
A Pseudomonas aeruginosa-Derived Particulate Vaccine Protects against P. aeruginosa Infection

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
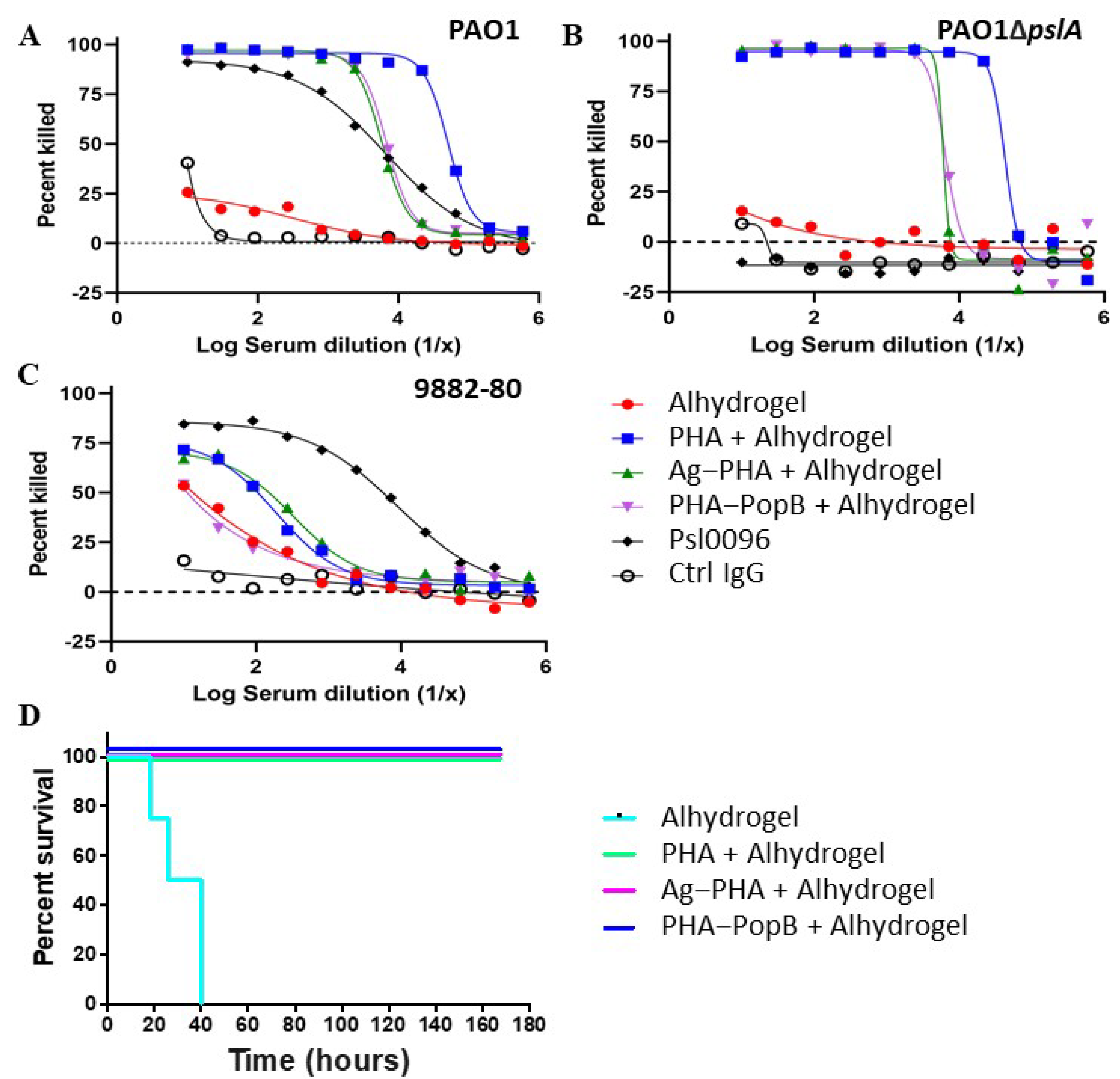{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Plasmid Construction for In Vivo Production of PHA Beads
2.3. PHA Bead Isolation and Purification
2.4. PHA Bead Characterization
2.5. Analysis of the Selected Epitopes/Antigens and HCPs Coating the PHA Beads
2.6. Mouse Vaccination and Challenge
2.7. Serum Antibody Analysis
2.8. Statistical Analysis
3. Results
3.1. Evaluation of the PHA Beads as Particulate Vaccines
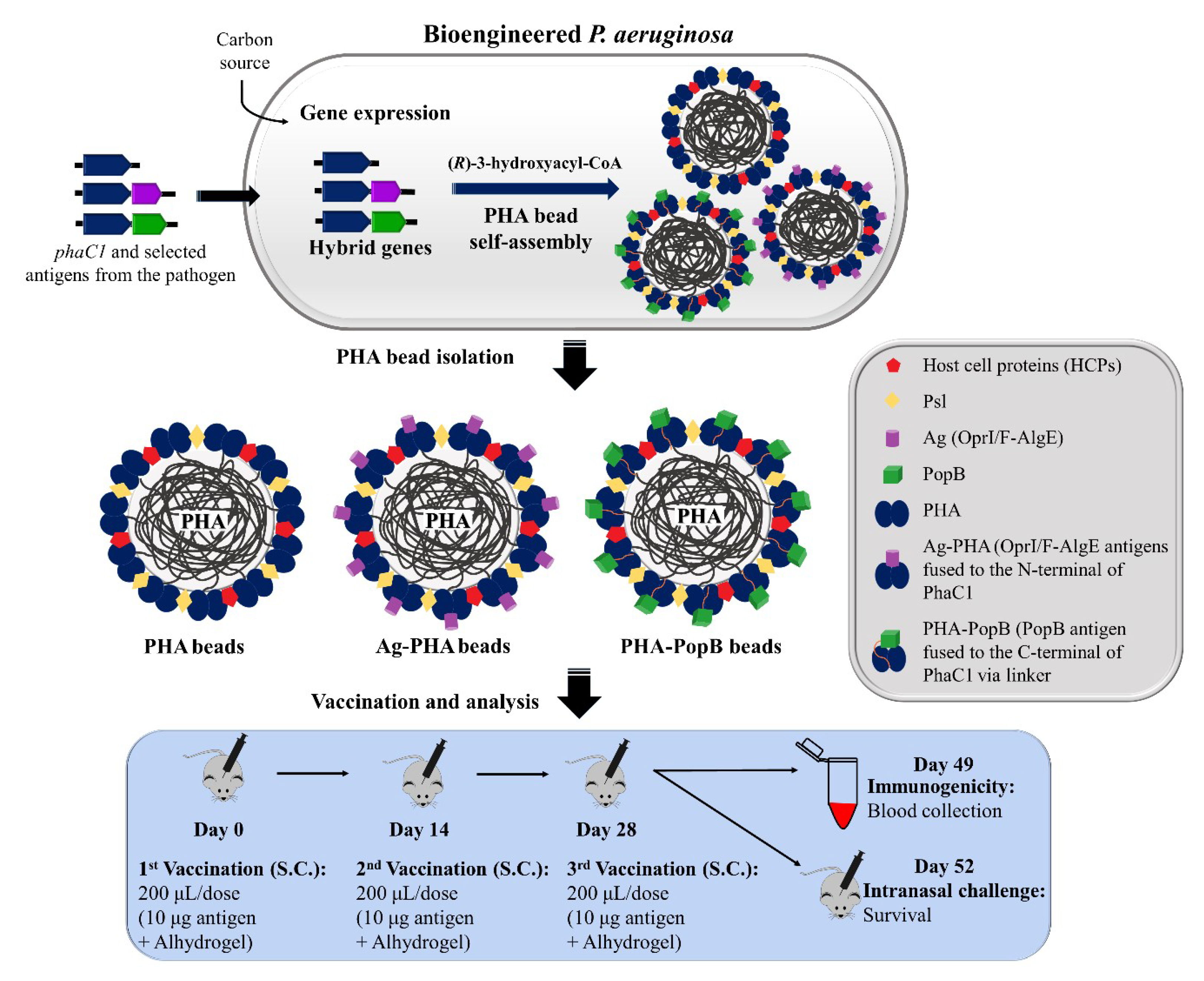
3.2. Bioengineering P. aeruginosa for the Production of PHA Bead Vaccines
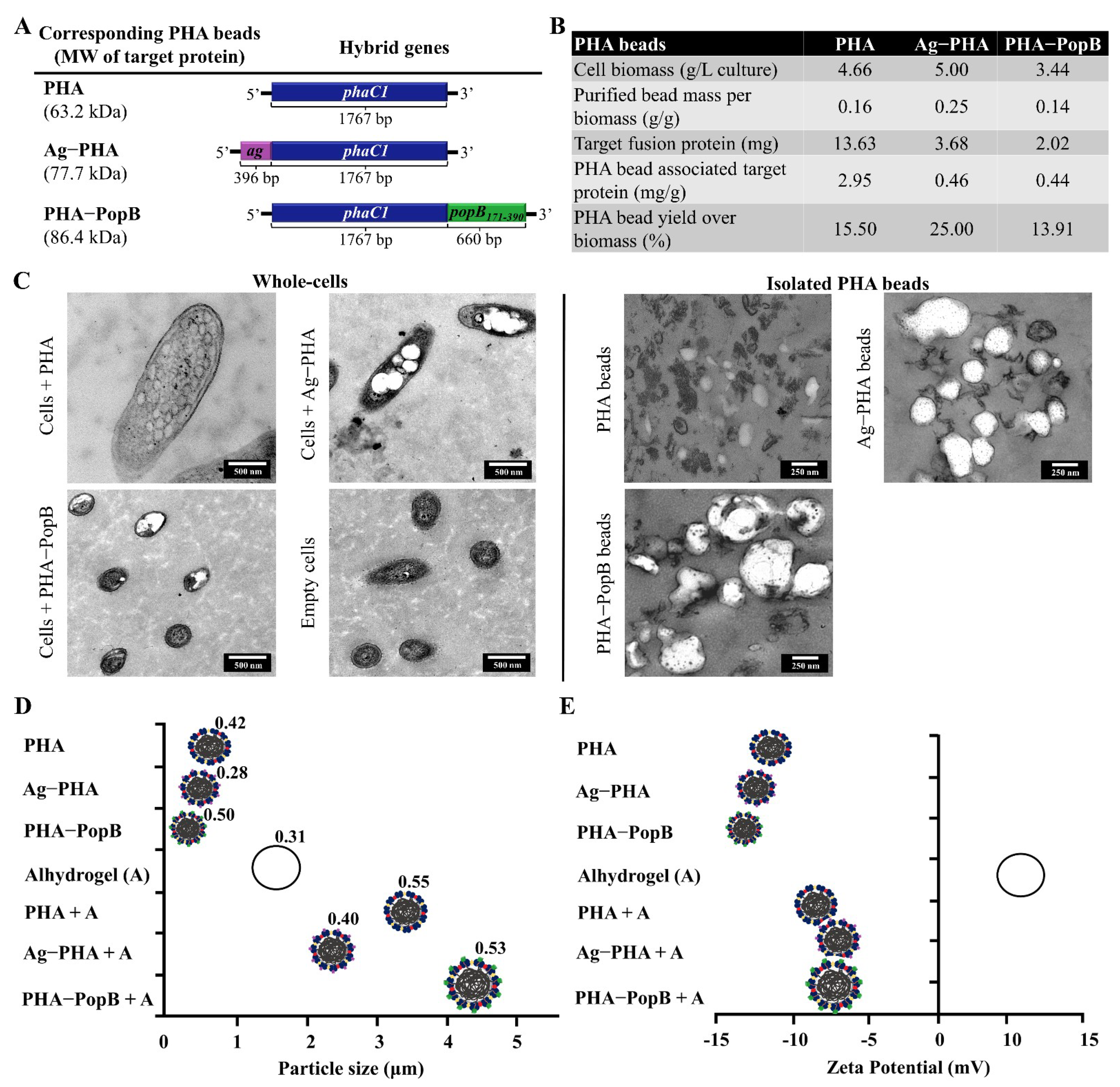
3.3. Biophysical Characterization of PHA Bead Vaccines
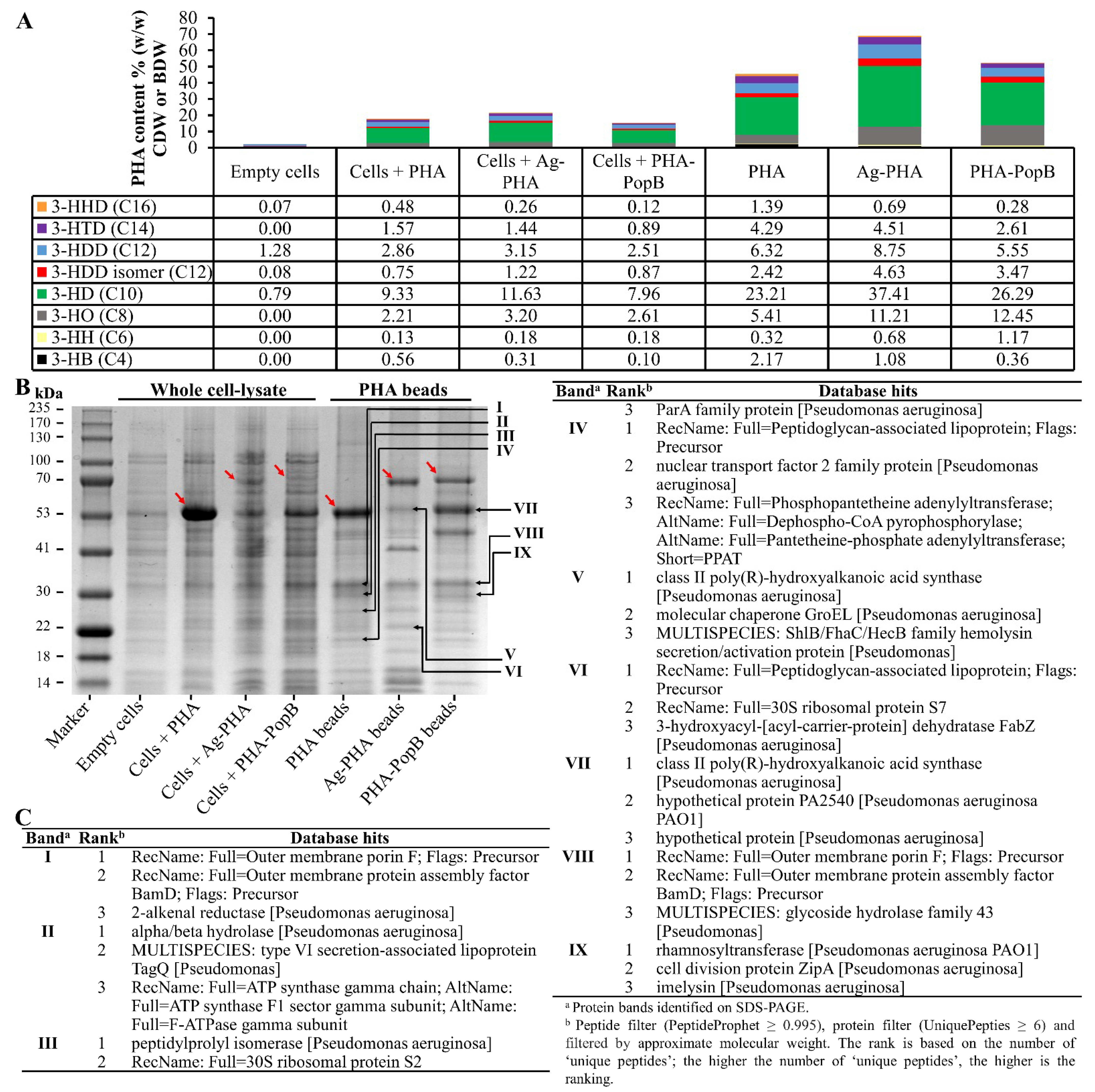
3.4. Biochemical Characterization of PHA Bead Vaccines
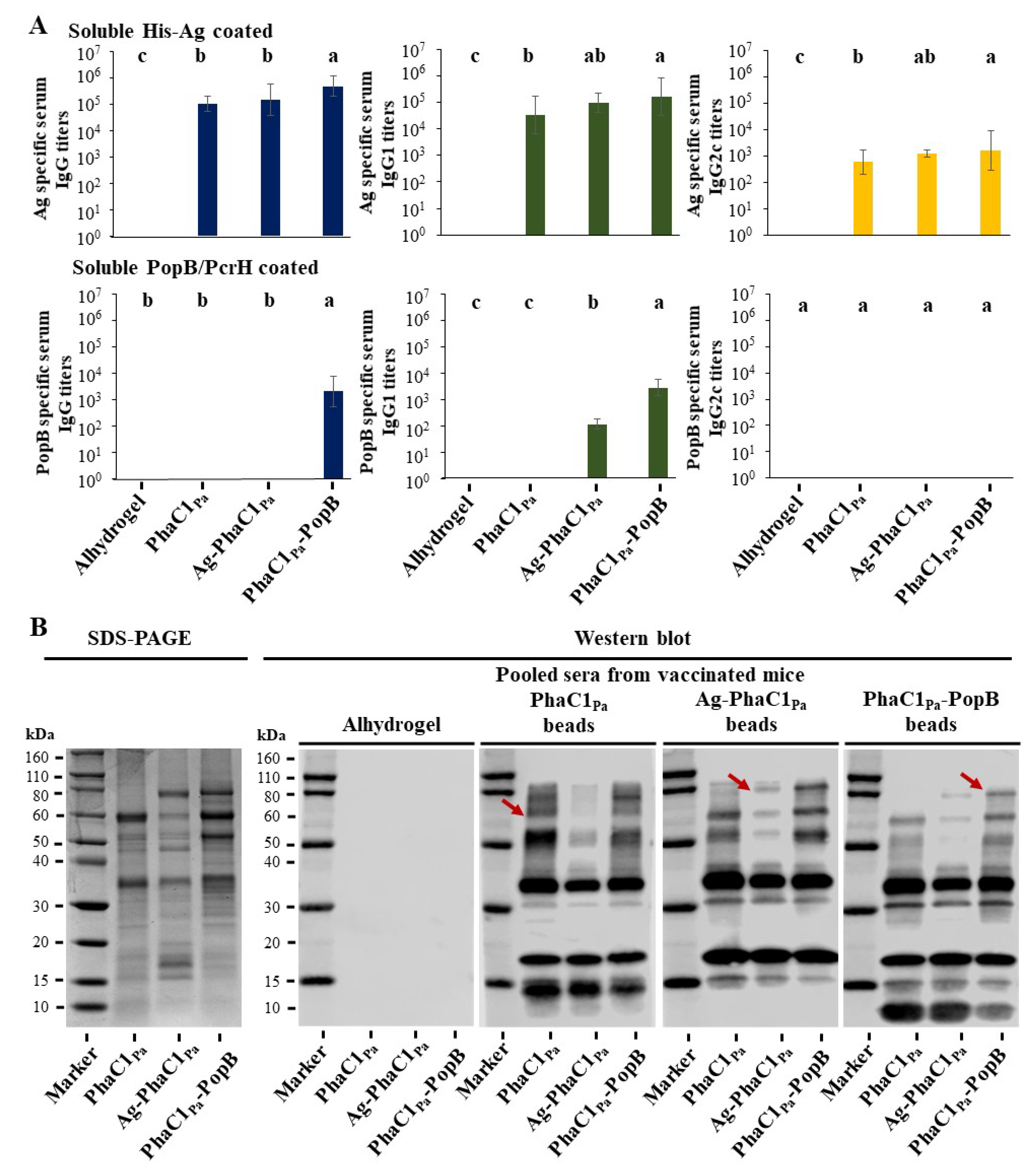
3.5. Vaccine Performance of PHA Beads
3.6. Opsonophagocytic Assay Shows Induction of Functional Antibodies
3.7. PHA Bead Vaccine Formulations Induce Protective Immunity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharma, A.; Krause, A.; Worgall, S. Recent developments for Pseudomonas vaccines. Hum. Vaccines 2011, 7, 999–1011. [Google Scholar] [CrossRef] [Green Version]
- Grimwood, K.; Kyd, J.M.; Owen, S.J.; Massa, H.M.; Cripps, A.W. Vaccination against respiratory Pseudomonas aeruginosa infection. Hum. Vacc. Immunother. 2015, 11, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Moradali, M.F.; Ghods, S.; Rehm, B.H.A. Pseudomonas aeruginosa Lifestyle: A Paradigm for Adaptation, Survival, and Persistence. Front. Cell. Infect. Microbiol. 2017, 7, 39. [Google Scholar] [CrossRef] [Green Version]
- Thi, M.T.T.; Wibowo, D.; Rehm, B.H.A. Pseudomonas aeruginosa Biofilms. Int. J. Mol. Sci. 2020, 21, 8671. [Google Scholar] [CrossRef]
- Brooun, A.; Liu, S.; Lewis, K. A Dose-Response Study of Antibiotic Resistance in Pseudomonas aeruginosa Biofilms. Antimicrob. Agents Chemother. 2000, 44, 640–646. [Google Scholar] [CrossRef] [Green Version]
- Ghafoor, A.; Hay, I.; Rehm, B.H.A. Role of Exopolysaccharides in Pseudomonas aeruginosa Biofilm Formation and Architecture. Appl. Environ. Microbiol. 2011, 77, 5238–5246. [Google Scholar] [CrossRef] [Green Version]
- Di Giandomenico, A.; Warrener, P.; Hamilton, M.; Guillard, S.; Ravn, P.; Minter, R.; Camara, M.M.; Venkatraman, V.; MacGill, R.S.; Lin, J.; et al. Identification of broadly protective human antibodies to Pseudomonas aeruginosa exopolysaccharide Psl by phenotypic screening. J. Exp. Med. 2012, 209, 1273–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merakou, C.; Schaefers, M.; Priebe, G.P. Progress Toward the ElusivePseudomonas aeruginosaVaccine. Surg. Infect. 2018, 19, 757–768. [Google Scholar] [CrossRef] [PubMed]
- Gellatly, S.L.; Hancock, R.E.W. Pseudomonas aeruginosa: New insights into pathogenesis and host defenses. Pathog. Dis. 2013, 67, 159–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priebe, G.P.; Goldberg, J.B. Vaccines for Pseudomonas aeruginosa: A long and winding road. Expert Rev. Vaccines 2014, 13, 507–519. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.W.; Parlane, N.A.; Wedlock, N.; Rehm, B.H.A. Bioengineering a bacterial pathogen to assemble its own particulate vaccine capable of inducing cellular immunity. Sci. Rep. 2017, 7, srep41607. [Google Scholar] [CrossRef] [Green Version]
- Peters, V.; Rehm, B.H.A. In Vivo Enzyme Immobilization by Use of Engineered Polyhydroxyalkanoate Synthase. Appl. Environ. Microbiol. 2006, 72, 1777–1783. [Google Scholar] [CrossRef] [Green Version]
- Parlane, N.A.; Grage, K.; Mifune, J.; Basaraba, R.J.; Wedlock, N.; Rehm, B.; Buddle, B.M. Vaccines Displaying Mycobacterial Proteins on Biopolyester Beads Stimulate Cellular Immunity and Induce Protection against Tuberculosis. Clin. Vaccine Immunol. 2012, 19, 37–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Miro, M.; Rodríguez-Noda, L.; Fariñas-Medina, M.; García-Rivera, D.; Vérez-Bencomo, V.; Rehm, B.H. Self-assembled particulate PsaA as vaccine against Streptococcus pneumoniae infection. Heliyon 2017, 3, e00291. [Google Scholar] [CrossRef] [Green Version]
- González-Miró, M.; Rodríguez-Noda, L.M.; Fariñas-Medina, M.; Cedré-Marrero, B.; Madariaga-Zarza, S.; Zayas-Vignier, C.; Hernández-Cedeño, M.; Kleffmann, T.; García-Rivera, D.; Vérez-Bencomo, V.; et al. Bioengineered polyester beads co-displaying protein and carbohydrate-based antigens induce protective immunity against bacterial infection. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Donato, G.; Piniella, B.; Aguilar, D.; Olivera, S.; Pérez, A.; Castañedo, Y.; Alvarez-Lajonchere, L.; Dueñas-Carrera, S.; Lee, J.W.; Burr, N.; et al. Protective T Cell and Antibody Immune Responses against Hepatitis C Virus Achieved Using a Biopolyester-Bead-Based Vaccine Delivery System. Clin. Vaccine Immunol. 2016, 23, 370–378. [Google Scholar] [CrossRef] [Green Version]
- Moradali, M.F.; Rehm, B.H.A. Bacterial biopolymers: From pathogenesis to advanced materials. Nat. Rev. Microbiol. 2020, 18, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Wibowo, D.; Jorritsma, S.H.; Gonzaga, Z.J.; Evert, B.; Chen, S.; Rehm, B.H. Polymeric nanoparticle vaccines to combat emerging and pandemic threats. Biomaterials 2021, 268, 120597. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Huang, J.; Duan, B.; Traficante, D.C.; Hong, H.; Risech, M.; Lory, S.; Priebe, G.P. Th17-stimulating Protein Vaccines Confer Protection againstPseudomonas aeruginosaPneumonia. Am. J. Respir. Crit. Care Med. 2012, 186, 420–427. [Google Scholar] [CrossRef] [Green Version]
- Schaefers, M.M.; Duan, B.; Mizrahi, B.; Lu, R.; Reznor, G.; Kohane, D.S.; Priebe, G.P. PLGA-encapsulation of the Pseudomonas aeruginosa PopB vaccine antigen improves Th17 responses and confers protection against experimental acute pneumonia. Vaccine 2018, 36, 6926–6932. [Google Scholar] [CrossRef]
- Baumann, U.; Mansouri, E.; Von Specht, B.-U.; Baumann, U.; Mansouri, E.; Von Specht, B.-U. Recombinant OprF–OprI as a vaccine against Pseudomonas aeruginosa infections. Vaccine 2004, 22, 840–847. [Google Scholar] [CrossRef]
- Westritschnig, K.; Hochreiter, R.; Wallner, G.; Firbas, C.; Schwameis, M.; Jilma, B. A randomized, placebo-controlled phase I study assessing the safety and immunogenicity of a Pseudomonas aeruginosa hybrid outer membrane protein OprF/I vaccine (IC43) in healthy volunteers. Hum. Vaccines Immunother. 2014, 10, 170–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Z.; Han, D.; Sun, X.; Zhang, M.; Feng, X.; Sun, C.; Gu, J.; Tong, C.; Lei, L.; Han, W. Mannose-modified chitosan microspheres enhance OprF-OprI-mediated protection of mice against Pseudomonas aeruginosa infection via induction of mucosal immunity. Appl. Microbiol. Biotechnol. 2015, 99, 667–680. [Google Scholar] [CrossRef] [PubMed]
- Weimer, E.; Lu, H.; Kock, N.D.; Wozniak, D.J.; Mizel, S.B. A Fusion Protein Vaccine Containing OprF Epitope 8, OprI, and Type A and B Flagellins Promotes Enhanced Clearance of Nonmucoid Pseudomonas aeruginosa. Infect. Immun. 2009, 77, 2356–2366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehm, B.; Boheim, G.; Tommassen, J.; Winkler, U.K. Overexpression of algE in Escherichia coli: Subcellular localization, purification, and ion channel properties. J. Bacteriol. 1994, 176, 5639–5647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehm, B.H.; Grabert, E.; Hein, J.; Winkler, U.K. Antibody response of rabbits and cystic fibrosis patients to an alginate-specific outer membrane protein of a mucoid strain of Pseudomonas aeruginosa. Microb. Pathog. 1994, 16, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Choi, K.-H.; Kumar, A.; Schweizer, H.P. A 10-min method for preparation of highly electrocompetent Pseudomonas aeruginosa cells: Application for DNA fragment transfer between chromosomes and plasmid transformation. J. Microbiol. Methods 2006, 64, 391–397. [Google Scholar] [CrossRef]
- Spiekermann, P.; Rehm, B.; Kalscheuer, R.; Baumeister, D.; Steinbüchel, A. A sensitive, viable-colony staining method using Nile red for direct screening of bacteria that accumulate polyhydroxyalkanoic acids and other lipid storage compounds. Arch. Microbiol. 1999, 171, 73–80. [Google Scholar] [CrossRef]
- Evert, B.; Vezina, B.; Rehm, B.H.A. Catalytically Active Bioseparation Resin Utilizing a Covalent Intermediate for Tagless Protein Purification. ACS Appl. Bio Mater. 2020, 3, 8911–8922. [Google Scholar] [CrossRef]
- Brandl, H.; Gross, R.A.; Lenz, R.W.; Fuller, R.C. Pseudomonas oleovorans as a Source of Poly(β-Hydroxyalkanoates) for Potential Applications as Biodegradable Polyesters. Appl. Environ. Microbiol. 1988, 54, 1977–1982. [Google Scholar] [CrossRef] [Green Version]
- Rubio Reyes, P.; Parlane, N.A.; Wedlock, D.N.; Rehm, B.H.A. Immunogencity of antigens from Mycobacterium tuberculosis self-assembled as particulate vaccines. Int. J. Med. Microbiol. 2016, 306, 624–632. [Google Scholar] [CrossRef]
- Hay, I.D.; Hooks, D.O.; Rehm, B.H.A. Use of Bacterial Polyhydroxyalkanoates in Protein Display Technologies. In Hydrocarbon and Lipid Microbiology Protocols; Springer: Berlin/Heidelberg, Germany, 2014; pp. 71–86. [Google Scholar]
- Shevchenko, A.; Tomas, H.; Havlis, J.; Olsen, J.; Mann, M.J. In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 2006, 1, 2856–2860. [Google Scholar] [CrossRef] [PubMed]
- Hatano, K.; Boisot, S.; DesJardins, D.; Wright, D.C.; Brisker, J.; Pier, G.B.; Hatano, K.; Boisot, S.; DesJardins, D.; Wright, D.C.; et al. Immunogenic and antigenic properties of a heptavalent high-molecular-weight O-polysaccharide vaccine derived from Pseudomonas aeruginosa. Infect. Immun. 1994, 62, 3608–3616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priebe, G.P.; Brinig, M.M.; Hatano, K.; Grout, M.; Coleman, F.T.; Pier, G.B.; Goldberg, J.B. Construction and Characterization of a Live, Attenuated aroA Deletion Mutant of Pseudomonas aeruginosa as a Candidate Intranasal Vaccine. Infect. Immun. 2002, 70, 1623–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, K.-H.; Gaynor, J.B.; White, K.G.; Lopez, C.; Bosio, C.M.; Karkhoff-Schweizer, R.R.; Schweizer, H.P. A Tn 7-based broad-range bacterial cloning and expression system. Nat. Methods 2005, 2, 443–448. [Google Scholar] [CrossRef]
- Hoffmann, N.; Rehm, B.H. Regulation of polyhydroxyalkanoate biosynthesis in Pseudomonas putida and Pseudomonas aeruginosa. FEMS Microbiol. Lett. 2004, 237, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Draper, J.L.; Rehm, B.H. Engineering bacteria to manufacture functionalized polyester beads. Bioengineered 2012, 3, 203–208. [Google Scholar] [CrossRef]
- Pham, T.H.; Webb, J.; Rehm, B.H.A. The role of polyhydroxyalkanoate biosynthesis by Pseudomonas aeruginosa in rhamnolipid and alginate production as well as stress tolerance and biofilm formation. Microbiology 2004, 150, 3405–3413. [Google Scholar] [CrossRef] [Green Version]
- Discola, K.F.; Förster, A.; Boulay, F.; Simorre, J.-P.; Attree, I.; Dessen, A.; Job, V. Membrane and Chaperone Recognition by the Major Translocator Protein PopB of the Type III Secretion System of Pseudomonas aeruginosa. J. Biol. Chem. 2014, 289, 3591–3601. [Google Scholar] [CrossRef] [Green Version]
- Dey, S.; Basu, A.; Datta, S. Characterization of Molten Globule PopB in Absence and Presence of Its Chaperone PcrH. Protein J. 2012, 31, 401–416. [Google Scholar] [CrossRef]
- Das, S.; Howlader, D.R.; Zheng, Q.; Ratnakaram, S.S.K.; Whittier, S.K.; Lu, T.; Keith, J.D.; Picking, W.D.; Birket, S.E.; Picking, W.L. Development of a Broadly Protective, Self-Adjuvanting Subunit Vaccine to Prevent Infections by Pseudomonas aeruginosa. Front. Immunol. 2020, 11, 583008. [Google Scholar] [CrossRef]
- Grage, K.; Jahns, A.C.; Parlane, N.; Palanisamy, R.; Rasiah, I.A.; Atwood, J.A.; Rehm, B. Bacterial Polyhydroxyalkanoate Granules: Biogenesis, Structure, and Potential Use as Nano-/Micro-Beads in Biotechnological and Biomedical Applications. Biomacromolecules 2009, 10, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Wang, W. Factors affecting alum–protein interactions. Int. J. Pharm. 2014, 466, 139–146. [Google Scholar] [CrossRef]
- Rehm, B.H. Bioengineering towards self-assembly of particulate vaccines. Curr. Opin. Biotechnol. 2017, 48, 42–53. [Google Scholar] [CrossRef] [PubMed]
- De Temmerman, M.-L.; Rejman, J.; Demeester, J.; Irvine, D.J.; Gander, B.; De Smedt, S.C. Particulate vaccines: On the quest for optimal delivery and immune response. Drug Discov. Today 2011, 16, 569–582. [Google Scholar] [CrossRef]
- Ackerman, A.L.; Kyritsis, C.; Tampé, R.; Cresswell, P. Early phagosomes in dendritic cells form a cellular compartment sufficient for cross presentation of exogenous antigens. Proc. Natl. Acad. Sci. USA 2003, 100, 12889–12894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foged, C.; Brodin, B.; Frokjaer, S.; Sundblad, A. Particle size and surface charge affect particle uptake by human dendritic cells in an in vitro model. Int. J. Pharm. 2005, 298, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Limbach, L.K.; Li, Y.; Grass, R.N.; Brunner, T.J.; Hintermann, M.A.; Muller, M.; Gunther, D.; Stark, W.J. Oxide nanoparticle uptake in human lung fibroblasts: Effects of particle size, agglomeration, and diffusion at low concentrations. Environ. Sci. Technol. 2005, 39, 9370–9376. [Google Scholar] [CrossRef]
- Rehm, B.H.A. Polyester synthases: Natural catalysts for plastics. Biochem. J. 2003, 376, 15–33. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Yang, F.; Zou, J.; Wu, W.; Jing, H.; Gou, Q.; Li, H.; Gu, J.; Zou, Q.; Zhang, J. Immunization with Pseudomonas aeruginosa outer membrane vesicles stimulates protective immunity in mice. Vaccine 2018, 36, 1047–1054. [Google Scholar] [CrossRef]
- Mizuno, T.; Kageyama, M. Separation and Characterization of the Outer Membrane of Pseudomonas aeruginosa. J. Biochem. 1978, 84, 179–191. [Google Scholar] [CrossRef]
- Koh, A.Y.; Priebe, G.P.; Ray, C.; Van Rooijen, N.; Pier, G.B. Inescapable Need for Neutrophils as Mediators of Cellular Innate Immunity to Acute Pseudomonas aeruginosa Pneumonia. Infect. Immun. 2009, 77, 5300–5310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Giandomenico, A.; Rao, J.; Goldberg, J.B. Oral Vaccination of BALB/c Mice with Salmonella enterica Serovar Typhimurium Expressing Pseudomonas aeruginosa O Antigen Promotes Increased Survival in an Acute Fatal Pneumonia Model. Infect. Immun. 2004, 72, 7012–7021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pier, G.B.; Boyer, D.; Preston, M.; Coleman, F.T.; Llosa, N.; Mueschenborn-Koglin, S.; Theilacker, C.; Goldenberg, H.; Uchin, J.; Priebe, G.; et al. Human Monoclonal Antibodies to Pseudomonas aeruginosaAlginate That Protect against Infection by Both Mucoid and Nonmucoid Strains. J. Immunol. 2004, 173, 5671–5678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehm, B.H.A. Bacterial polymers: Biosynthesis, modifications and applications. Nat. Rev. Microbiol. 2010, 8, 578–592. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Sandford, S.; Kirman, J.R.; Rehm, B.H.A. Innovative antigen carrier system for the development of tuberculosis vaccines. FASEB J. 2019, 33, 7505–7518. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Reyes, P.; Parlane, N.A.; Buddle, B.M.; Wedlock, D.N.; Rehm, B.H.A. Immunological properties and protective efficacy of a single mycobacterial antigen displayed on polyhydroxybutyrate beads. Microb. Biotechnol. 2017, 10, 1434–1440. [Google Scholar] [CrossRef] [Green Version]
- Parlane, N.A.; Wedlock, D.N.; Buddle, B.M.; Rehm, B.H.A. Bacterial Polyester Inclusions Engineered to Display Vaccine Candidate Antigens for Use as a Novel Class of Safe and Efficient Vaccine Delivery Agents. Appl. Environ. Microbiol. 2009, 75, 7739–7744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinman, L. A brief history of TH17, the first major revision in the TH1/TH2 hypothesis of T cell–mediated tissue damage. Nat. Med. 2007, 13, 139–145. [Google Scholar] [CrossRef]
- Kolls, J.K.; Kanaly, S.T.; Ramsay, A.J. Interleukin-17: An emerging role in lung inflammation. Am. J. Respir. Cell Mol. Biol. 2003, 28, 9–11. [Google Scholar] [CrossRef]
- Chen, K.; McAleer, J.P.; Lin, Y.; Paterson, D.; Zheng, M.; Alcorn, J.F.; Weaver, C.; Kolls, J.K. Th17 Cells Mediate Clade-Specific, Serotype-Independent Mucosal Immunity. Immunity 2011, 35, 997–1009. [Google Scholar] [CrossRef] [Green Version]
- Komiyama, Y.; Nakae, S.; Matsuki, T.; Nambu, A.; Ishigame, H.; Kakuta, S.; Sudo, K.; Iwakura, Y. IL-17 Plays an Important Role in the Development of Experimental Autoimmune Encephalomyelitis. J. Immunol. 2006, 177, 566–573. [Google Scholar] [CrossRef] [Green Version]
- Pier, G.B.; Saunders, J.M.; Ames, P.; Edwards, M.S.; Auerbach, H.; Goldfarb, J.; Speert, D.P.; Hurwitch, S. Opsonophagocytic Killing Antibody toPseudomonas aeruginosaMucoid Exopolysaccharide in Older Noncolonized Patients with Cystic Fibrosis. N. Engl. J. Med. 1987, 317, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Knirel, Y.A. Polysaccharide Antigens of Pseudomonas aeruginosa. Crit. Rev. Microbiol. 1990, 17, 273–304. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.W.; Devlin, H.B.; Gnabasik, F.J. New immunotype schema for Pseudomonas aeruginosa based on protective antigens. J. Bacteriol. 1969, 98, 835–836. [Google Scholar] [CrossRef] [Green Version]
- Moser, C.; Jensen, P.O.; Kobayashi, O.; Hougen, H.P.; Song, Z.; Rygaard, J.; Kharazmi, A.; Hby, N. Improved outcome of chronic Pseudomonas aeruginosa lung infection is associated with induction of a Th1-dominated cytokine response. Clin. Exp. Immunol. 2002, 127, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Priebe, G.P.; Walsh, R.L.; Cederroth, T.A.; Kamei, A.; Coutinho-Sledge, Y.S.; Goldberg, J.B.; Pier, G. IL-17 Is a Critical Component of Vaccine-Induced Protection against Lung Infection by Lipopolysaccharide-Heterologous Strains ofPseudomonas aeruginosa. J. Immunol. 2008, 181, 4965–4975. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Gu, J.; Yang, L.; Gao, C.; Jing, H.; Wang, Y.; Zeng, H.; Zou, Q.; Lv, F.; Zhang, J. Protective Efficacy of the Trivalent Pseudomonas aeruginosa Vaccine Candidate PcrV-OprI-Hcp1 in Murine Pneumonia and Burn Models. Sci. Rep. 2017, 7, 3957. [Google Scholar] [CrossRef] [Green Version]
- Sainz-Mejías, M.; Jurado-Martín, I.; McClean, S. Understanding Pseudomonas aeruginosa–Host Interactions: The Ongoing Quest for an Efficacious Vaccine. Cells 2020, 9, 2617. [Google Scholar] [CrossRef]
- Wan, C.; Gao, C.; Xie, Q.; Wang, Y.; Cheng, X.; Fang, Y.; Liu, Z.; Zhang, W.; Zou, Q.; Lu, G.; et al. Flagella hook protein FlgE is a novel vaccine candidate of Pseudomonas aeruginosa identified by a genomic approach. Vaccine 2021, 22, 2386–2395. [Google Scholar] [CrossRef]
- Bayes, H.K.; Bicknell, S.; MacGregor, G.; Evans, T.J. T Helper Cell Subsets Specific for Pseudomonas aeruginosa in Healthy Individuals and Patients with Cystic Fibrosis. PLoS ONE 2014, 9, e90263. [Google Scholar] [CrossRef] [PubMed]
- Roux, D.; Gaudry, S.; Dreyfuss, D.; El-Benna, J.; de Prost, N.; Denamur, E.; Saumon, G.; Ricard, J.D. Candida albicans impairs macrophage function and facilitates Pseudomonas aeruginosa pneumonia in rat. Crit. Care Med. 2009, 37, 1062–1067. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzaga, Z.J.C.; Merakou, C.; DiGiandomenico, A.; Priebe, G.P.; Rehm, B.H.A. A Pseudomonas aeruginosa-Derived Particulate Vaccine Protects against P. aeruginosa Infection. Vaccines 2021, 9, 803. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9070803
Gonzaga ZJC, Merakou C, DiGiandomenico A, Priebe GP, Rehm BHA. A Pseudomonas aeruginosa-Derived Particulate Vaccine Protects against P. aeruginosa Infection. Vaccines. 2021; 9(7):803. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9070803
Chicago/Turabian StyleGonzaga, Zennia Jean C., Christina Merakou, Antonio DiGiandomenico, Gregory P. Priebe, and Bernd H. A. Rehm. 2021. "A Pseudomonas aeruginosa-Derived Particulate Vaccine Protects against P. aeruginosa Infection" Vaccines 9, no. 7: 803. https://0-doi-org.brum.beds.ac.uk/10.3390/vaccines9070803