
TLR4 Agonist Combined with Trivalent Protein JointS of Streptococcus suis Provides Immunological Protection in Animals

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Bacterial Strains and Culture Conditions
2.3. Gene Amplification and Construction of Expression Vectors
2.4. The Expression, Purification, and Antigenicity Identification of GAPDH, MRP, DLD, and GAPDH-MRP-DLD (JointS)
2.5. Zebrafish Immunization with JointS and Challenge with S. suis
2.5.1. A 50% Lethal Dose of S. suis Determination in Zebrafish
2.5.2. Immunization and Challenge of Zebrafish
2.6. The Cytokine Expression of Immune Cells Induced by TLR Agonists Using qRT-PCR
2.7. Mice Immunization with JointS and Challenge with SS2
2.8. Piglets Immunization with JointS and Challenge with SS2
2.9. Enzyme-Linked Immunosorbent Assay (ELISA)
2.10. Statistical Analysis
3. Results
3.1. Preparation and Immunoreactivity of GAPDH, MRP, DLD, and JointS
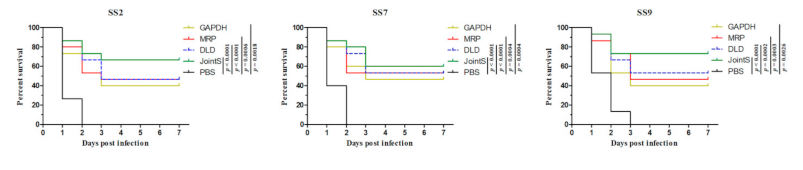
3.2. The Immune-Protective Effect of Recombinant Protein against Different Serotypes of S. suis in Zebrafish
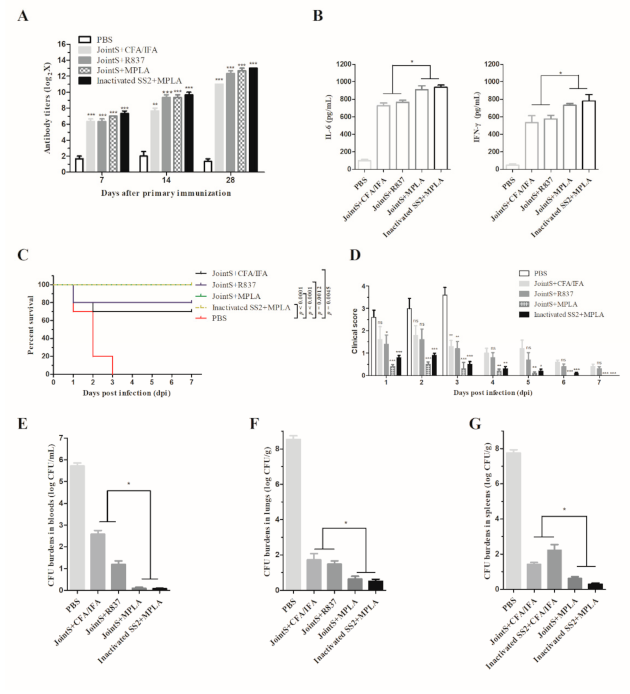
3.3. TLR-4 Agonist (MPLA) Stimulates Immune Cells to Produce Cytokines
3.4. MPLA Combined with JointS to Prevent S. suis Infection in Mice
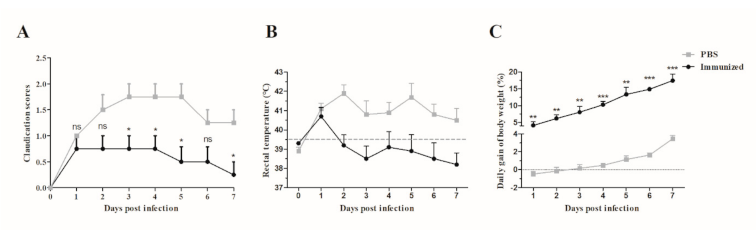
3.5. Application of JointS in Prevention of S. suis Infection In Piglets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tan, C.; Zhang, A.; Chen, H.; Zhou, R. Recent proceedings on prevalence and pathogenesis of Streptococcus suis. Curr. Issues Mol. Biol. 2019, 32, 473–520. [Google Scholar] [CrossRef]
- Segura, M. Streptococcus suis research: Progress and challenges. Pathogens 2020, 9, 707. [Google Scholar] [CrossRef]
- Xing, X.; Bi, S.; Fan, X.; Jin, M.; Liu, W.; Wang, B. Intranasal vaccination with multiple virulence factors promotes mucosal clearance of Streptococcus suis across serotypes and protects against meningitis in mice. J. Infect. Dis. 2019, 220, 1679–1687. [Google Scholar] [CrossRef]
- Estrada, A.A.; Gottschalk, M.; Rossow, S.; Rendahl, A.; Gebhart, C.; Marthaler, D.G. Serotype and genotype (multilocus sequence type) of Streptococcus suis isolates from the United States serve as predictors of pathotype. J. Clin. Microbiol. 2019, 57. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, Y.; Liu, B.; Wang, S.; Li, J.; Gong, S.; Sun, L.; Yi, L. pdh modulate virulence through reducing stress tolerance and biofilm formation of Streptococcus suis serotype 2. Virulence 2019, 10, 588–599. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Li, R.; Zhang, A.; He, H.; Hua, Y.; Xia, J.; Cai, X.; Chen, H.; Jin, M. Characterization of Streptococcus suis isolates from the diseased pigs in China between 2003 and 2007. Vet. Microbiol. 2009, 137, 196–201. [Google Scholar] [CrossRef]
- Tang, J.; Wang, C.; Feng, Y.; Yang, W.; Song, H.; Chen, Z.; Yu, H.; Pan, X.; Zhou, X.; Wang, H.; et al. Streptococcal toxic shock syndrome caused by Streptococcus suis serotype 2. PLoS Med. 2006, 3, e151. [Google Scholar] [CrossRef] [Green Version]
- Segura, M. Streptococcus suis vaccines: Candidate antigens and progress. Expert Rev. Vaccines 2015, 14, 1587–1608. [Google Scholar] [CrossRef]
- Yi, L.; Du, Y.; Mao, C.; Li, J.; Jin, M.; Sun, L.; Wang, Y. Immunogenicity and protective ability of RpoE against Streptococcus suis serotype 2. J. Appl. Microbiol. 2020. [Google Scholar] [CrossRef]
- Wang, H.; Qin, Z.; Li, M. Recent Advances in pathogenic streptococcus vaccine development. Curr. Issues Mol. Biol. 2019, 32, 645–700. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, G.; Tang, F.; Shao, J.; Lu, Y.; Bao, Y.; Yao, H.; Lu, C. Pre-absorbed immunoproteomics: A novel method for the detection of Streptococcus suis surface proteins. PLoS ONE 2011, 6, e21234. [Google Scholar] [CrossRef] [PubMed]
- Geng, H.; Zhu, L.; Yuan, Y.; Zhang, W.; Li, W.; Wang, J.; Zheng, Y.; Wei, K.; Cao, W.; Wang, H.; et al. Identification and characterization of novel immunogenic proteins of Streptococcus suis serotype 2. J. Proteome Res. 2008, 7, 4132–4142. [Google Scholar] [CrossRef]
- Baums, C.G.; Kock, C.; Beineke, A.; Bennecke, K.; Goethe, R.; Schroder, C.; Waldmann, K.H.; Valentin-Weigand, P. Streptococcus suis bacterin and subunit vaccine immunogenicities and protective efficacies against serotypes 2 and 9. Clin. Vaccine Immunol. 2009, 16, 200–208. [Google Scholar] [CrossRef] [Green Version]
- Wisselink, H.J.; Vecht, U.; Stockhofe-Zurwieden, N.; Smith, H.E. Protection of pigs against challenge with virulent Streptococcus suis serotype 2 strains by a muramidase-released protein and extracellular factor vaccine. Vet. Rec. 2001, 148, 473–477. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Lu, C. Adhesion activity of glyceraldehyde-3-phosphate dehydrogenase in a Chinese Streptococcus suis type 2 strain. Berl. Und Munch. Tierarztl. Wochenschr. 2007, 120, 207–209. [Google Scholar]
- Devasundaram, S.; Raja, A. Dihydrolipoamide dehydrogenase-Lpd (Rv0462)-specific T cell recall responses are higher in healthy household contacts of TB: A novel immunodominant antigen from M. tuberculosis. J. Leukoc. Biol. 2017, 102, 135–151. [Google Scholar] [CrossRef] [Green Version]
- Pang, H.; Chen, L.; Hoare, R.; Huang, Y.; Wu, Z.; Jian, J. Identification of DLD, by immunoproteomic analysis and evaluation as a potential vaccine antigen against three Vibrio species in Epinephelus coioides. Vaccine 2016, 34, 1225–1231. [Google Scholar] [CrossRef] [PubMed]
- Reed, S.G.; Tomai, M.; Gale, M.J., Jr. New horizons in adjuvants for vaccine development. Curr. Opin. Immunol. 2020, 65, 97–101. [Google Scholar] [CrossRef]
- Wilk, M.M.; Mills, K.H.G. CD4 TRM Cells Following infection and immunization: Implications for more effective vaccine design. Front. Immunol. 2018, 9, 1860. [Google Scholar] [CrossRef]
- Bharucha, T.; Ming, D.; Breuer, J. A critical appraisal of ‘Shingrix’, a novel herpes zoster subunit vaccine (HZ/Su or GSK1437173A) for varicella zoster virus. Hum. Vaccines Immunother. 2017, 13, 1789–1797. [Google Scholar] [CrossRef] [Green Version]
- Celis, E. Toll-like receptor ligands energize peptide vaccines through multiple paths. Cancer Res. 2007, 67, 7945–7947. [Google Scholar] [CrossRef] [Green Version]
- Kanzler, H.; Barrat, F.J.; Hessel, E.M.; Coffman, R.L. Therapeutic targeting of innate immunity with Toll-like receptor agonists and antagonists. Nat. Med. 2007, 13, 552–559. [Google Scholar] [CrossRef]
- Meng, X.; Shi, Y.; Ji, W.; Meng, X.; Zhang, J.; Wang, H.; Lu, C.; Sun, J.; Yan, Y. Application of a bacteriophage lysin to disrupt biofilms formed by the animal pathogen Streptococcus suis. Appl. Environ. Microbiol. 2011, 77, 8272–8279. [Google Scholar] [CrossRef] [Green Version]
- Aida, Y.; Pabst, M.J. Removal of endotoxin from protein solutions by phase separation using Triton X-114. J. Immunol. Methods 1990, 132, 191–195. [Google Scholar] [CrossRef]
- Patterson, H.; Saralahti, A.; Parikka, M.; Dramsi, S.; Trieu-Cuot, P.; Poyart, C.; Rounioja, S.; Ramet, M. Adult zebrafish model of bacterial meningitis in Streptococcus agalactiae infection. Dev. Comp. Immunol. 2012, 38, 447–455. [Google Scholar] [CrossRef]
- Stanic, M. A simplification of the estimation of the 50 percent endpoints according to the Reed and Muench method. Pathol. Microbiol. 1963, 26, 298–302. [Google Scholar]
- Zhang, Y.; Li, H.; Xiang, X.; Lu, Y.; Sharma, M.; Li, Z.; Liu, K.; Wei, J.; Shao, D.; Li, B.; et al. Identification of DNMT3B2 as the predominant isoform of DNMT3B in porcine alveolar macrophages and its involvement in LPS-stimulated TNF-alpha expression. Genes 2020, 11, 1065. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Wetherall, B.L.; Hallsworth, P.G.; McDonald, P.J. Enzyme-linked immunosorbent assay for detection of Haemophilus influenzae type b antigen. J. Clin. Microbiol. 1980, 11, 573–578. [Google Scholar] [CrossRef] [Green Version]
- Staton, G.J.; Carter, S.D.; Ainsworth, S.; Mullin, J.; Smith, R.F.; Evans, N.J. Putative beta-Barrel Outer Membrane Proteins of the Bovine Digital Dermatitis-Associated Treponemes: Identification, Functional characterization, and immunogenicity. Infect. Immun. 2020, 88. [Google Scholar] [CrossRef] [Green Version]
- Hallstrom, T.; Morgelin, M.; Barthel, D.; Raguse, M.; Kunert, A.; Hoffmann, R.; Skerka, C.; Zipfel, P.F. Dihydrolipoamide dehydrogenase of Pseudomonas aeruginosa is a surface-exposed immune evasion protein that binds three members of the factor H family and plasminogen. J. Immunol. 2012, 189, 4939–4950. [Google Scholar] [CrossRef] [Green Version]
- Dai, F.; Zhang, W.; Zhuang, Q.; Shao, Y.; Zhao, X.; Lv, Z.; Li, C. Dihydrolipoamide dehydrogenase of Vibrio splendidus is involved in adhesion to Apostichopus japonicus. Virulence 2019, 10, 839–848. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, X.; Ali, F.; Li, Z.; Fu, Y.; Yang, X.; Lin, W.; Lin, X. Comparative Extracellular Proteomics of Aeromonas hydrophila Reveals Iron-regulated secreted proteins as potential vaccine candidates. Front. Immunol. 2019, 10, 256. [Google Scholar] [CrossRef] [Green Version]
- Weir, H.; Chen, P.L.; Deiss, T.C.; Jacobs, N.; Nabity, M.B.; Young, M.; Criscitiello, M.F. DNP-KLH yields changes in leukocyte populations and immunoglobulin isotype use with different immunization routes in zebrafish. Front. Immunol. 2015, 6, 606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neely, M.N.; Pfeifer, J.D.; Caparon, M. Streptococcus-zebrafish model of bacterial pathogenesis. Infect. Immun. 2002, 70, 3904–3914. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Zhang, Y.; Ma, J.; Dong, W.; Zhong, X.; Pan, Z.; Yao, H. ICESsuHN105, a novel multiple antibiotic resistant ICE in Streptococcus suis serotype 5 strain HN105. Front. Microbiol. 2019, 10, 274. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, Y.; Dechao, D.; Yanfei, Y.; Zhang, W. Characterization and functional analysis of PnuC that is involved in the oxidative stress tolerance and virulence of Streptococcus suis serotype 2. Vet. Microbiol. 2018, 216, 198–206. [Google Scholar] [CrossRef]
- Vreman, S.; McCaffrey, J.; Popma-de Graaf, D.J.; Nauwynck, H.; Savelkoul, H.F.J.; Moore, A.; Rebel, J.M.J.; Stockhofe-Zurwieden, N. Toll-like receptor agonists as adjuvants for inactivated porcine reproductive and respiratory syndrome virus (PRRSV) vaccine. Vet. Immunol. Immunopathol. 2019, 212, 27–37. [Google Scholar] [CrossRef]
- Choi, J.H.; You, S.H.; Ko, M.K.; Jo, H.E.; Shin, S.H.; Jo, H.; Lee, M.J.; Kim, S.M.; Kim, B.; Lee, J.S.; et al. Improved immune responses and safety of foot-and-mouth disease vaccine containing immunostimulating components in pigs. J. Vet. Sci. 2020, 21, e74. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Vancott, J.L.; Okahashi, N.; Marinaro, M.; Kiyono, H.; Fujihashi, K.; Jackson, R.J.; Chatfield, S.N.; Bluethmann, H.; McGhee, J.R. The role of Th1 and Th2 cells for mucosal IgA responses. Ann. N. Y. Acad. Sci. 1996, 778, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A. Type 2 cytokines: Mechanisms and therapeutic strategies. Nat. Rev. Immunol. 2015, 15, 271–282. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′–3′) a | Restriction Site |
|---|---|---|
| P1 | GTAGGATCCGAACAGGTAACATCAGA | BamH I |
| P2 | GTACTCGAGCAAAGAGTAACGAATGTA | Sac I |
| P3 | CAGGATCCGTAGTTAAAGTTGGTA | BamH I |
| P4 | AAGAATTCTTTAGCGATTTTTGCG | EcoR I |
| P5 | ACGAATTCATCAAAGGTCGTAGCA | EcoR I |
| P6 | AACTCGAGGAACATCAAGAAAGGC | Sac I |
| P7 | GTAGGATCCGAACAGGTAACATCAGA | BamH I |
| P8 | TCCACCTCCACCTCCCAAAGAGTAACGAATGTA | |
| P9 | GGAGGTGGAGGTGGAGTAGTTAAAGTTGGTA | |
| P10 | TCCACCTCCACCTCCTTTAGCGATTTTTGCG | |
| P11 | GGAGGTGGAGGTGGAATCAAAGGTCGTAGCA | |
| P12 | AACTCGAGGAACATCAAGAAAGGC | Sac I |
| CFU/Fish | Zebrafish No. | Death No. | Mortality (100%) |
|---|---|---|---|
| 109 | 15 | 15 | 100 |
| 108 | 15 | 15 | 100 |
| 107 | 15 | 14 | 93.3 |
| 106 | 15 | 10 | 66.7 |
| 105 | 15 | 6 | 40 |
| 104 | 15 | 2 | 13.3 |
| PBS | 15 | 0 | 0 |
| LD50 | 2.36 × 105 CFU | ||
| CFU/Fish | Zebrafish No. | Death No. | Mortality (100%) |
|---|---|---|---|
| 109 | 15 | 15 | 100 |
| 108 | 15 | 15 | 100 |
| 107 | 15 | 14 | 93.3 |
| 106 | 15 | 8 | 53.3 |
| 105 | 15 | 3 | 20.0 |
| 104 | 15 | 1 | 6.7 |
| PBS | 15 | 0 | 0 |
| LD50 | 7.96 × 105 CFU | ||
| CFU/Fish | Zebrafish No. | Death No. | Mortality (100%) |
|---|---|---|---|
| 109 | 15 | 15 | 100 |
| 108 | 15 | 15 | 100 |
| 107 | 15 | 11 | 73.3 |
| 106 | 15 | 6 | 40.0 |
| 105 | 15 | 1 | 6.7 |
| 104 | 15 | 0 | 0 |
| PBS | 15 | 0 | 0 |
| LD50 | 2.00 × 106 CFU | ||
| Primer | Sequence (5′–3′) | Function |
|---|---|---|
| MIL − 6-F | TCCAGTTGCCTTCTTGGGAC | A fragment for murine IL − 6 gene |
| MIL − 6-R | GTGTAATTAAGCCTCCGACTTG | |
| MIL − 8-F | ATGGCTGCTGAACCAGTAGA | A fragment for murine IL − 8 gene |
| MIL − 8-R | CTAGTCTTCGTTTTGAACAG | |
| MIFN-γ-F | AGCTCCCAGAAACTGAACGA | A fragment for murine IFN-γ gene |
| MIFN-γ-R | AGGGTTCAAAGCATGAATGG | |
| PIL − 6-F | ACTGGCAGAAAACAACCTGA | A fragment for porcine IL − 6 gene |
| PIL − 6-R | CTAATCTGCACAGCCTCGAC | |
| PIL − 8-F | ATAAATACGCATTCCACACC | A fragment for porcine IL − 8 gene |
| PIL − 8-R | GTACAACCTTCTGCACCCA | |
| PIFN-γ-F | AGCCTCAATGACGACCTA | A fragment for porcine IFN-γ gene |
| PIFN-γ-R | ATCTTTGTTGGAGGGTGA | |
| MGAPDH-F | CCACAGTCCATGCCATCAC | A fragment for murine reference gene |
| MGAPDH-R | TCCACCACCCTGTTGCTGTA | |
| PGAPDH-F | GCTGGTGCTGAGTATGTCGT | A fragment for porcine reference gene |
| PGAPDH-R | AAGCAGTTGGTGGTACAGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Guo, M.; Kong, L.; Gao, Y.; Ma, J.; Cheng, Y.; Wang, H.; Yan, Y.; Sun, J. TLR4 Agonist Combined with Trivalent Protein JointS of Streptococcus suis Provides Immunological Protection in Animals. Vaccines 2021, 9, 184. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9020184
Wang Z, Guo M, Kong L, Gao Y, Ma J, Cheng Y, Wang H, Yan Y, Sun J. TLR4 Agonist Combined with Trivalent Protein JointS of Streptococcus suis Provides Immunological Protection in Animals. Vaccines. 2021; 9(2):184. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9020184
Chicago/Turabian StyleWang, Zhaofei, Mengting Guo, Licheng Kong, Ya Gao, Jingjiao Ma, Yuqiang Cheng, Henan Wang, Yaxian Yan, and Jianhe Sun. 2021. "TLR4 Agonist Combined with Trivalent Protein JointS of Streptococcus suis Provides Immunological Protection in Animals" Vaccines 9, no. 2: 184. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9020184