sequential amino acid changes at key antigenic sites. J. Virol. 1983, 48, 52–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. WHO Recommendations on the Composition of Influenza Virus Vaccines. Available online: http://www.who.int/influenza/vaccines/virus/recommendations/en/2019 (accessed on 22 April 2019).
- Worldwide Influenza Centre. The Francis Crick Institute. Report Prepared for the WHO Annual Consultation on the Composition of Influenza Vaccine for the Northern Hemisphere 2019–2020. Available online: https://www.crick.ac.uk/sites/default/files/2019-04/Crick%20VCMFeb2019%20report_toPost.pdf (accessed on 21 April 2019).
- Centers for Disease Control and Prevention. Weekly U.S. Influenza Surveillance Report. 2018-2019 Influenza Season Week, 11 Ending March 16, 2019; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2019. [Google Scholar]
- ECDE-WHO. Flu News Europe. Virus Characteristics. Available online: https://flunewseurope.org/VirusCharacteristics (accessed on 23 March 2019).
- Pebody, R.G.; Whitaker, H.; Ellis, J.; Andrews, N.; Marques, D.F.; Cottrell, S.; Reynolds, A.J.; Gunson, R.; Thompson, C.; Galiano, M.; et al. End of season influenza vaccine effectiveness in primary care in adults and children in the United Kingdom in 2018/19. Vaccine 2020, 38, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Skowronski, D.M.; Sabaiduc, S.; Leir, S.; Rose, C.; Zou, M.; Murti, M.; Dickinson, J.; Olsha, R.; Gubbay, J.B.; Croxen, M.; et al. Paradoxical clade- and age-specific vaccine effectiveness during the 2018/19 influenza A(H3N2) epidemic in Canada: Potential imprint-regulated effect of vaccine (I-REV). Eurosurveillance 2019, 24, 1900585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flannery, B.; Kondor, R.J.G.; Chung, J.R.; Gaglani, M.; Reis, M.; Zimmerman, R.K.; Nowalk, M.P.; Jackson, M.L.; Jackson, L.; Monto, A.S.; et al. Spread of Antigenically Drifted Influenza A(H3N2) Viruses and Vaccine Effectiveness in the United States During the 2018–2019 Season. J. Infect. Dis. 2020, 221, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Kissling, E.; Pozo, F.; Buda, S.; Vilcu, A.-M.; Gherasim, A.; Brytting, M.; Domegan, L.; Gómez, V.; Meijer, A.; Lazar, M.; et al. Low 2018/19 vaccine effectiveness against influenza A(H3N2) among 15–64-year-olds in Europe: Exploration by birth cohort. Eurosurveillance 2019, 24, 1900604. [Google Scholar] [CrossRef] [PubMed]
- Flannery, B.; Smith, C.; Garten, R.J.; Levine, M.Z.; Chung, J.R.; Jackson, M.L.; Jackson, L.; Monto, A.S.; Martin, E.T.; Belongia, E.; et al. Influence of Birth cohort on effectiveness of 2015–2016 influenza vaccine against medically attended illness due to 2009 pandemic influenza A(H1N1) virus in the United States. J. Infect. Dis. 2018, 218, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Gostic, K.M.; Ambrose, M.; Worobey, M.; Lloyd-Smith, J.O. Potent protection against H5N1 and H7N9 influenza via childhood hemagglutinin imprinting. Science 2016, 354, 722–726. [Google Scholar] [CrossRef] [Green Version]
- Francis, M.E.; McNeil, M.; Dawe, N.J.; Foley, M.K.; King, M.L.; Ross, T.M.; Kelvin, A.A. Historical H1N1 influenza virus imprinting increases vaccine protection by influencing the activity and sustained production of antibodies elicited at vaccination in ferrets. Vaccines 2019, 7, 133. [Google Scholar] [CrossRef] [Green Version]
- Kosikova, M.; Li, L.; Radvak, P.; Ye, Z.; Wan, X.-F.; Xie, H. Imprinting of repeated influenza A/H3 exposures on antibody quantity and antibody quality: Implications for seasonal vaccine strain selection and vaccine performance. Clin. Infect. Dis. 2018, 67, 1523–1532. [Google Scholar] [CrossRef]
- Kelvin, A.; Zambon, M. Influenza imprinting in childhood and the influence on vaccine response later in life. Eurosurveillance 2019, 24, 1900720. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, S.G.; Price, O.H.; Regan, A.K. Burden, effectiveness and safety of influenza vaccines in elderly, paediatric and pregnant populations. Ther. Adv. Vaccines Immunother. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Glatman-Freedman, A.; Portelli, I.; Jacobs, S.K.; Mathew, J.I.; Slutzman, J.E.; Goldfrank, L.R.; Smith, S.W. Attack rates assessment of the 2009 pandemic H1N1 influenza a in children and their contacts: A systematic review and meta-analysis. PLoS ONE 2012, 7, e50228. [Google Scholar] [CrossRef] [Green Version]
- Baguelin, M.; Flasche, S.; Camacho, A.; Demiris, N.; Miller, E.; Edmunds, W.J. Assessing optimal target populations for influenza vaccination programmes: An evidence synthesis and modelling study. PLoS Med. 2013, 10, e1001527. [Google Scholar] [CrossRef]
- Mossong, J.; Hens, N.; Jit, M.; Beutels, P.; Auranen, K.; Mikolajczyk, R.; Massari, M.; Salmaso, S.; Tomba, G.S.; Wallinga, J.; et al. Social contacts and mixing patterns relevant to the spread of infectious diseases. PLoS Med. 2008, 5, e74. [Google Scholar] [CrossRef] [PubMed]
- Sah, P.; Medlock, J.; Fitzpatrick, M.C.; Singer, B.H.; Galvani, A.P. Optimizing the impact of low-efficacy influenza vaccines. Proc. Natl. Acad. Sci. USA 2018, 115, 5151–5156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization Regional Office for the Eastern Mediterranean. Influenza Monthly Update, March 2019. Available online: http://www.emro.who.int/pandemic-epidemic-diseases/influenza/influenza-monthly-update-march-2019.html (accessed on 9 December 2019).
- Saito, N.; Komori, K.; Suzuki, M.; Kishikawa, T.; Yasaka, T.; Ariyoshi, K. Dose-dependent negative effects of prior multiple vaccinations against influenza A and influenza b among schoolchildren: A study of kamigoto Island in japan during the 2011–2012, 2012–2013, and 2013–2014 influenza seasons. Clin. Infect. Dis. 2018, 67, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Saito, N.; Komori, K.; Suzuki, M.; Morimoto, K.; Kishikawa, T.; Yasaka, T.; Ariyoshi, K. Negative impact of prior influenza vaccination on current influenza vaccination among people infected and not infected in prior season: A test-negative case-control study in Japan. Vaccine 2017, 35, 687–693. [Google Scholar] [CrossRef]
- Castilla, J.; Navascués, A.; Casado, I.; Pérez-García, A.; Aguinaga, A.; Ezpeleta, G.; Pozo, F.; Ezpeleta, C.; Martínez-Baz, I. Interim effectiveness of trivalent influenza vaccine in a season dominated by lineage mismatched influenza B, northern Spain, 2017/18. Eurosurveillance 2018, 23. [Google Scholar] [CrossRef] [Green Version]
- Buchan, S.A.; Chung, H.; Campitelli, M.A.; Crowcroft, N.S.; Gubbay, J.B.; Karnauchow, T.; Katz, K.; McGeer, A.J.; McNally, J.D.; Richardson, D.; et al. Vaccine effectiveness against laboratory-confirmed influenza hospitalizations among young children during the 2010–2011 to 2013–2014 influenza seasons in Ontario, ON, Canada. PLoS ONE 2017, 12, e0187834. [Google Scholar] [CrossRef] [Green Version]
- Belongia, E.A.; Skowronski, D.M.; McLean, H.Q.; Chambers, C.; Sundaram, M.E.; De Serres, G. Repeated annual influenza vaccination and vaccine effectiveness: Review of evidence. Expert Rev. Vaccines 2017, 16, 723–736. [Google Scholar] [CrossRef] [Green Version]
- Petrie, J.G.; Monto, A.S. Untangling the effects of prior vaccination on subsequent influenza vaccine effectiveness. J. Infect. Dis. 2017, 215, 841–843. [Google Scholar] [CrossRef] [Green Version]
- Nichols, M.K.; Andrew, M.K.; Ye, L.; Hatchette, T.F.; Ambrose, A.; Boivin, G.; Haguinet, F. The impact of prior season vaccination on subsequent influenza Vaccine Effectiveness (VE) to prevent influenza-related hospitalizations over four influenza seasons in Canada. Clin. Infect. Dis. 2018, 69, 970–979. [Google Scholar] [CrossRef]
- Casado, I.; Domínguez, À.; Toledo, D.; Chamorro, J.; Astray, J.; Egurrola, M.; Fernández-Sierra, M.A.; Martín, V.; Morales-Suárez-Varela, M.; Godoy, P.; et al. Repeated influenza vaccination for preventing severe and fatal influenza infection in older adults: A multicentre case—Control study. Can. Med. Assoc. J. 2018, 190, E3–E12. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Conrol and Prevention. Vaccine Effectiveness: How Well do the Flu Vaccines Work? Available online: https://www.cdc.gov/flu/vaccines-work/vaccineeffect.htm (accessed on 15 January 2020).
- Amrhein, V.; Greenland, S.; McShane, B. Scientists rise up against statistical significance. Nature 2019, 567, 305–307. [Google Scholar] [CrossRef] [Green Version]
- Wasserstein, R.L.; Schirm, A.L.; Lazar, N.A. Moving to a world beyond “p <0.05”. Am. Stat. 2019, 73, 1–19. [Google Scholar]
- Monto, A.S.; Petrie, J.G. Improving influenza vaccine effectiveness: Ways to begin solving the problem. Clin. Infect. Dis. 2019, 69, 1824–1826. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Controls (n = 744) | Cases (n = 435) | Total (n = 1179) | p-Value a |
|---|---|---|---|---|
| Age group (years) | ||||
| 0.5-4 | 252 | 104 | 356 | >0.01 |
| 17-May | 136 | 183 | 319 | |
| 18-44 | 204 | 96 | 300 | |
| 45-64 | 97 | 29 | 126 | |
| ≥ 65 | 55 | 23 | 78 | |
| Sex | ||||
| Male | 368 | 218 | 586 | 0.83 |
| Female | 376 | 217 | 593 | |
| Background Disease | ||||
| No | 596 | 387 | 983 | >0.01 |
| Yes | 148 | 48 | 196 | |
| Days between Symptom onset and Swabbing | ||||
| 0-1 | 302 | 155 | 457 | 0.09 |
| 04-Feb | 379 | 250 | 629 | |
| 07-May | 63 | 30 | 93 | |
| Month of Sample Collection | ||||
| October | 107 | 1 | 108 | >0.01 |
| November | 150 | 18 | 168 | |
| December | 183 | 110 | 293 | |
| January | 230 | 135 | 365 | |
| February | 126 | 69 | 195 | |
| March | 43 | 7 | 50 | |
| Influenza Vaccine | ||||
| No | 634 | 360 | 994 | 0.26 |
| Yes | 110 | 75 | 185 | |
| Age (Years) | Adjustment | Cases | Controls | Adjusted Vaccine Effectiveness | |||||
|---|---|---|---|---|---|---|---|---|---|
| % | Vac a | All | % | Vac a | All | 95% CI | % | ||
| All ages | Crude | 17.2 | 75 | 435 | 14.8 | 110 | 744 | −65.5 to 12.9 | −20.1 |
| All ages | Adjusted a | 17.2 | 75 | 435 | 14.8 | 110 | 744 | −51.2. to 29.1 | −3.5 |
| 0.5-4 | Adjusted b | 14.4 | 15 | 104 | 8.3 | 21 | 252 | −201.8 to 5 | −28.8 |
| 17-May | Adjusted b | 15.3 | 28 | 183 | 12.5 | 17 | 136 | −97.7 to 51.6 | 2.1 |
| 18-44 | Adjusted b | 13.5 | 13 | 96 | 8.3 | 17 | 204 | −200.2 to 44.5 | −29.1 |
| 45-64 | Adjusted b | 20.7 | 6 | 29 | 24.7 | 24 | 97 | −66.8 to 82.1 | 45.4 |
| 65≤ | Adjusted b | 56.5 | 13 | 23 | 56.4 | 31 | 55 | −68.5 to 83.0 | 46.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glatman-Freedman, A.; Pando, R.; Sefty, H.; Omer, I.; Rosenberg, A.; Drori, Y.; Nemet, I.; Mendelson, E.; Keinan-Boker, L.; Mandelboim, M., on behalf of the Israeli Influenza Surveillance Network (IISN). Predominance of a Drifted Influenza A (H3N2) Clade and Its Association with Age-Specific Influenza Vaccine Effectiveness Variations, Influenza Season 2018–2019. Vaccines 2020, 8, 78. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines8010078
Glatman-Freedman A, Pando R, Sefty H, Omer I, Rosenberg A, Drori Y, Nemet I, Mendelson E, Keinan-Boker L, Mandelboim M on behalf of the Israeli Influenza Surveillance Network (IISN). Predominance of a Drifted Influenza A (H3N2) Clade and Its Association with Age-Specific Influenza Vaccine Effectiveness Variations, Influenza Season 2018–2019. Vaccines. 2020; 8(1):78. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines8010078
Chicago/Turabian StyleGlatman-Freedman, Aharona, Rakefet Pando, Hanna Sefty, Itay Omer, Alina Rosenberg, Yaron Drori, Ital Nemet, Ella Mendelson, Lital Keinan-Boker, and Michal Mandelboim on behalf of the Israeli Influenza Surveillance Network (IISN). 2020. "Predominance of a Drifted Influenza A (H3N2) Clade and Its Association with Age-Specific Influenza Vaccine Effectiveness Variations, Influenza Season 2018–2019" Vaccines 8, no. 1: 78. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines8010078