
Phenotypic and Genetic Evolutions of a Porcine Reproductive and Respiratory Syndrome Modified Live Vaccine after Limited Passages in Pigs

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
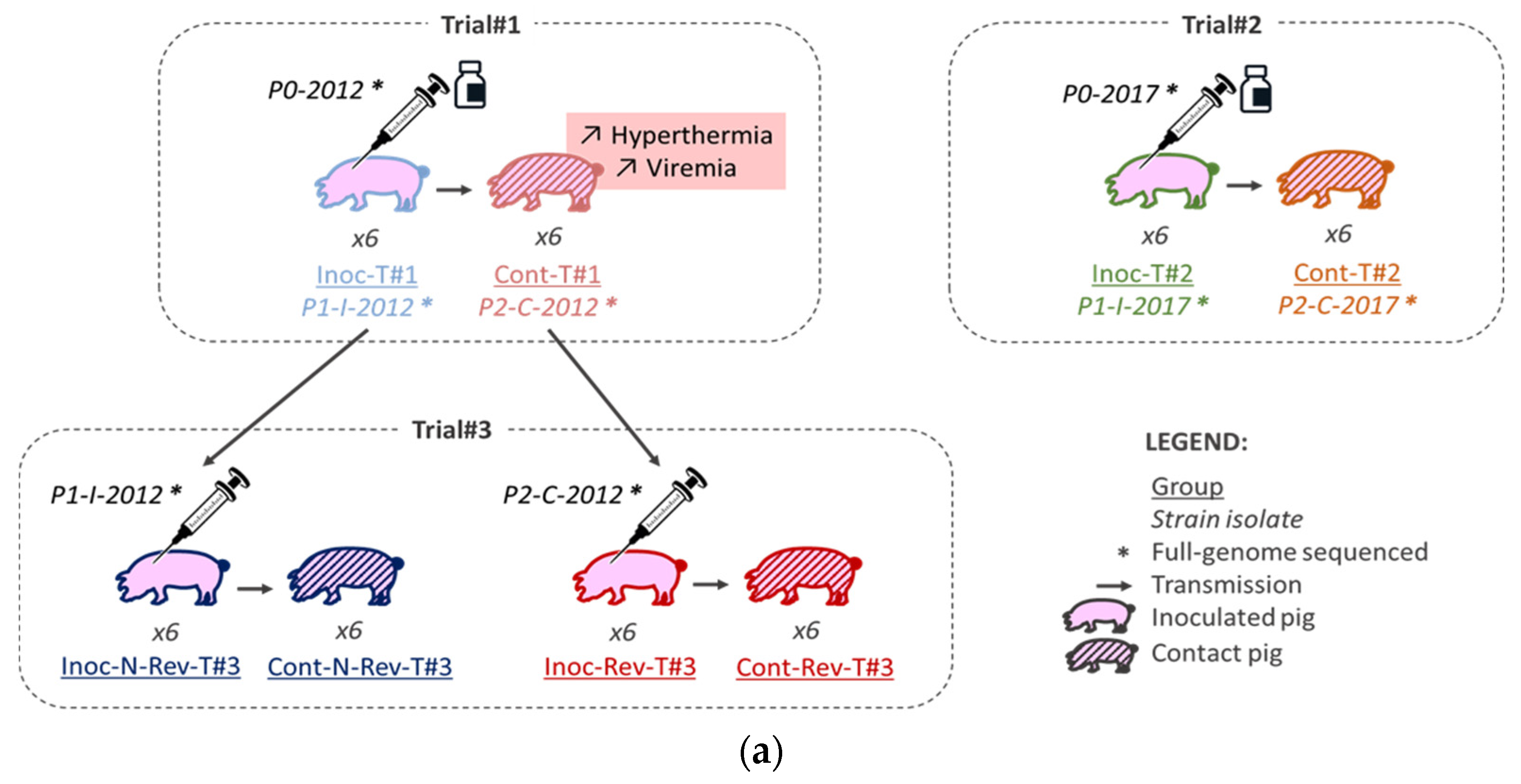
2.1. Animal Experiments
2.1.1. Viruses
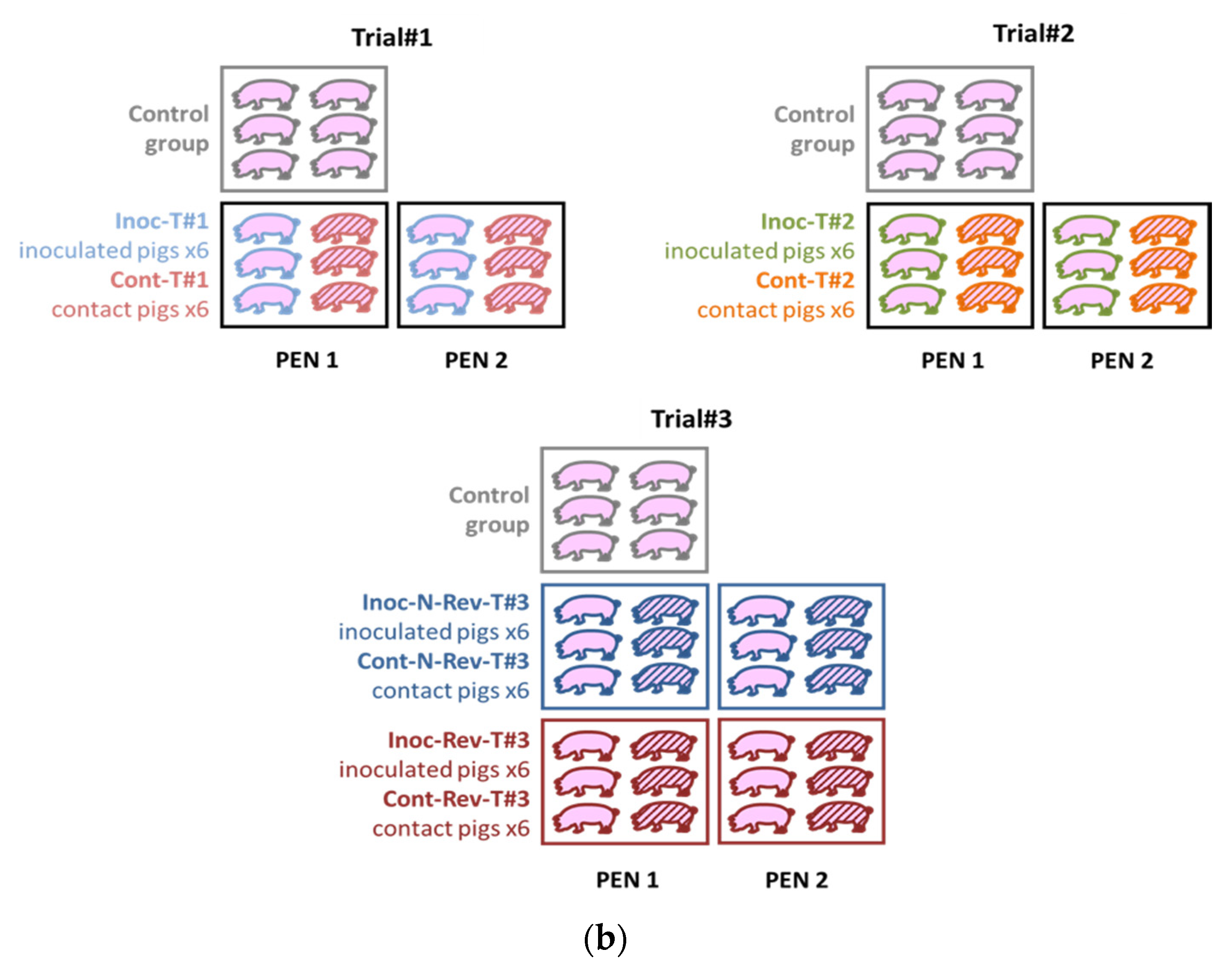
2.1.2. Animal Experiments
2.1.3. Ethical Statement
2.1.4. Quantification of PRRS Viral Genome Load
2.1.5. Statistical Analyses
2.1.6. Transmission Parameters
2.2. Genetic Characterization
2.2.1. Strains
2.2.2. Full-Genome Sequencing
2.2.3. Genomic Analysis
3. Results
3.1. Phenotypic Characterization
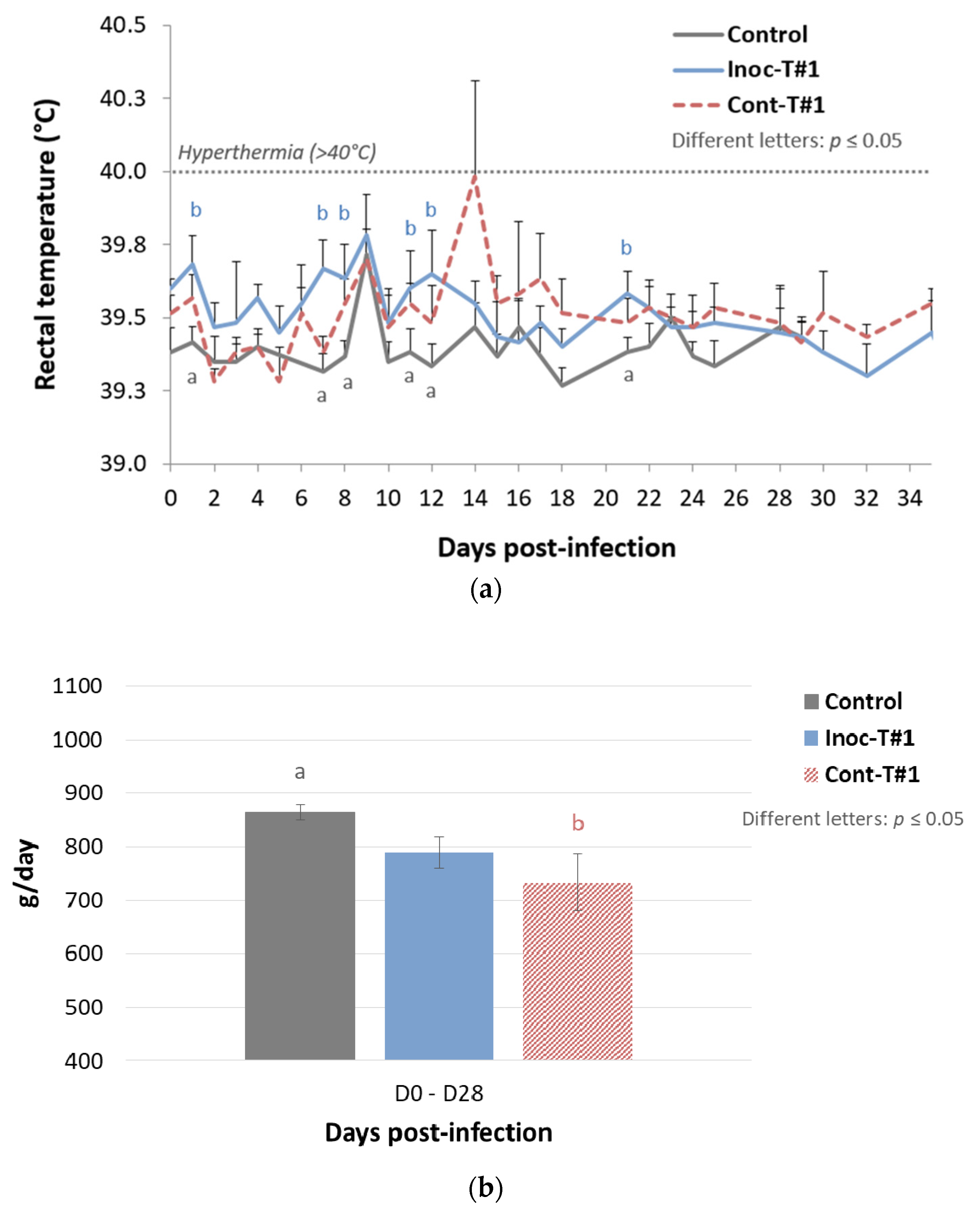
3.1.1. Trial#1
3.1.2. Trial#2
3.1.3. Trial#3
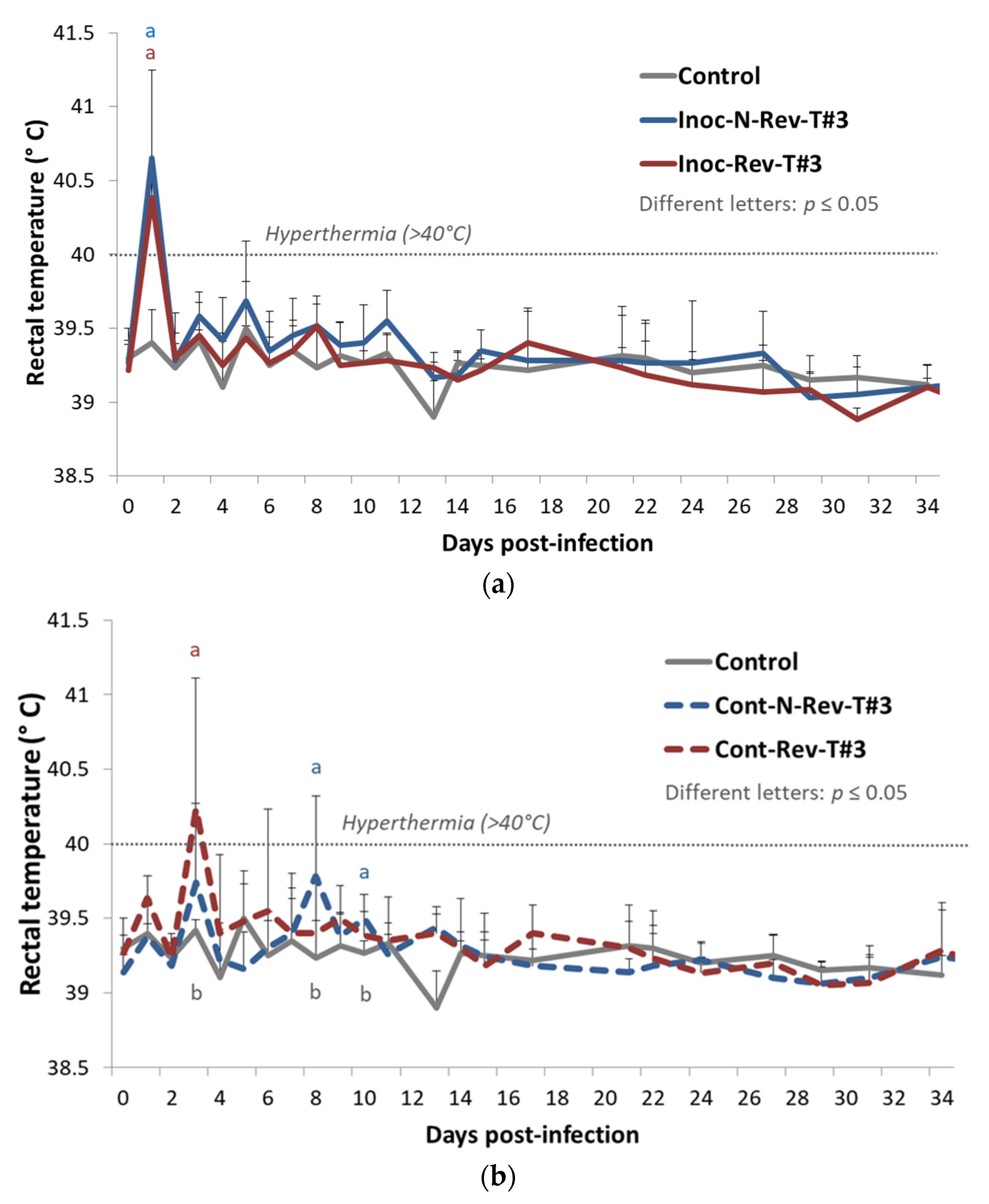
Rectal Temperatures
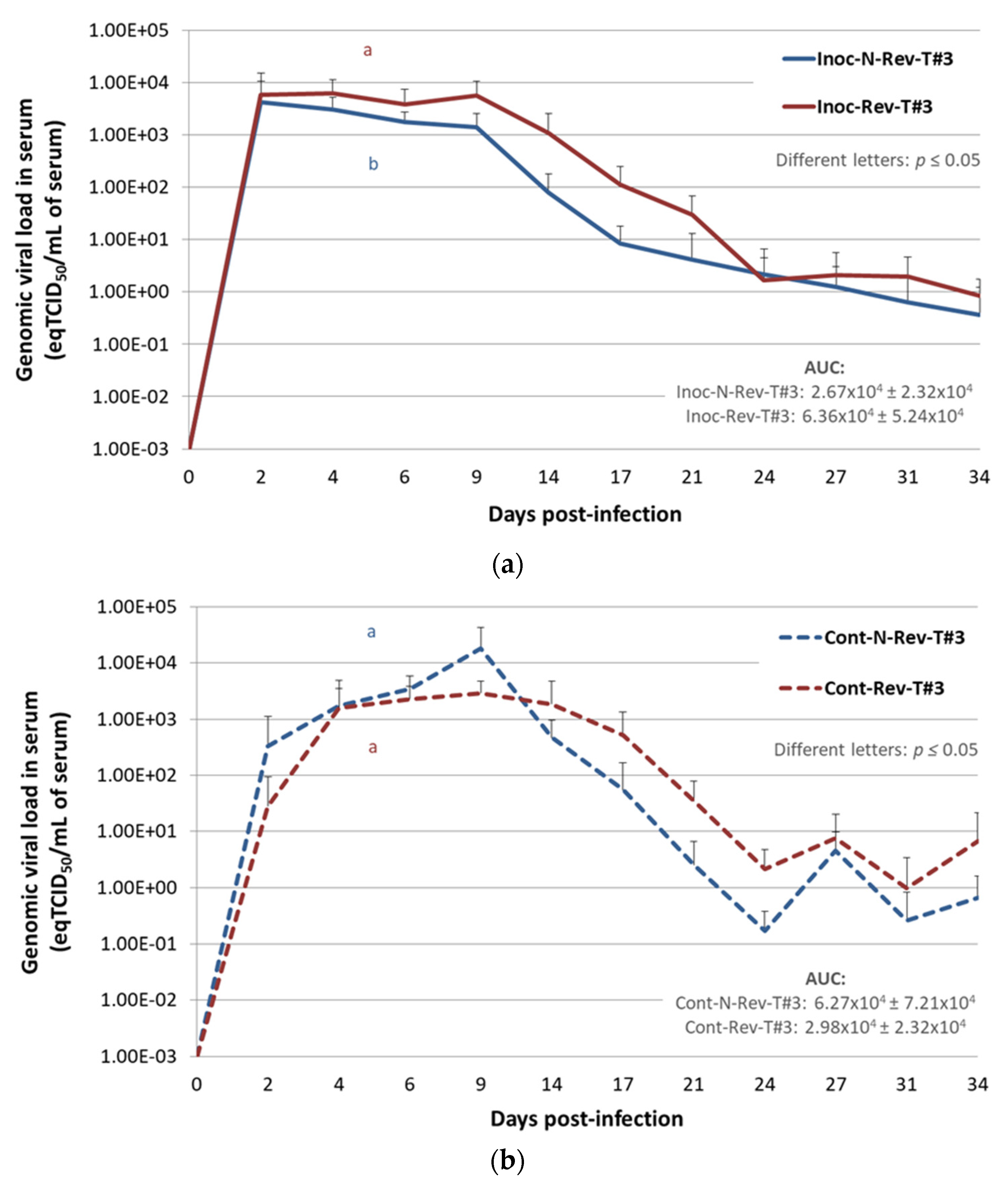
Viremia in Inoculated Pigs
Viral Nasal Excretion in Inoculated Pigs
Viremia in Contact Pigs
Transmission Parameters
3.2. Genomic Characterization
3.2.1. Coexistence of Different PRRSV Forms in Both DV Vaccine Batches
3.2.2. Selection of PRRSV Forms from Both DV Vaccine Batches Following Passages in Pigs
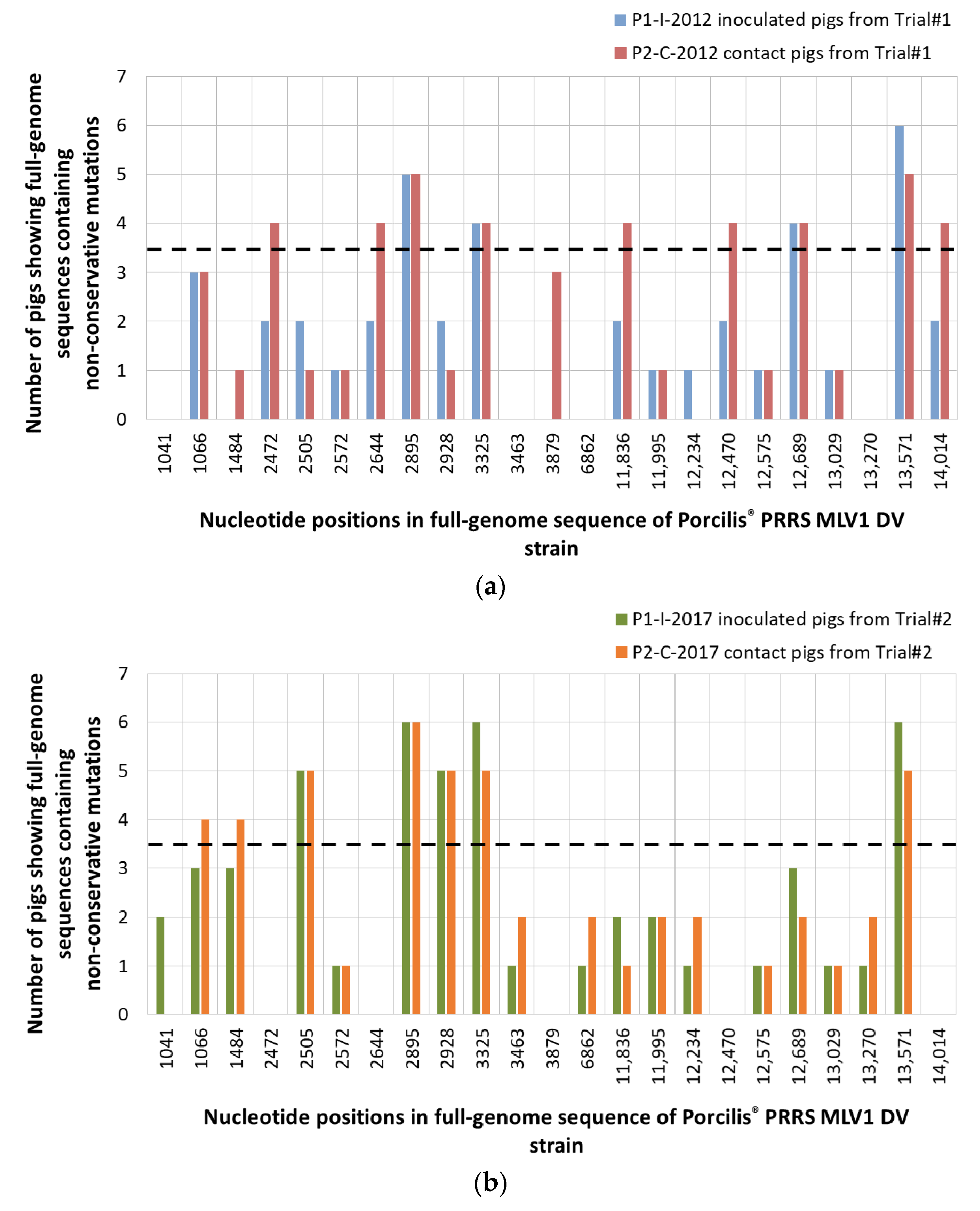
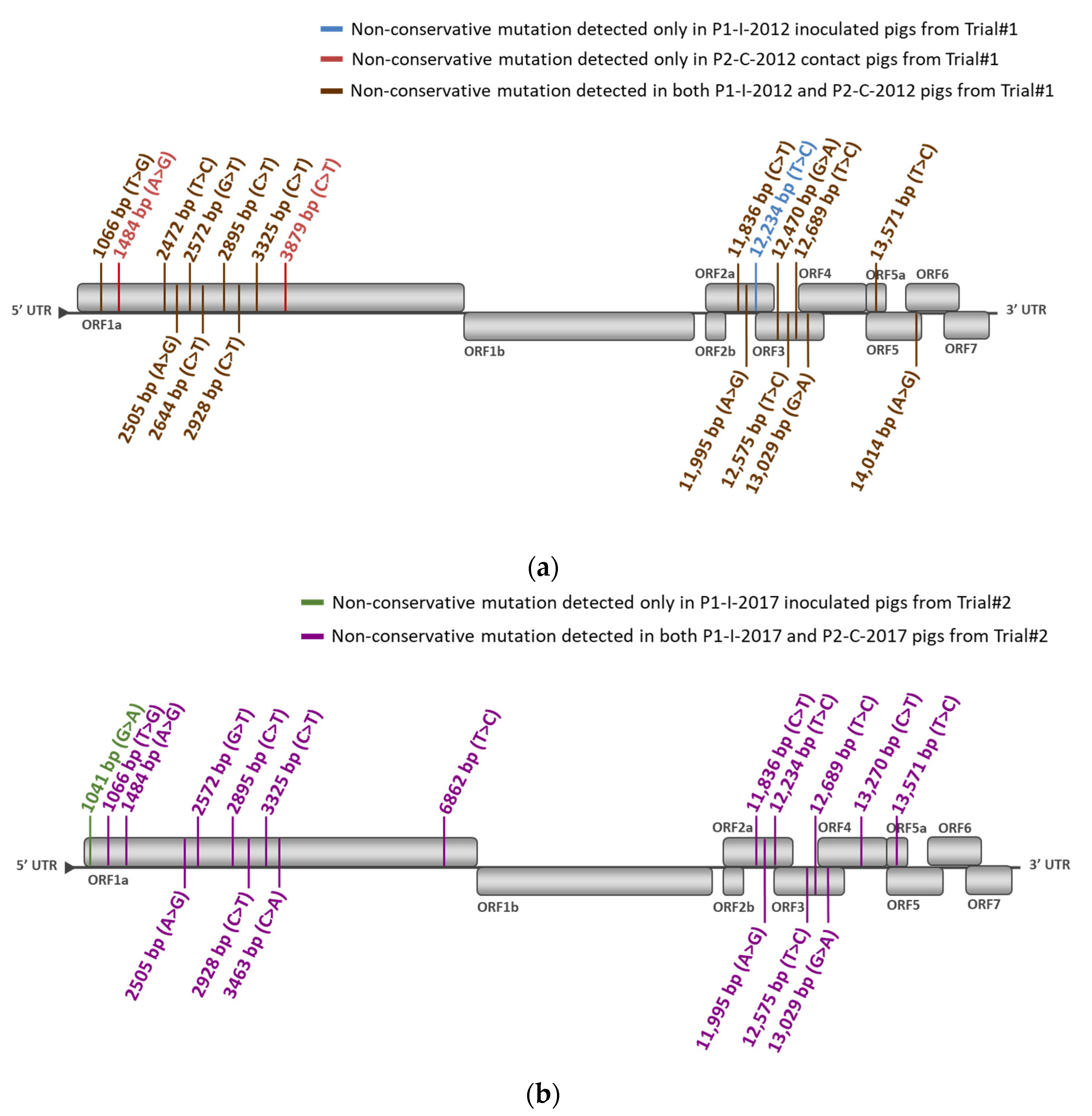
3.2.3. Identification of Non-Conservative Mutations during In Vivo Passages of DV Vaccine Strain
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holtkamp, D.J.; Kliebenstein, J.B.; Neumann, E.J.; Zimmerman, J.J.; Rotto, H.F.; Yoder, T.K.; Wang, C.; Yeske, P.E.; Mowrer, C.L.; Haley, C.A. Assessment of the economic impact of porcine reproductive and respiratory syndrome virus on United States pork producers. J. Swine Heal. Prod. 2013, 21, 72–84. [Google Scholar]
- Nathues, H.; Alarcon, P.; Rushton, J.; Jolie, R.; Fiebig, K.; Jimenez, M.; Geurts, V.; Nathues, C. Cost of porcine reproductive and respiratory syndrome virus at individual farm level—An economic disease model. Prev. Vet. Med. 2017, 142, 16–29. [Google Scholar] [CrossRef]
- Lunney, J.K.; Benfield, D.A.; Rowland, R.R. Porcine reproductive and respiratory syndrome virus: An update on an emerging and re-emerging viral disease of swine. Virus Res. 2010, 154, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Siddell, S.G.; Walker, P.J.; Lefkowitz, E.J.; Mushegian, A.R.; Adams, M.J.; Dutilh, B.E.; Gorbalenya, A.E.; Harrach, B.; Harrison, R.L.; Junglen, S.; et al. Additional changes to taxonomy ratified in a special vote by the International Committee on Taxonomy of Viruses (October 2018). Arch. Virol. 2019, 164, 943–946. [Google Scholar] [CrossRef] [Green Version]
- Balasuriya, U.B.; Carossino, M. Reproductive effects of arteriviruses: Equine arteritis virus and porcine reproductive and respiratory syndrome virus infections. Curr. Opin. Virol. 2017, 27, 57–70. [Google Scholar] [CrossRef]
- Kappes, M.A.; Faaberg, K.S. PRRSV structure, replication and recombination: Origin of phenotype and genotype diversity. Virology 2015, 479, 475–486. [Google Scholar] [CrossRef] [Green Version]
- Murtaugh, M.P.; Stadejek, T.; Abrahante, J.E.; Lam, T.T.; Leung, F.C. The ever-expanding diversity of porcine reproductive and respiratory syndrome virus. Virus Res. 2010, 154, 18–30. [Google Scholar] [CrossRef]
- OIE. Porcine Reproductive and Respiratory Syndrome. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; World Organisation for Animal Health (OIE): Paris, France, 2015. [Google Scholar]
- Nan, Y.; Wu, C.; Gu, G.; Sun, W.; Zhang, Y.J.; Zhou, E.M. Improved Vaccine against PRRSV: Current Progress and Future Perspective. Front. Microbiol. 2017, 8, 1635. [Google Scholar] [CrossRef]
- Charerntantanakul, W. Porcine reproductive and respiratory syndrome virus vaccines: Immunogenicity, efficacy and safety aspects. World J. Virol. 2012, 1, 23–30. [Google Scholar] [CrossRef]
- Botner, A.; Strandbygaard, B.; Sorensen, K.J.; Have, P.; Madsen, K.G.; Madsen, E.S.; Alexandersen, S. Appearance of acute PRRS-like symptoms in sow herds after vaccination with a modified live PRRS vaccine. Vet. Rec. 1997, 141, 497–499. [Google Scholar] [CrossRef]
- Nielsen, H.S.; Oleksiewicz, M.B.; Forsberg, R.; Stadejek, T.; Botner, A.; Storgaard, T. Reversion of a live porcine reproductive and respiratory syndrome virus vaccine investigated by parallel mutations. J. Gen. Virol. 2001, 82, 1263–1272. [Google Scholar] [CrossRef] [PubMed]
- Oh, W.T.; Nguyen, V.G.; Moon, H.J.; Lee, J.H.; Kim, H.K.; Park, S.J.; Chung, H.C.; Park, B.K. Investigation on Infectious Agents of Aborted Pig Fetuses and Its Correlation with PRRSV MLV Vaccine. J. Agric. Sci. Technol. 2017, 7, 282–287. [Google Scholar] [CrossRef] [Green Version]
- Grebennikova, T.V.; Clouser, D.F.; Vorwald, A.C.; Musienko, M.I.; Mengeling, W.L.; Lager, K.M.; Wesley, R.D.; Biketov, S.F.; Zaberezhny, A.D.; Aliper, T.I.; et al. Genomic characterization of virulent, attenuated, and revertant passages of a North American porcine reproductive and respiratory syndrome virus strain. Virology 2004, 321, 383–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, B.; Ansari, I.H.; Pattnaik, A.K.; Osorio, F.A. Identification of virulence determinants of porcine reproductive and respiratory syndrome virus through construction of chimeric clones. Virology 2008, 380, 371–378. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Chen, N.; Deng, X.; Cao, Z.; Han, W.; Hu, D.; Wu, J.; Zhang, S.; Wang, B.; Gu, X.; et al. Genomic sequencing reveals mutations potentially related to the overattenuation of a highly pathogenic porcine reproductive and respiratory syndrome virus. Clin. Vaccine Immunol. 2013, 20, 613–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ropp, S.L.; Wees, C.E.; Fang, Y.; Nelson, E.A.; Rossow, K.D.; Bien, M.; Arndt, B.; Preszler, S.; Steen, P.; Christopher-Hennings, J.; et al. Characterization of emerging European-like porcine reproductive and respiratory syndrome virus isolates in the United States. J. Virol. 2004, 78, 3684–3703. [Google Scholar] [CrossRef] [Green Version]
- Wensvoort, G.; Terpstra, C.; Pol, J.M.; ter Laak, E.A.; Bloemraad, M.; de Kluyver, E.P.; Kragten, C.; van Buiten, L.; den Besten, A.; Wagenaar, F.; et al. Mystery swine disease in The Netherlands: The isolation of Lelystad virus. Vet. Q. 1991, 13, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Charpin, C.; Mahe, S.; Keranflec’h, A.; Belloc, C.; Cariolet, R.; Le Potier, M.F.; Rose, N. Infectiousness of pigs infected by the Porcine Reproductive and Respiratory Syndrome virus (PRRSV) is time-dependent. Vet. Res. 2012, 43, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eclercy, J.; Renson, P.; Lebret, A.; Hirchaud, E.; Normand, V.; Andraud, M.; Paboeuf, F.; Blanchard, Y.; Rose, N.; Bourry, O. A Field Recombinant Strain Derived from Two Type 1 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV-1) Modified Live Vaccines Shows Increased Viremia and Transmission in SPF Pigs. Viruses 2019, 11, 296. [Google Scholar] [CrossRef] [Green Version]
- Rose, N.; Renson, P.; Andraud, M.; Paboeuf, F.; Le Potier, M.F.; Bourry, O. Porcine reproductive and respiratory syndrome virus (PRRSv) modified-live vaccine reduces virus transmission in experimental conditions. Vaccine 2015, 33, 2493–2499. [Google Scholar] [CrossRef] [PubMed]
- Chastagner, A.; Herve, S.; Bonin, E.; Queguiner, S.; Hirchaud, E.; Henritzi, D.; Beven, V.; Gorin, S.; Barbier, N.; Blanchard, Y.; et al. Spatiotemporal Distribution and Evolution of the A/H1N1 2009 Pandemic Influenza Virus in Pigs in France from 2009 to 2017: Identification of a Potential Swine-Specific Lineage. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Renson, P.; Fablet, C.; Andraud, M.; Normand, V.; Lebret, A.; Paboeuf, F.; Rose, N.; Bourry, O. Maternally-derived neutralizing antibodies reduce vaccine efficacy against porcine reproductive and respiratory syndrome virus infection. Vaccine 2019, 37, 4318–4324. [Google Scholar] [CrossRef]
- Khatun, A.; Shabir, N.; Seo, B.J.; Kim, B.S.; Yoon, K.J.; Kim, W.I. The Attenuation Phenotype of a Ribavirin-Resistant Porcine Reproductive and Respiratory Syndrome Virus Is Maintained during Sequential Passages in Pigs. J. Virol. 2016, 90, 4454–4468. [Google Scholar] [CrossRef] [Green Version]
- Eclercy, J.; Larcher, T.; Andraud, M.; Renson, P.; Bernard, C.; Bigault, L.; Ledevin, M.; Paboeuf, F.; Grasland, B.; Rose, N.; et al. PCV2 co-infection does not impact PRRSV MLV1 safety but enhances virulence of a PRRSV MLV1-like strain in infected SPF pigs. Vet. Microbiol. 2020, 244, 108656. [Google Scholar] [CrossRef] [PubMed]
- Kimman, T.G.; Cornelissen, L.A.; Moormann, R.J.; Rebel, J.M.; Stockhofe-Zurwieden, N. Challenges for porcine reproductive and respiratory syndrome virus (PRRSV) vaccinology. Vaccine 2009, 27, 3704–3718. [Google Scholar] [CrossRef]
- Martin-Valls, G.E.; Kvisgaard, L.K.; Tello, M.; Darwich, L.; Cortey, M.; Burgara-Estrella, A.J.; Hernandez, J.; Larsen, L.E.; Mateu, E. Analysis of ORF5 and full-length genome sequences of porcine reproductive and respiratory syndrome virus isolates of genotypes 1 and 2 retrieved worldwide provides evidence that recombination is a common phenomenon and may produce mosaic isolates. J. Virol. 2014, 88, 3170–3181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kvisgaard, L.K.; Kristensen, C.S.; Hjulsager, C.K.; Larsen, L.E. Characterization of the prrsv strain circulating in a prrsv type 1 positive herd before, during, and after vaccination with a prrsv type 1 vaccine. In Proceedings of the 7th International Symposium on Emerging and Re-emerging Pig Diseases, Kyoto, Japan, 21–24 June 2015. [Google Scholar]
- Cortey, M.; Arocena, G.; Pileri, E.; Martin-Valls, G.; Mateu, E. Bottlenecks in the transmission of porcine reproductive and respiratory syndrome virus (PRRSV1) to naive pigs and the quasi-species variation of the virus during infection in vaccinated pigs. Vet. Res. 2018, 49, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Allende, R.; Kutish, G.F.; Laegreid, W.; Lu, Z.; Lewis, T.L.; Rock, D.L.; Friesen, J.; Galeota, J.A.; Doster, A.R.; Osorio, F.A. Mutations in the genome of porcine reproductive and respiratory syndrome virus responsible for the attenuation phenotype. Arch. Virol. 2000, 145, 1149–1161. [Google Scholar] [CrossRef] [PubMed]
- Spear, A.; Wang, F.X.; Kappes, M.A.; Das, P.B.; Faaberg, K.S. Progress toward an enhanced vaccine: Eight marked attenuated viruses to porcine reproductive and respiratory disease virus. Virology 2018, 516, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Mickelson, D.; Murtaugh, M.P.; Faaberg, K.S. Complete genome comparison of porcine reproductive and respiratory syndrome virus parental and attenuated strains. Virus Res. 2001, 74, 99–110. [Google Scholar] [CrossRef]
- Song, J.; Gao, P.; Kong, C.; Zhou, L.; Ge, X.; Guo, X.; Han, J.; Yang, H. The nsp2 Hypervariable Region of Porcine Reproductive and Respiratory Syndrome Virus Strain JXwn06 Is Associated with Viral Cellular Tropism to Primary Porcine Alveolar Macrophages. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Hodgins, D.; Calvert, J.G.; Welch, S.K.; Jolie, R.; Yoo, D. Mutations within the nuclear localization signal of the porcine reproductive and respiratory syndrome virus nucleocapsid protein attenuate virus replication. Virology 2006, 346, 238–250. [Google Scholar] [CrossRef] [Green Version]
- Ni, Y.Y.; Opriessnig, T.; Zhou, L.; Cao, D.; Huang, Y.W.; Halbur, P.G.; Meng, X.J. Attenuation of porcine reproductive and respiratory syndrome virus by molecular breeding of virus envelope genes from genetically divergent strains. J. Virol. 2013, 87, 304–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pei, Y.; Hodgins, D.C.; Lee, C.; Calvert, J.G.; Welch, S.K.; Jolie, R.; Keith, M.; Yoo, D. Functional mapping of the porcine reproductive and respiratory syndrome virus capsid protein nuclear localization signal and its pathogenic association. Virus Res. 2008, 135, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Trus, I.; Oh, D.; Kvisgaard, L.K.; Rappe, J.C.F.; Ruggli, N.; Vanderheijden, N.; Larsen, L.E.; Lefevre, F.; Nauwynck, H.J. A Triple Amino Acid Substitution at Position 88/94/95 in Glycoprotein GP2a of Type 1 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV1) Is Responsible for Adaptation to MARC-145 Cells. Viruses 2019, 11, 36. [Google Scholar] [CrossRef] [Green Version]
- Botner, A.; Nielsen, J.; Oleksiewicz, M.B.; Storgaard, T. Heterologous challenge with porcine reproductive and respiratory syndrome (PRRS) vaccine virus: No evidence of reactivation of previous European-type PRRS virus infection. Vet. Microbiol. 1999, 68, 187–195. [Google Scholar] [CrossRef]
- Madsen, K.G.; Hansen, C.M.; Madsen, E.S.; Strandbygaard, B.; Botner, A.; Sorensen, K.J. Sequence analysis of porcine reproductive and respiratory syndrome virus of the American type collected from Danish swine herds. Arch. Virol. 1998, 143, 1683–1700. [Google Scholar] [CrossRef]
- Storgaard, T.; Oleksiewicz, M.; Botner, A. Examination of the selective pressures on a live PRRS vaccine virus. Arch. Virol. 1999, 144, 2389–2401. [Google Scholar] [CrossRef]
- Wang, F.X.; Song, N.; Chen, L.Z.; Cheng, S.P.; Wu, H.; Wen, Y.J. Non-structural protein 2 of the porcine reproductive and respiratory syndrome (PRRS) virus: A crucial protein in viral pathogenesis, immunity and diagnosis. Res. Vet. Sci. 2013, 95, 1–7. [Google Scholar] [CrossRef] [PubMed]
- An, T.Q.; Tian, Z.J.; Zhou, Y.J.; Xiao, Y.; Peng, J.M.; Chen, J.; Jiang, Y.F.; Hao, X.F.; Tong, G.Z. Comparative genomic analysis of five pairs of virulent parental/attenuated vaccine strains of PRRSV. Vet. Microbiol. 2011, 149, 104–112. [Google Scholar] [CrossRef]
- Kvisgaard, L.K.; Kristensen, C.S.; Ryt-Hansen, P.; Pedersen, K.; Stadejek, T.; Trebbien, R.; Andresen, L.O.; Larsen, L.E. A recombination between two Type 1 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV-1) vaccine strains has caused severe outbreaks in Danish pigs. Transbound. Emerg. Dis. 2020, 67, 1786–1796. [Google Scholar] [CrossRef] [PubMed] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trial | Trial#1 | Trial#2 | Trial#3 | |
|---|---|---|---|---|
| Number of pigs | 6 inoculated 6 contact | 6 inoculated 6 contact | 6 inoculated 6 contact | 6 inoculated 6 contact |
| Age of pigs (weeks) | 6 | 7 | 7 | 7 |
| Inoculated strain | Porcilis® PRRS (batch No. A142DB01) P0-2012 | Porcilis® PRRS (batch No. A207CB01) P0-2017 | DV isolate P1-I-2012 | DV isolate P2-C-2012 |
| Inoculum titer in MARC-145 cells (TCID50/mL) | 104.2 | 104.8 | 104.5 | 104.5 |
| Inoculation route | IM | IM | IM | IM |
| Transmission Parameters | N-Rev Strain | Rev Strain |
|---|---|---|
| Daily transmission rate (number of infected pigs per one infectious pig per day) | 0.58 a [0.23; 1.69] * | 0.75 b [0.29; 1.90] |
| Duration of latency (days) | 0.7 a [0.4; 1.7] | 0.7 a [0.4; 1.5] |
| Variant Name | Deletion Position (Nucleotide Base Pair) | Deletion Size (Base Pair) | Identity Rate with KF991509.2 (%) |
|---|---|---|---|
| P0-LONG-DEL | 2216–2437 | 222 | 99.95 |
| P0-SHORT-DEL | 2231–2365 | 135 | 99.93 |
| P0-SHIFT-DEL | 2344–2435 2446–2506 | 92 61 | 99.94 |
| Trial | Inoculated or Contact Pig | Pen | Pig No. | Full-Length Sequence | Variants | % Identity with Variants of Reference (P0) | ||
|---|---|---|---|---|---|---|---|---|
| Long(L) | Short(S) | Shift(F) | ||||||
| #1 | Inoc | 1 | 2012_1 | ND | <1% | >99% | ND | 99.79% |
| Inoc | 1 | 2012_2 | ND | >99% | ND | ND | 99.87% | |
| Inoc | 1 | 2012_3 | ND | ND | 32% | 67% | 99.86% | |
| Cont | 1 | 2012_4 | <1% | >99% | ND | ND | 99.85% | |
| Cont | 1 | 2012_5 | ND | >99% | ND | ND | 99.85% | |
| Cont | 1 | 2012_6 ** | ND | >99% | ND | ND | 99.84% | |
| Inoc | 2 | 2012_7 * | ND | ND | ND | >99% | 99.87% | |
| Inoc | 2 | 2012_8 | ND | >99% | ND | ND | 99.85% | |
| Inoc | 2 | 2012_9 | ND | ND | 50% | 50% | 99.90% | |
| Cont | 2 | 2012_10 | ND | >99% | ND | ND | 99.86% | |
| Cont | 2 | 2012_11 | ND | ND | >99% | ND | 99.85% | |
| Viral form detection frequency in Inoc pigs (≥1%) | 0/6 | 2/6 | 3/6 | 3/6 | ||||
| Viral form detection frequency in Cont pigs (≥1%) | 0/5 | 4/5 | 1/5 | 0/5 | ||||
| Viral form detection frequency in Trial#1 (≥1%) | 0/11 | 6/11 | 4/11 | 3/11 | ||||
| #2 | Inoc | 1 | 2017_12 | ND | >99% | ND | ND | 99.83% |
| Inoc | 1 | 2017_13 | ND | ND | >99% | ND | 99.83% | |
| Inoc | 1 | 2017_14 | ND | <1% | >99% | ND | 99.85% | |
| Cont | 1 | 2017_15 | ND | <1% | >99% | ND | 99.79% | |
| Cont | 1 | 2017_16 | ND | >99% | <1% | ND | 99.82% | |
| Cont | 1 | 2017_17 | ND | <1% | >99% | ND | 99.83% | |
| Inoc | 2 | 2017_18 | ND | ND | >99% | ND | 99.78% | |
| Inoc | 2 | 2017_19 | ND | >99% | ND | ND | 99.85% | |
| Inoc | 2 | 2017_20 | 5% | 44% | 51% | ND | 99.87% | |
| Cont | 2 | 2017_21 | ND | >99% | ND | ND | 99.83% | |
| Cont | 2 | 2017_22 | ND | >99% | ND | ND | 99.83% | |
| Cont | 2 | 2017_23 | ND | >99% | ND | ND | 99.87% | |
| Viral form detection frequency in Inoc pigs (≥1%) | 1/6 | 3/6 | 4/6 | 0/6 | ||||
| Viral form detection frequency in Cont pigs (≥1%) | 0/6 | 4/6 | 2/6 | 0/6 | ||||
| Viral form detection frequency in Trial#2 (≥1%) | 1/12 | 7/12 | 6/12 | 0/12 | ||||
| TOTAL VIRAL FORM DETECTION FREQUENCY (≥1%) | 1/23 | 13/23 | 10/23 | 3/23 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eclercy, J.; Renson, P.; Hirchaud, E.; Andraud, M.; Beven, V.; Paboeuf, F.; Rose, N.; Blanchard, Y.; Bourry, O. Phenotypic and Genetic Evolutions of a Porcine Reproductive and Respiratory Syndrome Modified Live Vaccine after Limited Passages in Pigs. Vaccines 2021, 9, 392. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9040392
Eclercy J, Renson P, Hirchaud E, Andraud M, Beven V, Paboeuf F, Rose N, Blanchard Y, Bourry O. Phenotypic and Genetic Evolutions of a Porcine Reproductive and Respiratory Syndrome Modified Live Vaccine after Limited Passages in Pigs. Vaccines. 2021; 9(4):392. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9040392
Chicago/Turabian StyleEclercy, Julie, Patricia Renson, Edouard Hirchaud, Mathieu Andraud, Véronique Beven, Frédéric Paboeuf, Nicolas Rose, Yannick Blanchard, and Olivier Bourry. 2021. "Phenotypic and Genetic Evolutions of a Porcine Reproductive and Respiratory Syndrome Modified Live Vaccine after Limited Passages in Pigs" Vaccines 9, no. 4: 392. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9040392