
Use of a Novel Peptide Welding Technology Platform for the Development of B- and T-Cell Epitope-Based Vaccines

 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
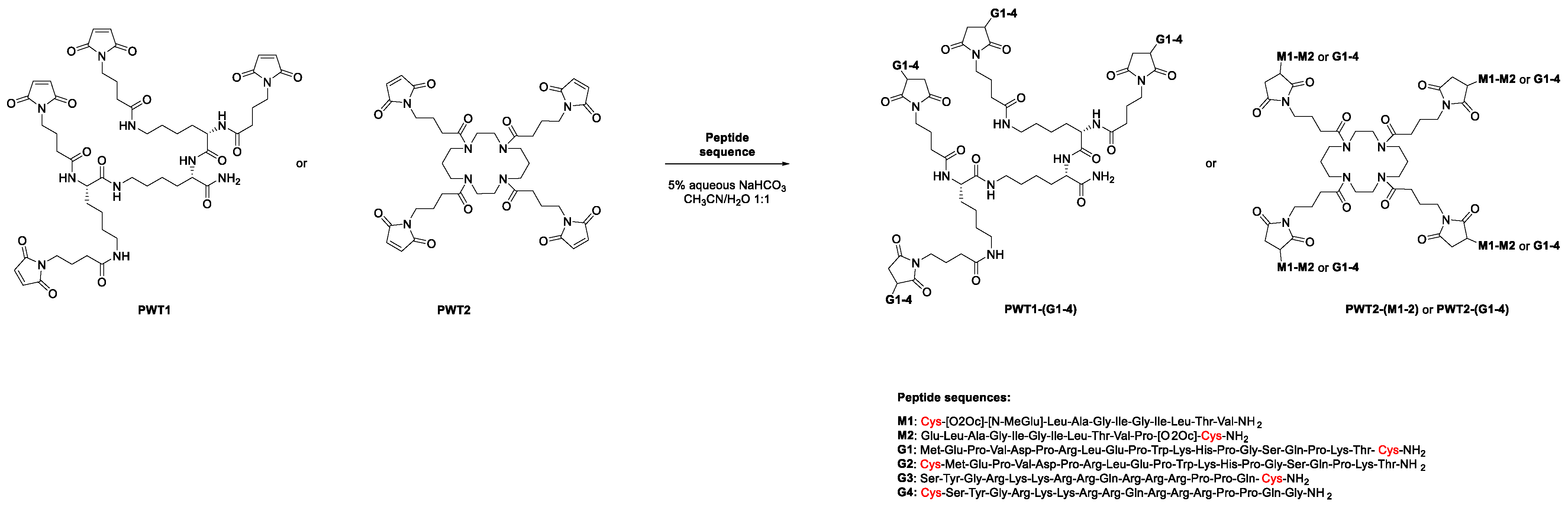
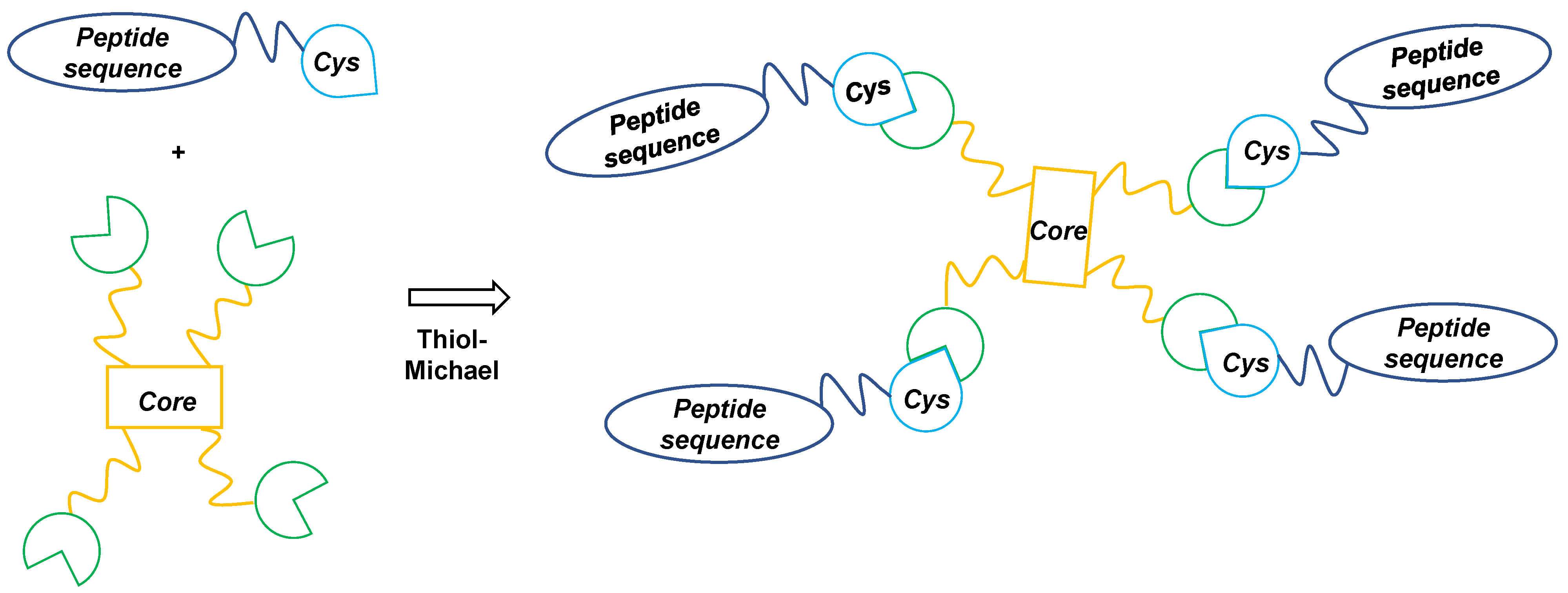
2.1. Peptides and PWT-Derivatives
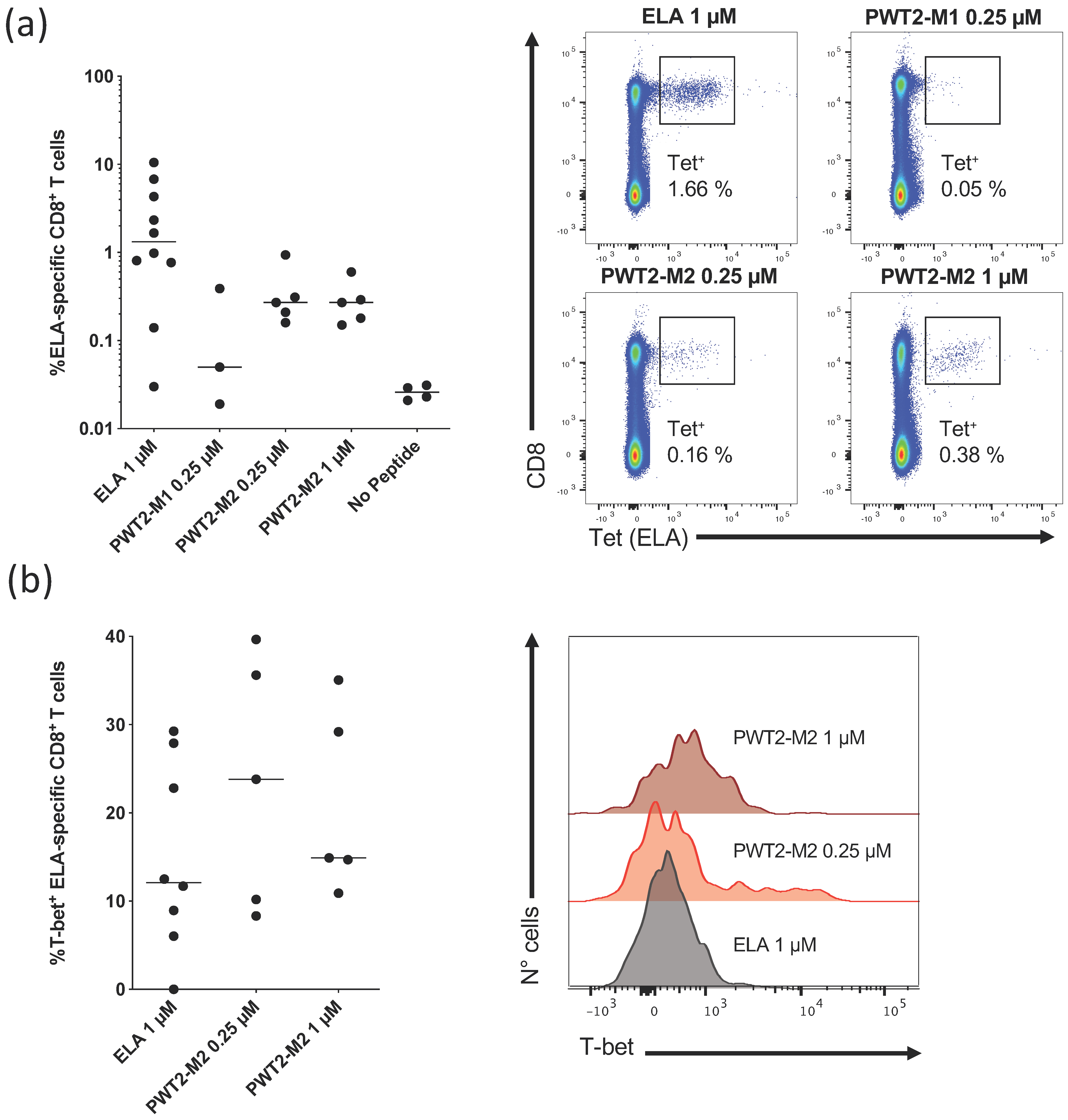
2.2. In Vitro Priming of Human Antigen-Specific CD8+ T Cells
2.3. Mouse Immunization
2.4. Serology
2.5. Statistical Analysis
3. Results
3.1. Chemistry
3.2. PWT Scaffolds Can Present T-Cell Epitopes
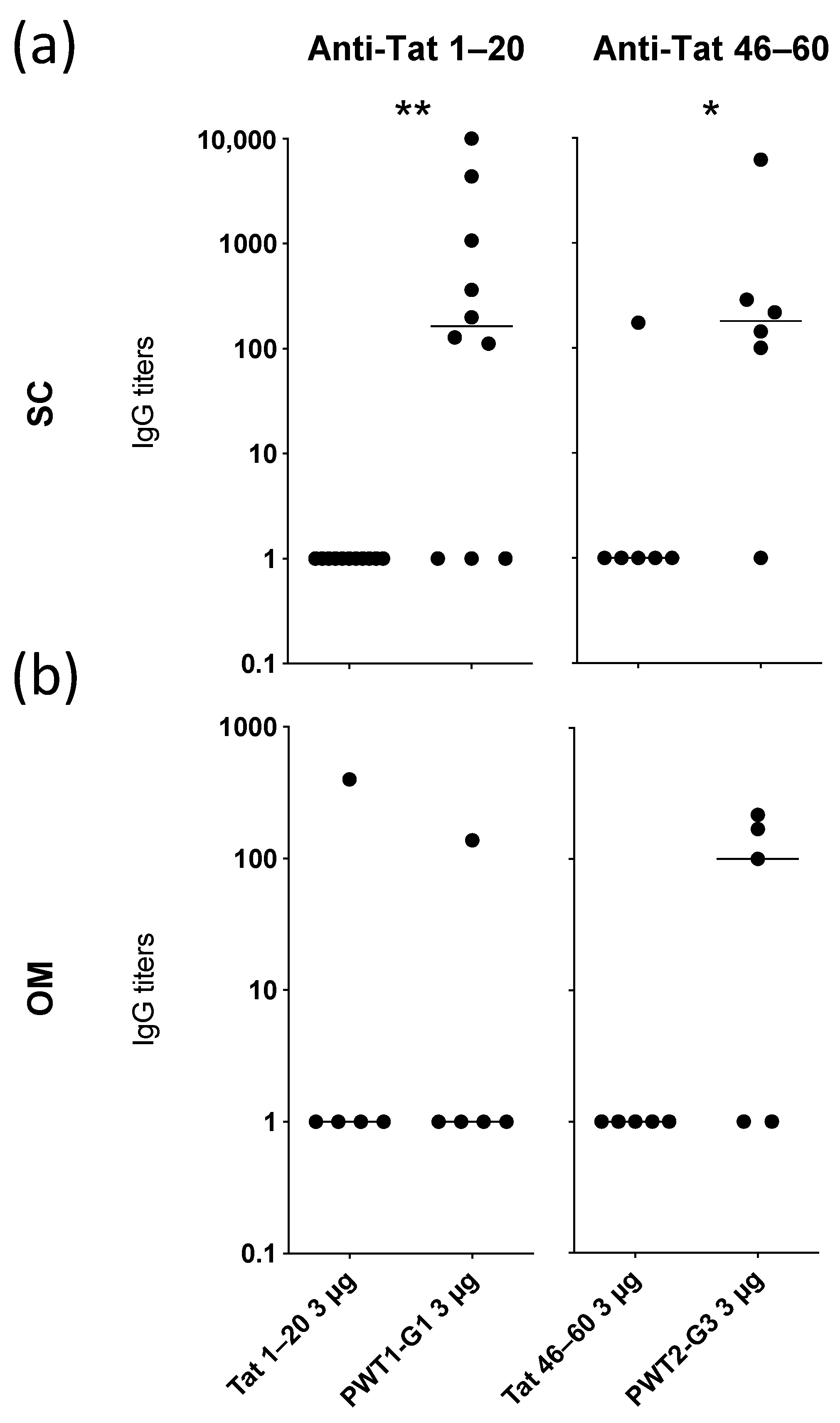
3.3. PWT Scaffolds Can Present B-Cell Epitopes
3.4. Conjugation of Peptide Epitopes to PWT Scaffolds Enhances Their Immunogenicity
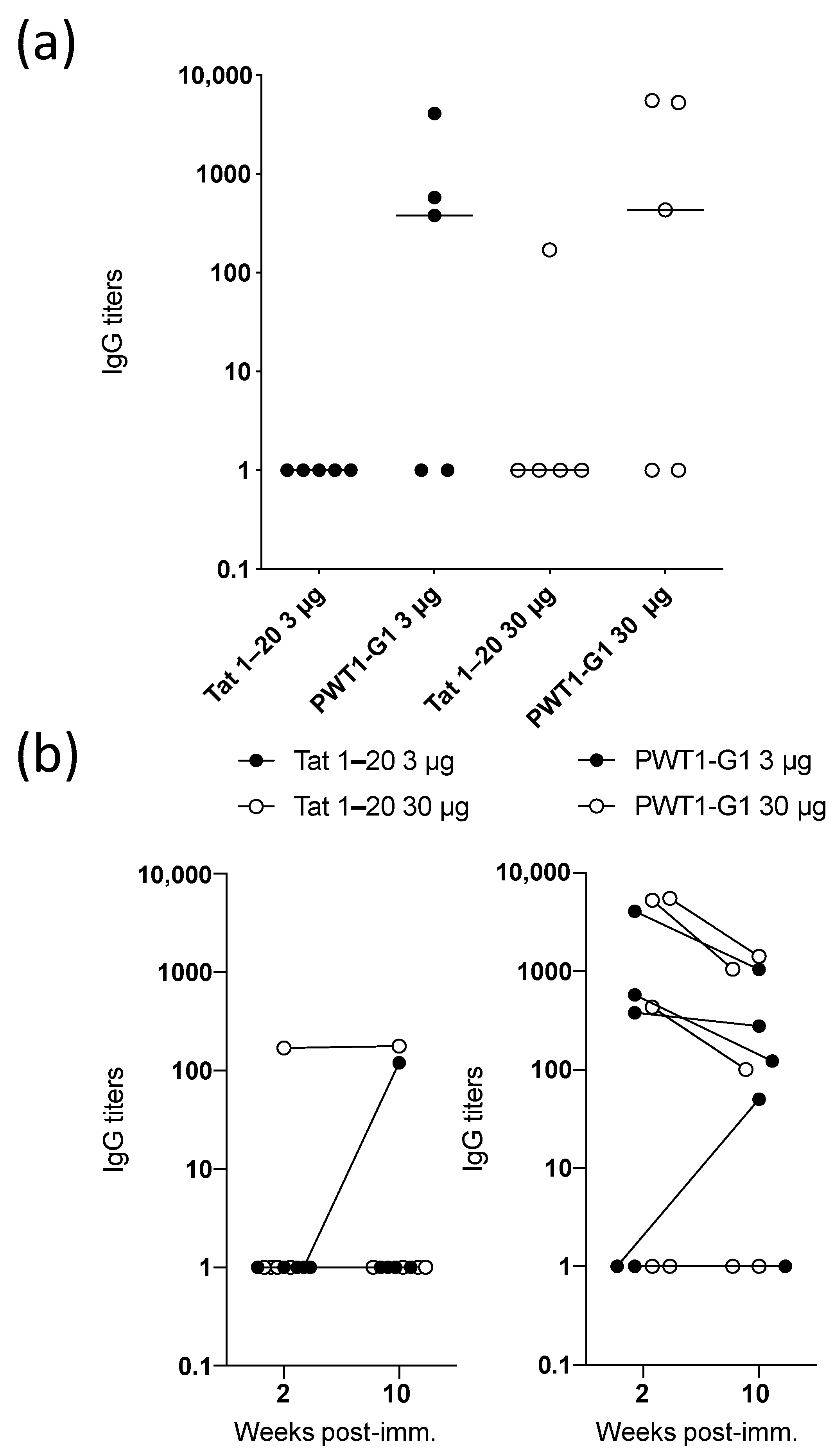
3.5. IgG Titers Induced by Peptides Conjugated to PWT Scaffolds Are Maintained Over Time
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Appendix A



















References
- Nicoli, F.; Appay, V. Immunological considerations regarding parental concerns on pediatric immunizations. Vaccine 2017, 35, 3012–3019. [Google Scholar] [CrossRef] [Green Version]
- Sallam, M. COVID-19 Vaccine Hesitancy Worldwide: A Concise Systematic Review of Vaccine Acceptance Rates. Vaccines 2021, 9, 160. [Google Scholar] [CrossRef] [PubMed]
- Sabatino, D. Medicinal Chemistry and Methodological Advances in the Development of Peptide-Based Vaccines. J. Med. Chem. 2020, 63, 14184–14196. [Google Scholar] [CrossRef]
- Yoshida, K.; Noguchi, M.; Mine, T.; Komatsu, N.; Yutani, S.; Ueno, T.; Yanagimoto, H.; Kawano, K.; Itoh, K.; Yamada, A. Characteristics of severe adverse events after peptide vaccination for advanced cancer patients: Analysis of 500 cases. Oncol. Rep. 2011, 25, 57–62. [Google Scholar]
- Nelde, A.; Rammensee, H.G.; Walz, J.S. The Peptide Vaccine of the Future. Mol. Cell. Proteom. 2021, 20, 100022. [Google Scholar] [CrossRef] [PubMed]
- Hilf, N.; Kuttruff-Coqui, S.; Frenzel, K.; Bukur, V.; Stevanovic, S.; Gouttefangeas, C.; Platten, M.; Tabatabai, G.; Dutoit, V.; van der Burg, S.H.; et al. Actively personalized vaccination trial for newly diagnosed glioblastoma. Nature 2019, 565, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Finessi, V.; Nicoli, F.; Gallerani, E.; Sforza, F.; Sicurella, M.; Cafaro, A.; Caputo, A.; Ensoli, B.; Gavioli, R. Effects of different routes of administration on the immunogenicity of the Tat protein and a Tat-derived peptide. Hum. Vaccin. Immunother. 2015, 11, 1489–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, J.P. Synthetic peptide vaccine design: Synthesis and properties of a high-density multiple antigenic peptide system. Proc. Natl. Acad. Sci. USA 1988, 85, 5409–5413. [Google Scholar] [CrossRef] [Green Version]
- Calo, G.; Rizzi, A.; Ruzza, C.; Ferrari, F.; Pacifico, S.; Gavioli, E.C.; Salvadori, S.; Guerrini, R. Peptide welding technology—A simple strategy for generating innovative ligands for G protein coupled receptors. Peptides 2018, 99, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Cerlesi, M.C.; Ding, H.; Bird, M.F.; Kiguchi, N.; Ferrari, F.; Malfacini, D.; Rizzi, A.; Ruzza, C.; Lambert, D.G.; Ko, M.C.; et al. Pharmacological studies on the NOP and opioid receptor agonist PWT2-[Dmt(1)]N/OFQ(1-13). Eur. J. Pharmacol. 2017, 794, 115–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bracci, L.; Falciani, C.; Lelli, B.; Lozzi, L.; Runci, Y.; Pini, A.; De Montis, M.G.; Tagliamonte, A.; Neri, P. Synthetic peptides in the form of dendrimers become resistant to protease activity. J. Biol. Chem. 2003, 278, 46590–46595. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.J.; Pugh, W.; Blanchard, S.G.; McDermed, J.; Tam, J.P. Antibody specific to the alpha subunit of the guanine nucleotide-binding regulatory protein Go: Developmental appearance and immunocytochemical localization in brain. Proc. Natl. Acad. Sci. USA 1988, 85, 4929–4933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behrendt, R.; White, P.; Offer, J. Advances in Fmoc solid-phase peptide synthesis. J. Pept. Sci. 2016, 22, 4–27. [Google Scholar] [CrossRef] [Green Version]
- Telatin, V.; Nicoli, F.; Frasson, C.; Menegotto, N.; Barbaro, F.; Castelli, E.; Erne, E.; Palu, G.; Caputo, A. In Chronic Hepatitis C Infection, Myeloid-Derived Suppressor Cell Accumulation and T Cell Dysfunctions Revert Partially and Late After Successful Direct-Acting Antiviral Treatment. Front. Cell. Infect. Microbiol. 2019, 9, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicoli, F.; Gallerani, E.; Sforza, F.; Finessi, V.; Chachage, M.; Geldmacher, C.; Cafaro, A.; Ensoli, B.; Caputo, A.; Gavioli, R. The HIV-1 Tat protein affects human CD4+ T-cell programing and activation, and favors the differentiation of naive CD4+ T cells. AIDS 2018, 32, 575–581. [Google Scholar] [CrossRef]
- Papagno, L.; Kuse, N.; Lissina, A.; Gostick, E.; Price, D.A.; Appay, V.; Nicoli, F. The TLR9 ligand CpG ODN 2006 is a poor adjuvant for the induction of de novo CD8+ T-cell responses in vitro. Sci. Rep. 2020, 10, 11620. [Google Scholar] [CrossRef] [PubMed]
- Nicoli, F.; Papagno, L.; Frere, J.J.; Cabral-Piccin, M.P.; Clave, E.; Gostick, E.; Toubert, A.; Price, D.A.; Caputo, A.; Appay, V. Naive CD8+ T-Cells Engage a Versatile Metabolic Program Upon Activation in Humans and Differ Energetically From Memory CD8+ T-Cells. Front. Immunol. 2018, 9, 2736. [Google Scholar] [CrossRef] [Green Version]
- Caputo, A.; Brocca-Cofano, E.; Castaldello, A.; Voltan, R.; Gavioli, R.; Srivastava, I.K.; Barnett, S.W.; Cafaro, A.; Ensoli, B. Characterization of immune responses elicited in mice by intranasal co-immunization with HIV-1 Tat, gp140 DeltaV2Env and/or SIV Gag proteins and the nontoxicogenic heat-labile Escherichia coli enterotoxin. Vaccine 2008, 26, 1214–1227. [Google Scholar] [CrossRef] [PubMed]
- Nicoli, F.; Finessi, V.; Sicurella, M.; Rizzotto, L.; Gallerani, E.; Destro, F.; Cafaro, A.; Marconi, P.; Caputo, A.; Ensoli, B.; et al. The HIV-1 Tat protein induces the activation of CD8+ T cells and affects in vivo the magnitude and kinetics of antiviral responses. PLoS ONE 2013, 8, e77746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicoli, F.; Chachage, M.; Clowes, P.; Bauer, A.; Kowour, D.; Ensoli, B.; Cafaro, A.; Maboko, L.; Hoelscher, M.; Gavioli, R.; et al. Association between different anti-Tat antibody isotypes and HIV disease progression: Data from an African cohort. BMC Infect. Dis. 2016, 16, 344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicoli, F.; Gallerani, E.; Sicurella, M.; Pacifico, S.; Cafaro, A.; Ensoli, B.; Marconi, P.; Caputo, A.; Gavioli, R. The Tat Protein of HIV-1 Prevents the Loss of HSV-Specific Memory Adaptive Responses and Favors the Control of Viral Reactivation. Vaccines 2020, 8, 274. [Google Scholar] [CrossRef]
- Sicurella, M.; Nicoli, F.; Gallerani, E.; Volpi, I.; Berto, E.; Finessi, V.; Destro, F.; Manservigi, R.; Cafaro, A.; Ensoli, B.; et al. An attenuated herpes simplex virus type 1 (HSV1) encoding the HIV-1 Tat protein protects mice from a deadly mucosal HSV1 challenge. PLoS ONE 2014, 9, e100844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicoli, F.; Gallerani, E.; Skarlis, C.; Sicurella, M.; Cafaro, A.; Ensoli, B.; Caputo, A.; Marconi, P.C.; Gavioli, R. Systemic immunodominant CD8 responses with an effector-like phenotype are induced by intravaginal immunization with attenuated HSV vectors expressing HIV Tat and mediate protection against HSV infection. Vaccine 2016, 34, 2216–2224. [Google Scholar] [CrossRef] [PubMed]
- Guerrini, R.; Marzola, E.; Trapella, C.; Pela, M.; Molinari, S.; Cerlesi, M.C.; Malfacini, D.; Rizzi, A.; Salvadori, S.; Calo, G. A novel and facile synthesis of tetra branched derivatives of nociceptin/orphanin FQ. Bioorg. Med. Chem. 2014, 22, 3703–3712. [Google Scholar] [CrossRef] [PubMed]
- Lambert, T.H.; Danishefsky, S.J. Total synthesis of UCS1025A. J. Am. Chem. Soc. 2006, 128, 426–427. [Google Scholar] [CrossRef] [PubMed]
- Song, H.Y.; Ngai, M.H.; Song, Z.Y.; MacAry, P.A.; Hobley, J.; Lear, M.J. Practical synthesis of maleimides and coumarin-linked probes for protein and antibody labelling via reduction of native disulfides. Org. Biomol. Chem. 2009, 7, 3400–3406. [Google Scholar] [CrossRef]
- Nair, D.P.; Podgorski, M.; Chatani, S.; Gong, T.; Xi, W.X.; Fenoli, C.R.; Bowman, C.N. The Thiol-Michael Addition Click Reaction: A Powerful and Widely Used Tool in Materials Chemistry. Chem. Mater. 2014, 26, 724–744. [Google Scholar] [CrossRef]
- Gayet, R.; Michaud, E.; Nicoli, F.; Chanut, B.; Paul, M.; Rochereau, N.; Guillon, C.; He, Z.; Papagno, L.; Bioley, G.; et al. Impact of IgA isoforms on their ability to activate dendritic cells and to prime T cells. Eur. J. Immunol. 2020, 50, 1295–1306. [Google Scholar] [CrossRef]
- Lissina, A.; Briceno, O.; Afonso, G.; Larsen, M.; Gostick, E.; Price, D.A.; Mallone, R.; Appay, V. Priming of Qualitatively Superior Human Effector CD8+ T Cells Using TLR8 Ligand Combined with FLT3 Ligand. J. Immunol. 2016, 196, 256–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longo, O.; Tripiciano, A.; Fiorelli, V.; Bellino, S.; Scoglio, A.; Collacchi, B.; Alvarez, M.J.; Francavilla, V.; Arancio, A.; Paniccia, G.; et al. Phase I therapeutic trial of the HIV-1 Tat protein and long term follow-up. Vaccine 2009, 27, 3306–3312. [Google Scholar] [CrossRef]
- Skwarczynski, M.; Toth, I. Peptide-based synthetic vaccines. Chem. Sci. 2016, 7, 842–854. [Google Scholar] [CrossRef] [Green Version]
- Tam, J.P. Recent advances in multiple antigen peptides. J. Immunol. Methods 1996, 196, 17–32. [Google Scholar] [CrossRef]
- Niederhafner, P.; Sebestik, J.; Jezek, J. Peptide dendrimers. J. Pept. Sci. 2005, 11, 757–788. [Google Scholar] [CrossRef] [PubMed]
- Schneble, E.; Clifton, G.T.; Hale, D.F.; Peoples, G.E. Peptide-Based Cancer Vaccine Strategies and Clinical Results. Methods Mol. Biol. 2016, 1403, 797–817. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Liu, J.; Jin, S.; Wang, L. Development of tumour peptide vaccines: From universalization to personalization. Scand. J. Immunol. 2020, 91, e12875. [Google Scholar] [CrossRef] [PubMed]
- Mukherji, B.; Chakraborty, N.G.; Yamasaki, S.; Okino, T.; Yamase, H.; Sporn, J.R.; Kurtzman, S.K.; Ergin, M.T.; Ozols, J.; Meehan, J.; et al. Induction of antigen-specific cytolytic T cells in situ in human melanoma by immunization with synthetic peptide-pulsed autologous antigen presenting cells. Proc. Natl. Acad. Sci. USA 1995, 92, 8078–8082. [Google Scholar] [CrossRef] [Green Version]
- Ott, P.A.; Hu, Z.; Keskin, D.B.; Shukla, S.A.; Sun, J.; Bozym, D.J.; Zhang, W.; Luoma, A.; Giobbie-Hurder, A.; Peter, L.; et al. An immunogenic personal neoantigen vaccine for patients with melanoma. Nature 2017, 547, 217–221. [Google Scholar] [CrossRef]
- Hu, Z.; Leet, D.E.; Allesoe, R.L.; Oliveira, G.; Li, S.; Luoma, A.M.; Liu, J.; Forman, J.; Huang, T.; Iorgulescu, J.B.; et al. Personal neoantigen vaccines induce persistent memory T cell responses and epitope spreading in patients with melanoma. Nat. Med. 2021, 27, 515–525. [Google Scholar] [CrossRef]
- Carretero-Iglesia, L.; Couturaud, B.; Baumgaertner, P.; Schmidt, J.; Maby-El Hajjami, H.; Speiser, D.E.; Hebeisen, M.; Rufer, N. High Peptide Dose Vaccination Promotes the Early Selection of Tumor Antigen-Specific CD8 T-Cells of Enhanced Functional Competence. Front. Immunol. 2019, 10, 3016. [Google Scholar] [CrossRef]
- Malonis, R.J.; Lai, J.R.; Vergnolle, O. Peptide-Based Vaccines: Current Progress and Future Challenges. Chem. Rev. 2020, 120, 3210–3229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azuar, A.; Jin, W.; Mukaida, S.; Hussein, W.M.; Toth, I.; Skwarczynski, M. Recent Advances in the Development of Peptide Vaccines and Their Delivery Systems Against Group A Streptococcus. Vaccines 2019, 7, 58. [Google Scholar] [CrossRef] [Green Version]
- Bellini, C.; Horvati, K. Recent Advances in the Development of Protein- and Peptide-Based Subunit Vaccines against Tuberculosis. Cells 2020, 9, 2673. [Google Scholar] [CrossRef]
- Ensoli, B.; Bellino, S.; Tripiciano, A.; Longo, O.; Francavilla, V.; Marcotullio, S.; Cafaro, A.; Picconi, O.; Paniccia, G.; Scoglio, A.; et al. Therapeutic immunization with HIV-1 Tat reduces immune activation and loss of regulatory T-cells and improves immune function in subjects on HAART. PLoS ONE 2010, 5, e13540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ensoli, B.; Nchabeleng, M.; Ensoli, F.; Tripiciano, A.; Bellino, S.; Picconi, O.; Sgadari, C.; Longo, O.; Tavoschi, L.; Joffe, D.; et al. HIV-Tat immunization induces cross-clade neutralizing antibodies and CD4+ T cell increases in antiretroviral-treated South African volunteers: A randomized phase II clinical trial. Retrovirology 2016, 13, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ensoli, F.; Cafaro, A.; Casabianca, A.; Tripiciano, A.; Bellino, S.; Longo, O.; Francavilla, V.; Picconi, O.; Sgadari, C.; Moretti, S.; et al. HIV-1 Tat immunization restores immune homeostasis and attacks the HAART-resistant blood HIV DNA: Results of a randomized phase II exploratory clinical trial. Retrovirology 2015, 12, 33. [Google Scholar] [CrossRef] [Green Version]
- Re, M.C.; Vignoli, M.; Furlini, G.; Gibellini, D.; Colangeli, V.; Vitone, F.; La Placa, M. Antibodies against full-length Tat protein and some low-molecular-weight Tat-peptides correlate with low or undetectable viral load in HIV-1 seropositive patients. J. Clin. Virol. 2001, 21, 81–89. [Google Scholar] [CrossRef]
- Butto, S.; Fiorelli, V.; Tripiciano, A.; Ruiz-Alvarez, M.J.; Scoglio, A.; Ensoli, F.; Ciccozzi, M.; Collacchi, B.; Sabbatucci, M.; Cafaro, A.; et al. Sequence conservation and antibody cross-recognition of clade B human immunodeficiency virus (HIV) type 1 Tat protein in HIV-1-infected Italians, Ugandans, and South Africans. J. Infect. Dis. 2003, 188, 1171–1180. [Google Scholar] [CrossRef]
- Moreau, E.; Belliard, G.; Partidos, C.D.; Pradezinsky, F.; Le Buanec, H.; Muller, S.; Desgranges, C. Important B-cell epitopes for neutralization of human immunodeficiency virus type 1 Tat in serum samples of humans and different animal species immunized with Tat protein or peptides. J. Gen. Virol. 2004, 85, 2893–2901. [Google Scholar] [CrossRef] [PubMed]
- Bachler, B.C.; Humbert, M.; Palikuqi, B.; Siddappa, N.B.; Lakhashe, S.K.; Rasmussen, R.A.; Ruprecht, R.M. Novel biopanning strategy to identify epitopes associated with vaccine protection. J. Virol. 2013, 87, 4403–4416. [Google Scholar] [CrossRef] [Green Version]
- Kurnaeva, M.A.; Sheval, E.V.; Musinova, Y.R.; Vassetzky, Y.S. Tat basic domain: A “Swiss army knife” of HIV-1 Tat? Rev. Med. Virol. 2019, 29, e2031. [Google Scholar] [CrossRef]
- Skwarczynski, M.; Toth, I. Non-invasive mucosal vaccine delivery: Advantages, challenges and the future. Expert Opin. Drug Deliv. 2020, 17, 435–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Construct | PWT | Conjugation Domain | IgG Responses vs. Tat1–20 | IgG Responses vs. Tat Protein |
| Tat1–20 | Peptide alone | NA 1 | 0/5 | 0/5 |
| PWT1-G1 | PWT1 | C-terminus | 3/5 | 2/5 |
| PWT1-G2 | PWT1 | N-terminus | 0/6 | 0/6 |
| PWT2-G1 | PWT2 | C-terminus | 0/6 | 0/6 |
| PWT2-G2 | PWT2 | N-terminus | 2/6 | 3/6 |
| Construct | PWT | Conjugation Domain | IgG Responses vs. Tat46–60 | IgG Responses vs. Tat Protein |
| Tat46–60 | Peptide alone | NA 1 | 1/6 | 1/6 |
| PWT1-G3 | PWT1 | C-terminus | 0/6 | 0/6 |
| PWT1-G4 | PWT1 | N-terminus | 3/6 | 3/6 |
| PWT2-G3 | PWT2 | C-terminus | 5/6 | 5/6 |
| PWT2-G4 | PWT2 | N-terminus | 2/6 | 6/6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nicoli, F.; Pacifico, S.; Gallerani, E.; Marzola, E.; Albanese, V.; Finessi, V.; Llewellyn-Lacey, S.; Price, D.A.; Appay, V.; Marconi, P.; et al. Use of a Novel Peptide Welding Technology Platform for the Development of B- and T-Cell Epitope-Based Vaccines. Vaccines 2021, 9, 526. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9050526
Nicoli F, Pacifico S, Gallerani E, Marzola E, Albanese V, Finessi V, Llewellyn-Lacey S, Price DA, Appay V, Marconi P, et al. Use of a Novel Peptide Welding Technology Platform for the Development of B- and T-Cell Epitope-Based Vaccines. Vaccines. 2021; 9(5):526. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9050526
Chicago/Turabian StyleNicoli, Francesco, Salvatore Pacifico, Eleonora Gallerani, Erika Marzola, Valentina Albanese, Valentina Finessi, Sian Llewellyn-Lacey, David A. Price, Victor Appay, Peggy Marconi, and et al. 2021. "Use of a Novel Peptide Welding Technology Platform for the Development of B- and T-Cell Epitope-Based Vaccines" Vaccines 9, no. 5: 526. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9050526