
Bacterial-Based Cancer Therapy (BBCT): Recent Advances, Current Challenges, and Future Prospects for Cancer Immunotherapy

 ,
,
Abstract
:1. Introduction
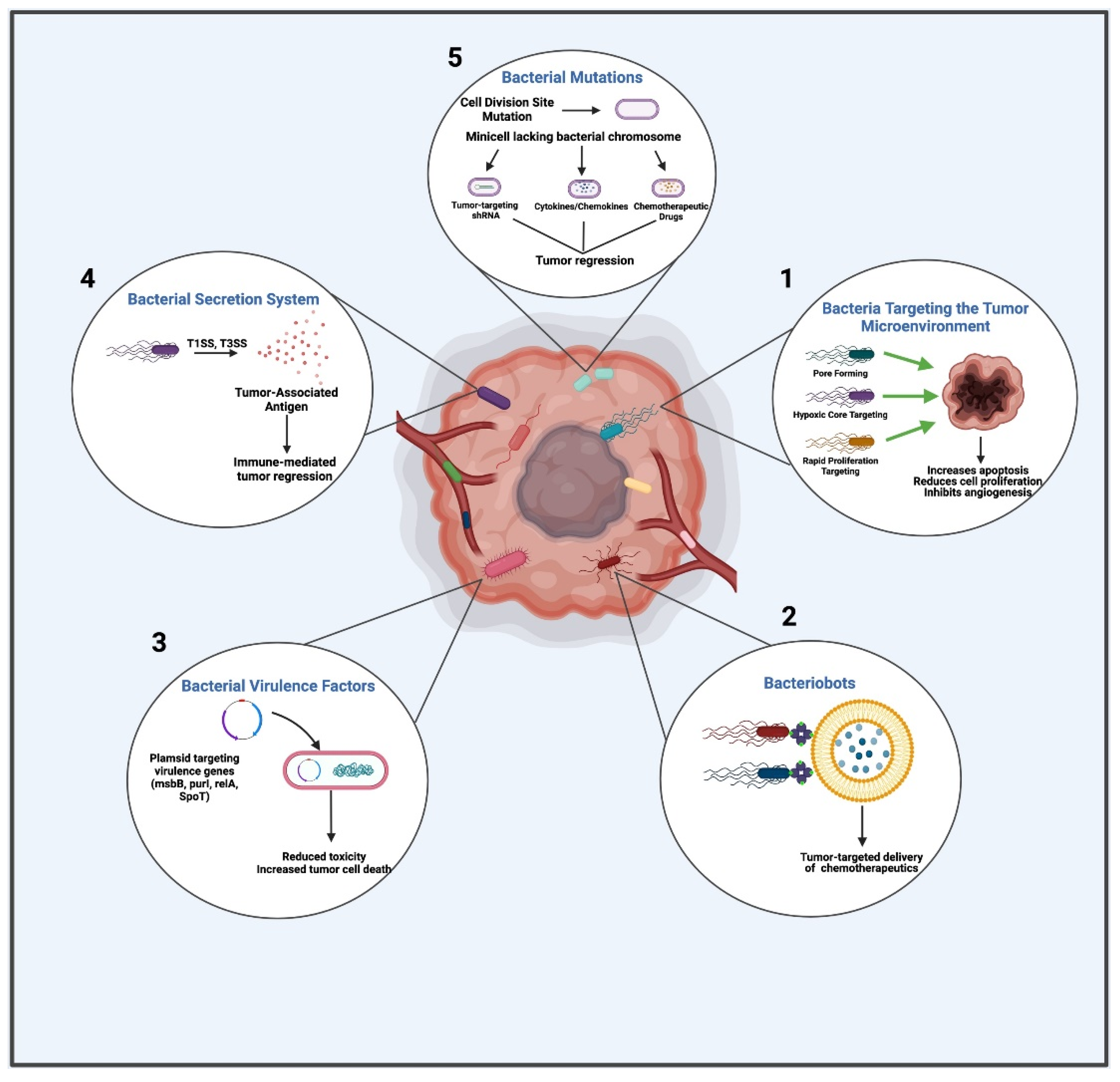
2. Bacterial Components and Products Targeted via BBCT
2.1. Bacterial Targeting of the Tumor Microenvironment
2.2. Bacteriobots
2.3. Bacterial Virulence Factors
2.4. The Bacterial Secretion System
2.5. Bacterial Mutations
3. Pathogenic and Non-Pathogenic Bacteria in BBCT
3.1. Pathogenic Bacteria in BBCT
3.1.1. Salmonella spp.
3.1.2. Escherichia spp.
3.1.3. Listeria spp.
3.1.4. Clostridium spp.
3.1.5. Corynebacterium spp.
3.1.6. Pseudomonas spp.
3.2. Non-Pathogenic Bacteria/Probiotics in BBCT
3.2.1. Lactic Acid Bacteria (LAB)
3.2.2. Lactobacillus spp.
3.2.3. Lactococcus spp.
3.2.4. Bifidobacterium spp.
3.2.5. Magnetococcus spp.

{kind=link}
| Bacteria | Strain | Mutated/Gene Modified | Cancer Type | Phenotypic Description | Ref |
|---|---|---|---|---|---|
| Pathogenic bacteria-mediated cancer immunotherapy | |||||
| Salmonella typhimurium | A1-R | ∆leu/∆arg | Prostate cancer | Auxotrophic strain defective in synthesis of leucine and arginine | [145] |
| VNP20009 | ∆msbB/∆purI | Metastatic melanoma, Glioblastoma, Pancreatic cancer, Colon cancer, Breast cancer | Modification of Lipid A structure; reduced ability to induce TNF-α secretion; deficiency in adenine synthesis | [146,147,148] | |
| SHJ2037 | ∆relA/∆spoT | ∆ppGpp (global regulator); reduction in bacterial invasion | [149,150] | ||
| SL3261, SL7207, BRD509, YB1 | aro- | Prostate cancer, Melanoma, Breast cancer | Mutations in aromatic amino acid biosynthesis | [19,151,152,153,154] | |
| LH430; VNP (Pho/Q-) | ∆phoP/∆phoQ | Colorectal cancer, Renal cancer | Reduced bacterial survival in macrophages | [37,155] | |
| MvP728 | ∆purD/∆htrA | Colon carcinoma, DBT glioblastoma, Melanoma | Defective in purine biosynthesis, produces heat-shock protein response to stress stimuli | [156] | |
| YB1; ST8 | ∆asd | Breast cancer, Colon cancer | Defective in diaminopimelic acid (DAP) synthesis | [19,157] | |
| X4550 | ∆cya/∆crp | Osteosarcoma | Disabled production of cAMP (cyclic adenosine monophosphate) synthetase and cAMP receptor protein | [158] | |
| RE88 | ∆dam | Breast carcinoma | Defective in DNA adenine methylase production | [159] | |
| SB824 | ∆sptP | Melanoma | Defective in pathogenicity island 1 (SPI-1) | [160] | |
| ST8 | ∆gmd | Colon cancer | Unable to replicate beyond the anaerobic regions of tumors | [137] | |
| SF100; SF200; S364 | rfa-, ΔpagP/ΔpagL/ΔlpxR | Colorectal cancer, Fibrosarcoma | Highly truncated LPS and attenuated bacterial virulence | [161,162] | |
| MPO378 | ∆purD/∆upp | Breast Cancer cell line | Inefficient in purine biosynthesis and uracil phosphoribosyl transferase | [162] | |
| FlaB | Vibrio vulnificusflagellin B | Colon cancer | Engineered FlaB from Vibrio vulnificus-secreting bacteria | [150] | |
| Listeria monocytogenes | rLM | Lm-LLO-E7 | Cervical cancer, Leukemia, Ovarian cancer, Prostate cancer, Colon cancer, Breast cancer | Secretes a fusion protein comprised of nonfunctional LLO joined with HPV protein E7 | [163] |
| XFL7 | Lm-LLO-PSA | Prostate cancer | Significantly higher number of IFN-γ-secreting cells | [164] | |
| DP-L4029 | ∆actA | Colon cancer, Lung cancer | Defective surface-bound ActA polypeptide, constitutes LLO activity at physiologic pH | [44,165,166] | |
| DP-L4017 | LLO L46IT, LLOD26 | Lung cancer | Cytotoxic, defective cell-to-cell spreading and greater percentages of splenic- and tumor-infiltrating, antigen-specific CD8+ lymphocytes | [5,42,167] | |
| DP-L4042 | ∆PEST | Colon cancer, Lung cancer | Cytotoxic, defective cell-to-cell spreading | [42,167] | |
| DP-L4405 DP-L4406 | ∆inIA/∆niB | Colon cancer | Impaired InIA-mediated infection | [168] | |
| CS-L0001 | ∆actA/∆inlB | Colon tumor lung metastases | Defective in cell-to-cell spreading | [44] | |
| CS-L0002 | ∆actA/∆lplA | L. monocytogenes vaccine vectors expressing influenza A nucleoprotein | [169] | ||
| DP-L4038 | ∆actA/L461T LLO | Inadequate surface-bound ActA polypeptide, constitutes LLO activity at physiologic pH | [165,166] | ||
| Mycobacterium bovis | BCG Pasteur | 1137P2 | Bladder cancer | Cancer cell phagocytosis by increasing proinflammatory cytokine activation and immune system | [47,170,171] |
| Clostridium novyi | NT | ∆toxA/∆toxB | Glioblastomas neuroshphere, Colon cancer | Produces specific enzymes and toxins capable of destroying cancer cells | [172,173,174] |
| Escherichia coli | MG1655 | 4T1 breast cancer | Optimized physicochemical properties for bacterial attachment; Low cost for bioconjugation | [175] | |
| Streptococcus pyogenes | OK-432 | Lymphangioma intraoral ranula | Including TNF, IL-8, IL-6, IFN-γ, and VEGF; increase in WBCs | [176,177,178,179] | |
| Pseudomonas aeruginosa | F10 | Lung cancer, Breast cancer, Cervical cancer, and Colon cancer | Anti-tumor effects of 2,4-diacetylphloroglucinol (DAPG) extracted | [180] | |
| (PA-MSHA) | Pancreatic cancer | Anti-tumor effect of P. aeruginosa-MSHA (mannose-sensitive hemagglutinin) inducing apoptosis by the EGFRa pathway and caspase signaling | [181] | ||
| Hepatocellular carcinoma | Anti-tumor effect of P. aeruginosa-MSHA (mannose-sensitive hemagglutinin) by EGFR/Akt/IκBβ/NF-κB pathway | [182] | |||
| Non-pathogenic bacteria-mediated cancer immunotherapy | |||||
| Lactobacillus reuteri | PTCC 1655 | WT | Gastric cancer | Probiotic-based strategies: inhibition of cell proliferation by downregulation of uPA/uPA receptors (uPARs) | [183] |
| FLRE5K1 | WT | Melanoma | Preventive effect of L. reuteri on melanoma | [129] | |
| Lactobacillus plantarum | WT | Colon cancer, Breast cancer, Oral cancer | Produces antioxidants, increases TNF-α, induces caspase-3 activity, inactivates Wnt/β-catenin signaling | [184,185] | |
| Lactobacillus rhamnosus | SHA111; SHA112; SHA113 | WT | Colorectal cancer, Cervical adenocarcinoma, Breast cancer | Apoptosis via up-regulation of BAD, BAX, Caspase3, Caspase8, and Caspase9, and down-regulation of BCL-2 genes | [183] |
| Lactococcus lactis | Head and neck tumor | Anti-tumor effect of nisin: by induction of apoptosis through a calpain-dependent pathway | [128] | ||
| Bifidobacterium bifidum | Lung cancer | Induction of immune responses, which leads to inhibition of tumor growth by activation of IL-12 and IFN-γ, lymphocyte proliferation, and CD8+ cytolytic induction | [186] | ||
| CGMCC 15068 | Colon cancer | B. bifidum growth in intestinal health by modulating dysbiosis and the gut metabolic profile | [187] | ||
| Bifidobacterium longum | NCC2705 | WT | Colon adenocarcinoma | B. longum as a vector of tumstatin (Tum) inducing significant anti-tumor effect | [137] |
| 420 and 440 | WT | Prostate cancer | B. longum-based vaccine inducing immune response against Wilms tumor 1 (WT1) antigen | [188] | |
| Bifidobacterium breve | UCC2003 | WT | Head and neck tumor | Strain expressing IL-24 gene: Apoptosis induction leads to anti-tumor activity | [136] |
4. Clinical Trials Using Bacteria as Delivery Vehicles
5. Current Challenges
6. Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coley, W.B. The treatment of malignant tumors by repeated inoculations of erysipelas. With a report of ten original cases. Clin. Orthop. Relat Res. 1893, 262, 487. [Google Scholar]
- McCarthy, E.F. The toxins of William B. Coley and the treatment of bone and soft-tissue sarcomas. Iowa Orthop. J. 2006, 26, 154–158. [Google Scholar] [PubMed]
- Lin, I.Y.; Van, T.T.; Smooker, P.M. Live-Attenuated Bacterial Vectors: Tools for Vaccine and Therapeutic Agent Delivery. Vaccines 2015, 3, 940–972. [Google Scholar] [CrossRef] [Green Version]
- Forbes, N.S. Engineering the perfect (bacterial) cancer therapy. Nat. Rev. Cancer 2010, 10, 785–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, L.M.; Guirnalda, P.D.; Seavey, M.M.; Paterson, Y. Cancer immunotherapy using Listeria monocytogenes and listerial virulence factors. Immunol. Res. 2008, 42, 233–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardlik, R.; Fruehauf, J.H. Bacterial vectors and delivery systems in cancer therapy. IDrugs 2010, 13, 701–706. [Google Scholar] [PubMed]
- Łukasiewicz, K.; Fol, M. Microorganisms in the Treatment of Cancer: Advantages and Limitations. J. Immunol. Res. 2018, 2018, 2397808. [Google Scholar] [CrossRef] [Green Version]
- St Jean, A.T.; Swofford, C.A.; Panteli, J.T.; Brentzel, Z.J.; Forbes, N.S. Bacterial delivery of Staphylococcus aureus α-hemolysin causes regression and necrosis in murine tumors. Mol. Ther. 2014, 22, 1266–1274. [Google Scholar] [CrossRef] [PubMed]
- Ryan, R.M.; Green, J.; Williams, P.J.; Tazzyman, S.; Hunt, S.; Harmey, J.H.; Kehoe, S.C.; Lewis, C.E. Bacterial delivery of a novel cytolysin to hypoxic areas of solid tumors. Gene Ther. 2009, 16, 329–339. [Google Scholar] [CrossRef] [Green Version]
- Kalaora, S.; Nagler, A.; Nejman, D.; Alon, M.; Barbolin, C.; Barnea, E.; Ketelaars, S.L.C.; Cheng, K.; Vervier, K.; Shental, N.; et al. Identification of bacteria-derived HLA-bound peptides in melanoma. Nature 2021, 592, 138–143. [Google Scholar] [CrossRef]
- Antonelli, A.C.; Binyamin, A.; Hohl, T.M.; Glickman, M.S.; Redelman-Sidi, G. Bacterial immunotherapy for cancer induces CD4-dependent tumor-specific immunity through tumor-intrinsic interferon-γ signaling. Proc. Natl. Acad. Sci. USA 2020, 117, 18627–18637. [Google Scholar] [CrossRef]
- Leventhal, D.S.; Sokolovska, A.; Li, N.; Plescia, C.; Kolodziej, S.A.; Gallant, C.W.; Christmas, R.; Gao, J.-R.; James, M.J.; Abin-Fuentes, A.; et al. Immunotherapy with engineered bacteria by targeting the STING pathway for anti-tumor immunity. Nat. Commun. 2020, 11, 2739. [Google Scholar] [CrossRef]
- Miyazaki, M.; Yuba, E.; Harada, A.; Kono, K. Hyaluronic acid derivative-modified liposomes as pH-sensitive anticancer drug delivery system. J. Control. Release 2015, 213, e73-4. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wang, Y.; Qin, M.; Zhang, X.; Zhang, Z.; Sun, X.; Gu, Z. Bacteria-Driven Hypoxia Targeting for Combined Biotherapy and Photothermal Therapy. ACS Nano 2018, 12, 5995–6005. [Google Scholar] [CrossRef]
- Toso, J.F.; Gill, V.J.; Hwu, P.; Marincola, F.M.; Restifo, N.P.; Schwartzentruber, D.J.; Sherry, R.M.; Topalian, S.L.; Yang, J.C.; Stock, F.; et al. Phase I study of the intravenous administration of attenuated Salmonella typhimurium to patients with metastatic melanoma. J. Clin. Oncol. 2002, 20, 142–152. [Google Scholar] [CrossRef]
- Fritz, S.E.; Henson, M.S.; Greengard, E.; Winter, A.L.; Stuebner, K.M.; Yoon, U.; Wilk, V.L.; Borgatti, A.; Augustin, L.B.; Modiano, J.F.; et al. A phase I clinical study to evaluate safety of orally administered, genetically engineered Salmonella enterica serovar Typhimurium for canine osteosarcoma. Vet. Med. Sci. 2016, 2, 179–190. [Google Scholar] [CrossRef]
- Heimann, D.M.; Rosenberg, S.A. Continuous intravenous administration of live genetically modified salmonella typhimurium in patients with metastatic melanoma. J. Immunother. 2003, 26, 179–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemunaitis, J.; Cunningham, C.; Senzer, N.; Kuhn, J.; Cramm, J.; Litz, C.; Cavagnolo, R.; Cahill, A.; Clairmont, C.; Sznol, M. Pilot trial of genetically modified, attenuated Salmonella expressing the E. coli cytosine deaminase gene in refractory cancer patients. Cancer Gene Ther. 2003, 10, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Yang, M.; Shi, L.; Yao, Y.; Jiang, Q.; Li, X.; Tang, L.H.; Zheng, B.J.; Yuen, K.Y.; Smith, D.K.; et al. Explicit hypoxia targeting with tumor suppression by creating an "obligate" anaerobic Salmonella Typhimurium strain. Sci. Rep. 2012, 2, 436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, M.Q.; Ellem, K.A.; Dunn, P.; West, M.J.; Bai, C.X.; Vogelstein, B. Facultative or obligate anaerobic bacteria have the potential for multimodality therapy of solid tumours. Eur. J. Cancer 2007, 43, 490–496. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Outschoorn, U.E.; Peiris-Pagés, M.; Pestell, R.G.; Sotgia, F.; Lisanti, M.P. Cancer metabolism: A therapeutic perspective. Nat. Rev. Clin. Oncol. 2017, 14, 11–31. [Google Scholar] [CrossRef]
- Sedighi, M.; Zahedi Bialvaei, A.; Hamblin, M.R.; Ohadi, E.; Asadi, A.; Halajzadeh, M.; Lohrasbi, V.; Mohammadzadeh, N.; Amiriani, T.; Krutova, M.; et al. Therapeutic bacteria to combat cancer; current advances, challenges, and opportunities. Cancer Med. 2019, 8, 3167–3181. [Google Scholar] [CrossRef]
- Jain, R.K.; Forbes, N.S. Can engineered bacteria help control cancer? Proc. Natl. Acad. Sci. USA 2001, 98, 14748–14750. [Google Scholar] [CrossRef] [Green Version]
- Carlisle, R.; Coussios, C.C. Mechanical approaches to oncological drug delivery. Ther. Deliv. 2013, 4, 1213–1215. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.M.; Giaccia, A.J. The unique physiology of solid tumors: Opportunities (and problems) for cancer therapy. Cancer Res. 1998, 58, 1408–1416. [Google Scholar]
- Cheong, I.; Zhou, S. Tumor-Specific Liposomal Drug Release Mediated by Liposomase. Methods Enzymol. 2009, 465, 251–265. [Google Scholar] [PubMed]
- Nallar, S.C.; Xu, D.Q.; Kalvakolanu, D.V. Bacteria and genetically modified bacteria as cancer therapeutics: Current advances and challenges. Cytokine 2017, 89, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Yazawa, K.; Fujimori, M.; Amano, J.; Kano, Y.; Taniguchi, S. Bifidobacterium longum as a delivery system for cancer gene therapy: Selective localization and growth in hypoxic tumors. Cancer Gene Ther. 2000, 7, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarty, A.M. Microorganisms and cancer: Quest for a therapy. J. Bacteriol. 2003, 185, 2683–2686. [Google Scholar] [CrossRef] [Green Version]
- Song, S.; Vuai, M.S.; Zhong, M. The role of bacteria in cancer therapy – enemies in the past, but allies at present. Infect. Agents Cancer 2018, 13, 9. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Park, S.-H.; Cho, S.; Kim, D.-M.; Lee, Y.; Ko, S.Y.; Hong, Y.; Choy, H.E.; Min, J.-J.; Park, J.-O.; et al. New paradigm for tumor theranostic methodology using bacteria-based microrobot. Sci. Rep. 2013, 3, 3394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, V.D.; Han, J.-W.; Choi, Y.J.; Cho, S.; Zheng, S.; Ko, S.Y.; Park, J.-O.; Park, S. Active tumor-therapeutic liposomal bacteriobot combining a drug (paclitaxel)-encapsulated liposome with targeting bacteria (Salmonella Typhimurium). Sens. Actuators B Chem. 2016, 224, 217–224. [Google Scholar] [CrossRef]
- Park, D.; Park, S.J.; Cho, S.; Lee, Y.; Lee, Y.K.; Min, J.-J.; Park, B.J.; Ko, S.Y.; Park, J.-O.; Park, S. Motility analysis of bacteria-based microrobot (bacteriobot) using chemical gradient microchamber. Biotechnol. Bioeng. 2014, 111, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A.; Pirofski, L.-a. Virulence factors and their mechanisms of action: The view from a damage–response framework. J. Water Health 2009, 7, S2–S18. [Google Scholar] [CrossRef]
- Cross, A.S. What is a virulence factor? Crit. Care 2008, 12, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.H.; Lin, S.T.; Liu, J.J.; Chang, W.W.; Hsieh, J.L.; Wang, W.K. Salmonella induce autophagy in melanoma by the downregulation of AKT/mTOR pathway. Gene Ther. 2014, 21, 309–316. [Google Scholar] [CrossRef]
- Frahm, M.; Felgner, S.; Kocijancic, D.; Rohde, M.; Hensel, M.; Curtiss, R., 3rd; Erhardt, M.; Weiss, S. Efficiency of conditionally attenuated Salmonella enterica serovar Typhimurium in bacterium-mediated tumor therapy. mBio 2015, 6, e00254-15. [Google Scholar] [CrossRef] [Green Version]
- Na, H.S.; Kim, H.J.; Lee, H.C.; Hong, Y.; Rhee, J.H.; Choy, H.E. Immune response induced by Salmonella typhimurium defective in ppGpp synthesis. Vaccine 2006, 24, 2027–2034. [Google Scholar] [CrossRef] [PubMed]
- Glomski, I.J.; Gedde, M.M.; Tsang, A.W.; Swanson, J.A.; Portnoy, D.A. The Listeria monocytogenes hemolysin has an acidic pH optimum to compartmentalize activity and prevent damage to infected host cells. J. Cell Biol. 2002, 156, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Glomski, I.J.; Decatur, A.L.; Portnoy, D.A. Listeria monocytogenes mutants that fail to compartmentalize listerolysin O activity are cytotoxic, avirulent, and unable to evade host extracellular defenses. Infect. Immun. 2003, 71, 6754–6765. [Google Scholar] [CrossRef] [Green Version]
- Camilli, A.; Tilney, L.G.; Portnoy, D.A. Dual roles of plcA in Listeria monocytogenes pathogenesis. Mol. Microbiol. 1993, 8, 143–157. [Google Scholar] [CrossRef]
- Decatur, A.L.; Portnoy, D.A. A PEST-like sequence in listeriolysin O essential for Listeria monocytogenes pathogenicity. Science 2000, 290, 992–995. [Google Scholar] [CrossRef] [Green Version]
- Bakardjiev, A.I.; Stacy, B.A.; Fisher, S.J.; Portnoy, D.A. Listeriosis in the pregnant guinea pig: A model of vertical transmission. Infect. Immun. 2004, 72, 489–497. [Google Scholar] [CrossRef] [Green Version]
- Brockstedt, D.G.; Giedlin, M.A.; Leong, M.L.; Bahjat, K.S.; Gao, Y.; Luckett, W.; Liu, W.; Cook, D.N.; Portnoy, D.A.; Dubensky, T.W., Jr. Listeria-based cancer vaccines that segregate immunogenicity from toxicity. Proc. Natl. Acad. Sci. USA 2004, 101, 13832–13837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chagnon, A.; Hudon, C.; McSween, G.; Vinet, G.; Fredette, V. Cytotoxicity and reduction of animal cell growth by Clostridium M-55 spores and their extracts. Cancer 1972, 29, 431–434. [Google Scholar] [CrossRef]
- Cheong, I.; Huang, X.; Bettegowda, C.; Diaz, L.A., Jr.; Kinzler, K.W.; Zhou, S.; Vogelstein, B. A bacterial protein enhances the release and efficacy of liposomal cancer drugs. Science 2006, 314, 1308–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felgner, S.; Kocijancic, D.; Frahm, M.; Weiss, S. Bacteria in Cancer Therapy: Renaissance of an Old Concept. Int. J. Microbiol. 2016, 2016, 8451728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fronzes, R.; Christie, P.J.; Waksman, G. The structural biology of type IV secretion systems. Nat. Rev. Microbiol. 2009, 7, 703–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singer, H.M.; Erhardt, M.; Steiner, A.M.; Zhang, M.M.; Yoshikami, D.; Bulaj, G.; Olivera, B.M.; Hughes, K.T. Selective purification of recombinant neuroactive peptides using the flagellar type III secretion system. mBio 2012, 3, e00115-12. [Google Scholar] [CrossRef] [Green Version]
- Farley, M.M.; Hu, B.; Margolin, W.; Liu, J. Minicells, Back in Fashion. J. Bacteriol. 2016, 198, 1186–1195. [Google Scholar] [CrossRef] [Green Version]
- Fensterle, J.; Bergmann, B.; Yone, C.L.; Hotz, C.; Meyer, S.R.; Spreng, S.; Goebel, W.; Rapp, U.R.; Gentschev, I. Cancer immunotherapy based on recombinant Salmonella enterica serovar Typhimurium aroA strains secreting prostate-specific antigen and cholera toxin subunit B. Cancer Gene Ther. 2008, 15, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, H.; Sato, E.; Briones, G.; Chen, L.M.; Matsuo, M.; Nagata, Y.; Ritter, G.; Jäger, E.; Nomura, H.; Kondo, S.; et al. In vivo antigen delivery by a Salmonella typhimurium type III secretion system for therapeutic cancer vaccines. J Clin. Investig. 2006, 116, 1946–1954. [Google Scholar] [CrossRef] [Green Version]
- Panthel, K.; Meinel, K.M.; Sevil Domènech, V.E.; Geginat, G.; Linkemann, K.; Busch, D.H.; Rüssmann, H. Prophylactic anti-tumor immunity against a murine fibrosarcoma triggered by the Salmonella type III secretion system. Microbes Infect. 2006, 8, 2539–2546. [Google Scholar] [CrossRef]
- Roider, E.; Jellbauer, S.; Köhn, B.; Berchtold, C.; Partilla, M.; Busch, D.H.; Rüssmann, H.; Panthel, K. Invasion and destruction of a murine fibrosarcoma by Salmonella-induced effector CD8 T cells as a therapeutic intervention against cancer. Cancer Immunol. Immunother. 2011, 60, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Epaulard, O.; Toussaint, B.; Quenee, L.; Derouazi, M.; Bosco, N.; Villiers, C.; Le Berre, R.; Guery, B.; Filopon, D.; Crombez, L.; et al. Anti-tumor immunotherapy via antigen delivery from a live attenuated genetically engineered Pseudomonas aeruginosa type III secretion system-based vector. Mol. Ther. 2006, 14, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Paton, A.W.; Morona, R.; Paton, J.C. Bioengineered microbes in disease therapy. Trends Mol. Med. 2012, 18, 417–425. [Google Scholar] [CrossRef]
- Grillot-Courvalin, C.; Goussard, S.; Courvalin, P. Wild-type intracellular bacteria deliver DNA into mammalian cells. Cell Microbiol. 2002, 4, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Akoachere, J.F.; Tanih, N.F.; Ndip, L.M.; Ndip, R.N. Phenotypic characterization of Salmonella typhimurium isolates from food-animals and abattoir drains in Buea, Cameroon. J. Health Popul. Nutr. 2009, 27, 612–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mi, Z.; Feng, Z.C.; Li, C.; Yang, X.; Ma, M.T.; Rong, P.F. Salmonella-Mediated Cancer Therapy: An Innovative Therapeutic Strategy. J. Cancer 2019, 10, 4765–4776. [Google Scholar] [CrossRef] [Green Version]
- Semenov, A.V.; van Overbeek, L.; Termorshuizen, A.J.; van Bruggen, A.H. Influence of aerobic and anaerobic conditions on survival of Escherichia coli O157:H7 and Salmonella enterica serovar Typhimurium in Luria-Bertani broth, farm-yard manure and slurry. J. Environ. Manag. 2011, 92, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, M.; Le’Negrate, G.; Krajewska, M.; Reed, J.C. Inhibition of tumor growth using salmonella expressing Fas ligand. J. Natl. Cancer Inst. 2008, 100, 1113–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganai, S.; Arenas, R.B.; Forbes, N.S. Tumour-targeted delivery of TRAIL using Salmonella typhimurium enhances breast cancer survival in mice. Br. J. Cancer 2009, 101, 1683–1691. [Google Scholar] [CrossRef]
- Yoon, W.S.; Chae, Y.S.; Hong, J.; Park, Y.K. Antitumor therapeutic effects of a genetically engineered Salmonella typhimurium harboring TNF-α in mice. Appl. Microbiol. Biotechnol. 2011, 89, 1807–1819. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yin, P.H.; Yang, S.S.; Li, Q.Y.; Chang, T.; Fang, L.; Shi, L.X.; Fang, G.E. Recombinant attenuated Salmonella typhimurium carrying a plasmid co-expressing ENDO-VEGI151 and survivin siRNA inhibits the growth of breast cancer in vivo. Mol. Med. Rep. 2013, 7, 1215–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, D.; Kim, K.S.; Kim, H.; Ko, K.C.; Song, J.J.; Choi, J.H.; Shin, M.; Min, J.J.; Jeong, J.H.; Choy, H.E. Anti-tumor activity of an immunotoxin (TGFα-PE38) delivered by attenuated Salmonella typhimurium. Oncotarget 2017, 8, 37550–37560. [Google Scholar] [CrossRef] [Green Version]
- Mansour, M.; Ismail, S.; Abou-Aisha, K. Bacterial delivery of the anti-tumor azurin-like protein Laz to glioblastoma cells. AMB Express 2020, 10, 59. [Google Scholar] [CrossRef]
- Gao, S.; Jung, J.H.; Lin, S.M.; Jang, A.Y.; Zhi, Y.; Bum Ahn, K.; Ji, H.J.; Hyang Lim, J.; Guo, H.; Choy, H.E.; et al. Development of Oxytolerant Salmonella typhimurium Using Radiation Mutation Technology (RMT) for Cancer Therapy. Sci. Rep. 2020, 10, 3764. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.N.; Phan, T.X.; Nam, T.K.; Nguyen, V.H.; Kim, H.S.; Bom, H.S.; Choy, H.E.; Hong, Y.; Min, J.J. Inhibition of tumor growth and metastasis by a combination of Escherichia coli-mediated cytolytic therapy and radiotherapy. Mol. Ther. 2010, 18, 635–642. [Google Scholar] [CrossRef]
- Chiang, C.-J.; Hong, Y.-H. In situ delivery of biobutyrate by probiotic Escherichia coli for cancer therapy. Sci. Rep. 2021, 11, 18172. [Google Scholar] [CrossRef]
- Zhang, Y.; Ji, W.; He, L.; Chen, Y.; Ding, X.; Sun, Y.; Hu, S.; Yang, H.; Huang, W.; Zhang, Y.; et al. E. coli Nissle 1917-Derived Minicells for Targeted Delivery of Chemotherapeutic Drug to Hypoxic Regions for Cancer Therapy. Theranostics 2018, 8, 1690–1705. [Google Scholar] [CrossRef]
- Chowdhury, S.; Castro, S.; Coker, C.; Hinchliffe, T.E.; Arpaia, N.; Danino, T. Programmable bacteria induce durable tumor regression and systemic antitumor immunity. Nat. Med. 2019, 25, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Radoshevich, L.; Cossart, P. Listeria monocytogenes: Towards a complete picture of its physiology and pathogenesis. Nat. Rev. Microbiol. 2018, 16, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Wood, L.M.; Paterson, Y. Attenuated Listeria monocytogenes: A powerful and versatile vector for the future of tumor immunotherapy. Front. Cell Infect. Microbiol. 2014, 4, 51. [Google Scholar] [CrossRef] [Green Version]
- Hense, M.; Domann, E.; Krusch, S.; Wachholz, P.; Dittmar, K.E.; Rohde, M.; Wehland, J.; Chakraborty, T.; Weiss, S. Eukaryotic expression plasmid transfer from the intracellular bacterium Listeria monocytogenes to host cells. Cell Microbiol. 2001, 3, 599–609. [Google Scholar] [CrossRef]
- Akin, D.; Sturgis, J.; Ragheb, K.; Sherman, D.; Burkholder, K.; Robinson, J.P.; Bhunia, A.K.; Mohammed, S.; Bashir, R. Bacteria-mediated delivery of nanoparticles and cargo into cells. Nat. Nanotechnol. 2007, 2, 441–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Pijkeren, J.P.; Morrissey, D.; Monk, I.R.; Cronin, M.; Rajendran, S.; O’Sullivan, G.C.; Gahan, C.G.; Tangney, M. A novel Listeria monocytogenes-based DNA delivery system for cancer gene therapy. Hum. Gene Ther. 2010, 21, 405–416. [Google Scholar] [CrossRef]
- Kim, S.H.; Castro, F.; Gonzalez, D.; Maciag, P.C.; Paterson, Y.; Gravekamp, C. Mage-b vaccine delivered by recombinant Listeria monocytogenes is highly effective against breast cancer metastases. Br. J. Cancer 2008, 99, 741–749. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.J.; Perumalsamy, H.; Markus, J.; Balusamy, S.R.; Wang, C.; Ho Kang, S.; Lee, S.; Park, S.Y.; Kim, S.; Castro-Aceituno, V.; et al. Development of Lactobacillus kimchicus DCY51(T)-mediated gold nanoparticles for delivery of ginsenoside compound K: In vitro photothermal effects and apoptosis detection in cancer cells. Artif. Cells Nanomed. Biotechnol. 2019, 47, 30–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kullberg, M.; McCarthy, R.; Anchordoquy, T.J. Gene delivery to Her-2+ breast cancer cells using a two-component delivery system to achieve specificity. Nanomedicine 2014, 10, 1253–1262. [Google Scholar] [CrossRef] [Green Version]
- Gilert, A.; Baruch, L.; Bronshtein, T.; Machluf, M. PLGA-Listeriolysin O microspheres: Opening the gate for cytosolic delivery of cancer antigens. Biomed. Microdevices 2016, 18, 23. [Google Scholar] [CrossRef]
- Mann, K.; Kullberg, M. Trastuzumab-targeted gene delivery to Her2-overexpressing breast cancer cells. Cancer Gene Ther. 2016, 23, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Al-Ani, A.W.; Zhang, L.; Ferreira, L.; Turyanska, L.; Bradshaw, T.D.; Thomas, N.R. Listeria innocua Dps as a nanoplatform for bioluminescence based photodynamic therapy utilizing Gaussia princeps luciferase and zinc protoporphyrin IX. Nanomedicine 2019, 20, 102005. [Google Scholar] [CrossRef] [PubMed]
- Barbé, S.; Van Mellaert, L.; Anné, J. The use of clostridial spores for cancer treatment. J. Appl. Microbiol. 2006, 101, 571–578. [Google Scholar] [CrossRef]
- Van Mellaert, L.; Barbé, S.; Anné, J. Clostridium spores as anti-tumour agents. Trends Microbiol. 2006, 14, 190–196. [Google Scholar] [CrossRef]
- Mengesha, A.; Dubois, L.; Chiu, R.K.; Paesmans, K.; Wouters, B.G.; Lambin, P.; Theys, J. Potential and limitations of bacterial-mediated cancer therapy. Front. Biosci. 2007, 12, 3880–3891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- St Jean, A.T.; Zhang, M.; Forbes, N.S. Bacterial therapies: Completing the cancer treatment toolbox. Curr. Opin. Biotechnol. 2008, 19, 511–517. [Google Scholar] [CrossRef] [Green Version]
- Wei, M.Q.; Mengesha, A.; Good, D.; Anné, J. Bacterial targeted tumour therapy-dawn of a new era. Cancer Lett. 2008, 259, 16–27. [Google Scholar] [CrossRef]
- Zu, C.; Wang, J. Tumor-colonizing bacteria: A potential tumor targeting therapy. Crit. Rev. Microbiol. 2014, 40, 225–235. [Google Scholar] [CrossRef]
- Connell, H.C. The Study and Treatment of Cancer by Proteolytic Enzymes: Preliminary Report. Can. Med. Assoc. J. 1935, 33, 364–370. [Google Scholar] [PubMed]
- Parker, R.C.; Plummer, H.C.; Siebenmann, C.O.; Chapman, M.G. Effect of histolyticus infection and toxin on transplantable mouse tumors. Proc. Soc. Exp. Biol. Med. 1947, 66, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Fox, M.E.; Lemmon, M.J.; Mauchline, M.L.; Davis, T.O.; Giaccia, A.J.; Minton, N.P.; Brown, J.M. Anaerobic bacteria as a delivery system for cancer gene therapy: In vitro activation of 5-fluorocytosine by genetically engineered clostridia. Gene Ther. 1996, 3, 173–178. [Google Scholar]
- Theys, J.; Nuyts, S.; Landuyt, W.; Van Mellaert, L.; Dillen, C.; Böhringer, M.; Dürre, P.; Lambin, P.; Anné, J. Stable Escherichia coli-Clostridium acetobutylicum shuttle vector for secretion of murine tumor necrosis factor alpha. Appl. Environ. Microbiol. 1999, 65, 4295–4300. [Google Scholar] [CrossRef] [Green Version]
- Barbé, S.; Van Mellaert, L.; Theys, J.; Geukens, N.; Lammertyn, E.; Lambin, P.; Anné, J. Secretory production of biologically active rat interleukin-2 by Clostridium acetobutylicum DSM792 as a tool for anti-tumor treatment. FEMS Microbiol. Lett. 2005, 246, 67–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casares, N.; Pequignot, M.O.; Tesniere, A.; Ghiringhelli, F.; Roux, S.; Chaput, N.; Schmitt, E.; Hamai, A.; Hervas-Stubbs, S.; Obeid, M.; et al. Caspase-dependent immunogenicity of doxorubicin-induced tumor cell death. J. Exp. Med. 2005, 202, 1691–1701. [Google Scholar] [CrossRef] [PubMed]
- Michaud, M.; Martins, I.; Sukkurwala, A.Q.; Adjemian, S.; Ma, Y.; Pellegatti, P.; Shen, S.; Kepp, O.; Scoazec, M.; Mignot, G.; et al. Autophagy-dependent anticancer immune responses induced by chemotherapeutic agents in mice. Science 2011, 334, 1573–1577. [Google Scholar] [CrossRef]
- Obeid, M.; Tesniere, A.; Ghiringhelli, F.; Fimia, G.M.; Apetoh, L.; Perfettini, J.L.; Castedo, M.; Mignot, G.; Panaretakis, T.; Casares, N.; et al. Calreticulin exposure dictates the immunogenicity of cancer cell death. Nat. Med. 2007, 13, 54–61. [Google Scholar] [CrossRef]
- Huang, T.; Li, S.; Li, G.; Tian, Y.; Wang, H.; Shi, L.; Perez-Cordon, G.; Mao, L.; Wang, X.; Wang, J.; et al. Utility of Clostridium difficile toxin B for inducing anti-tumor immunity. PLoS ONE 2014, 9, e110826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groot, A.J.; Verheesen, P.; Westerlaken, E.J.; Gort, E.H.; van der Groep, P.; Bovenschen, N.; van der Wall, E.; van Diest, P.J.; Shvarts, A. Identification by phage display of single-domain antibody fragments specific for the ODD domain in hypoxia-inducible factor 1alpha. Lab. Investig. 2006, 86, 345–356. [Google Scholar] [CrossRef] [Green Version]
- Collier, R.J. Diphtheria toxin: Mode of action and structure. Bacteriol. Rev. 1975, 39, 54–85. [Google Scholar] [CrossRef] [PubMed]
- Shafiee, F.; Aucoin, M.G.; Jahanian-Najafabadi, A. Targeted Diphtheria Toxin-Based Therapy: A Review Article. Front. Microbiol. 2019, 10, 2340. [Google Scholar] [CrossRef] [PubMed]
- Shapira, A.; Benhar, I. Toxin-based therapeutic approaches. Toxins 2010, 2, 2519–2583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Q.; Wang, Z.; Zhang, H.; Huang, Q.; Madsen, J.C.; Sachs, D.H.; Huang, C.A.; Wang, Z. Diphtheria toxin-based anti-human CD19 immunotoxin for targeting human CD19(+) tumors. Mol. Oncol. 2017, 11, 584–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.M.; Hall, W.A. Targeted toxins in brain tumor therapy. Toxins 2010, 2, 2645–2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsayad, K.; Kriz, J.; Moustakis, C.; Scobioala, S.; Reinartz, G.; Haverkamp, U.; Willich, N.; Weishaupt, C.; Stadler, R.; Sunderkötter, C.; et al. Total Skin Electron Beam for Primary Cutaneous T-cell Lymphoma. Int. J. Radiat. Oncol.Biol.Phys. 2015, 93, 1077–1086. [Google Scholar] [CrossRef] [PubMed]
- Zahaf, N.I.; Schmidt, G. Bacterial Toxins for Cancer Therapy. Toxins 2017, 9, 236. [Google Scholar] [CrossRef] [Green Version]
- Kiyokawa, T.; Shirono, K.; Hattori, T.; Nishimura, H.; Yamaguchi, K.; Nichols, J.C.; Strom, T.B.; Murphy, J.R.; Takatsuki, K. Cytotoxicity of interleukin 2-toxin toward lymphocytes from patients with adult T-cell leukemia. Cancer Res. 1989, 49, 4042–4046. [Google Scholar]
- Zhang, Y.; Schulte, W.; Pink, D.; Phipps, K.; Zijlstra, A.; Lewis, J.D.; Waisman, D.M. Sensitivity of cancer cells to truncated diphtheria toxin. PLoS ONE 2010, 5, e10498. [Google Scholar] [CrossRef]
- Leshem, Y.; Pastan, I. Pseudomonas Exotoxin Immunotoxins and Anti-Tumor Immunity: From Observations at the Patient’s Bedside to Evaluation in Preclinical Models. Toxins 2019, 11, 20. [Google Scholar] [CrossRef] [Green Version]
- Michalska, M.; Wolf, P. Pseudomonas Exotoxin A: Optimized by evolution for effective killing. Front. Microbiol. 2015, 6, 963. [Google Scholar] [CrossRef] [Green Version]
- Wolf, P.; Elsässer-Beile, U. Pseudomonas exotoxin A: From virulence factor to anti-cancer agent. Int. J. Med. Microbiol. 2009, 299, 161–176. [Google Scholar] [CrossRef]
- Iglewski, B.H.; Liu, P.V.; Kabat, D. Mechanism of action of Pseudomonas aeruginosa exotoxin Aiadenosine diphosphate-ribosylation of mammalian elongation factor 2 in vitro and in vivo. Infect. Immun. 1977, 15, 138–144. [Google Scholar] [CrossRef] [Green Version]
- Kreitman, R.J.; Hassan, R.; Fitzgerald, D.J.; Pastan, I. Phase I trial of continuous infusion anti-mesothelin recombinant immunotoxin SS1P. Clin. Cancer Res. 2009, 15, 5274–5279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Wang, B.; Jin, Z.; Ma, D.; Yang, W.; Zhao, R.; Jing, X.; Shen, B.; Peng, C.; Qiu, W. Pseudomonas aeruginosa-mannose-sensitive hemagglutinin inhibits pancreatic cancer cell proliferation and induces apoptosis via the EGFR pathway and caspase signaling. Oncotarget 2016, 7, 77916–77925. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Dong, Z.R.; Guo, Z.Y.; Wang, C.H.; Zhi, X.T.; Zhou, J.W.; Li, D.K.; Chen, Z.T.; Chen, Z.Q.; Hu, S.Y. Mannose-mediated inhibitory effects of PA-MSHA on invasion and metastasis of hepatocellular carcinoma via EGFR/Akt/IκBβ/NF-κB pathway. Liver Int. 2015, 35, 1416–1429. [Google Scholar] [CrossRef]
- van Geel-Schutten, G.H.; Flesch, F.; ten Brink, B.; Smith, M.R.; Dijkhuizen, L. Screening and characterization of Lactobacillus strains producing large amounts of exopolysaccharides. Appl. Microbiol. Biotechnol. 1998, 50, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Dethlefsen, L.; Huse, S.; Sogin, M.L.; Relman, D.A. The pervasive effects of an antibiotic on the human gut microbiota, as revealed by deep 16S rRNA sequencing. PLoS Biol. 2008, 6, e280. [Google Scholar] [CrossRef] [PubMed]
- Tamang, J.P. BIOCHEMICAL AND MODERN IDENTIFICATION TECHNIQUES | Microfloras of Fermented Foods. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Academic Press: Oxford, UK, 2014; pp. 250–258. [Google Scholar]
- Cano-Garrido, O.; Seras-Franzoso, J.; Garcia-Fruitós, E. Lactic acid bacteria: Reviewing the potential of a promising delivery live vector for biomedical purposes. Microb Cell Fact. 2015, 14, 137. [Google Scholar] [CrossRef] [Green Version]
- Oh, N.S.; Joung, J.Y.; Lee, J.Y.; Kim, Y. Probiotic and anti-inflammatory potential of Lactobacillus rhamnosus 4B15 and Lactobacillus gasseri 4M13 isolated from infant feces. PLoS ONE 2018, 13, e0192021. [Google Scholar] [CrossRef]
- Hibberd, A.A.; Lyra, A.; Ouwehand, A.C.; Rolny, P.; Lindegren, H.; Cedgård, L.; Wettergren, Y. Intestinal microbiota is altered in patients with colon cancer and modified by probiotic intervention. BMJ Open Gastroenterol. 2017, 4, e000145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tannock, I.F.; Rotin, D. Acid pH in tumors and its potential for therapeutic exploitation. Cancer Res. 1989, 49, 4373–4384. [Google Scholar]
- Chang, W.H.; Liu, J.J.; Chen, C.H.; Huang, T.S.; Lu, F.J. Growth inhibition and induction of apoptosis in MCF-7 breast cancer cells by fermented soy milk. Nutr. Cancer 2002, 43, 214–226. [Google Scholar] [CrossRef]
- Ohta, T.; Nakatsugi, S.; Watanabe, K.; Kawamori, T.; Ishikawa, F.; Morotomi, M.; Sugie, S.; Toda, T.; Sugimura, T.; Wakabayashi, K. Inhibitory effects of Bifidobacterium-fermented soy milk on 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine-induced rat mammary carcinogenesis, with a partial contribution of its component isoflavones. Carcinogenesis 2000, 21, 937–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takagi, A.; Kano, M.; Kaga, C. Possibility of breast cancer prevention: Use of soy isoflavones and fermented soy beverage produced using probiotics. Int. J. Mol. Sci. 2015, 16, 10907–10920. [Google Scholar] [CrossRef] [Green Version]
- Dróżdż, M.; Makuch, S.; Cieniuch, G.; Woźniak, M.; Ziółkowski, P. Obligate and facultative anaerobic bacteria in targeted cancer therapy: Current strategies and clinical applications. Life Sci. 2020, 261, 118296. [Google Scholar] [CrossRef]
- Park, J.; Kwon, M.; Lee, J.; Park, S.; Seo, J.; Roh, S. Anti-Cancer Effects of Lactobacillus plantarum L-14 Cell-Free Extract on Human Malignant Melanoma A375 Cells. Molecules 2020, 25, 3895. [Google Scholar] [CrossRef]
- Shida, K.; Nomoto, K. Probiotics as efficient immunopotentiators: Translational role in cancer prevention. Indian J. Med. Res. 2013, 138, 808–814. [Google Scholar] [PubMed]
- Kamarajan, P.; Hayami, T.; Matte, B.; Liu, Y.; Danciu, T.; Ramamoorthy, A.; Worden, F.; Kapila, S.; Kapila, Y. Nisin ZP, a Bacteriocin and Food Preservative, Inhibits Head and Neck Cancer Tumorigenesis and Prolongs Survival. PLoS ONE 2015, 10, e0131008. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Hu, M.; Feng, X.; XiaoLi, W.; Dong, D.; Wang, W. Preventive effect of Lactobacillus reuteri on melanoma. Biomed. Pharmacother. 2020, 126, 109929. [Google Scholar] [CrossRef] [PubMed]
- Yazdi, M.H.; Mahdavi, M.; Setayesh, N.; Esfandyar, M.; Shahverdi, A.R. Selenium nanoparticle-enriched Lactobacillus brevis causes more efficient immune responses in vivo and reduces the liver metastasis in metastatic form of mouse breast cancer. Daru 2013, 21, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raj, R.; Das, S. Development and application of anticancer fluorescent CdS nanoparticles enriched Lactobacillus bacteria as therapeutic microbots for human breast carcinoma. Appl. Microbiol. Biotechnol. 2017, 101, 5439–5451. [Google Scholar] [CrossRef] [PubMed]
- Steidler, L.; Neirynck, S.; Huyghebaert, N.; Snoeck, V.; Vermeire, A.; Goddeeris, B.; Cox, E.; Remon, J.P.; Remaut, E. Biological containment of genetically modified Lactococcus lactis for intestinal delivery of human interleukin 10. Nat. Biotechnol. 2003, 21, 785–789. [Google Scholar] [CrossRef]
- Bahey-El-Din, M.; Gahan, C.G.; Griffin, B.T. Lactococcus lactis as a cell factory for delivery of therapeutic proteins. Curr. Gene Ther. 2010, 10, 34–45. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, A.; van Sinderen, D. Bifidobacteria and Their Role as Members of the Human Gut Microbiota. Front. Microbiol. 2016, 7, 925. [Google Scholar] [CrossRef] [Green Version]
- Ngo, N.; Choucair, K.; Creeden, J.F.; Qaqish, H.; Bhavsar, K.; Murphy, C.; Lian, K.; Albrethsen, M.T.; Stanbery, L.; Phinney, R.C.; et al. Bifidobacterium spp: The promising Trojan Horse in the era of precision oncology. Future Oncol. 2019, 15, 3861–3876. [Google Scholar] [CrossRef]
- Wang, L.; Vuletic, I.; Deng, D.; Crielaard, W.; Xie, Z.; Zhou, K.; Zhang, J.; Sun, H.; Ren, Q.; Guo, C. Bifidobacterium breve as a delivery vector of IL-24 gene therapy for head and neck squamous cell carcinoma in vivo. Gene Therapy 2017, 24, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Xun, A.Y.; Wei, X.X.; Yao, J.; Wang, J.Y.; Shi, R.Y.; Yang, G.H.; Li, Y.X.; Xu, Z.L.; Lai, M.G.; et al. Bifidobacteria Expressing Tumstatin Protein for Antitumor Therapy in Tumor-Bearing Mice. Technol. Cancer Res. Treat. 2016, 15, 498–508. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Fu, G.-F.; Fan, Y.-R.; Liu, W.-H.; Liu, X.-J.; Wang, J.-J.; Xu, G.-X. Bifidobacterium adolescentis as a delivery system of endostatin for cancer gene therapy: Selective inhibitor of angiogenesis and hypoxic tumor growth. Cancer Gene Therapy 2003, 10, 105–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimori, M. Genetically engineered bifidobacterium as a drug delivery system for systemic therapy of metastatic breast cancer patients. Breast Cancer 2006, 13, 27–31. [Google Scholar] [CrossRef]
- Cronin, M.; Morrissey, D.; Rajendran, S.; El Mashad, S.M.; van Sinderen, D.; O’Sullivan, G.C.; Tangney, M. Orally administered bifidobacteria as vehicles for delivery of agents to systemic tumors. Mol. Ther. 2010, 18, 1397–1407. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, T.; Shimizu, H.; Akiyama, Y.; Taniguchi, S. In situ delivery and production system of trastuzumab scFv with Bifidobacterium. Biochem. Biophys. Res. Commun. 2017, 493, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Bazylinski, D.A.; Williams, T.J.; Lefèvre, C.T.; Berg, R.J.; Zhang, C.L.; Bowser, S.S.; Dean, A.J.; Beveridge, T.J. Magnetococcus marinus gen. nov., sp. nov., a marine, magnetotactic bacterium that represents a novel lineage (Magnetococcaceae fam. nov., Magnetococcales ord. nov.) at the base of the Alphaproteobacteria. Int. J. Syst. Evol. Microbiol. 2013, 63, 801–808. [Google Scholar] [CrossRef]
- Afkhami, F.; Taherkhani, S.; Mohammadi, M.; Martel, S. Encapsulation of magnetotactic bacteria for targeted and controlled delivery of anticancer agents for tumor therapy. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2011, 2011, 6668–6671. [Google Scholar] [CrossRef]
- Felfoul, O.; Mohammadi, M.; Taherkhani, S.; de Lanauze, D.; Zhong Xu, Y.; Loghin, D.; Essa, S.; Jancik, S.; Houle, D.; Lafleur, M.; et al. Magneto-aerotactic bacteria deliver drug-containing nanoliposomes to tumour hypoxic regions. Nat. Nanotechnol. 2016, 11, 941–947. [Google Scholar] [CrossRef]
- Zhao, M.; Yang, M.; Li, X.M.; Jiang, P.; Baranov, E.; Li, S.; Xu, M.; Penman, S.; Hoffman, R.M. Tumor-targeting bacterial therapy with amino acid auxotrophs of GFP-expressing Salmonella typhimurium. Proc. Natl. Acad. Sci. USA 2005, 102, 755–760. [Google Scholar] [CrossRef] [Green Version]
- Clairmont, C.; Lee, K.C.; Pike, J.; Ittensohn, M.; Low, K.B.; Pawelek, J.; Bermudes, D.; Brecher, S.M.; Margitich, D.; Turnier, J.; et al. Biodistribution and genetic stability of the novel antitumor agent VNP20009, a genetically modified strain of Salmonella typhimurium. J. Infect. Dis. 2000, 181, 1996–2002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawelek, J.M.; Low, K.B.; Bermudes, D. Tumor-targeted Salmonella as a novel anticancer vector. Cancer Res. 1997, 57, 4537–4544. [Google Scholar] [PubMed]
- Zheng, J.H.; Min, J.J. Targeted Cancer Therapy Using Engineered Salmonella typhimurium. Chonnam Med. J. 2016, 52, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, V.H.; Kim, H.S.; Ha, J.M.; Hong, Y.; Choy, H.E.; Min, J.J. Genetically engineered Salmonella typhimurium as an imageable therapeutic probe for cancer. Cancer Res. 2010, 70, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.H.; Nguyen, V.H.; Jiang, S.N.; Park, S.H.; Tan, W.; Hong, S.H.; Shin, M.G.; Chung, I.J.; Hong, Y.; Bom, H.S.; et al. Two-step enhanced cancer immunotherapy with engineered Salmonella typhimurium secreting heterologous flagellin. Sci. Transl. Med. 2017, 9, 9537. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.Z.; Dong, Y.J.; Huang, H.; Li, S.; Zhong, Y.; Liu, S.L.; Wang, Y.D. Oral vaccination with attenuated Salmonella enterica strains encoding T-cell epitopes from tumor antigen NY-ESO-1 induces specific cytotoxic T-lymphocyte responses. Clin. Vaccine Immunol. 2010, 17, 889–894. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.; Casey, G.; Cronin, M.; Rajendran, S.; Sweeney, P.; Tangney, M.; O’Sullivan, G.C. Induction of effective antitumor response after mucosal bacterial vector mediated DNA vaccination with endogenous prostate cancer specific antigen. J. Urol. 2011, 186, 687–693. [Google Scholar] [CrossRef] [PubMed]
- al-Ramadi, B.K.; Fernandez-Cabezudo, M.J.; El-Hasasna, H.; Al-Salam, S.; Bashir, G.; Chouaib, S. Potent anti-tumor activity of systemically-administered IL2-expressing Salmonella correlates with decreased angiogenesis and enhanced tumor apoptosis. Clin. Immunol. 2009, 130, 89–97. [Google Scholar] [CrossRef]
- Felgner, S.; Frahm, M.; Kocijancic, D.; Rohde, M.; Eckweiler, D.; Bielecka, A.; Bueno, E.; Cava, F.; Abraham, W.R.; Curtiss, R., 3rd; et al. aroA-Deficient Salmonella enterica Serovar Typhimurium Is More Than a Metabolically Attenuated Mutant. mBio 2016, 7, e01220-16. [Google Scholar] [CrossRef] [Green Version]
- Rüssmann, H.; Shams, H.; Poblete, F.; Fu, Y.; Galán, J.E.; Donis, R.O. Delivery of epitopes by the Salmonella type III secretion system for vaccine development. Science 1998, 281, 565–568. [Google Scholar] [CrossRef] [Green Version]
- Xiong, G.; Husseiny, M.I.; Song, L.; Erdreich-Epstein, A.; Shackleford, G.M.; Seeger, R.C.; Jäckel, D.; Hensel, M.; Metelitsa, L.S. Novel cancer vaccine based on genes of Salmonella pathogenicity island 2. Int. J. Cancer 2010, 126, 2622–2634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.; Yu, B.; Cai, C.H.; Huang, J.D. Angiogenic inhibitors delivered by the type III secretion system of tumor-targeting Salmonella typhimurium safely shrink tumors in mice. AMB Express 2016, 6, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorenson, B.S.; Banton, K.L.; Frykman, N.L.; Leonard, A.S.; Saltzman, D.A. Attenuated Salmonella typhimurium with IL-2 gene reduces pulmonary metastases in murine osteosarcoma. Clin. Orthop. Relat. Res. 2008, 466, 1285–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewēn, S.; Zhou, H.; Hu, H.D.; Cheng, T.; Markowitz, D.; Reisfeld, R.A.; Xiang, R.; Luo, Y. A Legumain-based minigene vaccine targets the tumor stroma and suppresses breast cancer growth and angiogenesis. Cancer Immunol. Immunother. 2008, 57, 507–515. [Google Scholar] [CrossRef]
- Jellbauer, S.; Panthel, K.; Hetrodt, J.H.; Rüssmann, H. CD8 T-cell induction against vascular endothelial growth factor receptor 2 by Salmonella for vaccination purposes against a murine melanoma. PLoS ONE 2012, 7, e34214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, K.; Liu, Q.; Li, P.; Han, Y.; Bian, X.; Tang, Y.; Kong, Q. Endostatin gene therapy delivered by attenuated Salmonella typhimurium in murine tumor models. Cancer Gene Ther. 2018, 25, 167–183. [Google Scholar] [CrossRef]
- Mesa-Pereira, B.; Medina, C.; Camacho, E.M.; Flores, A.; Santero, E. Improved cytotoxic effects of Salmonella-producing cytosine deaminase in tumour cells. Microb Biotechnol. 2015, 8, 169–176. [Google Scholar] [CrossRef] [Green Version]
- Gunn, G.R.; Zubair, A.; Peters, C.; Pan, Z.K.; Wu, T.C.; Paterson, Y. Two Listeria monocytogenes vaccine vectors that express different molecular forms of human papilloma virus-16 (HPV-16) E7 induce qualitatively different T cell immunity that correlates with their ability to induce regression of established tumors immortalized by HPV-16. J. Immunol. 2001, 167, 6471–6479. [Google Scholar] [CrossRef] [Green Version]
- Shahabi, V.; Reyes-Reyes, M.; Wallecha, A.; Rivera, S.; Paterson, Y.; Maciag, P. Development of a Listeria monocytogenes based vaccine against prostate cancer. Cancer Immunol. Immunother. 2008, 57, 1301–1313. [Google Scholar] [CrossRef]
- Carvalho, F.; Sousa, S.; Cabanes, D. How Listeria monocytogenes organizes its surface for virulence. Front. Cell Infect. Microbiol. 2014, 4, 48. [Google Scholar] [CrossRef] [PubMed]
- Rafelski, S.M.; Theriot, J.A. Mechanism of polarization of Listeria monocytogenes surface protein ActA. Mol. Microbiol. 2006, 59, 1262–1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sewell, D.A.; Shahabi, V.; Gunn, G.R., 3rd; Pan, Z.K.; Dominiecki, M.E.; Paterson, Y. Recombinant Listeria vaccines containing PEST sequences are potent immune adjuvants for the tumor-associated antigen human papillomavirus-16 E7. Cancer Res. 2004, 64, 8821–8825. [Google Scholar] [CrossRef] [Green Version]
- Olino, K.; Wada, S.; Edil, B.H.; Pan, X.; Meckel, K.; Weber, W.; Slansky, J.; Tamada, K.; Lauer, P.; Brockstedt, D.; et al. Tumor-associated antigen expressing Listeria monocytogenes induces effective primary and memory T-cell responses against hepatic colorectal cancer metastases. Ann. Surg. Oncol. 2012, 19 (Suppl. 3), S597–S607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, P.V.; Blair, B.M.; Zeller, S.; Kotton, C.N.; Hohmann, E.L. Attenuated Listeria monocytogenes vaccine vectors expressing influenza A nucleoprotein: Preclinical evaluation and oral inoculation of volunteers. Microbiol. Immunol. 2011, 55, 304–317. [Google Scholar] [CrossRef] [Green Version]
- Morales, A.; Eidinger, D.; Bruce, A.W. Intracavitary Bacillus Calmette-Guerin in the treatment of superficial bladder tumors. J. Urol. 1976, 116, 180–183. [Google Scholar] [CrossRef]
- Biot, C.; Rentsch, C.A.; Gsponer, J.R.; Birkhäuser, F.D.; Jusforgues-Saklani, H.; Lemaître, F.; Auriau, C.; Bachmann, A.; Bousso, P.; Demangel, C.; et al. Preexisting BCG-specific T cells improve intravesical immunotherapy for bladder cancer. Sci. Transl. Med. 2012, 4, 137ra172. [Google Scholar] [CrossRef]
- Roberts, N.J.; Zhang, L.; Janku, F.; Collins, A.; Bai, R.Y.; Staedtke, V.; Rusk, A.W.; Tung, D.; Miller, M.; Roix, J.; et al. Intratumoral injection of Clostridium novyi-NT spores induces antitumor responses. Sci. Transl. Med. 2014, 6, 249ra111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theys, J.; Lambin, P. Clostridium to treat cancer: Dream or reality? Ann. Transl. Med. 2015, 3, S21. [Google Scholar] [CrossRef]
- Dang, L.H.; Bettegowda, C.; Huso, D.L.; Kinzler, K.W.; Vogelstein, B. Combination bacteriolytic therapy for the treatment of experimental tumors. Proc. Natl. Acad. Sci. USA 2001, 98, 15155–15160. [Google Scholar] [CrossRef] [Green Version]
- Park, B.W.; Zhuang, J.; Yasa, O.; Sitti, M. Multifunctional Bacteria-Driven Microswimmers for Targeted Active Drug Delivery. ACS Nano 2017, 11, 8910–8923. [Google Scholar] [CrossRef]
- Ohta, N.; Fukase, S.; Watanabe, T.; Ito, T.; Aoyagi, M. Effects and mechanism of OK-432 therapy in various neck cystic lesions. Acta Otolaryngol. 2010, 130, 1287–1292. [Google Scholar] [CrossRef]
- Ohta, N.; Fukase, S.; Suzuki, Y.; Ishida, A.; Aoyagi, M. Treatments of various otolaryngological cystic diseases by OK-4321: Its indications and limitations. Laryngoscope 2010, 120, 2193–2196. [Google Scholar] [CrossRef] [PubMed]
- Kono, M.; Satomi, T.; Abukawa, H.; Hasegawa, O.; Watanabe, M.; Chikazu, D. Evaluation of OK-432 Injection Therapy as Possible Primary Treatment of Intraoral Ranula. J. Oral Maxillofac. Surg 2017, 75, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Ogita, S.; Tsuto, T.; Nakamura, K.; Deguchi, E.; Tokiwa, K.; Iwai, N. OK-432 therapy for lymphangioma in children: Why and how does it work? J. Pediatr. Surg. 1996, 31, 477–480. [Google Scholar] [CrossRef]
- Veena, V.K.; Popavath, R.N.; Kennedy, K.; Sakthivel, N. In vitro antiproliferative, pro-apoptotic, antimetastatic and anti-inflammatory potential of 2,4-diacteylphloroglucinol (DAPG) by Pseudomonas aeruginosa strain FP10. Apoptosis 2015, 20, 1281–1295. [Google Scholar] [CrossRef]
- Riaz Rajoka, M.S.; Zhao, H.; Mehwish, H.M.; Li, N.; Lu, Y.; Lian, Z.; Shao, D.; Jin, M.; Li, Q.; Zhao, L.; et al. Anti-tumor potential of cell free culture supernatant of Lactobacillus rhamnosus strains isolated from human breast milk. Food Res. Int. 2019, 123, 286–297. [Google Scholar] [CrossRef]
- Liu, C.F.; Pan, T.M. In Vitro Effects of Lactic Acid Bacteria on Cancer Cell Viability and Antioxidant Activity. J. Food Drug Anal. 2010, 18, 77–86. [Google Scholar] [CrossRef]
- Asoudeh-Fard, A.; Barzegari, A.; Dehnad, A.; Bastani, S.; Golchin, A.; Omidi, Y. Lactobacillus plantarum induces apoptosis in oral cancer KB cells through upregulation of PTEN and downregulation of MAPK signalling pathways. Bioimpacts 2017, 7, 193–198. [Google Scholar] [CrossRef]
- Abdolalipour, E.; Mahooti, M.; Salehzadeh, A.; Torabi, A.; Mohebbi, S.R.; Gorji, A.; Ghaemi, A. Evaluation of the antitumor immune responses of probiotic Bifidobacterium bifidum in human papillomavirus-induced tumor model. Microb. Pathog. 2020, 145, 104207. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, K.; Wu, W.; Lv, L.; Bian, X.; Yang, L.; Wang, Q.; Li, Y.; Ye, J.; Fang, D.; et al. Administration of Bifidobacterium bifidum CGMCC 15068 modulates gut microbiota and metabolome in azoxymethane (AOM)/dextran sulphate sodium (DSS)-induced colitis-associated colon cancer (CAC) in mice. Appl. Microbiol. Biotechnol. 2020, 104, 5915–5928. [Google Scholar] [CrossRef]
- Kitagawa, K.; Gonoi, R.; Tatsumi, M.; Kadowaki, M.; Katayama, T.; Hashii, Y.; Fujisawa, M.; Shirakawa, T. Preclinical Development of a WT1 Oral Cancer Vaccine Using a Bacterial Vector to Treat Castration-Resistant Prostate Cancer. Mol. Cancer Ther. 2019, 18, 980–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coley, W.B., II. Contribution to the Knowledge of Sarcoma. Ann. Surg 1891, 14, 199–220. [Google Scholar] [CrossRef]
- Heppner, F.; Möse, J.R. The liquefaction (oncolysis) of malignant gliomas by a non pathogenic Clostridium. Acta Neurochir (Wien.) 1978, 42, 123–125. [Google Scholar] [CrossRef] [PubMed]
- Basu, P.; Mehta, A.; Jain, M.; Gupta, S.; Nagarkar, R.V.; John, S.; Petit, R. A Randomized Phase 2 Study of ADXS11-001 Listeria monocytogenes-Listeriolysin O Immunotherapy With or Without Cisplatin in Treatment of Advanced Cervical Cancer. Int. J. Gynecol Cancer 2018, 28, 764–772. [Google Scholar] [CrossRef] [Green Version]
- Le, D.T.; Wang-Gillam, A.; Picozzi, V.; Greten, T.F.; Crocenzi, T.; Springett, G.; Morse, M.; Zeh, H.; Cohen, D.; Fine, R.L.; et al. Safety and survival with GVAX pancreas prime and Listeria Monocytogenes-expressing mesothelin (CRS-207) boost vaccines for metastatic pancreatic cancer. J. Clin. Oncol. 2015, 33, 1325–1333. [Google Scholar] [CrossRef] [Green Version]
- Gniadek, T.J.; Augustin, L.; Schottel, J.; Leonard, A.; Saltzman, D.; Greeno, E.; Batist, G. A Phase I, Dose Escalation, Single Dose Trial of Oral Attenuated Salmonella typhimurium Containing Human IL-2 in Patients With Metastatic Gastrointestinal Cancers. J. Immunother. 2020, 43, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Schmitz-Winnenthal, F.H.; Hohmann, N.; Schmidt, T.; Podola, L.; Friedrich, T.; Lubenau, H.; Springer, M.; Wieckowski, S.; Breiner, K.M.; Mikus, G.; et al. A phase 1 trial extension to assess immunologic efficacy and safety of prime-boost vaccination with VXM01, an oral T cell vaccine against VEGFR2, in patients with advanced pancreatic cancer. Oncoimmunology 2018, 7, e1303584. [Google Scholar] [CrossRef] [PubMed]
- Griffin, M.E.; Espinosa, J.; Becker, J.L.; Luo, J.D.; Carroll, T.S.; Jha, J.K.; Fanger, G.R.; Hang, H.C. Enterococcus peptidoglycan remodeling promotes checkpoint inhibitor cancer immunotherapy. Science 2021, 373, 1040–1046. [Google Scholar] [CrossRef] [PubMed]
- Duong, M.T.-Q.; Qin, Y.; You, S.-H.; Min, J.-J. Bacteria-cancer interactions: Bacteria-based cancer therapy. Exp. Mol. Med. 2019, 51, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyake, K.; Murata, T.; Murakami, T.; Zhao, M.; Kiyuna, T.; Kawaguchi, K.; Igarashi, K.; Miyake, M.; Lwin, T.M.; Hozumi, C.; et al. Tumor-targeting Salmonella typhimurium A1-R overcomes nab-paclitaxel resistance in a cervical cancer PDOX mouse model. Arch. Gynecol. Obstet. 2019, 299, 1683–1690. [Google Scholar] [CrossRef] [PubMed]

| Bacterial Strain | Type of Cancer | Clinical Phase | Identifier No. | Reference (All Links Were Accessed on 16 December 2021) |
|---|---|---|---|---|
| C. histolyticum | Lipoma | I | NCT01613313 | https://clinicaltrials.gov/ct2/show/NCT01613313 |
| Lipoma | I | NCT02249052 | https://clinicaltrials.gov/ct2/show/NCT02249052 | |
| C. butyricum M55 | Vascular glioblastoma | I | - | [190] |
| C. novyi-NT | Solid tumor malignancies | I | NCT01924689 | [173] |
| Colorectal cancer | I | NCT00358397 | https://clinicaltrials.gov/ct2/show/NCT00358397 | |
| Solid tumor malignancies | I | NCT01118819 | https://clinicaltrials.gov/ct2/show/NCT01118819 | |
| Refractory advanced solid tumors | Ib | NCT03435952 | https://clinicaltrials.gov/ct2/show/NCT03435952 | |
| L. monocytogenes | Cervical cancer | II | - | [191] |
| Cervical cancer | III | NCT02853604 | https://clinicaltrials.gov/ct2/show/NCT02853604 | |
| Metastatic pancreatic tumors | II | - | [192] | |
| Malignant epithelial mesothelioma Adenocarcinoma of the pancreas, Non-small cell lung adenocarcinoma of the ovaries | I | NCT00585845 | https://clinicaltrials.gov/ct2/show/NCT00585845 | |
| HPV-16 +ve oropharyngeal carcinoma | I | NCT01598792 | https://clinicaltrials.gov/ct2/show/NCT01598792 | |
| L. monocytogenes (LADD) | Prostatic neoplasms (castration resistant) | II | NCT01613313 | https://clinicaltrials.gov/ct2/show/NCT02625857 |
| Non-small cell lung carcinoma | I | NCT02592967 | https://clinicaltrials.gov/ct2/show/NCT02592967 | |
| S. typhimurium (χ4550) | Hepatocellular carcinoma | I | - | [193] |
| S. typhimurium VNP20009 | Metastatic melanoma, metastatic renal cell carcinoma | I | - | [15] |
| S. typhimurium VNP20009 | Melanoma | I | - | [17] |
| S. typhimurium VNP20009 expressing TAPET-CD (cytosine deaminase) | Head and neck, and esophageal adenocarcinoma | I | - | [[18]] |
| S. typhimurium VNP20009 | Advanced metastatic solid tumors | I | NCT00004216 | https://clinicaltrials.gov/ct2/show/NCT00004216 |
| Solid tumors | I | NCT00006254 | https://clinicaltrials.gov/ct2/show/NCT00006254 | |
| Neoplasm metastatic tumor | I | NCT00004988 | https://clinicaltrials.gov/ct2/show/NCT00004988 | |
| S. typhimurium expressing IL-2 | Liver cancer | I | NCT01099631 | https://clinicaltrials.gov/ct2/show/NCT01099631 |
| S. typhimurium Ty21a VXM01 | Pancreatic cancer | I | - | [194] |
| Mixed Bacterial Vaccine | Malignant tumors | I | NCT00623831 | https://clinicaltrials.gov/ct2/show/NCT00623831 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gupta, K.H.; Nowicki, C.; Giurini, E.F.; Marzo, A.L.; Zloza, A. Bacterial-Based Cancer Therapy (BBCT): Recent Advances, Current Challenges, and Future Prospects for Cancer Immunotherapy. Vaccines 2021, 9, 1497. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9121497
Gupta KH, Nowicki C, Giurini EF, Marzo AL, Zloza A. Bacterial-Based Cancer Therapy (BBCT): Recent Advances, Current Challenges, and Future Prospects for Cancer Immunotherapy. Vaccines. 2021; 9(12):1497. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9121497
Chicago/Turabian StyleGupta, Kajal H., Christina Nowicki, Eileena F. Giurini, Amanda L. Marzo, and Andrew Zloza. 2021. "Bacterial-Based Cancer Therapy (BBCT): Recent Advances, Current Challenges, and Future Prospects for Cancer Immunotherapy" Vaccines 9, no. 12: 1497. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9121497