
Radiation-Inactivated Acinetobacter baumannii Vaccine Candidates

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Bacterial Growth Conditions
2.3. Formulation of Vaccine Candidates in MDP Prior to Irradiation
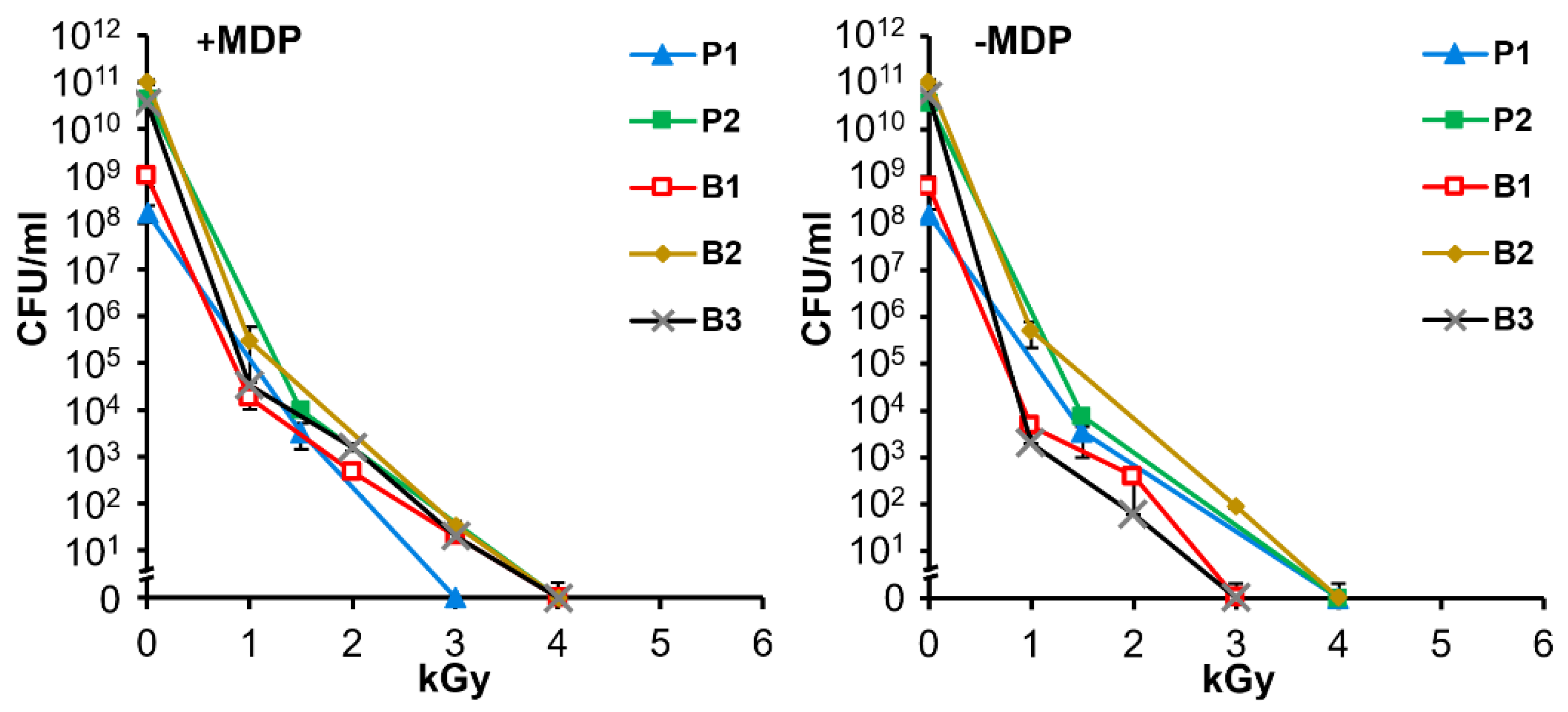
2.4. Cobalt-60 Irradiations and Inactivation Curves
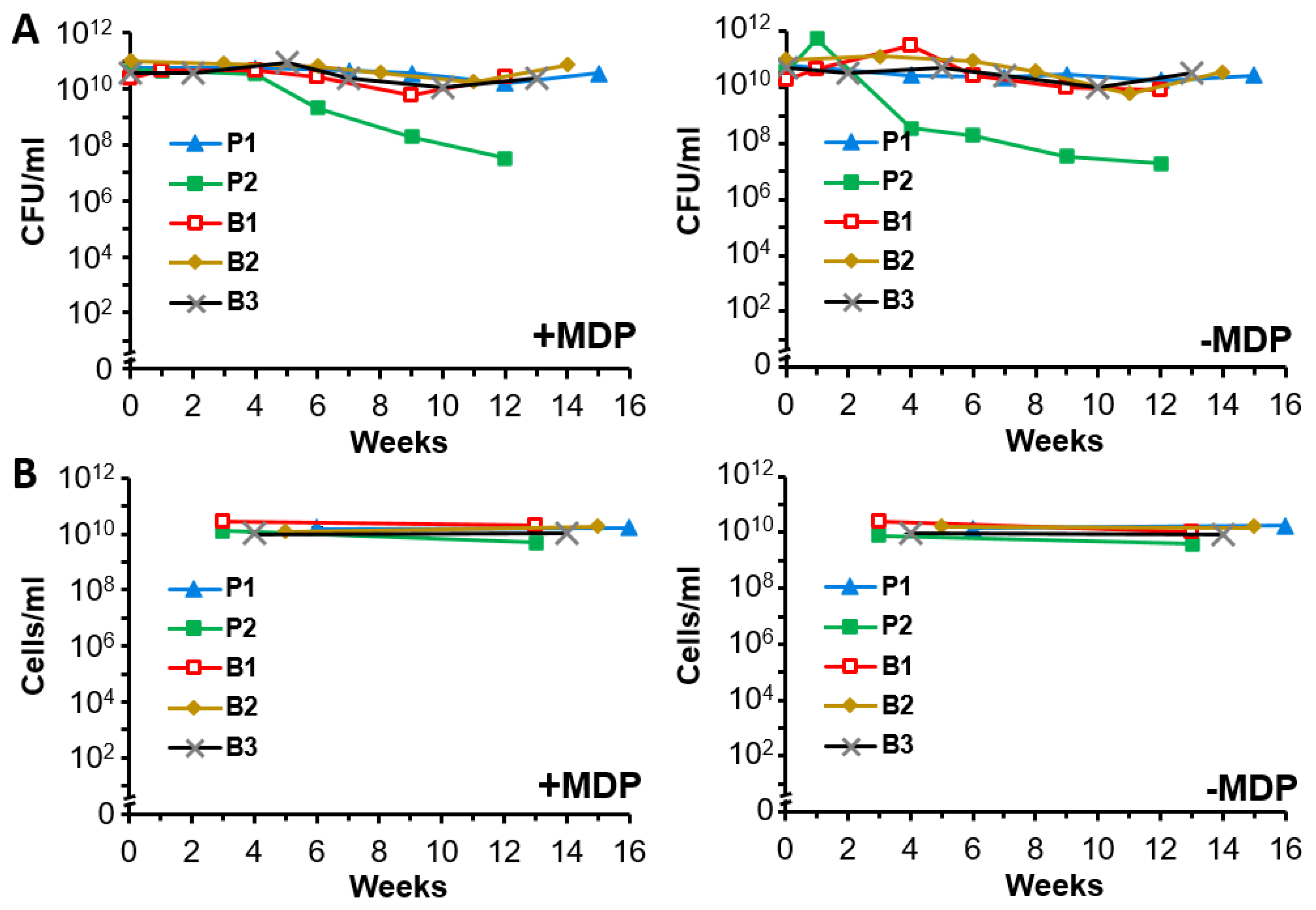
2.5. Stability Assays
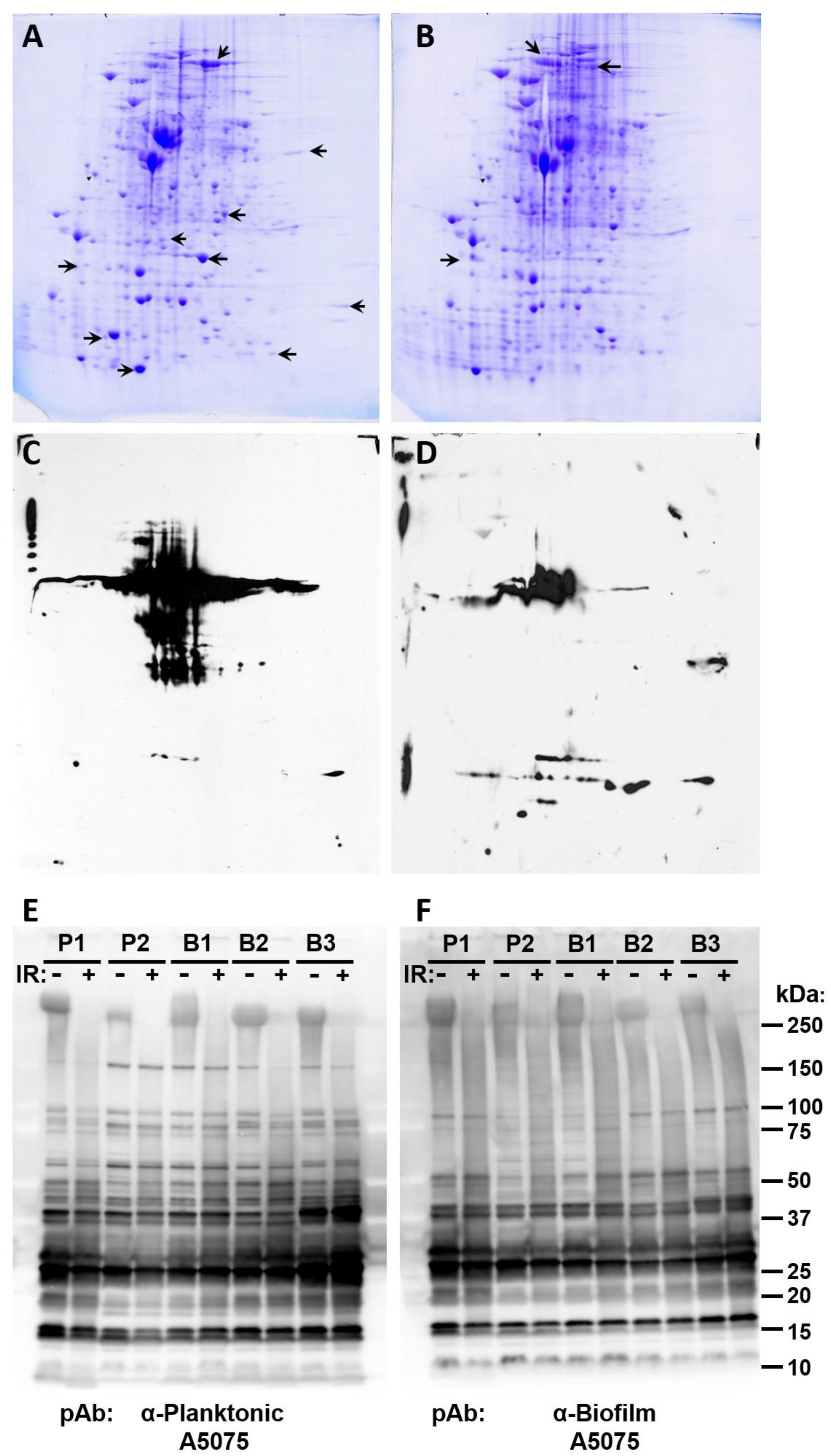
2.6. Coomassie-Stained Protein Gels
2.7. Two-Dimensional Electrophoresis
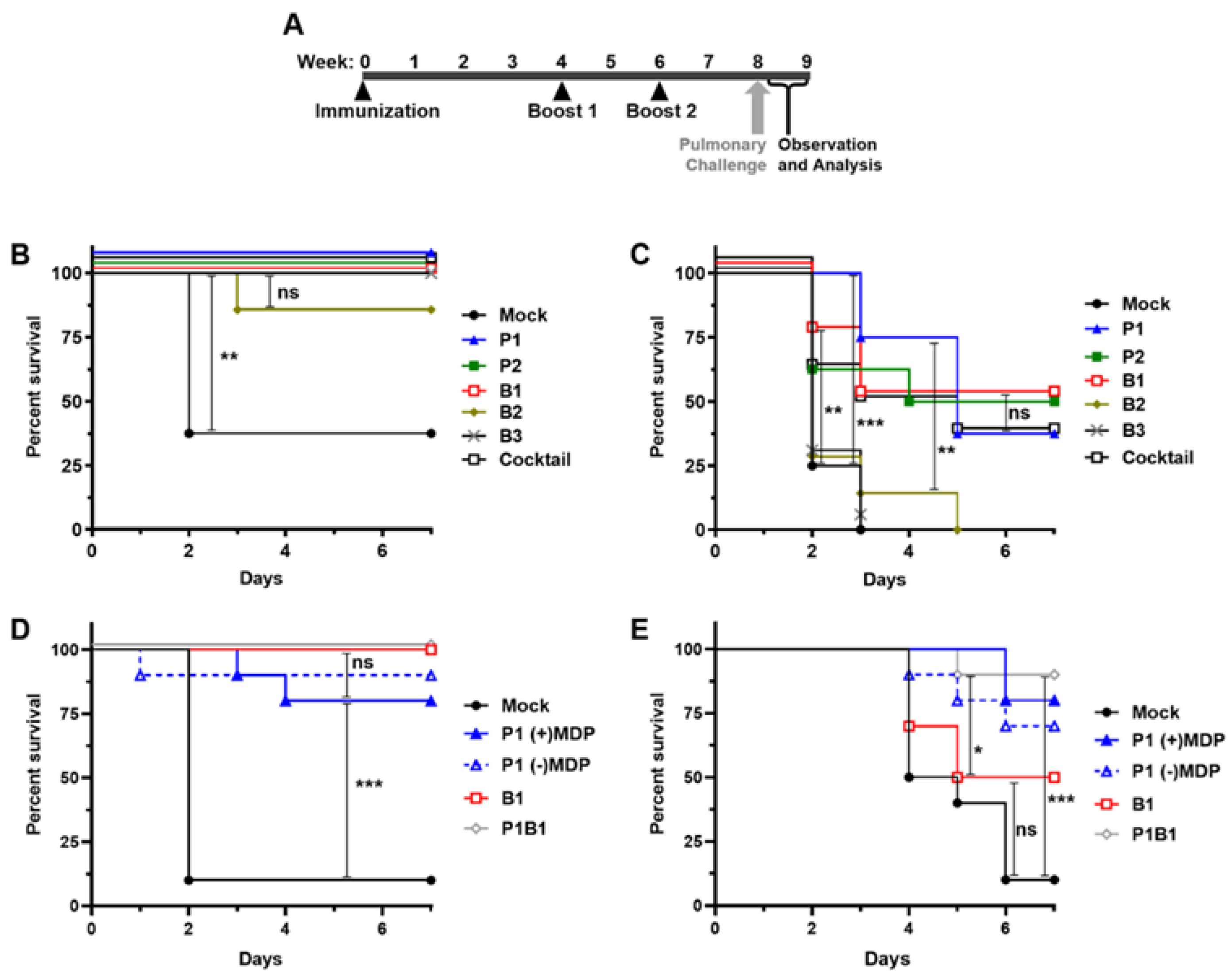
2.8. Challenge in the Murine Pulmonary Models
3. Results
3.1. Growth of A. baumannii Cultures with Differing Protein Expression Profiles
3.2. Antigenic Analysis of A. baumannii Cultures
3.3. Gamma Irradiation Inactivates Replication of Planktonic and Biofilm Cultures of A. baumannii in the Presence or Absence of the Protective MDP Complex
3.4. Irradiated and Non-Irradiated A. baumannii Cultures Display Stability for at Least 4 Months at 4 °C
3.5. Irradiated A. baumannii Vaccines Are Highly Protective in Healthy Mice
3.6. Select A. baumannii Vaccines Are Highly Protective in Neutropenic Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- WHO. Priority Pathogens List for R&D of New Antibiotics. Available online: https://www.testtargettreat.com/en/home/news-events/who_published_list_bacteria.html (accessed on 21 January 2021).
- CDC. Antibiotic Resistance Threats in the United States, 2019; 2019 AR Threats Report; U.S. Center for Disease Control: Atlanta, GA, USA, 2019.
- Lee, C.R.; Lee, J.H.; Park, M.; Park, K.S.; Bae, I.K.; Kim, Y.B.; Cha, C.J.; Jeong, B.C.; Lee, S.H. Biology of Acinetobacter baumannii: Pathogenesis, Antibiotic Resistance Mechanisms, and Prospective Treatment Options. Front. Cell. Infect. Microbiol. 2017, 7, 55. [Google Scholar] [CrossRef] [Green Version]
- Anderson, S.E.; Sherman, E.X.; Weiss, D.S.; Rather, P.N. Aminoglycoside Heteroresistance in Acinetobacter baumannii AB5075. mSphere 2018, 3, e00271-00218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, M.S.; Don, M.; Merkier, A.K.; Bistue, A.J.; Zorreguieta, A.; Centron, D.; Tolmasky, M.E. Naturally competent Acinetobacter baumannii clinical isolate as a convenient model for genetic studies. J. Clin. Microbiol. 2010, 48, 1488–1490. [Google Scholar] [CrossRef] [Green Version]
- Touchon, M.; Cury, J.; Yoon, E.J.; Krizova, L.; Cerqueira, G.C.; Murphy, C.; Feldgarden, M.; Wortman, J.; Clermont, D.; Lambert, T.; et al. The genomic diversification of the whole Acinetobacter genus: Origins, mechanisms, and consequences. Genome Biol. Evol. 2014, 6, 2866–2882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traglia, G.M.; Chua, K.; Centron, D.; Tolmasky, M.E.; Ramirez, M.S. Whole-genome sequence analysis of the naturally competent Acinetobacter baumannii clinical isolate A118. Genome Biol. Evol. 2014, 6, 2235–2239. [Google Scholar] [CrossRef] [Green Version]
- Spellberg, B.; Rex, J.H. The value of single-pathogen antibacterial agents. Nat. Rev. Drug Discov. 2013, 12, 963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lob, S.H.; Hoban, D.J.; Sahm, D.F.; Badal, R.E. Regional differences and trends in antimicrobial susceptibility of Acinetobacter baumannii. Int. J. Antimicrob. Agents 2016, 47, 317–323. [Google Scholar] [CrossRef] [PubMed]
- da Silva, K.E.; Maciel, W.G.; Croda, J.; Cayo, R.; Ramos, A.C.; de Sales, R.O.; Kurihara, M.N.L.; Vasconcelos, N.G.; Gales, A.C.; Simionatto, S. A high mortality rate associated with multidrug-resistant Acinetobacter baumannii ST79 and ST25 carrying OXA-23 in a Brazilian intensive care unit. PLoS ONE 2018, 13, e0209367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falagas, M.E.; Rafailidis, P.I. Attributable mortality of Acinetobacter baumannii: No longer a controversial issue. Crit. Care 2007, 11, 134. [Google Scholar] [CrossRef] [Green Version]
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: Emergence of a successful pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef] [Green Version]
- Harding, C.M.; Hennon, S.W.; Feldman, M.F. Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat. Rev. Microbiol. 2018, 16, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Lashinsky, J.N.; Henig, O.; Pogue, J.M.; Kaye, K.S. Minocycline for the Treatment of Multidrug and Extensively Drug-Resistant A. baumannii: A Review. Infect. Dis. Ther. 2017, 6, 199–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, T.; Wang, R.; Liu, D.; Walsh, T.R.; Zhang, R.; Lv, Y.; Ke, Y.; Ji, Q.; Wei, R.; Liu, Z.; et al. Emergence of plasmid-mediated high-level tigecycline resistance genes in animals and humans. Nat. Microbiol. 2019, 4, 1450–1456. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, T.B.; Pantapalangkoor, P.; Luna, B.M.; Bruhn, K.W.; Yan, J.; Dekitani, K.; Hsieh, S.; Yeshoua, B.; Pascual, B.; Vinogradov, E.; et al. Monoclonal Antibody Protects Against Acinetobacter baumannii Infection by Enhancing Bacterial Clearance and Evading Sepsis. J. Infect. Dis. 2017, 216, 489–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regeimbal, J.M.; Jacobs, A.C.; Corey, B.W.; Henry, M.S.; Thompson, M.G.; Pavlicek, R.L.; Quinones, J.; Hannah, R.M.; Ghebremedhin, M.; Crane, N.J.; et al. Personalized Therapeutic Cocktail of Wild Environmental Phages Rescues Mice from Acinetobacter baumannii Wound Infections. Antimicrob. Agents Chemother. 2016, 60, 5806–5816. [Google Scholar] [CrossRef] [Green Version]
- Lipsitch, M.; Siber, G.R. How Can Vaccines Contribute to Solving the Antimicrobial Resistance Problem? mBio 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Mishra, R.P.; Oviedo-Orta, E.; Prachi, P.; Rappuoli, R.; Bagnoli, F. Vaccines and antibiotic resistance. Curr. Opin. Microbiol. 2012, 15, 596–602. [Google Scholar] [CrossRef]
- WHO. Why Is Vaccination Important for Addressing Antibiotic Resistance? Available online: https://www.who.int/news-room/q-a-detail/why-is-vaccination-important-for-addressing-antibiotic-resistance (accessed on 18 December 2020).
- Gaidamakova, E.K.; Myles, I.A.; McDaniel, D.P.; Fowler, C.J.; Valdez, P.A.; Naik, S.; Gayen, M.; Gupta, P.; Sharma, A.; Glass, P.J.; et al. Preserving immunogenicity of lethally irradiated viral and bacterial vaccine epitopes using a radio- protective Mn2+-Peptide complex from Deinococcus. Cell Host Microbe 2012, 12, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Gayen, M.; Gupta, P.; Morazzani, E.M.; Gaidamakova, E.K.; Knollmann-Ritschel, B.; Daly, M.J.; Glass, P.J.; Maheshwari, R.K. Deinococcus Mn2+-peptide complex: A novel approach to alphavirus vaccine development. Vaccine 2017, 35, 3672–3681. [Google Scholar] [CrossRef]
- Tobin, G.J.; Tobin, J.K.; Gaidamakova, E.K.; Wiggins, T.J.; Bushnell, R.V.; Lee, W.M.; Matrosova, V.Y.; Dollery, S.J.; Meeks, H.N.; Kouiavskaia, D.; et al. A novel gamma radiation-inactivated sabin-based polio vaccine. PLoS ONE 2020, 15, e0228006. [Google Scholar] [CrossRef] [Green Version]
- Moore, H.N.; Kersten, H. Preliminary Note on the Preparation of Non-toxic Shiga Dysentery Vaccines by Irradiation with Soft X-rays. J. Med. Bacteriol. 1936, 31, 581–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dabral, N.; Martha Moreno, L.; Sriranganathan, N.; Vemulapalli, R. Oral immunization of mice with gamma-irradiated Brucella neotomae induces protection against intraperitoneal and intranasal challenge with virulent B. abortus 2308. PLoS ONE 2014, 9, e107180. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.S.; Bakken, R.R.; Lind, C.M.; Garcia, P.; Jenkins, E.; Glass, P.J.; Parker, M.D.; Hart, M.K.; Fine, D.L. Comparison of the immunological responses and efficacy of gamma-irradiated V3526 vaccine formulations against subcutaneous and aerosol challenge with Venezuelan equine encephalitis virus subtype IAB. Vaccine 2010, 28, 1031–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daly, M.J. A new perspective on radiation resistance based on Deinococcus radiodurans. Nat. Rev. Microbiol. 2009, 7, 237–245. [Google Scholar] [CrossRef]
- Bruckbauer, S.T.; Minkoff, B.B.; Yu, D.; Cryns, V.L.; Cox, M.M.; Sussman, M.R. Ionizing radiation-induced proteomic oxidation in Escherichia coli. Mol. Cell. Proteom. 2020, 19, 1375–1395. [Google Scholar] [CrossRef]
- Anderson, A.; Nordon, H.; Cain, R.F.; Parrish, G.; Duggan, D.; Nordan, H.; Parish, G.; Cullum-Dugan, D. Studies on a radio-resistant micrococcus. I. Isolation, morphology, cultural characteristics, and resistance to gamma radiation. Food Technol. 1956, 10, 575–578. [Google Scholar]
- Daly, M.J.; Gaidamakova, E.K.; Matrosova, V.Y.; Kiang, J.G.; Fukumoto, R.; Lee, D.Y.; Wehr, N.B.; Viteri, G.A.; Berlett, B.S.; Levine, R.L. Small-molecule antioxidant proteome-shields in Deinococcus radiodurans. PLoS ONE 2010, 5, e12570. [Google Scholar] [CrossRef]
- Daly, M.J.; Gaidamakova, E.K.; Matrosova, V.Y.; Vasilenko, A.; Zhai, M.; Venkateswaran, A.; Hess, M.; Omelchenko, M.V.; Kostandarithes, H.M.; Makarova, K.S.; et al. Accumulation of Mn(II) in Deinococcus radiodurans facilitates gamma-radiation resistance. Science 2004, 306, 1025–1028. [Google Scholar] [CrossRef] [Green Version]
- Daly, M.J. Death by protein damage in irradiated cells. DNA Repair 2012, 11, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Gaidamakova, E.K.; Grichenko, O.; Matrosova, V.Y.; Hoeke, V.; Klimenkova, P.; Conze, I.H.; Volpe, R.P.; Tkavc, R.; Gostinčar, C.; et al. Across the tree of life, radiation resistance is governed by antioxidant Mn2+, gauged by paramagnetic resonance. Proc. Natl. Acad. Sci. USA 2017, 114, E9253–E9260. [Google Scholar] [CrossRef] [Green Version]
- Peana, M.; Gumienna-Kontecka, E.; Piras, F.; Ostrowska, M.; Piasta, K.; Krzywoszynska, K.; Medici, S.; Zoroddu, M.A. Exploring the Specificity of Rationally Designed Peptides Reconstituted from the Cell-Free Extract of Deinococcus radiodurans toward Mn(II) and Cu(II). Inorg. Chem. 2020, 59, 4661–4684. [Google Scholar] [CrossRef] [PubMed]
- Rumbo-Feal, S.; Gomez, M.J.; Gayoso, C.; Alvarez-Fraga, L.; Cabral, M.P.; Aransay, A.M.; Rodriguez-Ezpeleta, N.; Fullaondo, A.; Valle, J.; Tomas, M.; et al. Whole transcriptome analysis of Acinetobacter baumannii assessed by RNA-sequencing reveals different mRNA expression profiles in biofilm compared to planktonic cells. PLoS ONE 2013, 8, e72968. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.L.; Neu, H.M.; Gilbreath, J.J.; Michel, S.L.J.; Zurawski, D.V.; Merrell, D.S. Copper Resistance of the Emerging Pathogen Acinetobacter baumannii. Appl. Environ. Microbiol. 2016, 82, 6174–6188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, A.C.; Thompson, M.G.; Black, C.C.; Kessler, J.L.; Clark, L.P.; McQueary, C.N.; Gancz, H.Y.; Corey, B.W.; Moon, J.K.; Si, Y.; et al. AB5075, a Highly Virulent Isolate of Acinetobacter baumannii, as a Model Strain for the Evaluation of Pathogenesis and Antimicrobial Treatments. mBio 2014, 5, e01076-14. [Google Scholar] [CrossRef] [Green Version]
- Chin, C.Y.; Tipton, K.A.; Farokhyfar, M.; Burd, E.M.; Weiss, D.S.; Rather, P.N. A high-frequency phenotypic switch links bacterial virulence and environmental survival in Acinetobacter baumannii. Nat. Microbiol. 2018, 3, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.G.; Black, C.C.; Pavlicek, R.L.; Honnold, C.L.; Wise, M.C.; Alamneh, Y.A.; Moon, J.K.; Kessler, J.L.; Si, Y.; Williams, R.; et al. Validation of a novel murine wound model of Acinetobacter baumannii infection. Antimicrob. Agents Chemother. 2014, 58, 1332–1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barki, K.G.; Das, A.; Dixith, S.; Ghatak, P.D.; Mathew-Steiner, S.; Schwab, E.; Khanna, S.; Wozniak, D.J.; Roy, S.; Sen, C.K. Electric Field Based Dressing Disrupts Mixed-Species Bacterial Biofilm Infection and Restores Functional Wound Healing. Ann. Surg. 2019, 269, 756–766. [Google Scholar] [CrossRef] [PubMed]
- Di Venanzio, G.; Flores-Mireles, A.L.; Calix, J.J.; Haurat, M.F.; Scott, N.E.; Palmer, L.D.; Potter, R.F.; Hibbing, M.E.; Friedman, L.; Wang, B.; et al. Urinary tract colonization is enhanced by a plasmid that regulates uropathogenic Acinetobacter baumannii chromosomal genes. Nat. Commun. 2019, 10, 2763. [Google Scholar] [CrossRef] [PubMed]
- de Breij, A.; Dijkshoorn, L.; Lagendijk, E.; van der Meer, J.; Koster, A.; Bloemberg, G.; Wolterbeek, R.; van den Broek, P.; Nibbering, P. Do biofilm formation and interactions with human cells explain the clinical success of Acinetobacter baumannii? PLoS ONE 2010, 5, e10732. [Google Scholar] [CrossRef]
- Gaddy, J.A.; Arivett, B.A.; McConnell, M.J.; López-Rojas, R.; Pachón, J.; Actis, L.A. Role of acinetobactin-mediated iron acquisition functions in the interaction of Acinetobacter baumannii strain ATCC 19606T with human lung epithelial cells, Galleria mellonella caterpillars, and mice. Infect. Immun. 2012, 80, 1015–1024. [Google Scholar] [CrossRef] [Green Version]
- Eijkelkamp, B.A.; Stroeher, U.H.; Hassan, K.A.; Papadimitrious, M.S.; Paulsen, I.T.; Brown, M.H. Adherence and motility characteristics of clinical Acinetobacter baumannii isolates. FEMS Microbiol. Lett. 2011, 323, 44–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvarez-Fraga, L.; Pérez, A.; Rumbo-Feal, S.; Merino, M.; Vallejo, J.A.; Ohneck, E.J.; Edelmann, R.E.; Beceiro, A.; Vázquez-Ucha, J.C.; Valle, J.; et al. Analysis of the role of the LH92_11085 gene of a biofilm hyper-producing Acinetobacter baumannii strain on biofilm formation and attachment to eukaryotic cells. Virulence 2016, 7, 443–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Guitián, M.; Vázquez-Ucha, J.C.; Álvarez-Fraga, L.; Conde-Pérez, K.; Vallejo, J.A.; Perina, A.; Bou, G.; Poza, M.; Beceiro, A. Global Transcriptomic Analysis During Murine Pneumonia Infection Reveals New Virulence Factors in Acinetobacter baumannii. J. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Harro, J.M.; Achermann, Y.; Freiberg, J.A.; Allison, D.L.; Brao, K.J.; Marinos, D.P.; Sanjari, S.; Leid, J.G.; Shirtliff, M.E. Clearance of Staphylococcus aureus from In Vivo Models of Chronic Infection by Immunization Requires Both Planktonic and Biofilm Antigens. Infect. Immun. 2019, 88. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.K.; Okamoto, S.; Hayashi, T.; Shin, S.S.; Mihajlov, I.; Fermin, A.; Guiney, D.G.; Fierer, J.; Raz, E. Vaccination with irradiated Listeria induces protective T cell immunity. Immunity 2006, 25, 143–152. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture Designation | Growth Phase | Nutritional Source | Time of Harvest | Growth Platform |
|---|---|---|---|---|
| P1 | Planktonic | TSB | Overnight | Agitated Broth |
| P2 | Planktonic | TSB | 2 h | Agitated Broth |
| B1 | (Submerged) Biofilm | M9 | 2 days | Stationary Broth |
| B2 | (Colony) Biofilm | M9 | Overnight | Stationary Agar |
| B3 | (Colony) Biofilm | TSA + SB | Overnight | Stationary Agar |
| Culture Designation: | P1 | P2 | B1 | B2 | B3 |
|---|---|---|---|---|---|
| 150 | 150 | 150 | |||
| 130 | 130 | ||||
| 115 | 115 | ||||
| Unique/Distinct Bands kDa: | 80 | 80 | 80 | ||
| 75 | 75 | ||||
| 65 | |||||
| 40 | 40 | ||||
| 22 | 22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dollery, S.J.; Zurawski, D.V.; Gaidamakova, E.K.; Matrosova, V.Y.; Tobin, J.K.; Wiggins, T.J.; Bushnell, R.V.; MacLeod, D.A.; Alamneh, Y.A.; Abu-Taleb, R.; et al. Radiation-Inactivated Acinetobacter baumannii Vaccine Candidates. Vaccines 2021, 9, 96. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9020096
Dollery SJ, Zurawski DV, Gaidamakova EK, Matrosova VY, Tobin JK, Wiggins TJ, Bushnell RV, MacLeod DA, Alamneh YA, Abu-Taleb R, et al. Radiation-Inactivated Acinetobacter baumannii Vaccine Candidates. Vaccines. 2021; 9(2):96. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9020096
Chicago/Turabian StyleDollery, Stephen J., Daniel V. Zurawski, Elena K. Gaidamakova, Vera Y. Matrosova, John K. Tobin, Taralyn J. Wiggins, Ruth V. Bushnell, David A. MacLeod, Yonas A. Alamneh, Rania Abu-Taleb, and et al. 2021. "Radiation-Inactivated Acinetobacter baumannii Vaccine Candidates" Vaccines 9, no. 2: 96. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9020096