
Update on the Inactivation Procedures for the Vaccine Development Prospects of a New Highly Virulent RGNNV Isolate

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Virus
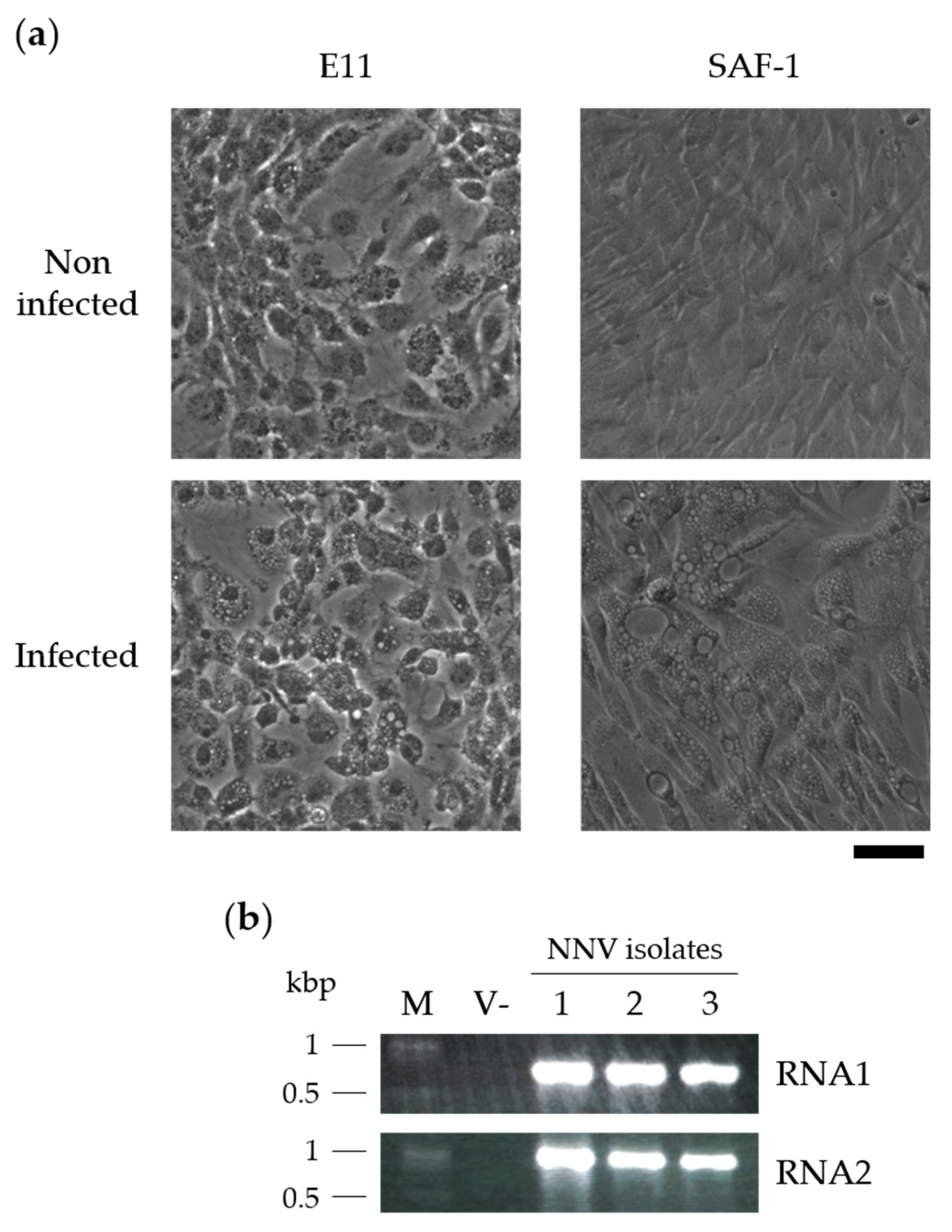
2.2. Microscopy
2.3. Isolation of NNV
2.4. RNA Isolation and cDNA Synthesis
2.5. PCR and Sequencing
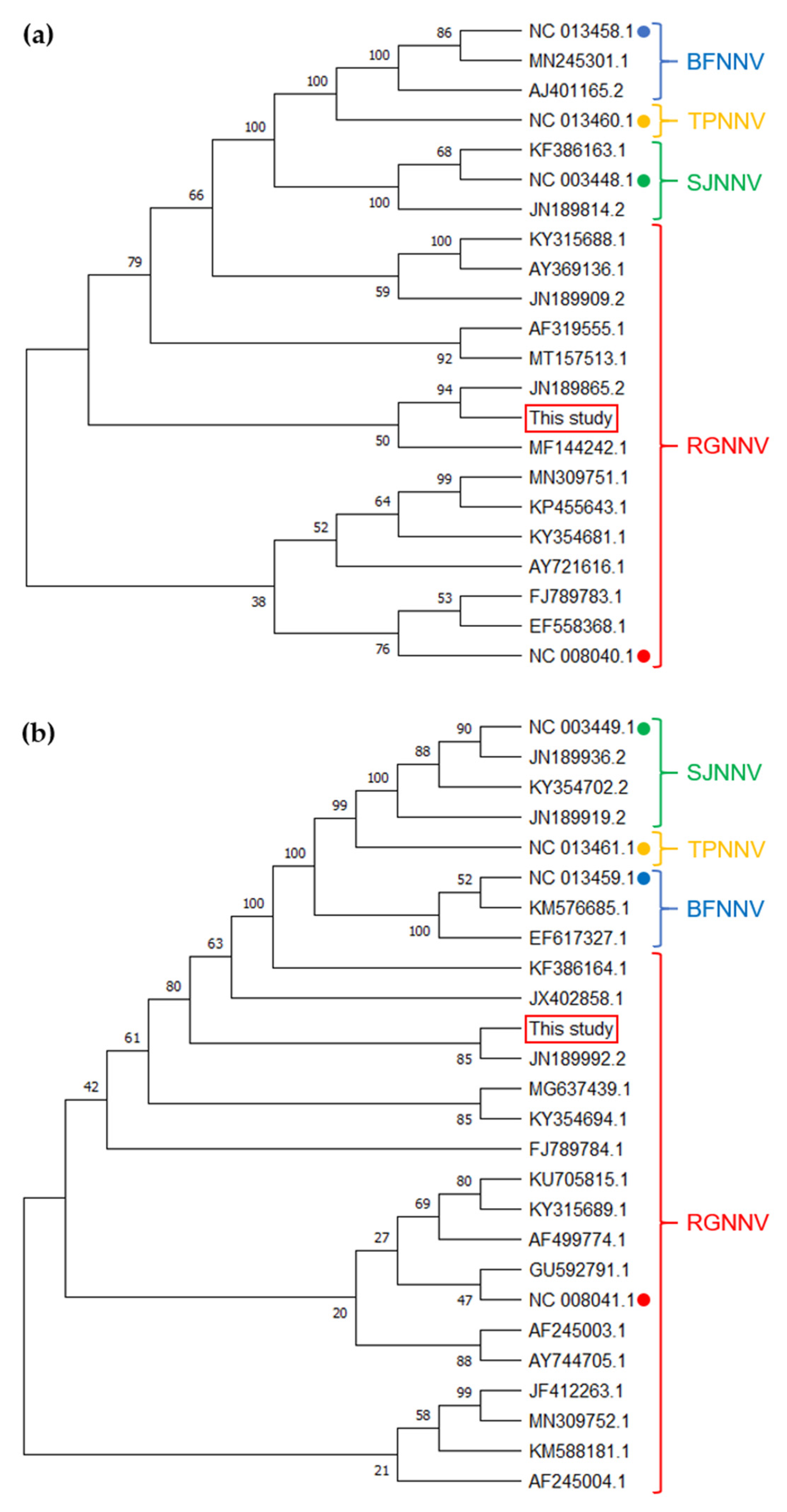
2.6. Phylogenetic Analysis
2.7. NNV Inactivation
2.8. qPCR
2.9. Statistical Analysis and Graphics
3. Results and Discussion
3.1. NNV Isolation and Identification
3.2. NNV Physical Inactivations
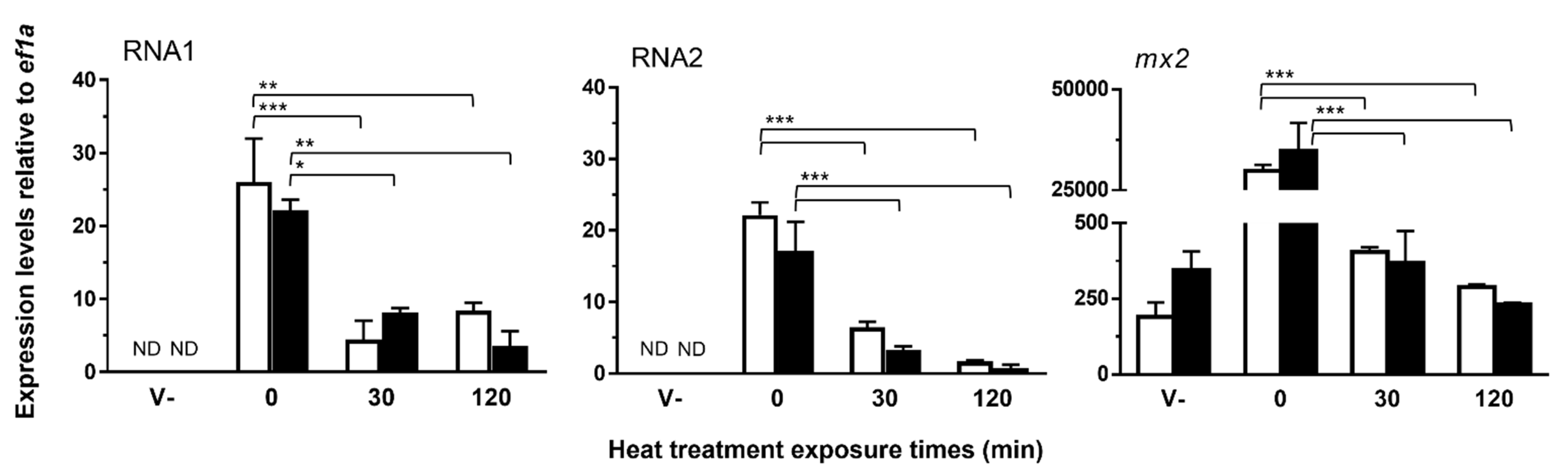
3.2.1. Heat Treatment
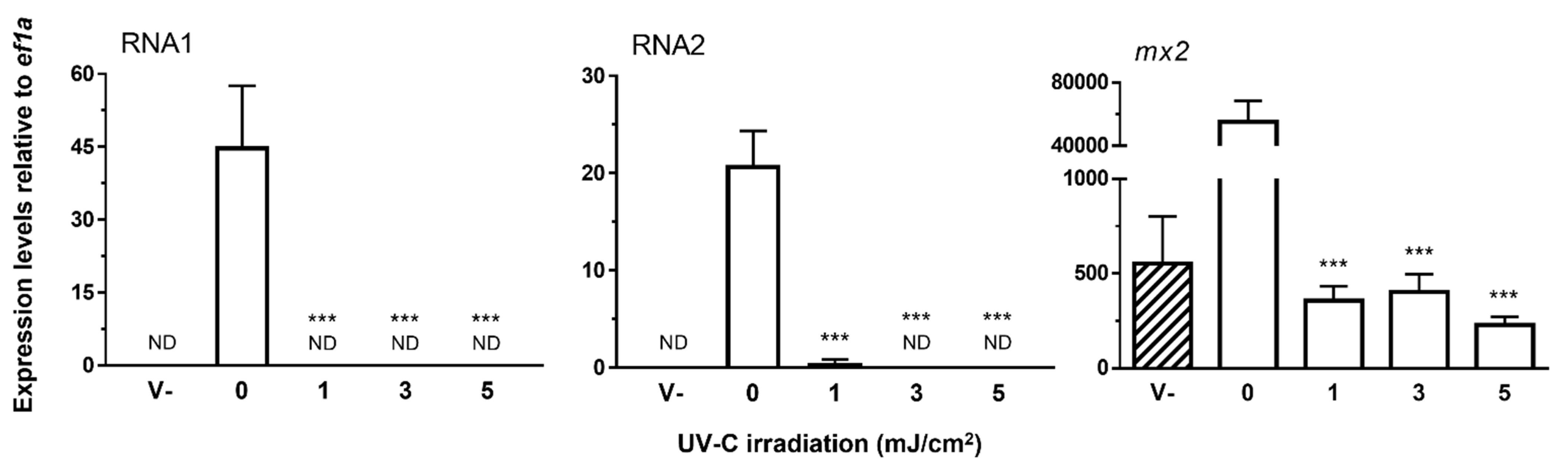
3.2.2. UV-C Irradiation
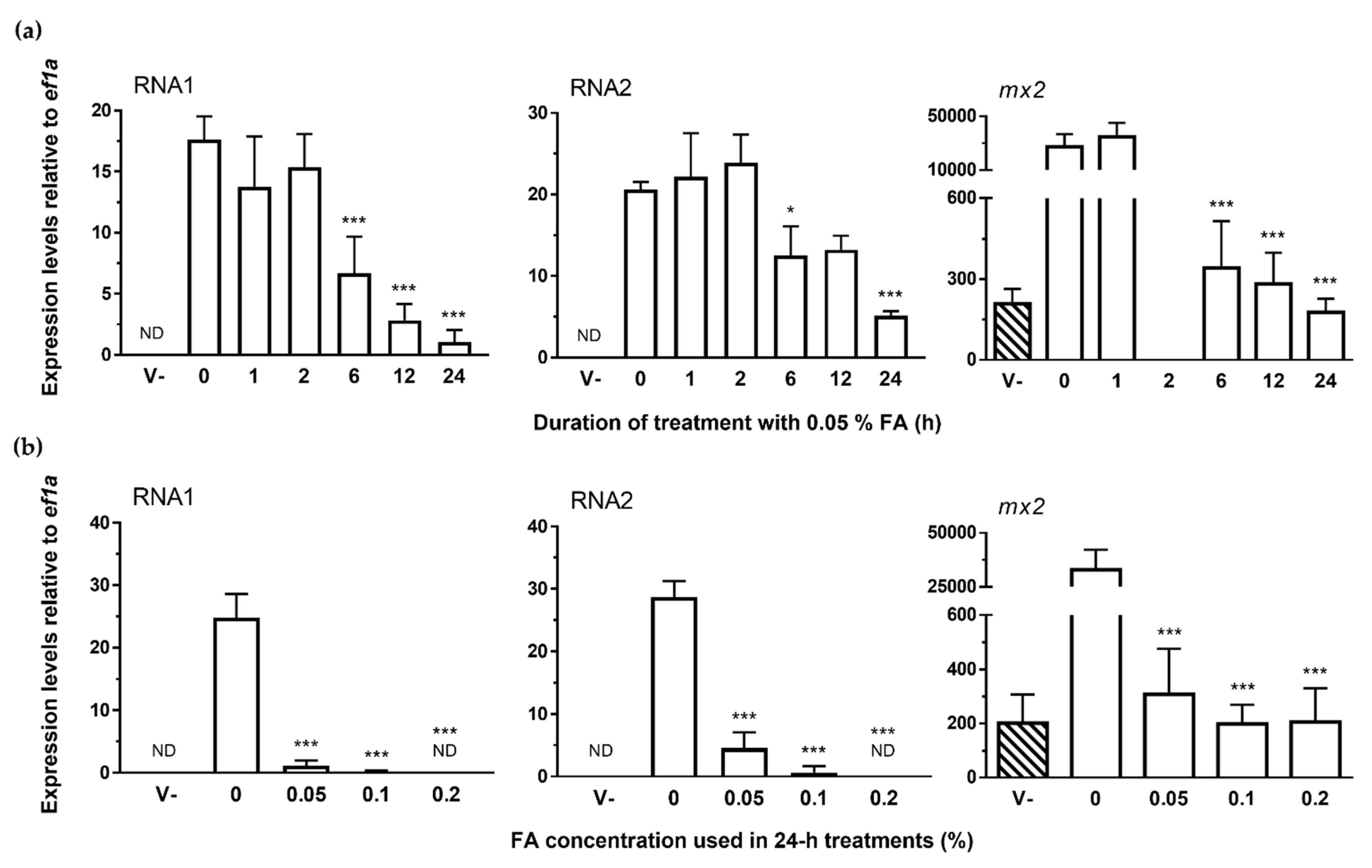
3.3. NNV Chemical Inactivation: Formalin Treatment
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Clercq, E.; Li, G. Approved antiviral drugs over the past 50 years. Clin. Microbiol. Rev. 2016, 29, 695–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clem, A.S. Fundamentals of vaccine immunology. J. Glob. Infect. Dis. 2011, 3, 73. [Google Scholar] [CrossRef]
- Salgado-Miranda, C.; Loza-Rubio, E.; Rojas-Anaya, E.; García-Espinosa, G. Viral vaccines for bony fish: Past, present and future. Expert Rev. Vaccines 2013, 12, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Dhar, A.K.; Manna, S.K.; Allnutt, F.T. Viral vaccines for farmed finfish. Virusdisease 2014, 25, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Chambers, M.A.; Graham, S.P.; La Ragione, R.M. Challenges in veterinary vaccine development and immunization. Vaccine Des. 2016, 3–35. [Google Scholar]
- Ma, J.; Bruce, T.J.; Jones, E.M.; Cain, K.D. A review of fish vaccine development strategies: Conventional methods and modern biotechnological approaches. Microorganisms 2019, 7, 569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ike, A.C.; Ononugbo, C.M.; Obi, O.J.; Onu, C.J.; Olovo, C.V.; Muo, S.O.; Chukwu, O.S.; Reward, E.E.; Omeke, O.P. Towards Improved Use of Vaccination in the Control of Infectious Bronchitis and Newcastle Disease in Poultry: Understanding the Immunological Mechanisms. Vaccines 2021, 9, 20. [Google Scholar] [CrossRef] [PubMed]
- Sanders, B.; Koldijk, M.; Schuitemaker, H. Inactivated viral vaccines. In Vaccine Analysis: Strategies, Principles, and Control; Springer: Berlin/Heidelberg, Germany, 2015; pp. 45–80. [Google Scholar]
- Kai, Y.-H.; Chi, S.-C. Efficacies of inactivated vaccines against betanodavirus in grouper larvae (Epinephelus coioides) by bath immunization. Vaccine 2008, 26, 1450–1457. [Google Scholar] [CrossRef]
- Rivas-Aravena, A.; Fuentes, Y.; Cartagena, J.; Brito, T.; Poggio, V.; La Torre, J.; Mendoza, H.; Gonzalez-Nilo, F.; Sandino, A.M.; Spencer, E. Development of a nanoparticle-based oral vaccine for Atlantic salmon against ISAV using an alphavirus replicon as adjuvant. Fish Shellfish Immunol. 2015, 45, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Wang, Y.; Hu, H.; Wang, Q.; Bergmann, S.M.; Wang, Y.; Li, B.; Lv, Y.; Li, H.; Yin, J. Cell Culture-Derived Tilapia Lake Virus-Inactivated Vaccine Containing Montanide Adjuvant Provides High Protection against Viral Challenge for Tilapia. Vaccines 2021, 9, 86. [Google Scholar] [CrossRef]
- Zhang, L.; Ma, J.; Fan, Y.; Zhou, Y.; Xu, J.; Liu, W.; Gu, Z.; Zeng, L. Immune response and protection in gibel carp, Carassius gibelio, after vaccination with β-propiolactone inactivated cyprinid herpesvirus 2. Fish Shellfish Immunol. 2016, 49, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Doriga, M.; Smail, D.; Smith, R.; Domenech, A.; Castric, J.; Smith, P.; Ellis, A. Isolation of salmon pancreas disease virus (SPDV) in cell culture and its ability to protect against infection by the ‘wild-type’agent. Fish Shellfish Immunol. 2001, 11, 505–522. [Google Scholar] [CrossRef]
- Hwang, J.Y.; Kwon, M.G.; Jung, S.-H.; Park, M.A.; Kim, D.-W.; Cho, W.S.; Park, J.W.; Son, M.-H. RNA-Seq transcriptome analysis of the olive flounder (Paralichthys olivaceus) kidney response to vaccination with heat-inactivated viral hemorrhagic septicemia virus. Fish Shellfish Immunol. 2017, 62, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.Y.; Kwon, M.-G.; Kim, Y.J.; Jung, S.-H.; Park, M.-A.; Son, M.-H. Montanide IMS 1312 VG adjuvant enhances the efficacy of immersion vaccine of inactivated viral hemorrhagic septicemia virus (VHSV) in olive flounder, Paralichthys olivaceus. Fish Shellfish Immunol. 2017, 60, 420–425. [Google Scholar] [CrossRef]
- Valero, Y.; Mokrani, D.; Chaves-Pozo, E.; Arizcun, M.; Oumouna, M.; Meseguer, J.; Esteban, M.Á.; Cuesta, A. Vaccination with UV-inactivated nodavirus partly protects European sea bass against infection, while inducing few changes in immunity. Dev. Comp. Immunol. 2018, 86, 171–179. [Google Scholar] [CrossRef]
- Brudeseth, B.E.; Wiulsrød, R.; Fredriksen, B.N.; Lindmo, K.; Løkling, K.-E.; Bordevik, M.; Steine, N.; Klevan, A.; Gravningen, K. Status and future perspectives of vaccines for industrialised fin-fish farming. Fish Shellfish Immunol. 2013, 35, 1759–1768. [Google Scholar] [CrossRef] [PubMed]
- Schulz, P.; Terech-Majewska, E.; Siwicki, A.K.; Kazuń, B.; Demska-Zakęś, K.; Rożyński, M.; Zakęś, Z. Effect of Different Routes of Vaccination against Aeromonas salmonicida on Rearing Indicators and Survival after an Experimental Challenge of Pikeperch (Sander lucioperca) in Controlled Rearing. Vaccines 2020, 8, 476. [Google Scholar] [CrossRef]
- Costa, J.Z.; Thompson, K.D. Understanding the interaction between Betanodavirus and its host for the development of prophylactic measures for viral encephalopathy and retinopathy. Fish Shellfish Immunol. 2016, 53, 35–49. [Google Scholar] [CrossRef] [Green Version]
- Yong, C.Y.; Yeap, S.K.; Omar, A.R.; Tan, W.S. Advances in the study of nodavirus. PeerJ 2017, 5, e3841. [Google Scholar] [CrossRef]
- Iwamoto, T.; Mise, K.; Takeda, A.; Okinaka, Y.; Mori, K.-I.; Arimoto, M.; Okuno, T.; Nakai, T. Characterization of Striped jack nervous necrosis virus subgenomic RNA3 and biological activities of its encoded protein B2. J. Gen. Virol. 2005, 86, 2807–2816. [Google Scholar] [CrossRef]
- Fenner, B.J.; Goh, W.; Kwang, J. Sequestration and protection of double-stranded RNA by the betanodavirus B2 protein. J. Virol. 2006, 80, 6822–6833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, N.-C.; Yoshimura, M.; Guan, H.-H.; Wang, T.-Y.; Misumi, Y.; Lin, C.-C.; Chuankhayan, P.; Nakagawa, A.; Chan, S.I.; Tsukihara, T. Crystal structures of a piscine betanodavirus: Mechanisms of capsid assembly and viral infection. PLoS Pathog. 2015, 11, e1005203. [Google Scholar] [CrossRef] [Green Version]
- NaveenKumar, S.; Shekar, M.; Karunasagar, I.; Karunasagar, I. Genetic analysis of RNA1 and RNA2 of Macrobrachium rosenbergii nodavirus (MrNV) isolated from India. Virus Res. 2013, 173, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Yoshikoshi, K.; Inoue, K. Viral nervous necrosis in hatchery-reared larvae and juveniles of Japanese parrotfish, Oplegnathus fasciatus (Temminck & Schlegel). J. Fish Dis. 1990, 13, 69–77. [Google Scholar]
- Munday, B.; Kwang, J.; Moody, N. Betanodavirus infections of teleost fish: A review. J. Fish Dis. 2002, 25, 127–142. [Google Scholar] [CrossRef]
- Shetty, M.; Maiti, B.; Santhosh, K.S.; Venugopal, M.N.; Karunasagar, I. Betanodavirus of marine and freshwater fish: Distribution, genomic organization, diagnosis and control measures. Indian J. Virol. 2012, 23, 114–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arimoto, M.; Mushiake, K.; Mizuta, Y.; Nakai, T.; Muroge, K.; Furusawa, I. Detection of striped jack nervous necrosis virus (SJNNV) by enzyme-linked immunosorbent assay (ELISA). Fish Pathol. 1992, 27, 191–195. [Google Scholar] [CrossRef] [Green Version]
- Grotmol, S.; Totland, G.K. Surface disinfection of Atlantic halibut Hippoglossus hippoglossus eggs with ozonated sea-water inactivates nodavirus and increases survival of the larvae. Dis. Aquat. Org. 2000, 39, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Skliris, G.P.; Krondiris, J.V.; Sideris, D.C.; Shinn, A.P.; Starkey, W.G.; Richards, R.H. Phylogenetic and antigenic characterization of new fish nodavirus isolates from Europe and Asia. Virus Res. 2001, 75, 59–67. [Google Scholar] [CrossRef]
- Fukuda, Y.; Furuhashi, M.; Nakai, T. Mass mortality of cultured sevenband grouper, Epinephelus septemfasciatus, associated with viral nervous necrosis. Fish Pathol. 1996, 31, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Munday, B.; Nakai, T. Nodaviruses as pathogens in larval and juvenile marine finfish. World J. Microbiol. Biotechnol. 1997, 13, 375–381. [Google Scholar] [CrossRef]
- Le Breton, A.; Grisez, L.; Sweetman, J.; Ollevier, F. Viral nervous necrosis (VNN) associated with mass mortalities in cage-reared sea bass, Dicentrarchus labrax (L.). J. Fish Dis. 1997, 20, 145–151. [Google Scholar] [CrossRef]
- Glazebrook, J.; Campbell, R. Diseases of barramundi (Lates calcarifer) in Australia: A review. In Management of Wild and Cultured Sea Bass/Barramundi; Ruskin Press: Melbourne, Australia, 1987; pp. 204–206. [Google Scholar]
- Panzarin, V.; Fusaro, A.; Monne, I.; Cappellozza, E.; Patarnello, P.; Bovo, G.; Capua, I.; Holmes, E.C.; Cattoli, G. Molecular epidemiology and evolutionary dynamics of betanodavirus in southern Europe. Infect. Genet. Evol. 2012, 12, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.S.; Chen, C.W.; Lin, C.H.; Tzeng, C.S.; Chang, C.Y. Differential expression profiling of orange-spotted grouper larvae, Epinephelus coioides (Hamilton), that survived a betanodavirus outbreak. J. Fish Dis. 2012, 35, 215–225. [Google Scholar] [CrossRef]
- Tanaka, S.; Mori, K.; Arimoto, M.; Iwamoto, T.; Nakai, T. Protective immunity of sevenband grouper, Epinephelus septemfasciatus Thunberg, against experimental viral nervous necrosis. J. Fish Dis. 2001, 24, 15–22. [Google Scholar] [CrossRef]
- Nishizawa, T.; Mori, K.-i.; Furuhashi, M.; Nakai, T.; Furusawa, I.; Muroga, K. Comparison of the coat protein genes of five fish nodaviruses, the causative agents of viral nervous necrosis in marine fish. J. Gen. Virol. 1995, 76, 1563–1569. [Google Scholar] [CrossRef]
- Nishizawa, T.; Furuhashi, M.; Nagai, T.; Nakai, T.; Muroga, K. Genomic classification of fish nodaviruses by molecular phylogenetic analysis of the coat protein gene. Appl. Environ. Microbiol. 1997, 63, 1633–1636. [Google Scholar] [CrossRef] [Green Version]
- Toffolo, V.; Negrisolo, E.; Maltese, C.; Bovo, G.; Belvedere, P.; Colombo, L.; Dalla Valle, L. Phylogeny of betanodaviruses and molecular evolution of their RNA polymerase and coat proteins. Mol. Phylogenetics Evol. 2007, 43, 298–308. [Google Scholar] [CrossRef]
- Olveira, J.G.; Souto, S.; Dopazo, C.P.; Thiéry, R.; Barja, J.L.; Bandín, I. Comparative analysis of both genomic segments of betanodaviruses isolated from epizootic outbreaks in farmed fish species provides evidence for genetic reassortment. J. Gen. Virol. 2009, 90, 2940–2951. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.-i.; Mangyoku, T.; Iwamoto, T.; Arimoto, M.; Tanaka, S.; Nakai, T. Serological relationships among genotypic variants of betanodavirus. Dis. Aquat. Org. 2003, 57, 19–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, J.Z. B Cell Epitopes in Fish Nodavirus. 2005. Available online: https://dspace.stir.ac.uk/bitstream/1893/13240/1/COSTA%202005.pdf (accessed on 17 July 2021).
- Yamashita, H.; Fujita, Y.; Kawakami, H.; Nakai, T. The efficacy of inactivated virus vaccine against viral nervous necrosis (VNN). Fish Pathol. 2005, 40, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Pakingking, R., Jr.; Seron, R.; Dela Peña, L.; Mori, K.; Yamashita, H.; Nakai, T. Immune responses of Asian sea bass, Lates calcarifer Bloch, against an inactivated betanodavirus vaccine. J. Fish Dis. 2009, 32, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Pakingking, R., Jr.; Bautista, N.B.; de Jesus-Ayson, E.G.; Reyes, O. Protective immunity against viral nervous necrosis (VNN) in brown-marbled grouper (Epinephelus fuscogutattus) following vaccination with inactivated betanodavirus. Fish Shellfish Immunol. 2010, 28, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Bejar, J.; Borrego, J.J.; Alvarez, M.C. A continuous cell line from the cultured marine fish gilt-head seabream (Sparus aurata L.). Aquaculture 1997, 150, 143–153. [Google Scholar] [CrossRef]
- Iwamoto, T.; Nakai, T.; Mori, K.-i.; Arimoto, M.; Furusawa, I. Cloning of the fish cell line SSN-1 for piscine nodaviruses. Dis. Aquat. Org. 2000, 43, 81–89. [Google Scholar] [CrossRef]
- Frerichs, G.; Morgan, D.; Hart, D.; Skerrow, C.; Roberts, R.; Onions, D. Spontaneously productive C-type retrovirus infection of fish cell lines. J. Gen. Virol. 1991, 72, 2537–2539. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Cattoli, G. Viral encephalopathy and retinopathy. In Manual of Diagnostic Tests for Aquatic Animals; OIE: Paris, France, 2019. [Google Scholar]
- Falco, A.; Chico, V.; Marroqui, L.; Perez, L.; Coll, J.; Estepa, A. Expression and antiviral activity of a β-defensin-like peptide identified in the rainbow trout (Oncorhynchus mykiss) EST sequences. Mol. Immunol. 2008, 45, 757–765. [Google Scholar] [CrossRef]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.; Potter, S.C.; Finn, R.D. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Frerichs, G.; Tweedie, A.; Starkey, W.; Richards, R. Temperature, pH and electrolyte sensitivity, and heat, UV and disinfectant inactivation of sea bass (Dicentrarchus labrax) neuropathy nodavirus. Aquaculture 2000, 185, 13–24. [Google Scholar] [CrossRef]
- Bello-Perez, M.; Pereiro, P.; Coll, J.; Novoa, B.; Perez, L.; Falco, A. Zebrafish C-reactive protein isoforms inhibit SVCV replication by blocking autophagy through interactions with cell membrane cholesterol. Sci. Rep. 2020, 10, 566. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Chaves-Pozo, E.; Guardiola, F.A.; Meseguer, J.; Esteban, M.A.; Cuesta, A. Nodavirus infection induces a great innate cell-mediated cytotoxic activity in resistant, gilthead seabream, and susceptible, European sea bass, teleost fish. Fish Shellfish Immunol. 2012, 33, 1159–1166. [Google Scholar] [CrossRef]
- Leiva-Rebollo, R.; Labella, A.; Borrego, J.; Castro, D. Immune gene expression in gilthead seabream (Sparus aurata) after Lymphocystis disease virus (LCDV-Sa) challenge resulting in asymptomatic infection. J. Appl. Microbiol. 2020, 128, 41–53. [Google Scholar] [CrossRef] [Green Version]
- Cerezuela, R.; Meseguer, J.; Esteban, M.Á. Effects of dietary inulin, Bacillus subtilis and microalgae on intestinal gene expression in gilthead seabream (Sparus aurata L.). Fish Shellfish Immunol. 2013, 34, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Vendramin, N.; Toffan, A.; Mancin, M.; Cappellozza, E.; Panzarin, V.; Bovo, G.; Cattoli, G.; Capua, I.; Terregino, C. Comparative pathogenicity study of ten different betanodavirus strains in experimentally infected E uropean sea bass, Dicentrarchus labrax (L.). J. Fish Dis. 2014, 37, 371–383. [Google Scholar] [CrossRef]
- Toffan, A.; Panzarin, V.; Toson, M.; Cecchettin, K.; Pascoli, F. Water temperature affects pathogenicity of different betanodavirus genotypes in experimentally challenged Dicentrarchus labrax. Dis. Aquat. Org. 2016, 119, 231–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panzarin, V.; Cappellozza, E.; Mancin, M.; Milani, A.; Toffan, A.; Terregino, C.; Cattoli, G. In vitro study of the replication capacity of the RGNNV and the SJNNV betanodavirus genotypes and their natural reassortants in response to temperature. Vet. Res. 2014, 45, 56. [Google Scholar] [CrossRef] [Green Version]
- Barsøe, S.; Allal, F.; Vergnet, A.; Vandeputte, M.; Olesen, N.J.; Schmidt, J.G.; Larsen, C.A.; Cuenca, A.; Vendramin, N. Different survival of three populations of European sea bass (Dicentrarchus labrax) following challenge with two variants of nervous necrosis virus (NNV). Aquac. Rep. 2021, 19, 100621. [Google Scholar] [CrossRef]
- Dixon, P.; Smail, D.; Algoët, M.; Hastings, T.; Bayley, A.; Byrne, H.; Dodge, M.; Garden, A.; Joiner, C.; Roberts, E. Studies on the effect of temperature and pH on the inactivation of fish viral and bacterial pathogens. J. Fish Dis. 2012, 35, 51–64. [Google Scholar] [CrossRef]
- Dalla Valle, L.; Toffolo, V.; Lamprecht, M.; Maltese, C.; Bovo, G.; Belvedere, P.; Colombo, L. Development of a sensitive and quantitative diagnostic assay for fish nervous necrosis virus based on two-target real-time PCR. Vet. Microbiol. 2005, 110, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Panzarin, V.; Patarnello, P.; Mori, A.; Rampazzo, E.; Cappellozza, E.; Bovo, G.; Cattoli, G. Development and validation of a real-time TaqMan PCR assay for the detection of betanodavirus in clinical specimens. Arch. Virol. 2010, 155, 1193–1203. [Google Scholar] [CrossRef] [PubMed]
- Delrue, I.; Verzele, D.; Madder, A.; Nauwynck, H.J. Inactivated virus vaccines from chemistry to prophylaxis: Merits, risks and challenges. Expert Rev. Vaccines 2012, 11, 695–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gye, H.J.; Park, M.-J.; Kim, W.-S.; Oh, M.-J.; Nishizawa, T. Heat-denaturation of conformational structures on nervous necrosis virus for generating neutralization antibodies. Aquaculture 2018, 484, 65–70. [Google Scholar] [CrossRef]
- Yamashita, H.; Mori, K.; Kuroda, A.; Nakai, T. Neutralizing antibody levels for protection against betanodavirus infection in sevenband grouper, Epinephelus septemfasciatus (Thunberg), immunized with an inactivated virus vaccine. J. Fish Dis. 2009, 32, 767–775. [Google Scholar] [CrossRef]
- Nuñez-Ortiz, N.; Pascoli, F.; Picchietti, S.; Buonocore, F.; Bernini, C.; Toson, M.; Scapigliati, G.; Toffan, A. A formalin-inactivated immunogen against viral encephalopathy and retinopathy (VER) disease in European sea bass (Dicentrarchus labrax): Immunological and protection effects. Vet. Res. 2016, 47, 89. [Google Scholar] [CrossRef] [Green Version]
- Valero, Y.; Olveira, J.G.; López-Vázquez, C.; Dopazo, C.P.; Bandín, I. BEI Inactivated Vaccine Induces Innate and Adaptive Responses and Elicits Partial Protection upon Reassortant Betanodavirus Infection in Senegalese Sole. Vaccines 2021, 9, 458. [Google Scholar] [CrossRef]
- Tso, C.-H.; Lu, M.-W. Transcriptome profiling analysis of grouper during nervous necrosis virus persistent infection. Fish Shellfish Immunol. 2018, 76, 224–232. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Species | GB ID | Sequence (5′-3′) | Length | Ref. |
|---|---|---|---|---|---|
| rdrp | NNV | - | F: CCGATATCACGATGAGTTCAC R: GAAAGCGTAGGTGTAGCTGG | 139 | This study |
| cp | NNV | - | F: CGCTGCTAGAATCTTCCA R: CTTTCCCACCATTTGGC | 183 | This study |
| mx2 | Sea bream | FJ490555 | F: AAGAGGAGGACGAGGAGGAG R: TTCAGGTGCAGCATCAACTC | 148 | [58,59] |
| ef1a | Sea bream | AF184170 | F: CTGTCAAGGAAATCCGTCGT R: TGACCTGAGCGTTGAAGTTG | 87 | [60] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Falco, A.; Bello-Perez, M.; Díaz-Puertas, R.; Mold, M.; Adamek, M. Update on the Inactivation Procedures for the Vaccine Development Prospects of a New Highly Virulent RGNNV Isolate. Vaccines 2021, 9, 1441. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9121441
Falco A, Bello-Perez M, Díaz-Puertas R, Mold M, Adamek M. Update on the Inactivation Procedures for the Vaccine Development Prospects of a New Highly Virulent RGNNV Isolate. Vaccines. 2021; 9(12):1441. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9121441
Chicago/Turabian StyleFalco, Alberto, Melissa Bello-Perez, Rocío Díaz-Puertas, Matthew Mold, and Mikolaj Adamek. 2021. "Update on the Inactivation Procedures for the Vaccine Development Prospects of a New Highly Virulent RGNNV Isolate" Vaccines 9, no. 12: 1441. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9121441