Design and Characterization of a DNA Vaccine Based on Spike with Consensus Nucleotide Sequence against Infectious Bronchitis Virus

 , ,
, , 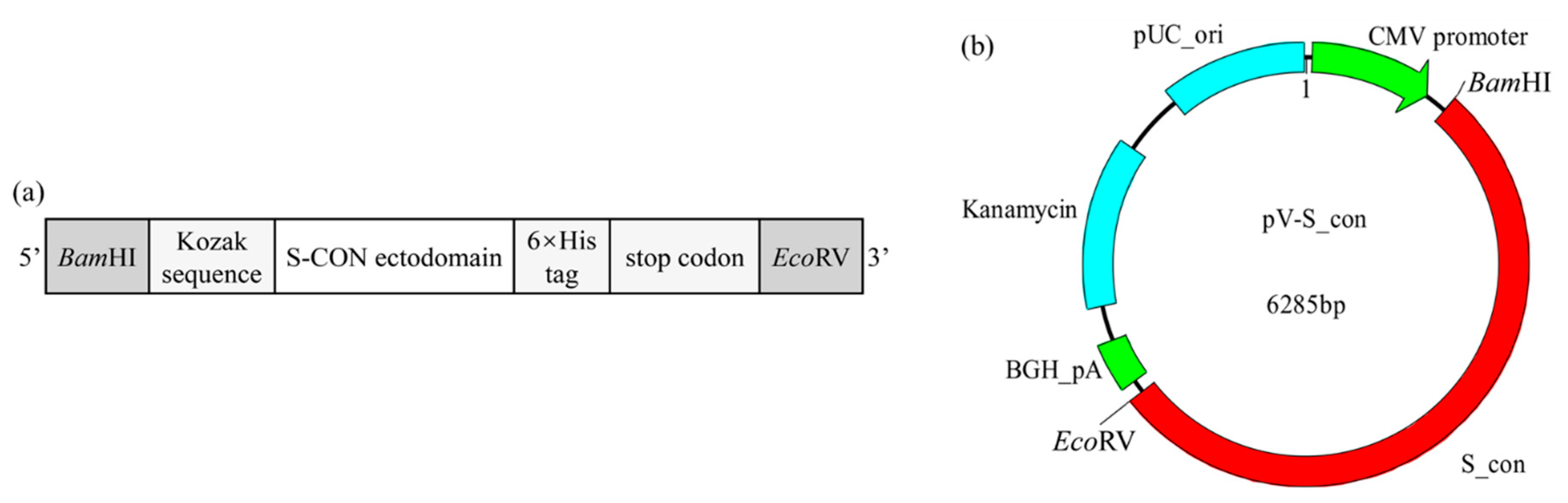{kind=link}
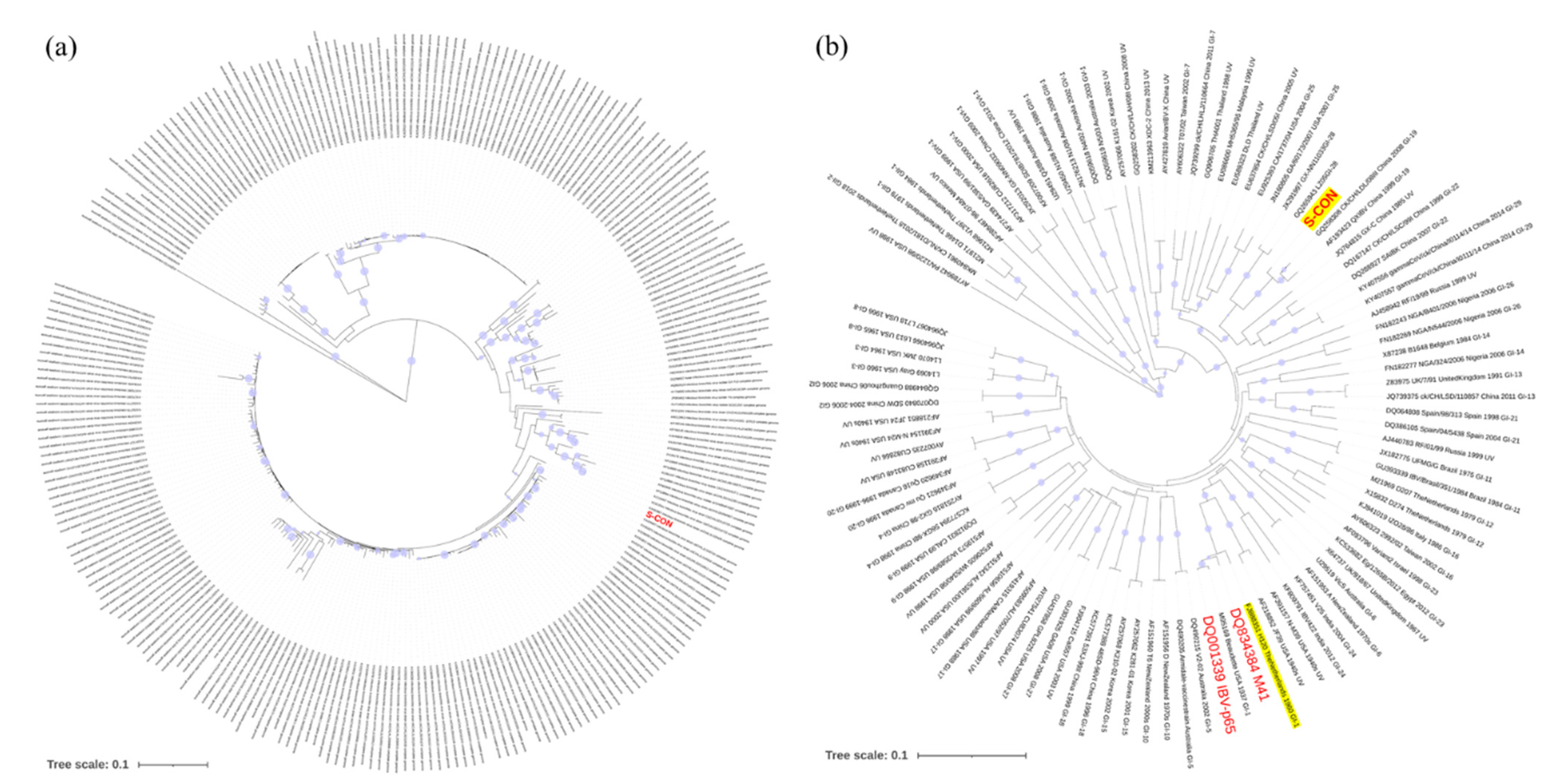{kind=link}
{kind=link}
{kind=link}
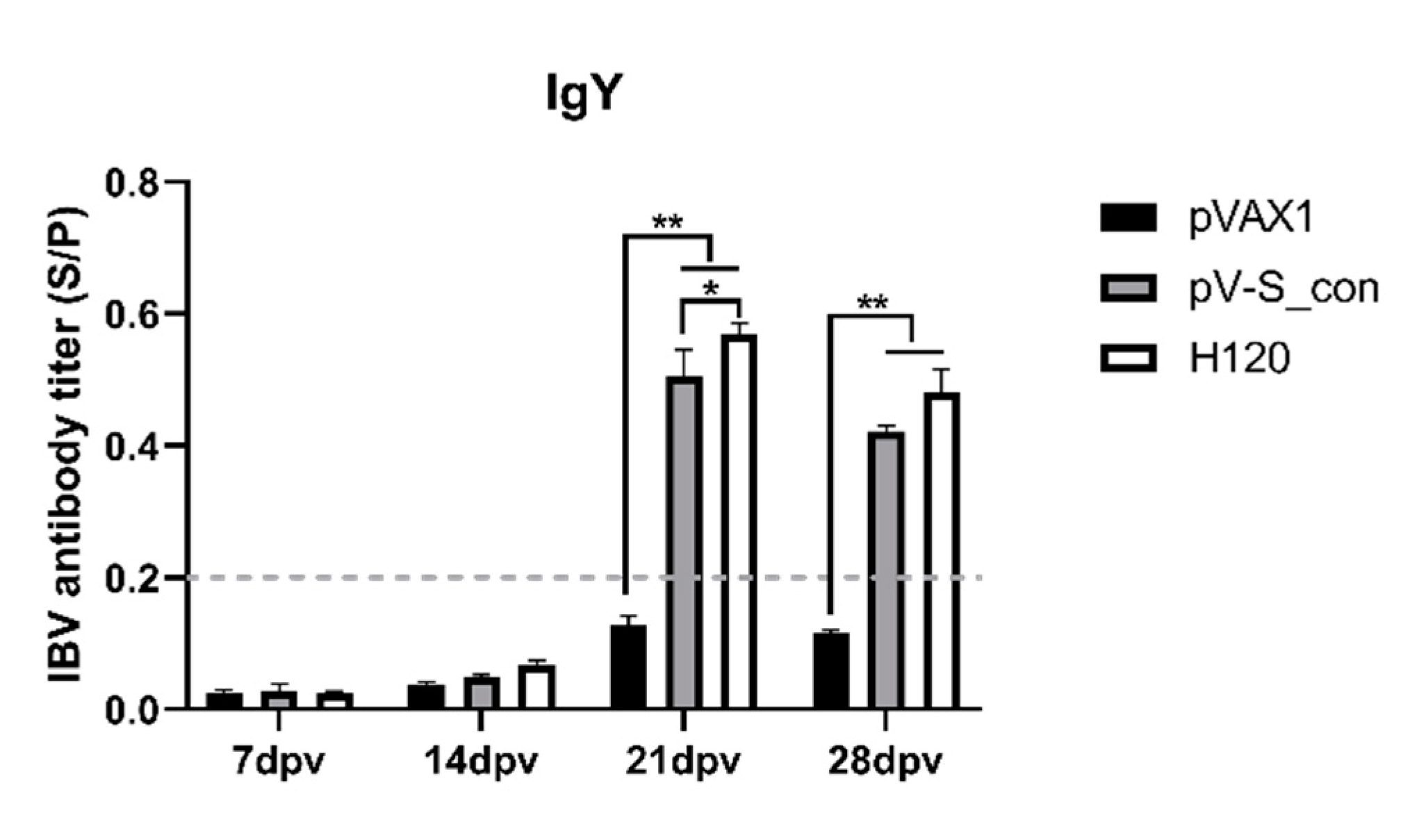{kind=link}
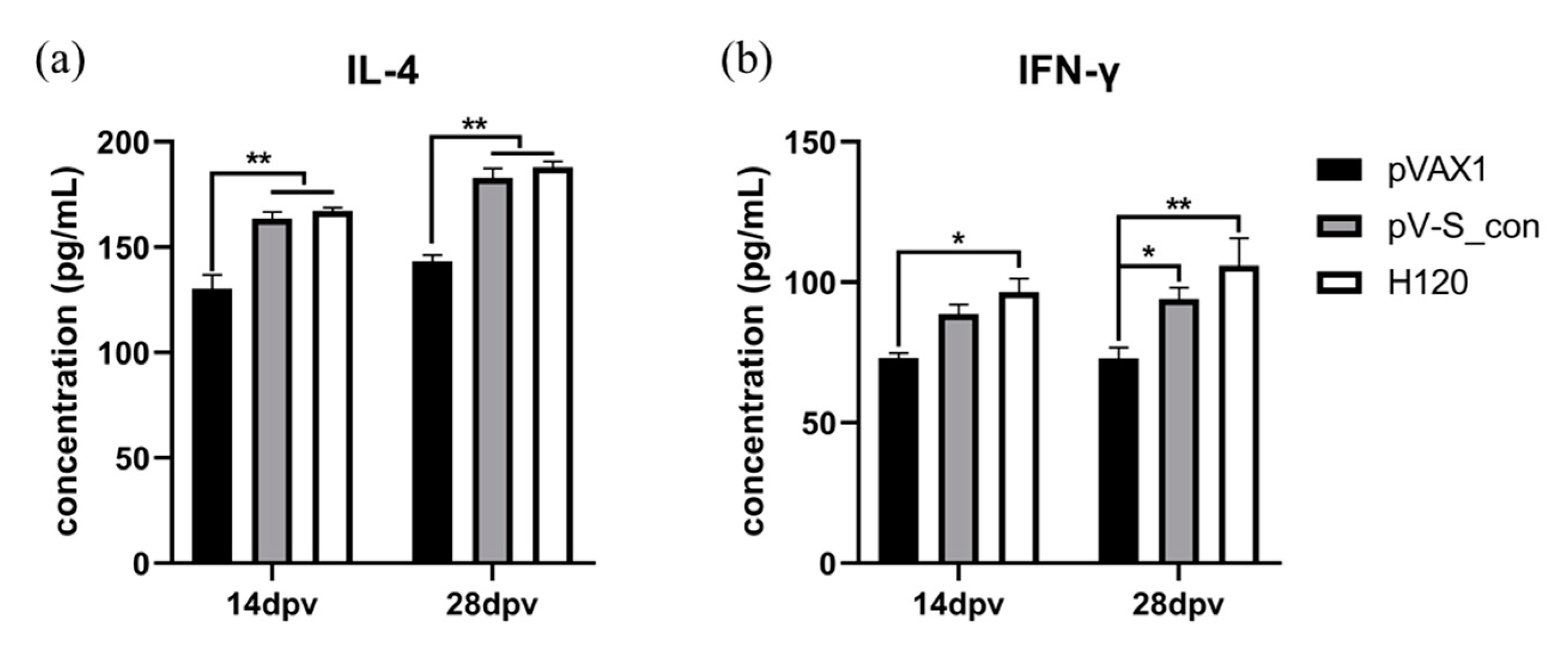{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Viruses and Cells
2.3. Design and Analysis of IBV S Protein Consensus Nucleotide Sequence
2.4. Design and Construction of IBV DNA Vaccine
2.5. Analysis of S_con Expression by Indirect Immunofluorescence Assay and Western Blot
2.6. Neutralizing Antibody Induced by IBV S_con DNA Vaccine in Mice
2.7. Immunization and Challenge
2.8. Sample Collection
2.9. Evaluation the Efficacy of IBV S_con DNA Vaccine in Chickens
2.10. Statistical Analysis
3. Results
3.1. Phylogenetic Analysis of S-con
3.2. Analyses of S_con Protein Expression in Vitro
3.3. Neutralizing Antibody Induced by IBV S_con DNA Vaccine in Mice
3.4. IBV-Specific Antibodies in Immunized Chickens
3.5. Cytokines after Immunization in Chickens
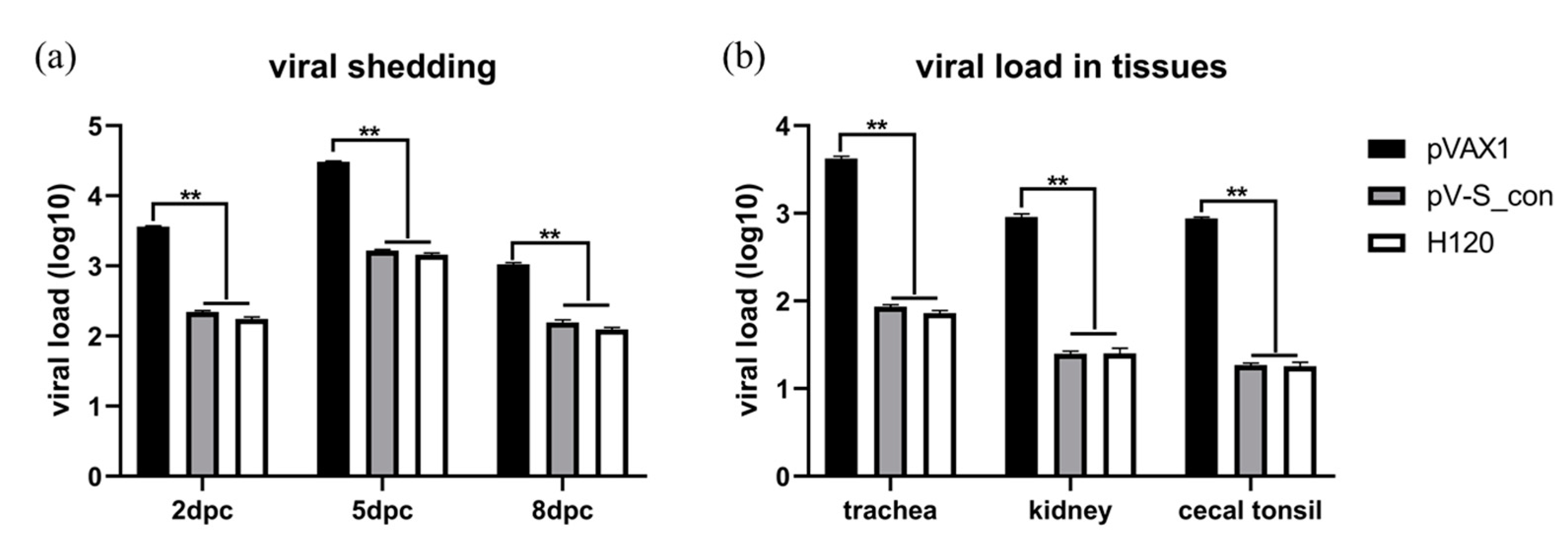
3.6. Evaluation of Protection Against IBV M41 Challenge
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cavanagh, D. Coronavirus avian infectious bronchitis virus. Vet. Res. 2007, 38, 281–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, J.K.A.; Jackwood, M.; Jones, R.C. The long view: 40 years of infectious bronchitis research. Avian Pathol. 2012, 41, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Eldemery, F.; Joiner, K.S.; Toro, H.; Van Santen, V.L. Protection against infectious bronchitis virus by spike ectodomain subunit vaccine. Vaccine 2017, 35, 5864–5871. [Google Scholar] [CrossRef]
- Ellis, S.; Keep, S.P.; Britton, P.; De Wit, S.; Bickerton, E.; Vervelde, L. Recombinant Infectious Bronchitis Viruses Expressing Chimeric Spike Glycoproteins Induce Partial Protective Immunity against Homologous Challenge despite Limited Replication In Vivo. J. Virol. 2018, 92, e01473-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickramasinghe, I.A.; Van Beurden, S.; Weerts, E.; Verheije, M.H. The avian coronavirus spike protein. Virus Res. 2014, 194, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Bande, F.; Arshad, S.S.; Omar, A.R.; Hair-Bejo, M.; Mahmuda, A.; Nair, V. Global distributions and strain diversity of avian infectious bronchitis virus: A review. Anim. Health Res. Rev. 2017, 18, 70–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Z.; Liwen, X.; Ren, M.; Sheng, J.; Ma, T.; Sun, J.; Zhao, Y.; Liu, S. Genetic, antigenic and pathogenic characterization of avian coronaviruses isolated from pheasants (Phasianus colchicus) in China. Vet. Microbiol. 2019, 240, 108513. [Google Scholar] [CrossRef] [PubMed]
- Molenaar, R.J.; Dijkman, R.; De Wit, J.J. Characterization of infectious bronchitis virus D181, a new serotype (GII-2). Avian Pathol. 2020, 49, 243–250. [Google Scholar] [CrossRef]
- Valastro, V.; Holmes, E.C.; Britton, P.; Fusaro, A.; Jackwood, M.W.; Cattoli, G.; Monne, I. S1 gene-based phylogeny of infectious bronchitis virus: An attempt to harmonize virus classification. Infect. Genet. Evol. 2016, 39, 349–364. [Google Scholar] [CrossRef]
- Jackwood, M.W. Review of Infectious Bronchitis Virus around the World. Avian Dis. 2012, 56, 634–641. [Google Scholar] [CrossRef] [Green Version]
- Bande, F.; Arshad, S.S.; Bejo, M.H.; Moeini, H.; Omar, A.R. Progress and Challenges toward the Development of Vaccines against Avian Infectious Bronchitis. J. Immunol. Res. 2015, 2015, 424860. [Google Scholar] [CrossRef] [Green Version]
- Bagarazzi, M.L.; Yan, M.P.J.; Morrow, X.; Shen, R.L.; Parker, J.C.; Lee, M.; Giffear, P.; Pankhong, A.S.; Khan, K.E.; Broderick, C.; et al. Immunotherapy against Hpv16/18 Generates Potent Th1 and Cytotoxic Cellular Immune Responses. Sci. Transl. Med. 2012, 4, 155ra38. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.-W.; Cheng, T.-J.R.; Huang, Y.; Jan, J.-T.; Ma, S.-H.; Yu, A.L.; Wong, C.-H.; Ho, D.D. A consensus-hemagglutinin-based DNA vaccine that protects mice against divergent H5N1 influenza viruses. Proc. Natl. Acad. Sci. USA 2008, 105, 13538–13543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muthumani, K.; Falzarano, D.; Reuschel, E.L.; Tingey, C.; Flingai, S.; Villarreal, D.O.; Wise, M.C.; Patel, A.; Izmirly, A.; Aljuaid, A.; et al. A synthetic consensus anti-spike protein DNA vaccine induces protective immunity against Middle East respiratory syndrome coronavirus in nonhuman primates. Sci. Transl. Med. 2015, 7, 301ra132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santra, S.; Korber, B.; Muldoon, M.R.; Barouch, D.H.; Nabel, G.J.; Gao, F.; Hahn, B.H.; Haynes, B.F.; Letvin, N.L. A centralized gene-based HIV-1 vaccine elicits broad cross-clade cellular immune responses in rhesus monkeys. Proc. Natl. Acad. Sci. USA 2008, 105, 10489–10494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vu, H.L.X.; Ma, F.; Laegreid, W.W.; Pattnaik, A.K.; Steffen, D.; Doster, A.R.; Osorio, F.A. A Synthetic Porcine Reproductive and Respiratory Syndrome Virus Strain Confers Unprecedented Levels of Heterologous Protection. J. Virol. 2015, 89, 12070–12083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Job, E.R.; Ysenbaert, T.; Smet, A.; Christopoulou, I.; Strugnell, T.; Oloo, E.O.; Oomen, R.P.; Kleanthous, H.; Vogel, T.U.; Saelens, X. Broadened immunity against influenza by vaccination with computationally designed influenza virus N1 neuraminidase constructs. NPJ Vaccines 2018, 3, 55. [Google Scholar] [CrossRef]
- Ping, X.; Hu, W.; Xiong, R.; Zhang, X.; Teng, Z.; Ding, M.; Li, L.; Chang, C.; Ke, X. Generation of a broadly reactive influenza H1 antigen using a consensus HA sequence. Vaccine 2018, 36, 4837–4845. [Google Scholar] [CrossRef]
- Xie, X.; Zhao, C.; He, Q.; Qiu, T.; Yuan, S.; Ding, L.; Liu, L.; Jiang, L.; Wang, J.; Zhang, L.; et al. Influenza Vaccine with Consensus Internal Antigens as Immunogens Provides Cross-Group Protection against Influenza a Viruses. Front. Microbiol. 2019, 10, 1630. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Morrow, M.P.; Chu, J.S.; Racine, T.; Reed, C.C.; Khan, A.S.; Broderick, K.E.; Kim, J.J.; Kobinger, G.; Sardesai, N.Y.; et al. Broad cross-protective anti-hemagglutination responses elicited by influenza microconsensus DNA vaccine. Vaccine 2018, 36, 3079–3089. [Google Scholar] [CrossRef]
- Guy, J.S. Isolation and Propagation of Coronaviruses in Embryonated Eggs. Comput. Biol. 2015, 1282, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Kint, J.; Maier, H.J.; Jagt, E. Quantification of Infectious Bronchitis Coronavirus by Titration in Vitro and In Ovo. Comput. Biol. 2014, 1282, 89–98. [Google Scholar] [CrossRef]
- Han, X.; Tian, Y.; Guan, R.; Gao, W.; Yang, X.; Zhou, L.; Wang, H.-N. Infectious Bronchitis Virus Infection Induces Apoptosis during Replication in Chicken Macrophage HD11 Cells. Viruses 2017, 9, 198. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. Mafft Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 4, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, S.; Peng, J.; Xiao, Y.; Liu, Y.; Hao, W.; Yang, X.; Wang, H.-N.; Gao, R. The Construction and Immunoadjuvant Activities of the Oral Interleukin-17B Expressed by Lactobacillus plantarum NC8 Strain in the Infectious Bronchitis Virus Vaccination of Chickens. Vaccines 2020, 8, 282. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Jiang, L.; Zhao, W.-J.; Liu, L.; Zhao, Y.; Shao, Y.; Li, H.; Han, Z.; Liu, S. Identification and molecular characterization of a novel serotype infectious bronchitis virus (GI-28) in China. Vet. Microbiol. 2017, 198, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Xu, L.; Ren, M.; Shen, J.; Han, Z.; Sun, J.; Zhao, Y.; Liu, S. Novel genotype of infectious bronchitis virus isolated in China. Vet. Microbiol. 2019, 230, 178–186. [Google Scholar] [CrossRef]
- Sheng, J.; Ren, M.; Han, Z.; Sun, J.; Zhao, Y.; Liu, S. Genetic and antigenic heterogeneity of GI-1/Massachusetts lineage infectious bronchitis virus variants recently isolated in China. Poult. Sci. 2020, 99, 5440–5451. [Google Scholar] [CrossRef]
- Farsang, A.; Ros, C.; Renström, L.H.M.; Baule, C.; Soos, T.; Belák, S. Molecular epizootiology of infectious bronchitis virus in Sweden indicating the involvement of a vaccine strain. Avian Pathol. 2002, 31, 229–236. [Google Scholar] [CrossRef] [Green Version]
- Han, Z.; Zhao, W.; Chen, Y.; Xu, Q.; Sun, J.; Zhang, T.; Zhao, Y.; Liang, S.; Gao, M.; Shengwang, L.; et al. Genetic, antigenic, and pathogenic characteristics of avian infectious bronchitis viruses genotypically related to 793/B in China. Vet. Microbiol. 2017, 203, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Han, Z.; Jiang, L.; Sun, J.; Zhao, Y.; Liu, S. Genetic diversity of avian infectious bronchitis virus in China in recent years. Infect. Genet. Evol. 2018, 66, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, H.; Zhao, J.; Guo-Zhong, Z.; Jin, J.-H.; Zhang, G.-Z. Evolution of infectious bronchitis virus in China over the past two decades. J. Gen. Virol. 2016, 97, 1566–1574. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D.; Davis, P.J.; Darbyshire, J.H.; Peters, R.W. Coronavirus Ibv—Virus Retaining Spike Glycopolypeptide-S2 but Not S1 Is Unable to Induce Virus-Neutralizing or Hemagglutination-Inhibiting Antibody, or Induce Chicken Tracheal Protection. J. Gen. Virol. 1986, 67, 1435–1442. [Google Scholar] [CrossRef]
- Cavanagh, D.; Davis, P.J.; Mockett, A. Amino acids within hypervariable region 1 of avian coronavirus IBV (Massachusetts serotype) spike glycoprotein are associated with neutralization epitopes. Virus Res. 1988, 11, 141–150. [Google Scholar] [CrossRef]
- Moore, K.M.; Jackwood, M.W.; Hilt, D.A. Identification of Amino Acids Involved in a Serotype and Neutralization Specific Epitope within the S1 Subunit of Avian Infectious Bronchitis Virus. Arch. Virol. 1997, 142, 2249–2256. [Google Scholar] [CrossRef]
- Wu, X.; Zhai, X.; Lai, Y.; Zuo, L.; Zhang, Y.; Mei, X.; Xiang, R.; Kang, Z.; Zhou, L.; Wang, H. Construction and Immunogenicity of Novel Chimeric Virus-Like Particles Bearing Antigens of Infectious Bronchitis Virus and Newcastle Disease Virus. Viruses 2019, 11, 254. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Chen, X.-W.; Tong, T.-Z.; Ye, Y.; Liao, M.; Fan, H. BacMam virus-based surface display of the infectious bronchitis virus (IBV) S1 glycoprotein confers strong protection against virulent IBV challenge in chickens. Vaccine 2014, 32, 664–670. [Google Scholar] [CrossRef]
- Zhao, R.; Sun, J.; Qi, T.; Zhao, W.; Han, Z.; Yang, X.; Liu, S. Recombinant Newcastle disease virus expressing the infectious bronchitis virus S1 gene protects chickens against Newcastle disease virus and infectious bronchitis virus challenge. Vaccine 2017, 35, 2435–2442. [Google Scholar] [CrossRef]
- Winter, C.; Schwegmann-Wessels, C.; Neumann, U.; Herrler, G. The Spike Protein of Infectious Bronchitis Virus Is Retained Intracellularly by a Tyrosine Motif. J. Virol. 2007, 82, 2765–2771. [Google Scholar] [CrossRef] [Green Version]
- Tan, L.; Zhang, Y.; Liu, F.; Yuan, Y.; Zhan, Y.; Sun, Y.; Qiu, X.; Meng, C.; Song, C.; Ding, C. Infectious bronchitis virus poly-epitope-based vaccine protects chickens from acute infection. Vaccine 2016, 34, 5209–5216. [Google Scholar] [CrossRef] [PubMed]
- Belkasmi, S.F.Z.; Fellahi, S.; Umar, S.; Delpont, M.; Delverdier, M.; Lucas, M.-N.; Bleuart, C.; Kichou, F.; Nassik, S.; Guerin, J.-L.; et al. Efficacy of Massachusetts and 793B Vaccines against Infectious Bronchitis Moroccan-Italy 02 Virus in Specific-Pathogen-Free Chickens and Commercial Broilers. Avian Dis. 2017, 61, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Okino, C.; Alessi, A.C.; Montassier, M.D.F.S.; Rosa, A.J.; Wang, X.; Montassier, H.J. Humoral and Cell-Mediated Immune Responses to Different Doses of Attenuated Vaccine against Avian Infectious Bronchitis Virus. Viral Immunol. 2013, 26, 259–267. [Google Scholar] [CrossRef] [PubMed]
- De Wit, J.J.; Cook, J.K.A. Factors influencing the outcome of infectious bronchitis vaccination and challenge experiments. Avian Pathol. 2014, 43, 485–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batlle, E.; Massague, J. Transforming Growth Factor-Beta Signaling in Immunity and Cancer. Immunity 2019, 50, 924–940. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuo, L.; Yan, W.; Song, Z.; Li, H.; Xie, X.; Gu, K.; Ma, P.; Tian, Y.; Zhou, C.; Zhao, Y.; et al. Design and Characterization of a DNA Vaccine Based on Spike with Consensus Nucleotide Sequence against Infectious Bronchitis Virus. Vaccines 2021, 9, 50. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9010050
Zuo L, Yan W, Song Z, Li H, Xie X, Gu K, Ma P, Tian Y, Zhou C, Zhao Y, et al. Design and Characterization of a DNA Vaccine Based on Spike with Consensus Nucleotide Sequence against Infectious Bronchitis Virus. Vaccines. 2021; 9(1):50. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9010050
Chicago/Turabian StyleZuo, Lei, Wenjun Yan, Zhou Song, Hao Li, Xin Xie, Kui Gu, Peng Ma, Yiming Tian, Changyu Zhou, Yu Zhao, and et al. 2021. "Design and Characterization of a DNA Vaccine Based on Spike with Consensus Nucleotide Sequence against Infectious Bronchitis Virus" Vaccines 9, no. 1: 50. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9010050