
HCV Core/NS3 Protein Immunization with “N-Terminal Heat Shock gp96 Protein (rNT (gp96))” Induced Strong and Sustained Th1-Type Cytokines in Immunized Mice

 and
and
Abstract
:1. Introduction
2. Materials and Methods
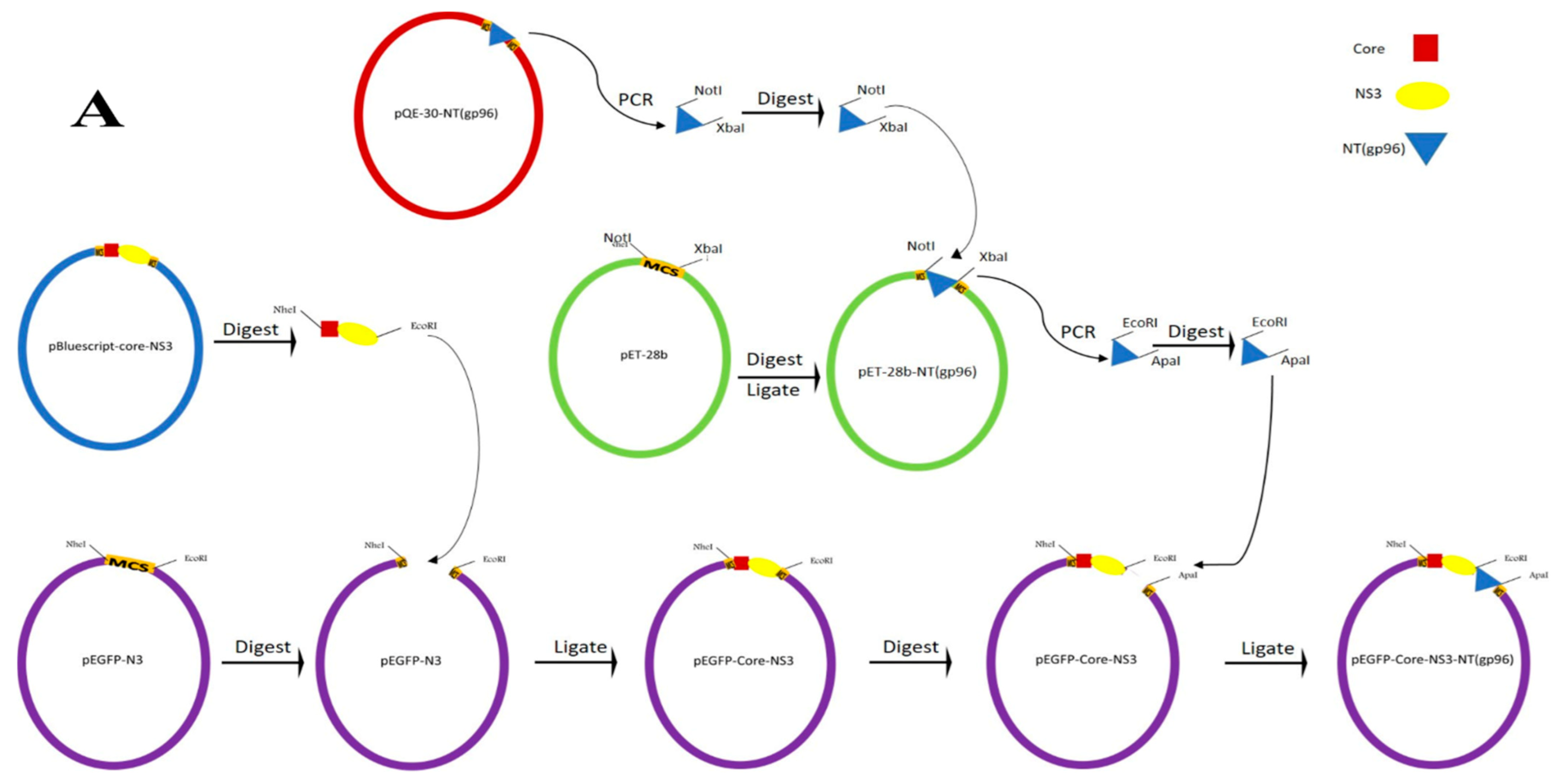
2.1. Constructs of the Expression Vector and Purification of Proteins
2.1.1. Construction of Recombinant Expression Vectors
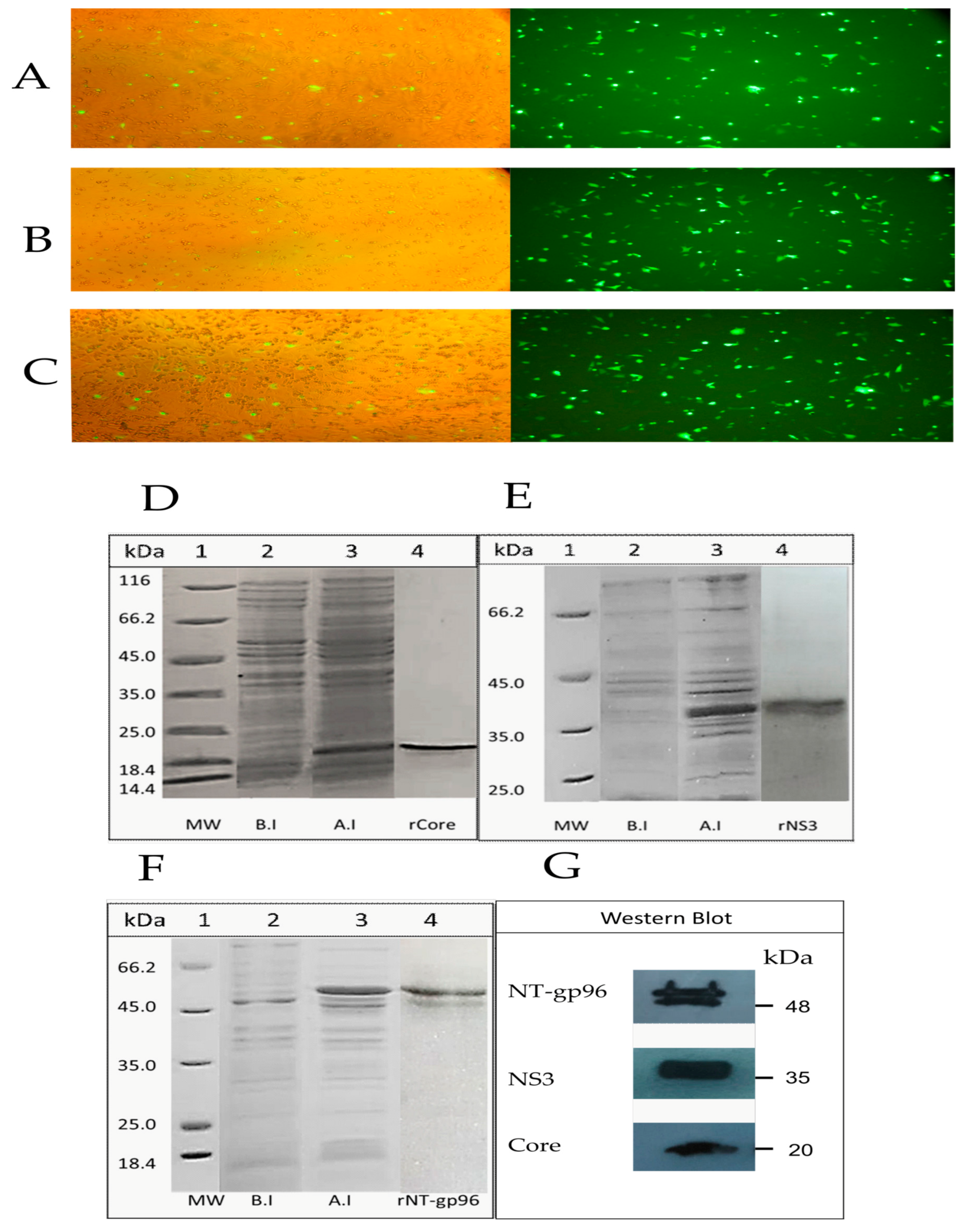
2.1.2. Expression and Purification of the Recombinant Proteins
2.1.3. Mice and Vaccination Regimens
2.2. Cytokine Assay
2.3. Antigen–Specific Antibody Responses
2.4. Statistical Analysis
3. Results
3.1. Analyses on Expression of the Recombinant Core, NS3, and NT (gp96) Proteins
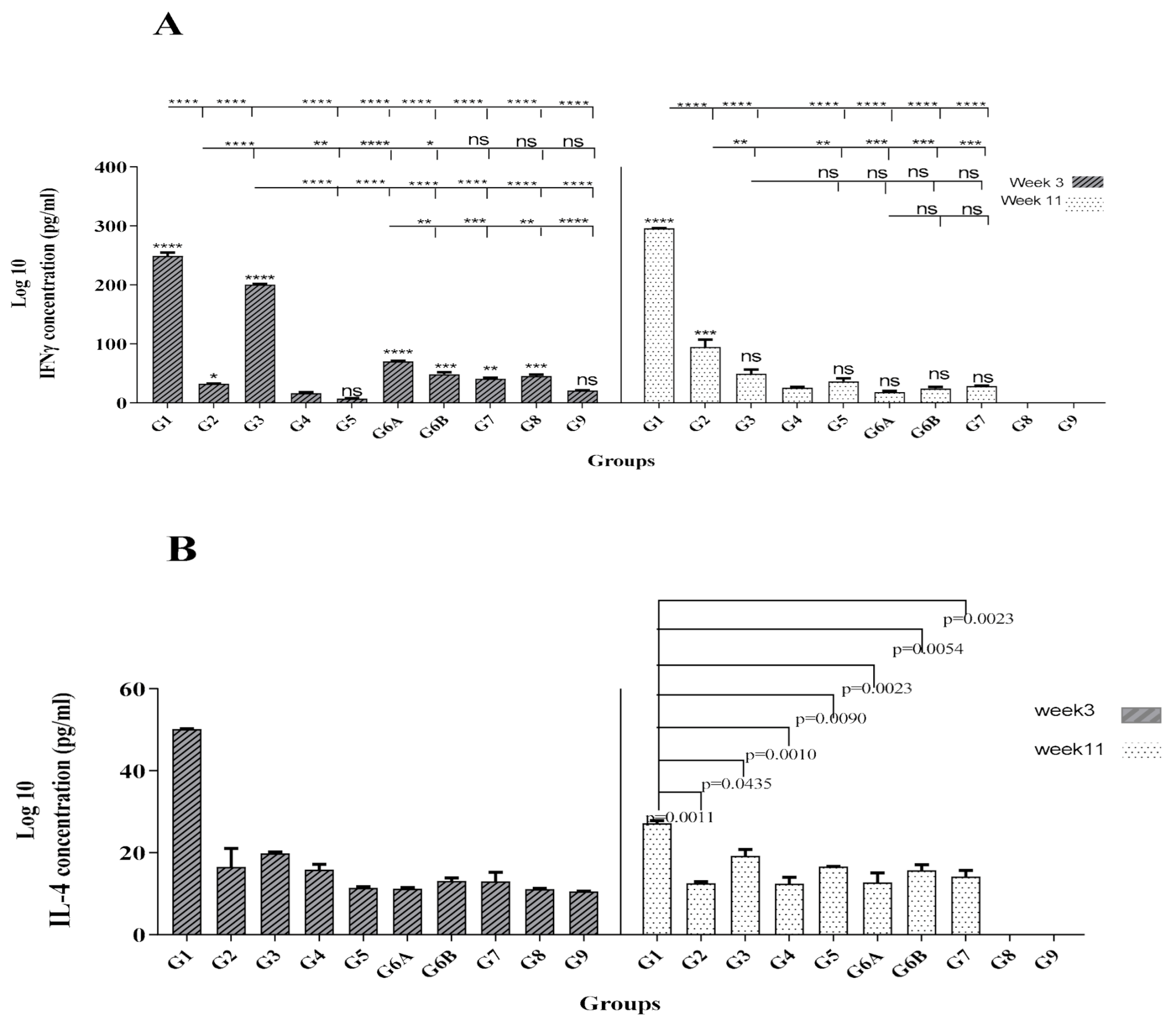
3.2. Protein/Protein + rNT (gp96) Vaccination Induced Strong and Long-Term Th1 Cytokine Profile
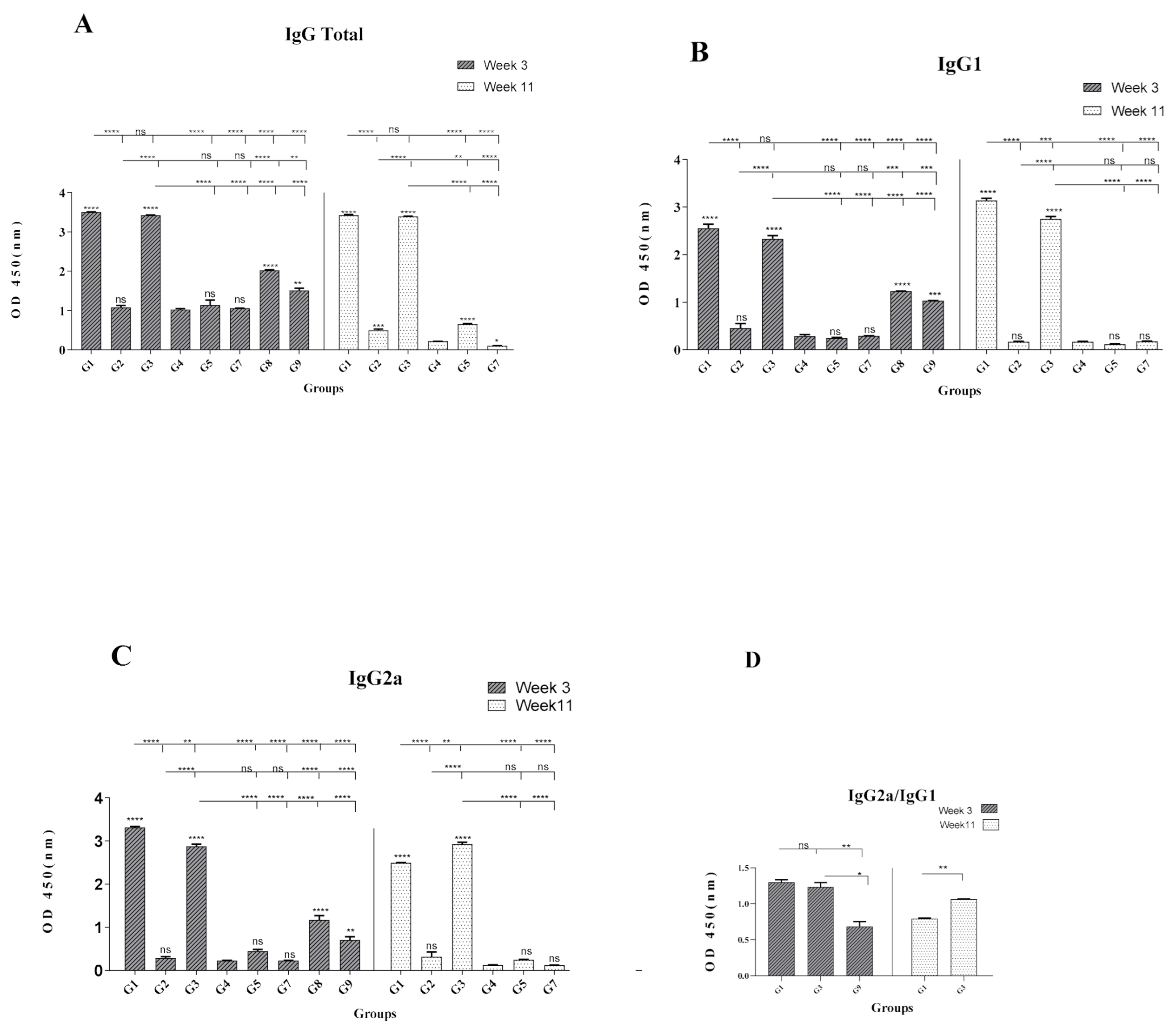
3.3. Formulation of HCV Core-NS3 Proteins with rNT(gp96) Elicited Th1-Phenotype Antibody Responses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Suwanthawornkul, T.; Anothaisintawee, T.; Sobhonslidsuk, A.; Thakkinstian, A.; Teerawattananon, Y. Efficacy of second gen-eration direct-acting antiviral agents for treatment naïve hepatitis C genotype 1: A systematic review and network meta-analysis. PLoS ONE 2015, 10, e0145953. [Google Scholar] [CrossRef]
- World Health Organization. Global Hepatitis Report 2017; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Gummow, J.; Masavuli, M.G.; Mekonnen, Z.A.; Li, Y.; Wijesundara, D.K.; Shrestha, A.C.; Voskoboinik, I.; Gowans, E.J.; Grubor-Bauk, B. Safety Profile of a Multi-Antigenic DNA Vaccine Against Hepatitis C Virus. Vaccines 2020, 8, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, A.L. Global control of hepatitis C virus. Science 2015, 349, 790–791. [Google Scholar] [CrossRef]
- Masavuli, M.G.; Wijesundara, D.K.; Underwood, A.; Christiansen, D.; Earnest-Silveira, L.; Bull, R.; Torresi, J.; Gowans, E.J.; Grubor-Bauk, B. A hepatitis C virus DNA vaccine encoding a secreted, oligomerized form of envelope proteins is highly immu-nogenic and elicits neutralizing antibodies in vaccinated mice. Front. Immunol. 2009, 10, 1145. [Google Scholar] [CrossRef] [Green Version]
- Conti, F.; Buonfiglioli, F.; Scuteri, A.; Crespi, C.; Bolondi, L.; Caraceni, P.; Foschi, F.G.; Lenzi, M.; Mazzella, G.; Verucchi, G.; et al. Early occurrence and recurrence of hepatocellular carcinoma in HCV-related cirrhosis treated with direct-acting antivirals. J. Hepatol. 2016, 65, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Tabata, K.; Neufeldt, C.J.; Bartenschlager, R. Hepatitis C Virus Replication. Cold Spring Harb. Perspect. Med. 2019, 10, a037093. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.R.; Barnes, E.; Cox, A.L. Approaches, Progress, and Challenges to Hepatitis C Vaccine Development. Gastroenterology 2019, 156, 418–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ugarte, S.O.; Perez, A.; Falcon, V.; Urquiza, D.; Pichardo, D.; Martinez-Donato, G. Protective cellular immune response against hepatitis C virus elicited by chimeric protein formulations in BALB/c mice. Arch. Virol. 2020, 165, 593–607. [Google Scholar] [CrossRef]
- Donnison, T.; Von Delft, A.; Brown, A.; Swadling, L.; Hutchings, C.; Hanke, T.; Chinnakannan, S.; Barnes, E. Viral vectored hepatitis C virus vaccines generate pan-genotypic T cell responses to conserved subdominant epitopes. Vaccine 2020, 38, 5036–5048. [Google Scholar] [CrossRef]
- Logvinoff, C.; Major, M.E.; Oldach, D.; Heyward, S.; Talal, A.; Balfe, P.; Feinstone, S.M.; Alter, H.; Rice, C.M.; McKeating, J.A. Neutralizing antibody response during acute and chronic hepatitis C virus infection. Proc. Natl. Acad. Sci. USA 2004, 101, 10149–10154. [Google Scholar] [CrossRef] [Green Version]
- Semmo, N.; Lucas, M.; Krashias, G.; Lauer, G.; Chapel, H.; Klenerman, P. Maintenance of HCV-specific T-cell responses in anti-body-deficient patients a decade after early therapy. Blood 2006, 107, 4570–4571. [Google Scholar] [CrossRef] [Green Version]
- Fauvelle, C.; Colpitts, C.C.; Keck, Z.-Y.; Pierce, B.G.; Foung, S.K.H.; Baumert, T.F. Hepatitis C virus vaccine candidates inducing protective neutralizing antibodies. Expert Rev. Vaccines 2016, 15, 1535–1544. [Google Scholar] [CrossRef]
- Samimi-Rad, K.; Sadeghi, F.; Amirzargar, A.; Eshraghian, M.R.; Alavian, S.-M.; Rahimnia, R. Association of HLA class II alleles with hepatitis C virus clearance and persistence in thalassemia patients from Iran. J. Med. Virol. 2015, 87, 1565–1572. [Google Scholar] [CrossRef] [PubMed]
- Habersetzer, F.; Baumert, T.F.; Stoll-Keller, F. Gl-5OO5, a yeast vector vaccine expressing an NS3-core fusion protein. Curr. Opin. Mol. Ther. 2009, 11, 456–462. [Google Scholar] [PubMed]
- Wiesch, J.S.Z.; Lauer, G.M.; Day, C.L.; Kim, A.Y.; Ouchi, K.; Duncan, J.E.; Wurcel, A.G.; Timm, J.; Jones, A.M.; Mothe, B.; et al. Broad Repertoire of the CD4+Th Cell Response in Spontaneously Controlled Hepatitis C Virus Infection Includes Dominant and Highly Promiscuous Epitopes. J. Immunol. 2005, 175, 3603–3613. [Google Scholar] [CrossRef] [Green Version]
- Bastola, R.; Noh, G.; Keum, T.; Bashyal, S.; Seo, J.-E.; Choi, J.; Oh, Y.; Cho, Y.; Lee, S. Vaccine adjuvants: Smart components to boost the immune system. Arch. Pharmacal. Res. 2017, 40, 1238–1248. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Kwon, T.; Polo, J.; Zhu, Y.-F.; Coates, S.; Crawford, K.; Dong, C.; Wininger, M.; Hall, J.; Selby, M. Induction of broad CD4+ and CD8+ T-cell responses and cross-neutralizing antibodies against hepatitis C virus by vaccination with Th1-adjuvanted poly-peptides followed by defective alphaviral particles expressing envelope glycoproteins gpE1 and gpE2 and nonstructural pro-teins 3, 4, and 5. J. Virol. 2008, 82, 7492–7503. [Google Scholar]
- Manickan, E.; Karem, K.L.; Rouse, B.T. DNA Vaccines–A Modern Gimmick or a Boon to Vaccinology. Crit. Rev. Immunol. 2017, 37, 139–154. [Google Scholar] [CrossRef]
- Klinman, D.M. CpG DNA as a vaccine adjuvant. Expert Rev. Vaccines 2003, 2, 305–315. [Google Scholar] [CrossRef]
- Alekseeva, E.; Sominskaya, I.; Skrastina, D.; Egorova, I.; Starodubova, E.; Kushners, E.; Mihailova, M.; Petrakova, N.; Bruvere, R.; Kozlovskaya, T.; et al. Enhancement of the expression of HCV core gene does not enhance core-specific immune response in DNA immunization: Advantages of the heterologous DNA prime, protein boost immunization regimen. Genet. Vaccines Ther. 2009, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Dale, C.J.; Thomson, S.; De Rose, R.; Ranasinghe, C.; Medveczky, C.J.; Pamungkas, J.; Boyle, D.B.; Ramshaw, I.A.; Kent, S.J. Prime-Boost Strategies in DNA Vaccines. DNA Vaccines 2006, 127, 171–198. [Google Scholar] [CrossRef]
- Srivastava, P. Interaction of heat shock proteins with peptides and antigen presenting cells: Chaperoning of the innate and adaptive immune responses. Annu. Rev. Immunol. 2002, 20, 395–425. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Qiu, L.; Liu, G.; Li, Y.; Zhang, X.; Jin, W.; Gao, G.F.; Kong, X.; Meng, S. Heat shock protein gp96 enhances humoral and T cell responses, decreases Treg frequency and potentiates the anti-HBV activity in BALB/c and transgenic mice. Vaccine 2011, 29, 6342–6351. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Li, J.; Bi, Y.; Jia, X.; Meng, S.; Sun, L.; Liu, W. Gp96 enhances the immunogenicity of subunit vaccine of porcine repro-ductive and respiratory syndrome virus. Virus Res. 2012, 167, 162–172. [Google Scholar] [CrossRef]
- Ju, Y.; Fan, H.; Liu, J.; Hu, J.; Li, X.; Li, C.; Chen, L.; Gao, Q.; Gao, G.F.; Meng, S. Heat shock protein gp96 adjuvant induces T cell responses and cross-protection to a split influenza vaccine. Vaccine 2014, 32, 2703–2711. [Google Scholar] [CrossRef] [PubMed]
- Pishraft-Sabet, L.; Kosinska, A.D.; Rafati, S.; Bolhassani, A.; Taheri, T.; Memarnejadian, A.; Alavian, S.-M.; Roggendorf, M.; Samimi-Rad, K. Enhancement of HCV polytope DNA vaccine efficacy by fusion to an N-terminal fragment of heat shock protein gp96. Arch. Virol. 2014, 160, 141–152. [Google Scholar] [CrossRef]
- Sugauchi, F.; Wang, R.Y.-H.; Qiu, Q.; Jin, B.; Alter, H.J.; Shih, J.W. Vigorous Hepatitis C Virus—Specific CD4+ and CD8+ T Cell Responses Induced by Protein Immunization in the Presence of Montanide ISA720 plus Synthetic Oligodeoxynucleotides Containing Immunostirnulatory Cytosine-Guanine Dinucleotide Motifs. J. Infect. Dis. 2006, 193, 563–572. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Q.; Wang, R.Y.-H.; Jiao, X.; Jin, B.; Sugauchi, F.; Grandinetti, T.; Alter, H.J.; Shih, J.W.-K. Induction of multispecific Th-1 type immune response against HCV in mice by protein immunization using CpG and Montanide ISA 720 as adjuvants. Vaccine 2008, 26, 5527–5534. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Russell, D.W. The Condensed Protocols from Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2006. [Google Scholar]
- Ansari, N.; Rafati, S.; Taheri, T.; Roohvand, F.; Farahmand, M.; Hajikhezri, Z.; Keshavarz, A.; Samimi-Rad, K.; Katayoun, S.-R. A non-pathogenic Leishmania tarentolae vector based- HCV polytope DNA vaccine elicits potent and long lasting Th1 and CTL responses in BALB/c mice model. Mol. Immunol. 2019, 111, 152–161. [Google Scholar] [CrossRef]
- Takeuchi, A.; Badr, M.E.S.G.; Miyauchi, K.; Ishihara, C.; Onishi, R.; Guo, Z.; Sasaki, Y.; Ike, H.; Takumi, A.; Tsuji, N.M.; et al. CRTAM determines the CD4+ cytotoxic T lymphocyte lineage. J. Exp. Med. 2016, 213, 123–138. [Google Scholar] [CrossRef]
- Bolhassani, A.; Zahedifard, F.; Taghikhani, M.; Rafati, S. Enhanced immunogenicity of HPV16E7 accompanied by Gp96 as an adjuvant in two vaccination strategies. Vaccine 2008, 26, 3362–3370. [Google Scholar] [CrossRef] [PubMed]
- Fernando, G.; Stewart, T.J.; Tindle, R.W.; Frazer, I.H. Th2-type CD4+ cells neither enhance nor suppress antitumor CTL activity in a mouse tumor model. J. Immunol. 1998, 161, 2421–2427. [Google Scholar] [PubMed]
- Garmory, H.S.; Brown, K.A.; Titball, R.W. DNA vaccines: Improving expression of antigens. Genet. Vaccines Ther. 2003, 1, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geissler, M.; Gesien, A.; Tokushige, K.; Wands, J.R. Enhancement of cellular and humoral immune responses to hepatitis C virus core protein using DNA-based vaccines augmented with cytokine-expressing plasmids. J. Immunol. 1997, 158, 1231–1237. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Sequence |
|---|---|
| Forward Kgp1 with NotI site (underlined) | 5′ATA TGC GGC CGC GAA GAT GAC GTT G3′ |
| Reverse Kgp2 with stop codon (bold) and XbaI site (underlined) | 5′GGG CTC TCT AGA TTA TTT GTA GAA GGC TTT G3′ |
| Forward KZ1 with EcoRI site (underlined) | 5′TCC GAA TTC GAA GAT GAC GTT GAA GTG3′ |
| Reverse KZ2 with ApaI site (underlined) | 5′CTC GGG CCC ATTTT GTA GAA GGC3′ |
| Reverse KZ3 with stop codon (bold) and ApaI site (underlined) | 5′CTC GGG CCC CTA TTT GTA GAA GGC3′ |
| Forward ZC1 with NcoI and NheI sites (underlined) | 5′ATC GAT GCT AGC GAG CTC ACC A3′ |
| Reverse ZC2 with BamH I site (underlined) | 5′CAA CAG CGG ACC GGA TCC AAG C3′ |
| Forward ZN1 with Nhe I and BamH I sites (underlined) | 5′CTT GCT AGC GGA TCC GGT CCG CTG TTG3′ |
| Reverse ZN2 with NotI sites (underlined) | 5′GTG GCG GCC GCT TAT CTA GAC TGC AG3′ |
| Groups | Prime 1st Week | Boost 4th Week | Boost 7th Week | Modeling |
|---|---|---|---|---|
| G1 | Core-NS3 + NT (gp96) | Core-NS3 + NT (gp96) | Protein/protein | |
| G2 | pcDNA-Core-NS3-NT (gp96) | pcDNA-Core-NS3-NT (gp96) | pcDNA-Core-NS3-NT (gp96) | DNA/DNA |
| G3 | pcDNA-Core-NS3-NT (gp96) | Core-NS3 + NT (gp96) | Core-NS3 + NT (gp96) | DNA/protein |
| G4 | PBS | PBS | PBS | Control |
| G5 | pcDNA-Core-NS3 | pcDNA-Core-NS3 | pcDNA-Core-NS3 | Control |
| G6A | Core-NS3 + CpG ODN | Core-NS3 + CpG ODN | protein/protein | |
| G6B | Core-NS3 + non-CpG | Core-NS3 + non-CpG | Control | |
| G7 | pcDNA3.1(+) | pcDNA3.1(+) | pcDNA3.1(+) | |
| G8 | NT (gp96) protein | NT (gp96) protein | Control | |
| G9 | Core-Ns3 protein | Core-Ns3 protein | Control |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hajikhezri, Z.; Roohvand, F.; Maleki, M.; Shahmahmoodi, S.; Amirzargar, A.A.; Keshavarz, A.; Seyed, N.; Farahmand, M.; Samimi-Rad, K. HCV Core/NS3 Protein Immunization with “N-Terminal Heat Shock gp96 Protein (rNT (gp96))” Induced Strong and Sustained Th1-Type Cytokines in Immunized Mice. Vaccines 2021, 9, 215. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9030215
Hajikhezri Z, Roohvand F, Maleki M, Shahmahmoodi S, Amirzargar AA, Keshavarz A, Seyed N, Farahmand M, Samimi-Rad K. HCV Core/NS3 Protein Immunization with “N-Terminal Heat Shock gp96 Protein (rNT (gp96))” Induced Strong and Sustained Th1-Type Cytokines in Immunized Mice. Vaccines. 2021; 9(3):215. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9030215
Chicago/Turabian StyleHajikhezri, Zamaneh, Farzin Roohvand, Monireh Maleki, Shohreh Shahmahmoodi, Ali Akbar Amirzargar, Abolfazl Keshavarz, Negar Seyed, Mohammad Farahmand, and Katayoun Samimi-Rad. 2021. "HCV Core/NS3 Protein Immunization with “N-Terminal Heat Shock gp96 Protein (rNT (gp96))” Induced Strong and Sustained Th1-Type Cytokines in Immunized Mice" Vaccines 9, no. 3: 215. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9030215