
Fusion Protein Vaccine Based on Ag85B and STEAP1 Induces a Protective Immune Response against Prostate Cancer

Abstract
:1. Introduction
2. Materials and Methods
2.1. Antigen Expression Analysis with Polymerase Chain Reaction (PCR)
2.2. Western Blotting
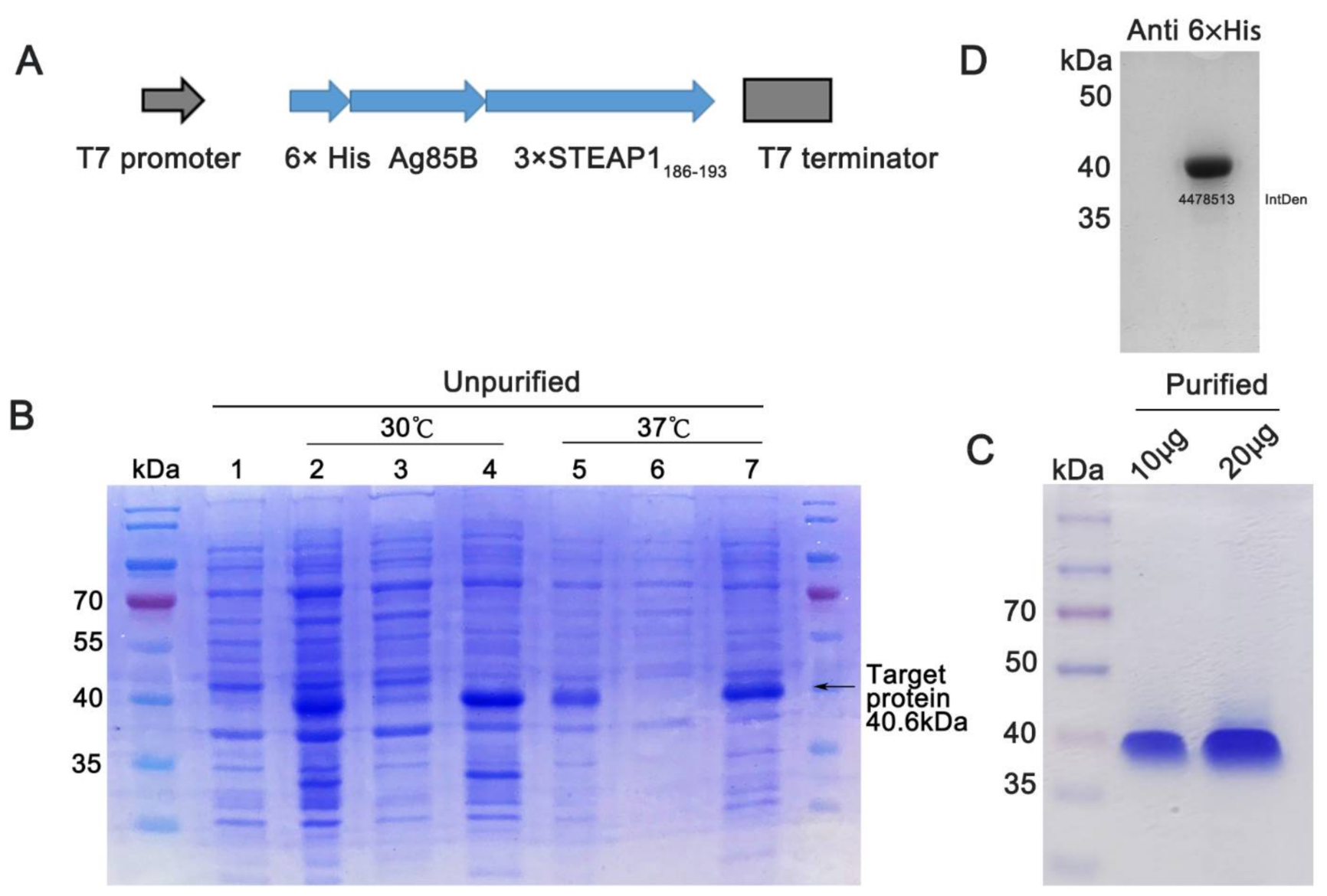
2.3. Expression Plasmid Construct
2.4. Fusion Protein Preparation
2.5. Fusion Protein Analysis with Coomassie Brilliant Blue
2.6. Cells
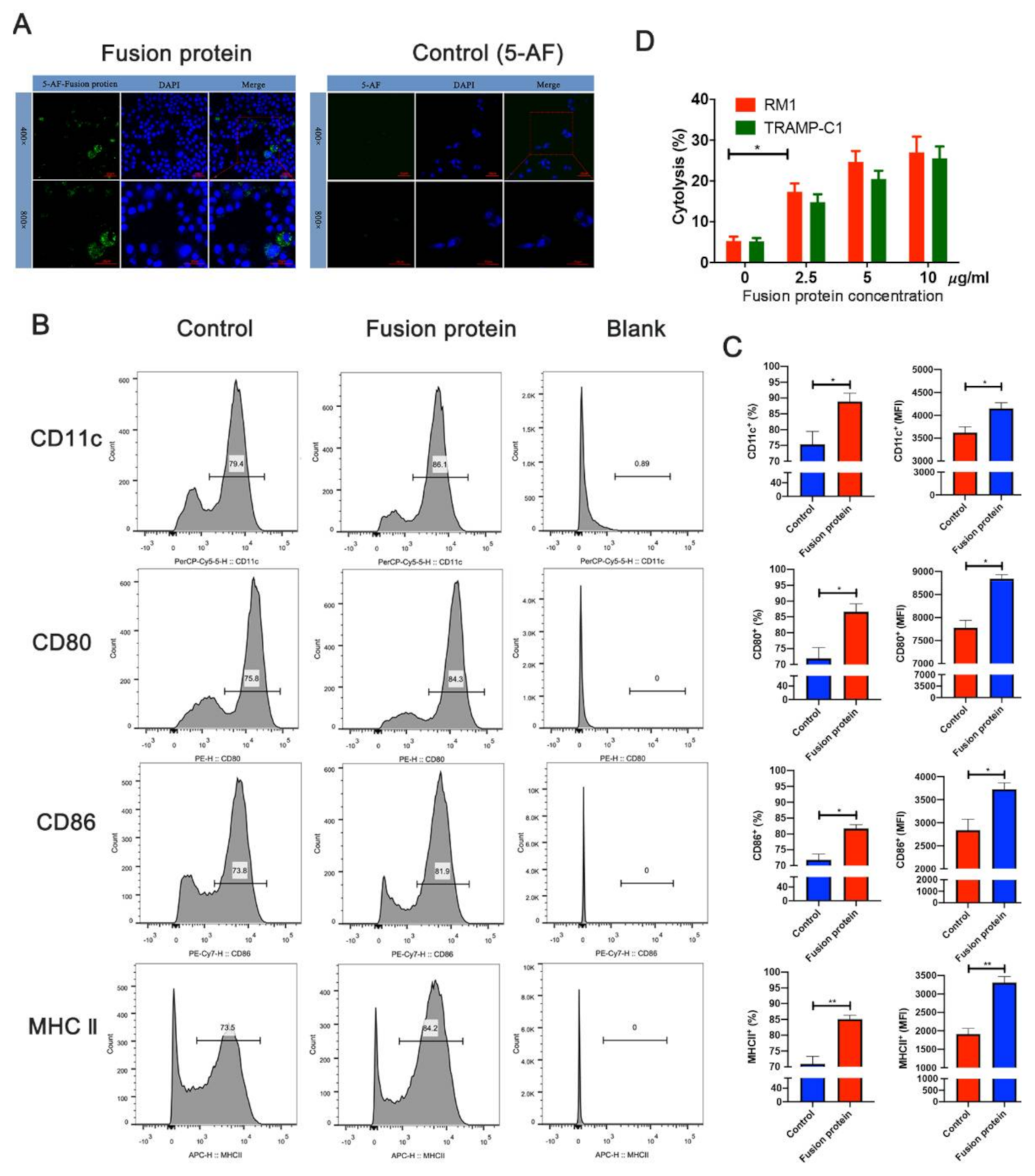
2.7. Fusion Protein Labeled with 5-Aminofluorescein (5-AF)
2.8. Uptake of Fusion Protein by DCs
2.9. Flow Cytometry
2.10. Cytotoxicity Assay
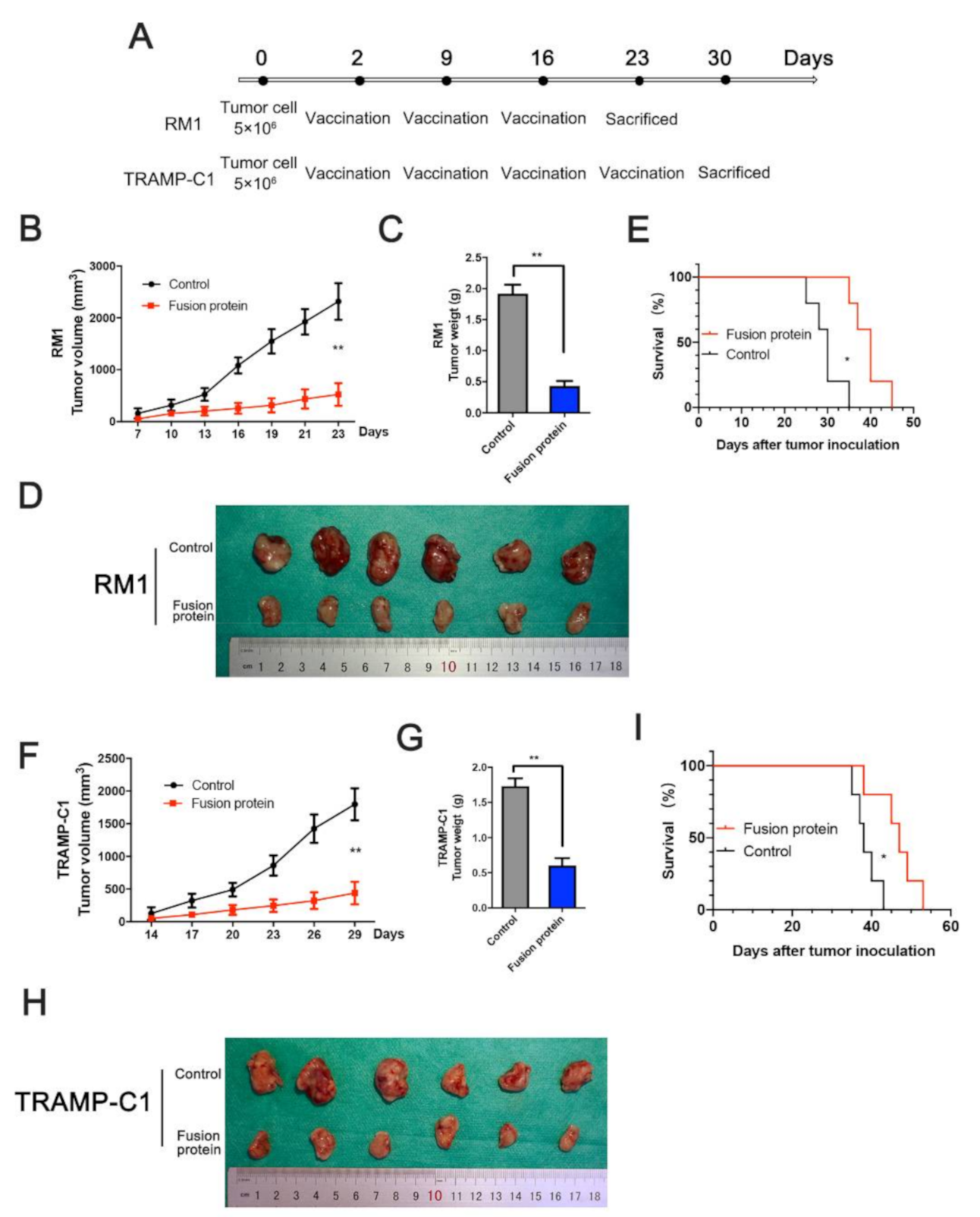
2.11. Mice
2.12. Immunohistochemistry (IHC)
2.13. Separation of Lymphocytes from the Spleen
2.14. T Cell Proliferation Assay
2.15. ELISA
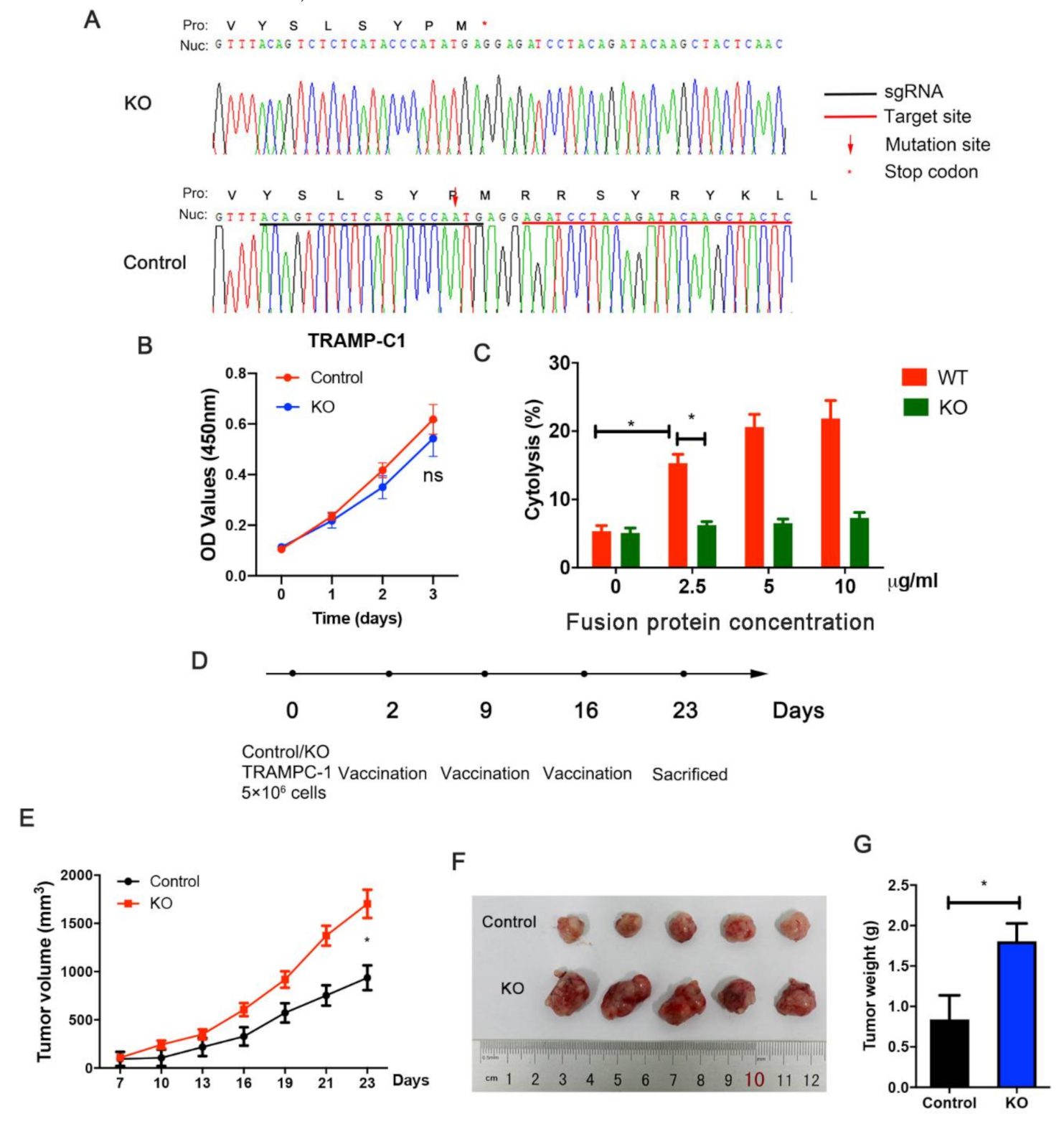
2.16. STEAP1 Knockout Using CRISPR-Cas9
2.17. Statistical Analysis
3. Results
3.1. STEAP1 Is Highly Expressed in Prostate Cancer and Gradually Increases as the Tumor Progresses
3.2. Preparation of Fusion Protein with pET28a
3.3. Fusion Protein Vaccine Activated DC Cells and Elicited Efficient Cytotoxic T Lymphocyte (CTL) Responses In Vitro
3.4. Immunization with Fusion Protein Inhibited Prostate Cancer Cells in Mice
3.5. Fusion Protein Mediated T Cell-Dependent Antitumor Effect
3.6. STEAP1 Knockout Suppresses Immunization Elicited by the Fusion Protein
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| STEAP1 | six-transmembrane epithelial antigen of the prostate 1 |
| CTL | cytotoxic T lymphocyte |
| CRPC | castration-resistant prostate cancer |
| TAA | tumor-associated antigen |
| PIN | prostatic intraepithelial neoplasia |
| LDH | lactate dehydrogenase-releasing cytotoxicity |
| CFSE | 5(6)-carboxyfluorescein diacetate succinimidyl ester |
| IPTG | isopropyl β-D-1-thiogalactopyranoside |
| PCR | polymerase chain reaction |
| CFA | complete Freund’s adjuvant |
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA A Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA A Cancer J. Clin. 2020, 70, 145–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandhi, J.; Afridi, A.; Vatsia, S.; Joshi, G.; Joshi, G.; Kaplan, S.A.; Smith, N.L.; Khan, S.A. The molecular biology of prostate cancer: Current understanding and clinical implications. Prostate Cancer Prostatic Dis. 2017, 21, 22–36. [Google Scholar] [CrossRef] [PubMed]
- Čapoun, O.; Mikulová, V.; Jančíková, M.; Honová, H.; Kološtová, K.; Sobotka, R.; Pešl, M.; Zima, T.; Hanuš, T.; Soukup, V. Prognosis of Castration-resistant Prostate Cancer Patients—Use of the AdnaTest(R) System for Detection of Circulating Tumor Cells. Anticancer Res. 2016, 36, 2019–2026. [Google Scholar]
- Ahmed, S.; Bae, Y.-S. Dendritic cell-based therapeutic cancer vaccines: Past, present and future. Clin. Exp. Vaccine Res. 2014, 3, 113–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maia, C.; Socorro, S.; Schmitt, F.; Santos, C.R.A. STEAP1 is over-expressed in breast cancer and down-regulated by 17β-estradiol in MCF-7 cells and in the rat mammary gland. Endocrine 2008, 34, 108–116. [Google Scholar] [CrossRef]
- Gomes, I.; Maia, C.; Santos, C.R. STEAP Proteins: From Structure to Applications in Cancer Therapy. Mol. Cancer Res. 2012, 10, 573–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulley, J.L.; Madan, R.A.; Tsang, K.Y.; Jochems, C.; Marté, J.L.; Farsaci, B.; Tucker, J.A.; Hodge, J.W.; Liewehr, D.J.; Steinberg, S.M.; et al. Immune Impact Induced by PROSTVAC (PSA-TRICOM), a Therapeutic Vaccine for Prostate Cancer. Cancer Immunol. Res. 2014, 2, 133–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, J.; Pais, J.; Santos, M.M.; Gonçalves, A.M.G.; Gomes, I.M.; Sousa, I.M.; Rocha, S.M.; Passarinha, L.; Maia, C. Targeting STEAP1 Protein in Human Cancer: Current Trends and Future Challenges. Curr. Cancer Drug Targets 2018, 18, 222–230. [Google Scholar] [CrossRef]
- Takatsu, K.; Kariyone, A. The immunogenic peptide for Th1 development. Int. Immunopharmacol. 2003, 3, 783–800. [Google Scholar] [CrossRef]
- Belisle, J.T.; Vissa, V.D.; Sievert, T.; Takayama, K.; Brennan, P.J.; Besra, G. Role of the Major Antigen of Mycobacterium tuberculosis in Cell Wall Biogenesis. Science 1997, 276, 1420–1422. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, R.; Chen, A.; Wang, Y.; Wang, Y.; Zhou, J.; Cao, R. Inhibition of mouse RM-1 prostate cancer and B16F10 melanoma by the fusion protein of HSP65 & STEAP1 186–193. Biomed. Pharmacother. 2019, 111, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Ihlaseh-Catalano, S.M.; A Drigo, S.; De Jesus, C.M.N.; Domingues, M.A.C.; Filho, J.C.S.T.; De Camargo, J.L.V.; Rogatto, S. STEAP1 protein overexpression is an independent marker for biochemical recurrence in prostate carcinoma. Histopathology 2013, 63, 678–685. [Google Scholar] [CrossRef] [PubMed]
- Gomes, I.; Arinto, P.; Lopes, C.; Santos, C.R.; Maia, C.J. STEAP1 is overexpressed in prostate cancer and prostatic intraepithelial neoplasia lesions, and it is positively associated with Gleason score. Urol. Oncol. Semin. Orig. Investig. 2014, 32, 53.e23–53.e29. [Google Scholar] [CrossRef]
- Garcia-Hernandez, M.D.L.L.; Gray, A.; Hubby, B.; Kast, W.M. In vivo Effects of Vaccination with Six-Transmembrane Epithelial Antigen of the Prostate: A Candidate Antigen for Treating Prostate Cancer. Cancer Res. 2007, 67, 1344–1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prendergast, K.A.; Counoupas, C.; Leotta, L.; Eto, C.; Bitter, W.; Winter, N.; Triccas, J.A. The Ag85B protein of the BCG vaccine facilitates macrophage uptake but is dispensable for protection against aerosol Mycobacterium tuberculosis infection. Vaccine 2016, 34, 2608–2615. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.-J.; Bell, H.; Hsieh, C.-L.; Ptak, C.P.; Chang, Y.-F. Novel Mycobacteria Antigen 85 Complex Binding Motif on Fibronectin. J. Biol. Chem. 2012, 287, 1892–1902. [Google Scholar] [CrossRef] [Green Version]
- Peake, P.; Gooley, A.; Britton, W.J. Mechanism of interaction of the 85B secreted protein of Mycobacterium bovis with fibronectin. Infect. Immun. 1993, 61, 4828–4834. [Google Scholar] [CrossRef] [Green Version]
- Abou-Zeid, C.; Ratliff, T.L.; Wiker, H.G.; Harboe, M.; Bennedsen, J.; A Rook, G. Characterization of fibronectin-binding antigens released by Mycobacterium tuberculosis and Mycobacterium bovis BCG. Infect. Immun. 1988, 56, 3046–3051. [Google Scholar] [CrossRef] [Green Version]
- Proctor, R.A. Fibronectin: An Enhancer of Phagocyte Function. Clin. Infect. Dis. 1987, 9, S412–S419. [Google Scholar] [CrossRef]
- Fan, X.; Gao, Q.; Fu, R. Differential immunogenicity and protective efficacy of DNA vaccines expressing proteins of Mycobacterium tuberculosis in a mouse model. Microbiol. Res. 2009, 164, 374–382. [Google Scholar] [CrossRef]
- Kariyone, A.; Tamura, T.; Kano, H.; Iwakura, Y.; Takeda, K.; Akira, S.; Takatsu, K. Immunogenicity of Peptide-25 of Ag85B in Th1 development: Role of IFN-γ. Int. Immunol. 2003, 15, 1183–1194. [Google Scholar] [CrossRef] [Green Version]
- Palma, C.; Iona, E.; Giannoni, F.; Pardini, M.; Brunori, L.; Orefici, G.; Fattorini, L.; Cassone, A. The Ag85B protein of Mycobacterium tuberculosis may turn a protective immune response induced by Ag85B-DNA vaccine into a potent but non-protective Th1immune response in mice. Cell. Microbiol. 2007, 9, 1455–1465. [Google Scholar] [CrossRef] [PubMed]
- Aung, H.; Toossi, Z.; Wisnieski, J.J.; Wallis, R.S.; A Culp, L.; Phillips, N.B.; Phillips, M.; E Averill, L.; Daniel, T.M.; Ellner, J.J. Induction of monocyte expression of tumor necrosis factor alpha by the 30-kD alpha antigen of Mycobacterium tuberculosis and synergism with fibronectin. J. Clin. Investig. 1996, 98, 1261–1268. [Google Scholar] [CrossRef]
- Kuromatsu, I.; Matsuo, K.; Takamura, S.; Kim, G.; Takebe, Y.; Kawamura, J.; Yasutomi, Y. Induction of effective antitumor immune responses in a mouse bladder tumor model by using DNA of an α antigen from mycobacteria. Cancer Gene Ther. 2001, 8, 483–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roche, P.W.; Peake, P.W.; Billman-Jacobe, H.; Doran, T.; Britton, W.J. T-cell determinants and antibody binding sites on the major mycobacterial secretory protein MPB59 of Mycobacterium bovis. Infect. Immun. 1994, 62, 5319–5326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mustafa, A.S.; Shaban, F.A.; Abal, A.T.; Al-Attiyah, R.; Wiker, H.G.; Lundin, K.E.A.; Oftung, F.; Huygen, K. Identification and HLA Restriction of Naturally Derived Th1-Cell Epitopes from the Secreted Mycobacterium tuberculosis Antigen 85B Recognized by Antigen-Specific Human CD4 + T-Cell Lines. Infect. Immun. 2000, 68, 3933–3940. [Google Scholar] [CrossRef] [Green Version]
- Huygen, K.; Abramowicz, D.; Vandenbussche, P.; Jacobs, F.; De Bruyn, J.; Kentos, A.; Drowart, A.; Van Vooren, J.P.; Goldman, M. Spleen cell cytokine secretion in Mycobacterium bovis BCG-infected mice. Infect. Immun. 1992, 60, 2880–2886. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Ji, G.; Xie, J.; Jiao, Z.; Zhang, H.; Chen, J. Six-transmembrane epithelial antigen of the prostate 1 is associated with tumor invasion and migration in endometrial carcinomas. J. Cell. Biochem. 2019, 120, 11172–11189. [Google Scholar] [CrossRef]
- Hayashi, S.; Kumai, T.; Matsuda, Y.; Aoki, N.; Sato, K.; Kimura, S.; Kitada, M.; Tateno, M.; Celis, E.; Kobayashi, H. Six-transmembrane epithelial antigen of the prostate and enhancer of zeste homolog 2 as immunotherapeutic targets for lung cancer. J. Transl. Med. 2011, 9, 191. [Google Scholar] [CrossRef] [Green Version]
- Valenti, M.T.; Carbonare, L.D.; Donatelli, L.; Bertoldo, F.; Giovanazzi, B.; Caliari, F.; Cascio, V.L. STEAP mRNA detection in serum of patients with solid tumours. Cancer Lett. 2009, 273, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Burnell, S.E.A.; Spencer-Harty, S.; Howarth, S.; Bodger, O.; Kynaston, H.; Morgan, C.; Doak, S.H. Utilisation of the STEAP protein family in a diagnostic setting may provide a more comprehensive prognosis of prostate cancer. PLoS ONE 2019, 14, e0220456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azumi, M.; Kobayashi, H.; Aoki, N.; Sato, K.; Kimura, S.; Kakizaki, H.; Tateno, M. Six-Transmembrane Epithelial Antigen of the Prostate as an Immunotherapeutic Target for Renal Cell and Bladder Cancer. J. Urol. 2010, 183, 2036–2044. [Google Scholar] [CrossRef] [PubMed]
- Manzo, T.; Michelini, R.H.; Basso, V.; Ricupito, A.; Chai, J.-G.; Simpson, E.; Bellone, M.; Mondino, A. Concurrent Allorecognition Has a Limited Impact on Posttransplant Vaccination. J. Immunol. 2011, 186, 1361–1368. [Google Scholar] [CrossRef] [PubMed]
- Atherton, M.J.; Stephenson, K.B.; Tzelepis, F.; Bakhshinyan, D.; Nikota, J.K.; Son, H.H.; Jirovec, A.; Lefebvre, C.; Dvorkin-Gheva, A.; Ashkar, A.A.; et al. Transforming the prostatic tumor microenvironment with oncolytic virotherapy. OncoImmunology 2018, 7, e1445459. [Google Scholar] [CrossRef] [Green Version]
- Cappuccini, F.; Stribbling, S.; Pollock, E.; Hill, A.; Redchenko, I. Immunogenicity and efficacy of the novel cancer vaccine based on simian adenovirus and MVA vectors alone and in combination with PD-1 mAb in a mouse model of prostate cancer. Cancer Immunol. Immunother. 2016, 65, 701–713. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Lee, J.-B.; Lee, G.K.; Chang, J. Vaccination with recombinant adenoviruses and dendritic cells expressing prostate-specific antigens is effective in eliciting CTL and suppresses tumor growth in the experimental prostate cancer. Prostate 2009, 69, 938–948. [Google Scholar] [CrossRef]
- Zaidi, M.R. The Interferon-Gamma Paradox in Cancer. J. Interf. Cytokine Res. 2019, 39, 30–38. [Google Scholar] [CrossRef]
- Castro, F.; Cardoso, A.P.; Goncalves, R.M.; Serre, K.; Oliveira, M.J. Interferon-Gamma at the Crossroads of Tumor Immune Surveillance or Evasion. Front. Immunol. 2018, 9, 847. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Kohli, K.; Black, R.; Yao, L.; Spadinger, S.M.; He, Q.; Pillarisetty, V.G.; Cranmer, L.D.; Van Tine, B.A.; Yee, C.; et al. Systemic Interferon-γ Increases MHC Class I Expression and T-cell Infiltration in Cold Tumors: Results of a Phase 0 Clinical Trial. Cancer Immunol. Res. 2019, 7, 1237–1243. [Google Scholar] [CrossRef] [Green Version]
- Ong, C.E.B.; Lyons, A.B.; Woods, G.M.; Flies, A.S. Inducible IFN-γ Expression for MHC-I Upregulation in Devil Facial Tumor Cells. Front. Immunol. 2019, 9, 3117. [Google Scholar] [CrossRef] [Green Version]
- Zhou, F. Molecular Mechanisms of IFN-γ to Up-Regulate MHC Class I Antigen Processing and Presentation. Int. Rev. Immunol. 2009, 28, 239–260. [Google Scholar] [CrossRef] [PubMed]
- Nappo, G.; Handle, F.; Santer, F.R.; McNeill, R.V.; I Seed, R.; Collins, A.T.; Morrone, G.; Culig, Z.; Maitland, N.J.; Erb, H.H.H. The immunosuppressive cytokine interleukin-4 increases the clonogenic potential of prostate stem-like cells by activation of STAT6 signalling. Oncogenesis 2017, 6, e342. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Pang, Y.; Gara, S.K.; Achyut, B.; Heger, C.; Goldsmith, P.K.; Lonning, S.; Yang, L. Gr-1+CD11b+ cells are responsible for tumor promoting effect of TGF-β in breast cancer progression. Int. J. Cancer 2012, 131, 2584–2595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trikha, P.; Carson, W.E. Signaling pathways involved in MDSC regulation. Biochim. Biophys. Acta Bioenerg. 2014, 1846, 55–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Name | Primer Sequences |
|---|---|---|
| STEAP1 | F1 | AGTCGCTGCCATCATATCATCC |
| R1 | AAGTCGGAGGCATGTACCATAC | |
| Ag85B + link | F2 | CGGCAGCCATATGGCTAGCATGACAGACGTGAGCCGAAAGA |
| R2 | GTAGGAACGTGATCCTCCACCTCCACCGC | |
| R3 | GGTGGTGGTGGTGCTCGAGCTACAGCAGTTTGTAACGGTAGGAACGC | |
| GAPDH | F4 | CATCACTGCCACCCAGAAGACTG |
| R4 | ATGCCAGTGAGCTTCCCGTTCAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, L.; Xie, H.; Zhang, Z.; Wang, Z.; Peng, S.; Niu, Y.; Shang, Z. Fusion Protein Vaccine Based on Ag85B and STEAP1 Induces a Protective Immune Response against Prostate Cancer. Vaccines 2021, 9, 786. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9070786
Guo L, Xie H, Zhang Z, Wang Z, Peng S, Niu Y, Shang Z. Fusion Protein Vaccine Based on Ag85B and STEAP1 Induces a Protective Immune Response against Prostate Cancer. Vaccines. 2021; 9(7):786. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9070786
Chicago/Turabian StyleGuo, Linpei, Hui Xie, Zheng Zhang, Zhun Wang, Shuanghe Peng, Yuanjie Niu, and Zhiqun Shang. 2021. "Fusion Protein Vaccine Based on Ag85B and STEAP1 Induces a Protective Immune Response against Prostate Cancer" Vaccines 9, no. 7: 786. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9070786