1. Introduction
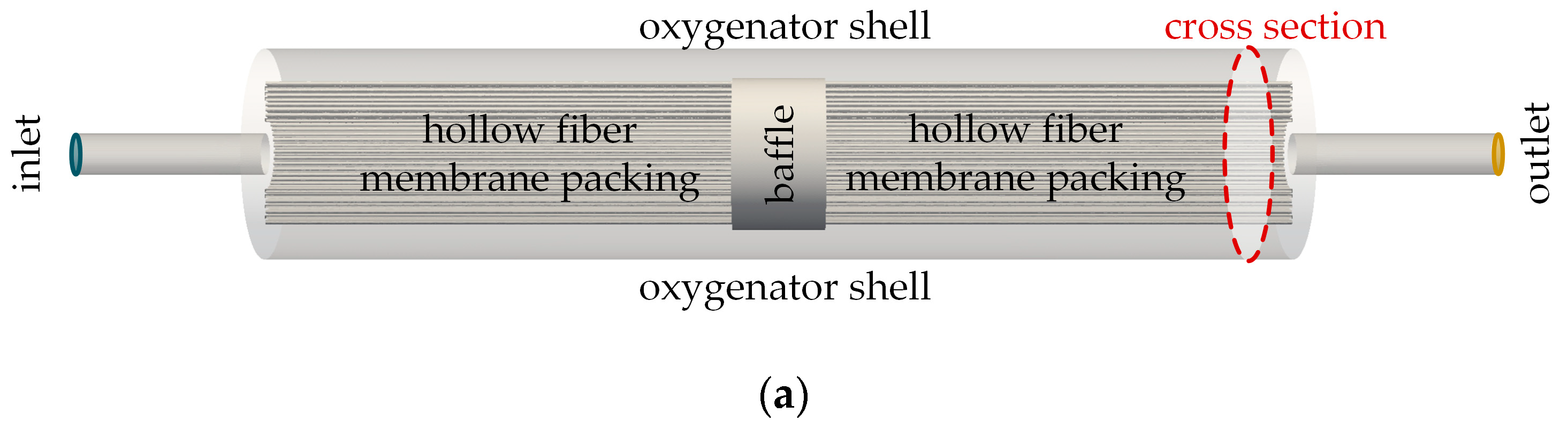
Blood oxygenators, also known as artificial lungs, are needed to supplement respiratory function during cardiopulmonary bypass or to support patients with respiratory failure. A prerequisite for sufficient gas exchange is a large gas-exchange surface and efficient contact between the blood and membrane. To provide a large gas-exchange surface at the lowest possible priming volume, hollow fiber membrane packings are used [
1]. In modern oxygenators, blood flows on the shell side of the packing while the fiber lumen are purged with O
2. The transmembrane transfer of the respiratory gases (CO
2 and O
2) is hereby facilitated by the partial pressure difference between the blood and gas phase [
2].
Although great efforts have been made to improve the biocompatibility of oxygenator circuits [
3], serious side effects occur due to contact of blood with the artificial polymer surfaces. These side effects ultimately include reduced platelet function and survival as well as prolonged bleeding times after perfusion [
4]. Consequently, the optimization of oxygenators aims for increasing gas exchange efficiency while reducing the membrane surface and blood priming volume [
5].
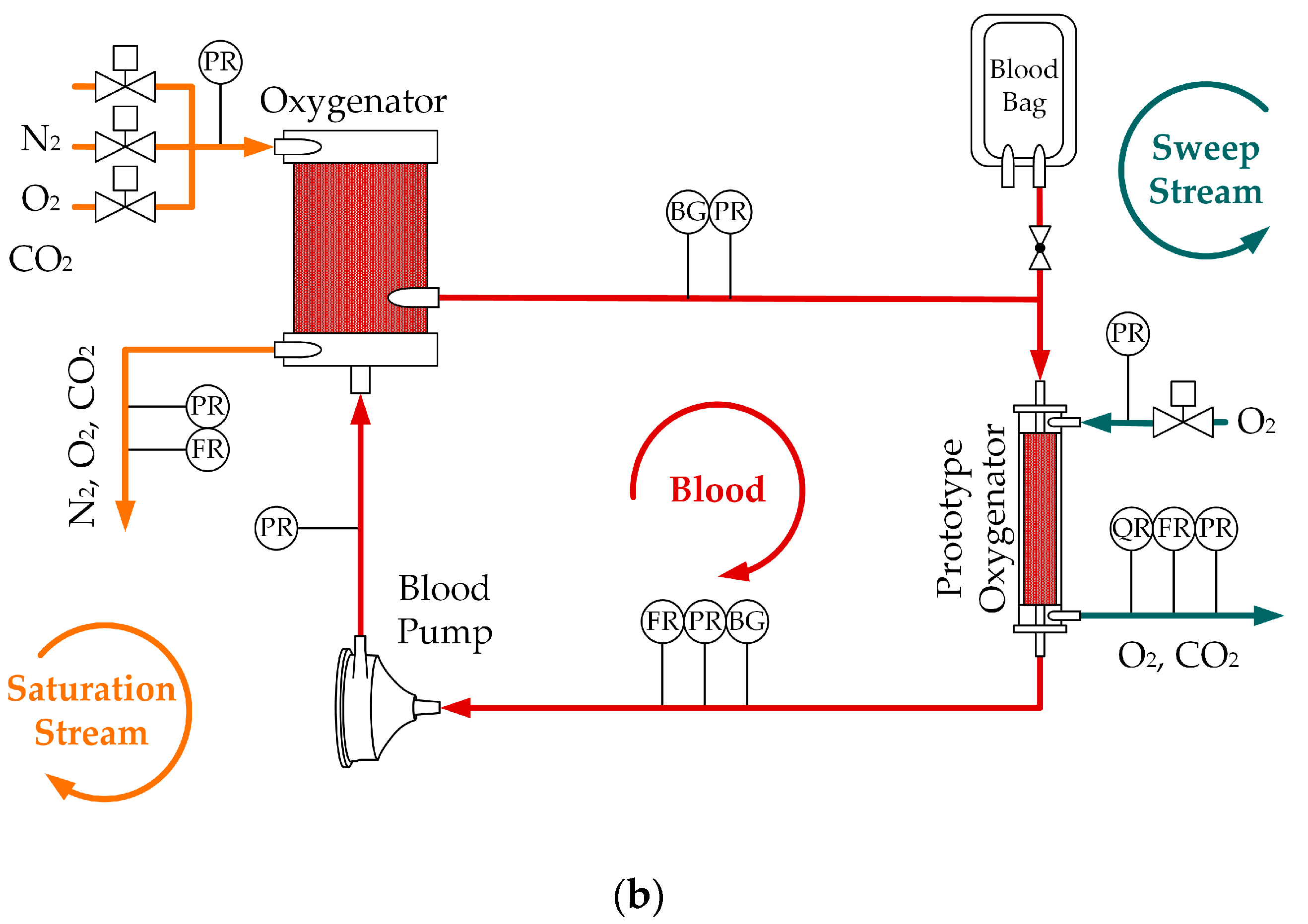
For reviewing and studying optimized oxygenator designs, experimental methods are the most reliable. However, the use of blood significantly increases the effort and costs in many ways:
The use of blood is accompanied by animal suffering.
The use of blood is not permitted in all technical laboratories.
Test circuits get contaminated and must be rebuilt due to blood deposits.
Additional logistical challenges due to limited durability of blood.
Ethics committee approval must be obtained.
Water is a cheap, save and easy-to-handle substitute. However, water tests must be able to reasonably represent the CO
2 removal performance observed with blood. Here, the difference in physical and chemical properties between blood and waster must be considered. While O
2 solubility of blood differs significantly from that of water due to the binding of O
2 to hemoglobin [
6], the CO
2 solubility in water and blood is subject to similar mechanisms. CO
2 first dissolves physically and then reacts to carbonic acid (H
2CO
3), which dissociates to bicarbonate (HCO
3−) [
7]:
In order to shift the reaction equilibrium toward the production of bicarbonate, blood buffers the pH decrease, which is caused by the dissociation reaction. Hence, the total CO
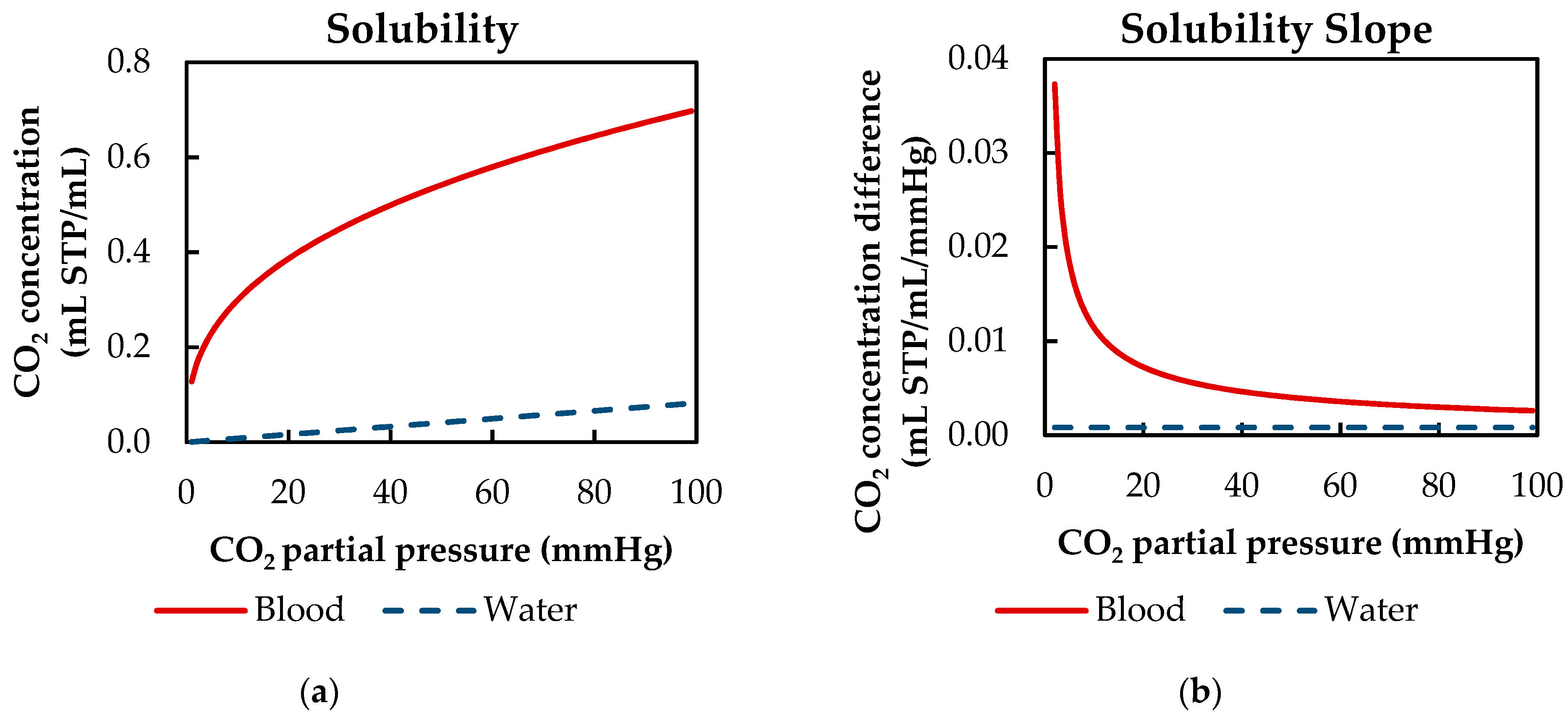
2 capacity of blood exceeds that of water. Examining the solubility curves of blood and water at clinically relevant CO
2 partial pressures (40 to 100 mmHg) shows that the CO
2 capacity of blood is up to 11 times higher (
Figure 1a). Additionally, the reaction from CO
2 to carbonic acid is accelerated by the catalytic enzyme carbonic anhydrase located in the red blood cells [
7].
However, the effect of the buffer system and the enzymatic catalyst on the overall CO
2 removal of oxygenators is limited because of two reasons. First, the slope of the solubility curve of blood and water at relevant venous levels is comparable. This is of importance, as the CO
2 removal is primarily dependent on the concentration difference. At CO
2 partial pressures from 40 to 100 mmHg, the slope of the solubility curve of blood is only four times higher than the slope of water (
Figure 1b). Second, outside the red blood cells and in the absence of the catalyst carbonic anhydrase, the reaction from physically dissolved CO
2 to carbonic acid and vice versa is slow, compared to the short residence time of blood in an oxygenator. Consequently, mostly physically dissolved CO
2 is removed at the membrane [
8].
Compared to water, the diffusion of CO
2 components in blood is hindered by the presence of proteins [
7]. This leads to an approximately 2.6 times higher diffusion rate for physically dissolved CO
2 in water than in blood plasma [
11].
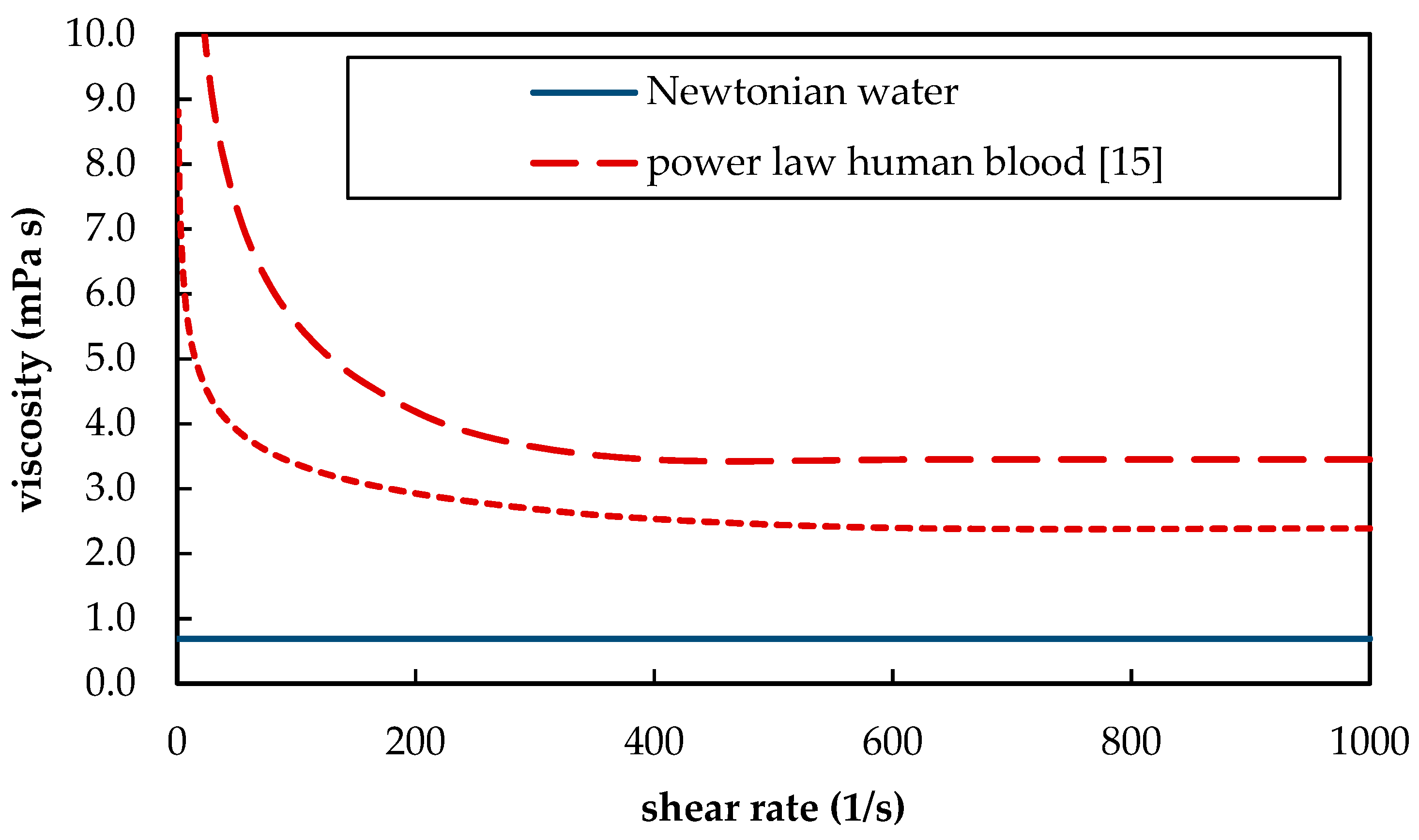
Furthermore, blood as a suspension of blood plasma and red blood cells shows shear thinning behavior. The shear thinning behavior hereby differs between different animal species [
12] and depends, among other parameters, on the hematocrit [
13]. At high shear rates, the whole blood viscosity converges toward its minimum (
Figure 2). For human blood, this minimum is approximately 3.5 mPa s. In contrast, water is a Newtonian fluid and has a viscosity of 0.69 mPa s at 37 °C [
14]. The different rheological behavior of blood and water (
Figure 2) has two opposing influences on CO
2 separation. On the one hand, lower viscosities result in more turbulent flow conditions. This could promote additional mixing. On the other hand, higher viscosities produce higher shear stress. Due to the higher shear stress, thinner boundary layers can be expected. Both additional mixing at lower viscosities (water) and thinner boundary layers at higher viscosities (blood) would result in an increase in CO
2 removal.
To conclude, blood and water differ in CO2 solubility, CO2 diffusion rate and viscosity. These important parameters have partially opposing effects on the CO2 removal of oxygenators. This makes it difficult to evaluate the suitability and limitations of water as a blood substitute without using in-depth studies.
In recent research, multiple in vitro studies were conducted, using water as a blood substitute to determine the CO
2 removal of an oxygenator. Hout et al. [
17] investigated the dependency of CO
2 removal on the sweep flow rate. This was done for two different oxygenator models. In order to be able to use water as a blood substitute, the gas exchange rate was normalized by the maximum CO
2 removal rate of the respective oxygenator. By doing so, the measured CO
2 removal rates can be made independent from the specific mass transfer characteristics of an oxygenator [
18], which, in turn, are dependent on the used test fluid (blood/water). Hattler et al. [
19] tested the CO
2 and O
2 transfer performance of a gas exchange catheter in vitro with water, and in vivo, using calves as a large animal model. However, the suitability of water is only discussed for the O
2 exchange. Additionally, the in vitro and in vivo results cannot be compared directly due to the different hydrodynamic conditions. Consequently, a representative deviation of the CO
2 removal rates determined with blood and water cannot be calculated. Svitek et al. [
11] proposed a Sherwood model, allowing to predict the CO
2 removal with blood, based on experiments with water, using an adapted diffusivity. To validate the model, in vitro tests with blood and water were conducted. Although the experimental results show comparable CO
2 removal rates for blood and water they are not compared directly by the authors. The suitability of water for prediction of the CO
2 removal of oxygenators is only discussed in the scope of the proposed conversion of water to blood data. Tabesh et al. [
20] determined the CO
2 removal rate of oxygenators via in vitro tests with porcine blood and water. To reduce the difference between the two CO
2 removal rates of blood and water, N
2 was blended into the saturation stream. This allowed to reduce the maximum deviation to 5%. Yet, the reason for the good fit between blood and water is not analyzed and discussed. Mihelc et al. [
21] and Jeffries et al. [
22] reported that the CO
2 removal measured for an intracorporal membrane catheter correlates well (within 10% deviation) when comparing in vitro trials with water to in vivo trials with calves. The good agreement is attributed to the opposing effects on the CO
2 removal rate induced by the different viscosities and CO
2 solubilities of blood and water. An in-depth examination of this phenomenon is not conducted. Furthermore, they explicitly limit the suitability of water to their devices and consider the good correlation between water and blood to be fortuitous. A recommendation to use water for the in vitro determination of the CO
2 removal rate is not given.
To summarize, water is commonly used as a blood substitute for the in vitro determination of the CO2 removal rate of oxygenators. Its suitability for determining the total CO2 removal rate has been confirmed by multiple independent research groups but has never been main focus of their published research. The CO2 solubility, CO2 diffusion rate and viscosity of water differ significantly from those of blood. Due to the complex interactions of these parameters, the reason for the suitability of water as a blood substitute for the determination of CO2 removal in oxygenators remains unclear.
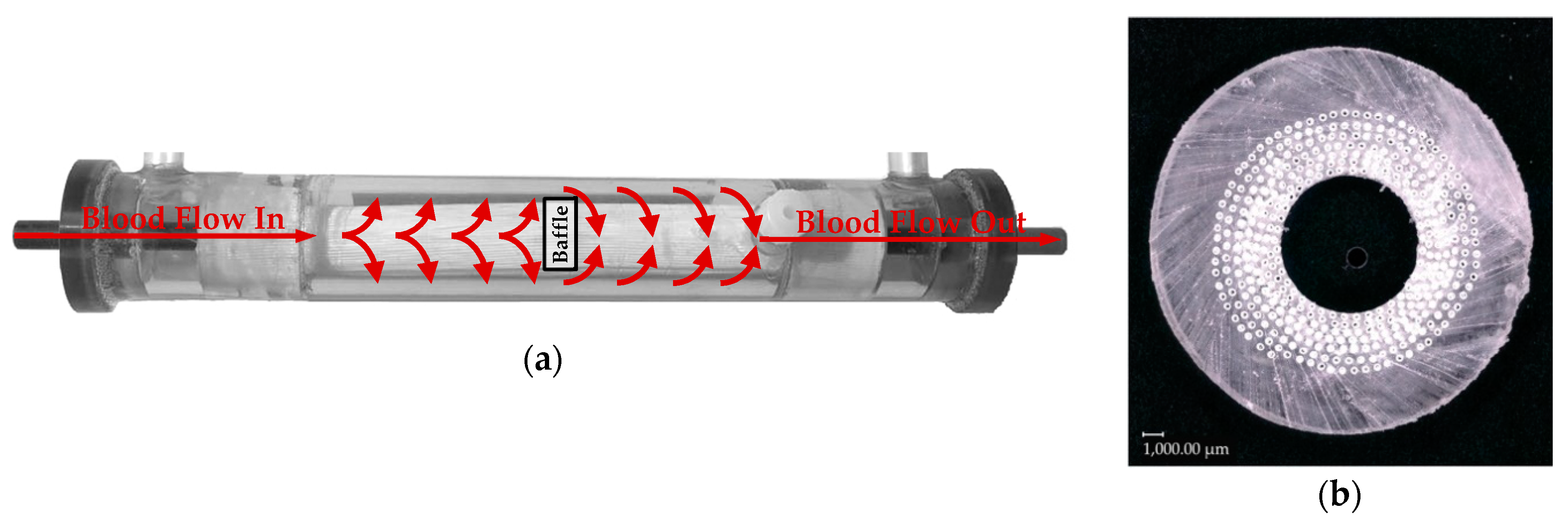
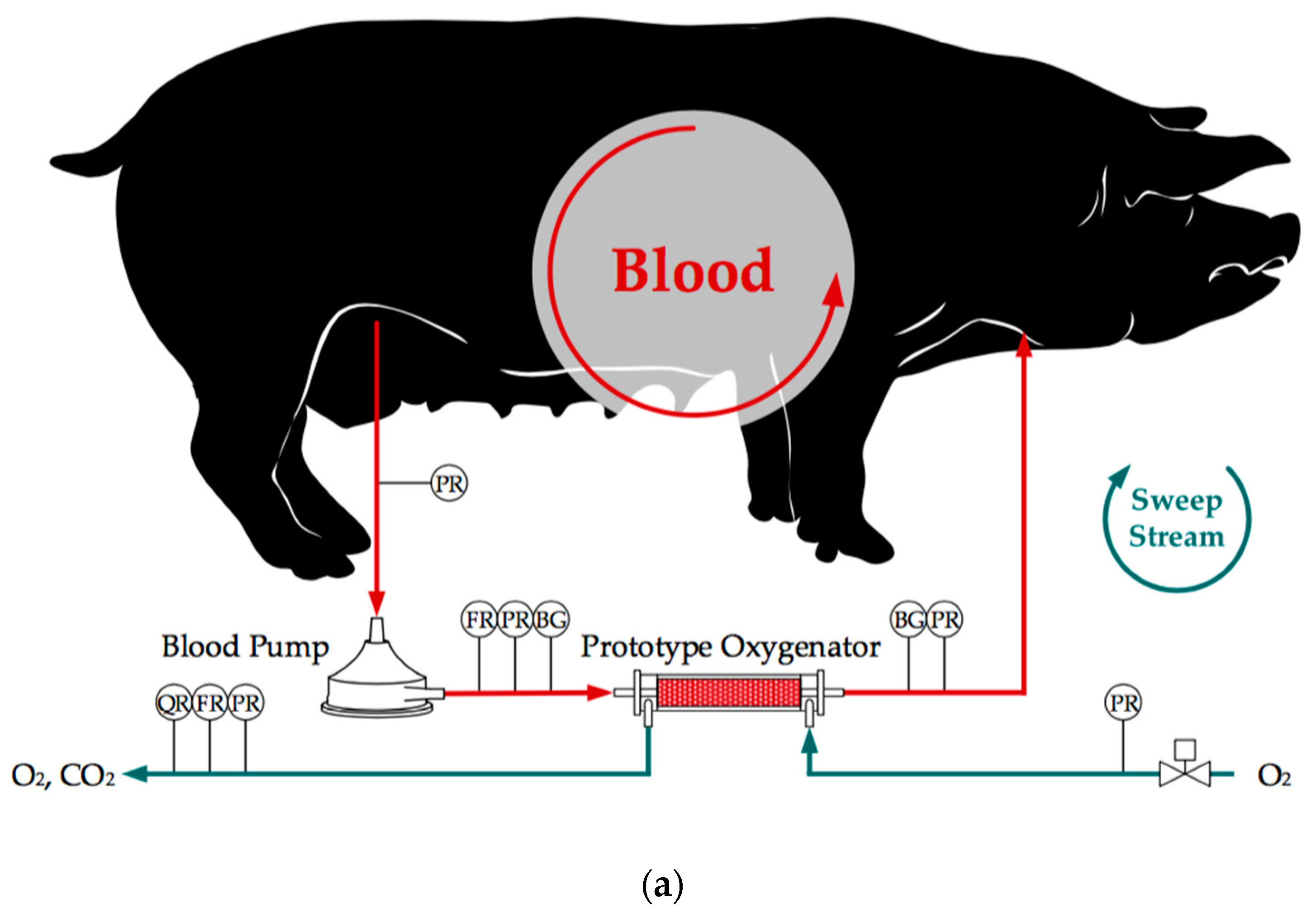
The scope of this research is to give a detailed comparison of CO
2 removal rates gained from in vitro tests using water with data from in vivo tests using pigs as large animal models. To better understand the contribution of the differing CO
2 solubility, CO
2 diffusion and viscosity on the CO
2 exchange, the CO
2 concentration polarization in the boundary layer attached to the membrane is studied. It represents the main CO
2 transport resistance and, therefore, characterizes the CO
2-removal performance of oxygenators [
2]. As the boundary layer cannot be resolved experimentally, computational fluid dynamic (CFD) simulations were conducted to investigate and compare the behavior of the boundary layer for blood and water. By comprising experimental and CFD results, this work aims to evaluate the suitability and limitations of water as a blood substitute for the determination of the CO
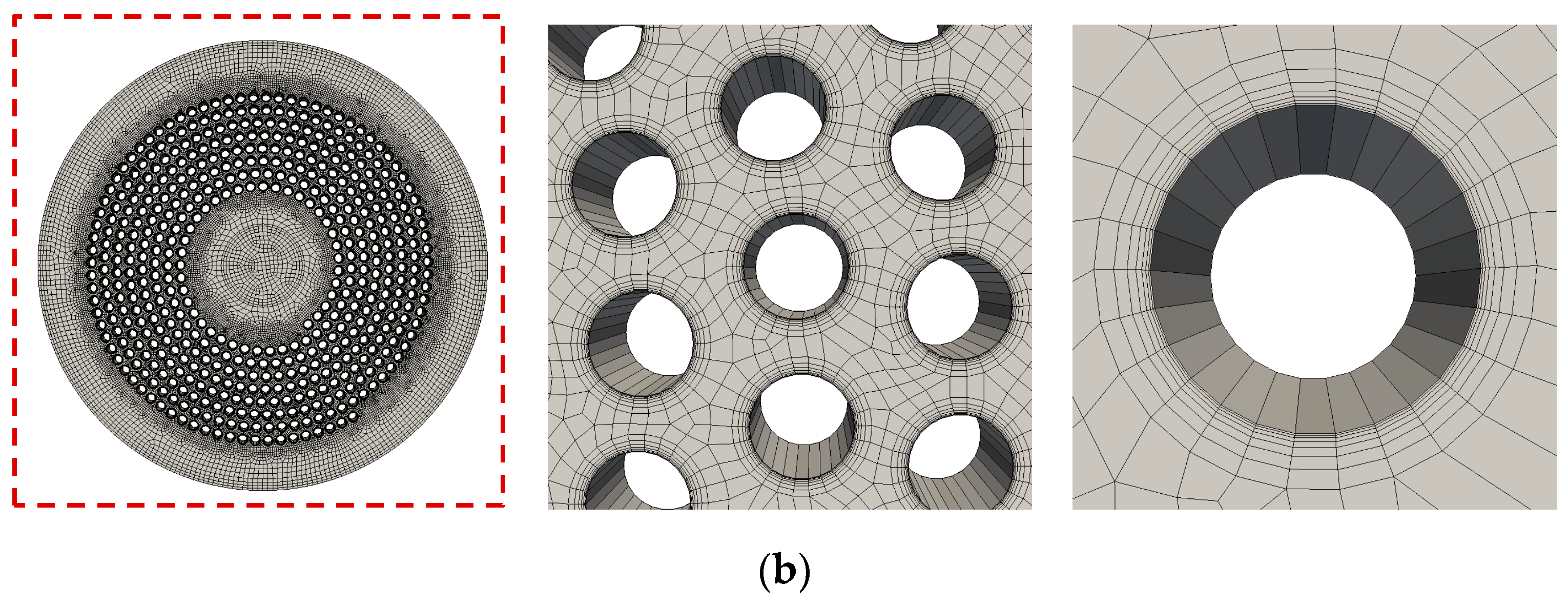
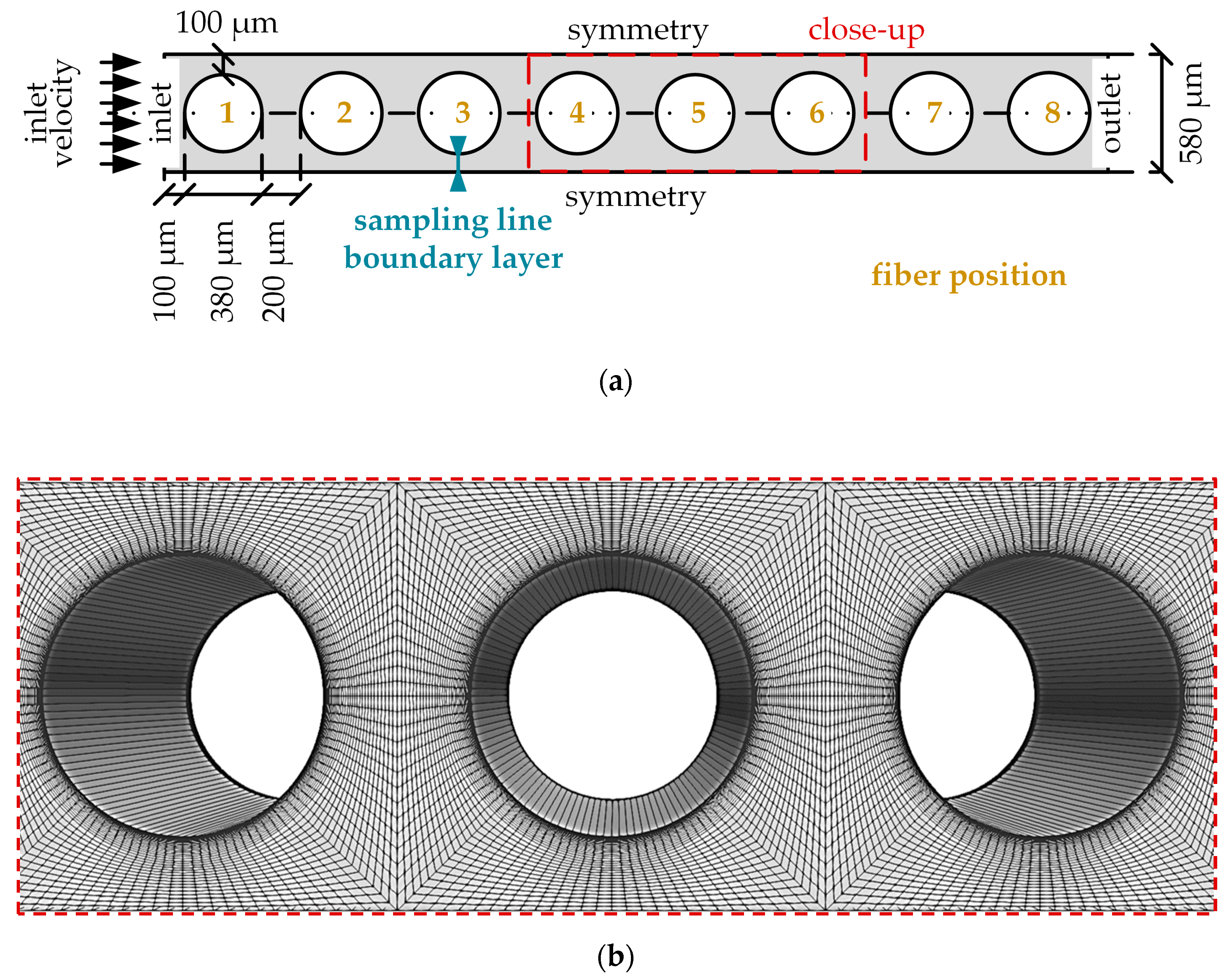
2 removal rate of oxygenators.
4. Conclusions
The CO
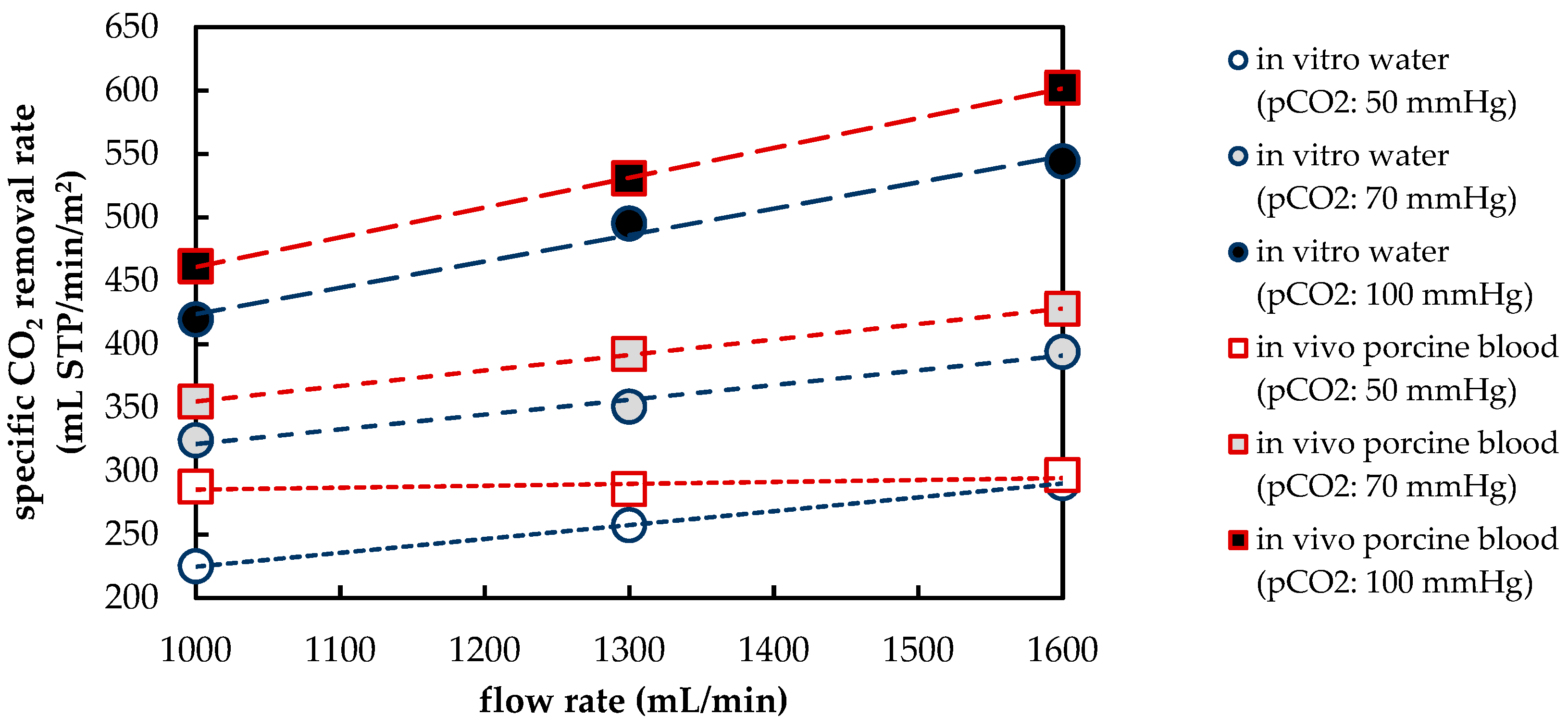
2 removal rate of a prototype oxygenator was measured in vivo using porcine blood and in vitro using water. In general, the CO
2 removal rates of porcine blood and water are comparable despite differing fluid properties (CO
2 diffusion, CO
2 solubility and viscosity). The deviation of the CO
2 removal rate determined with porcine blood from that determined with water amounts to approximately 10%. This deviation agrees well with the data found in recent literature [
20,
21,
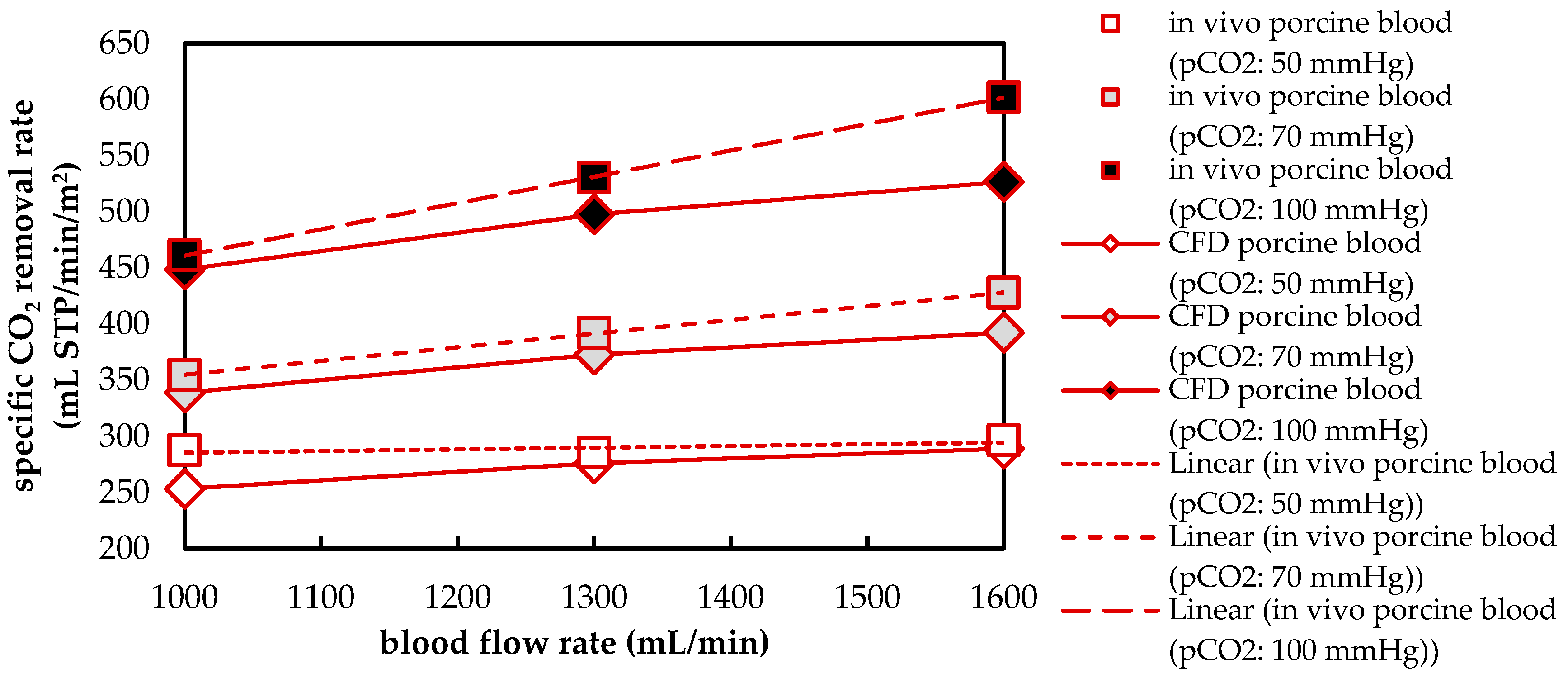
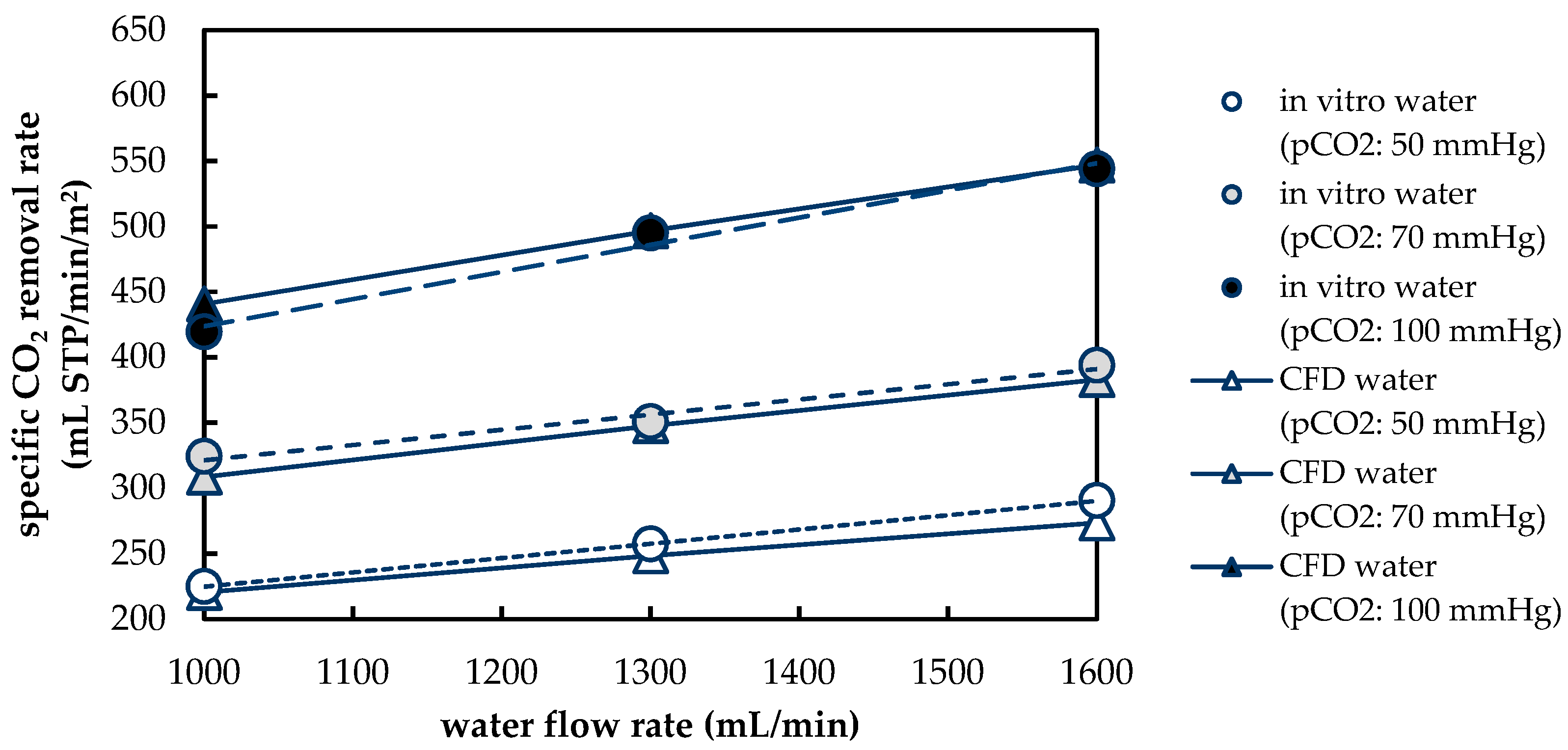
22]. Based on the results of the in vivo and in vitro tests, the CFD CO
2 transport simulations were validated.
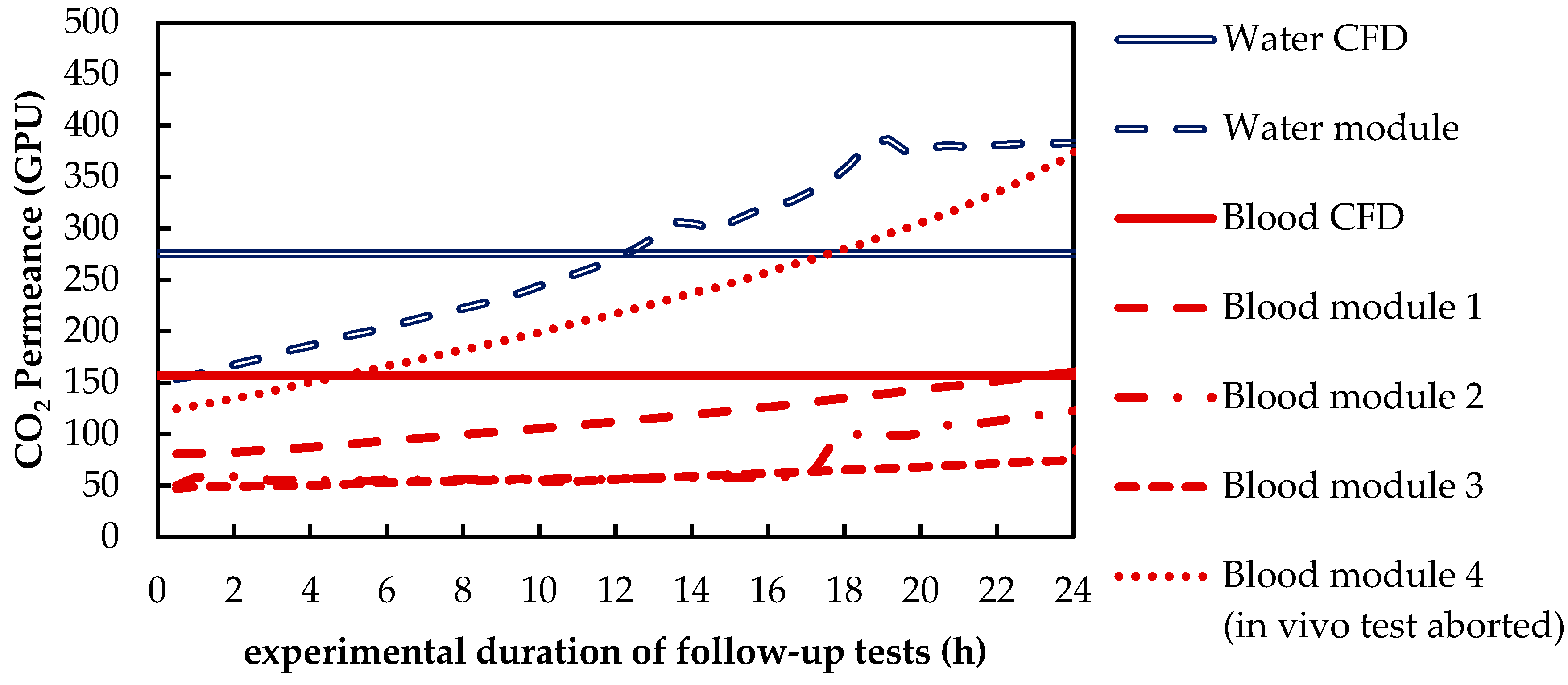
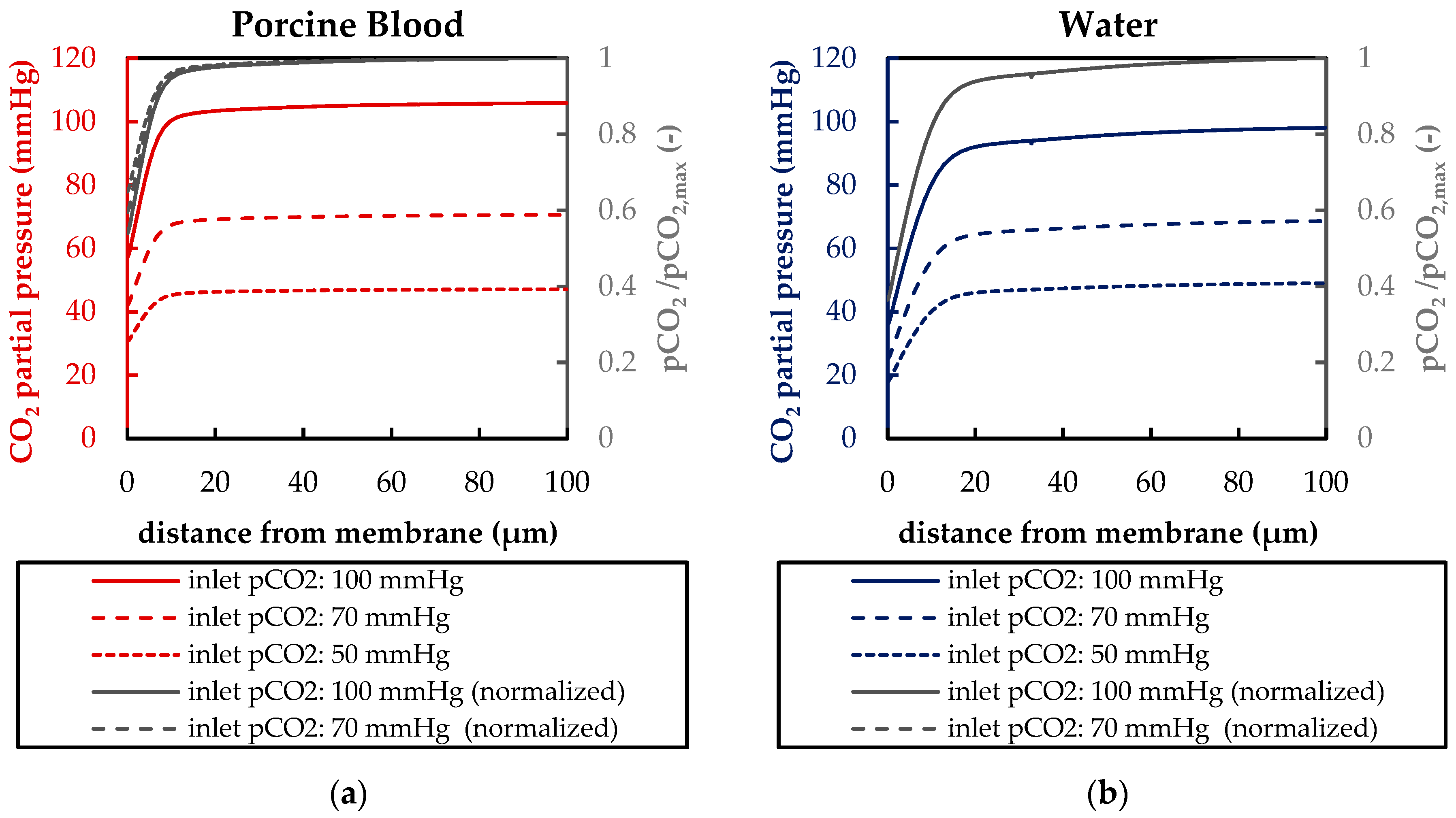
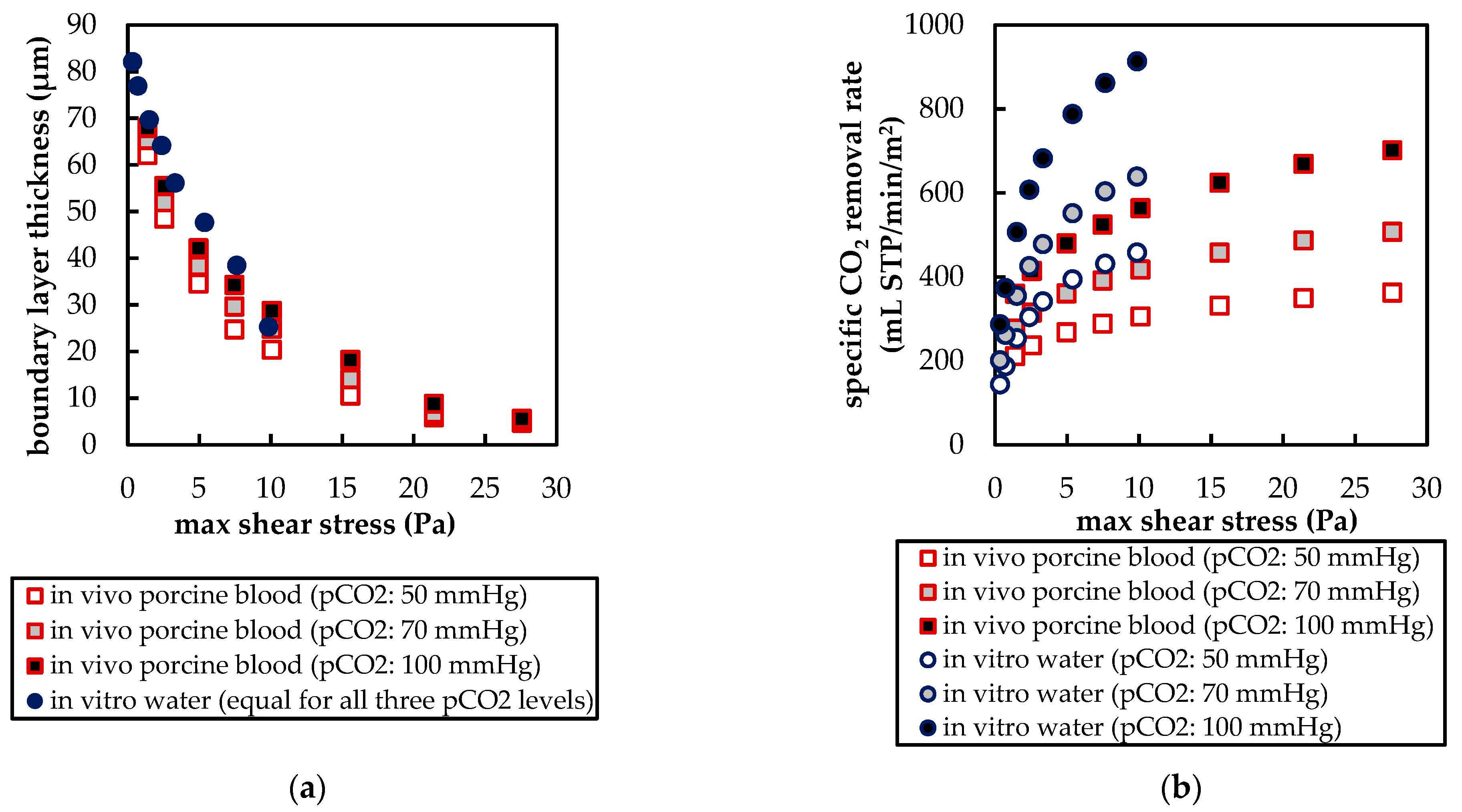
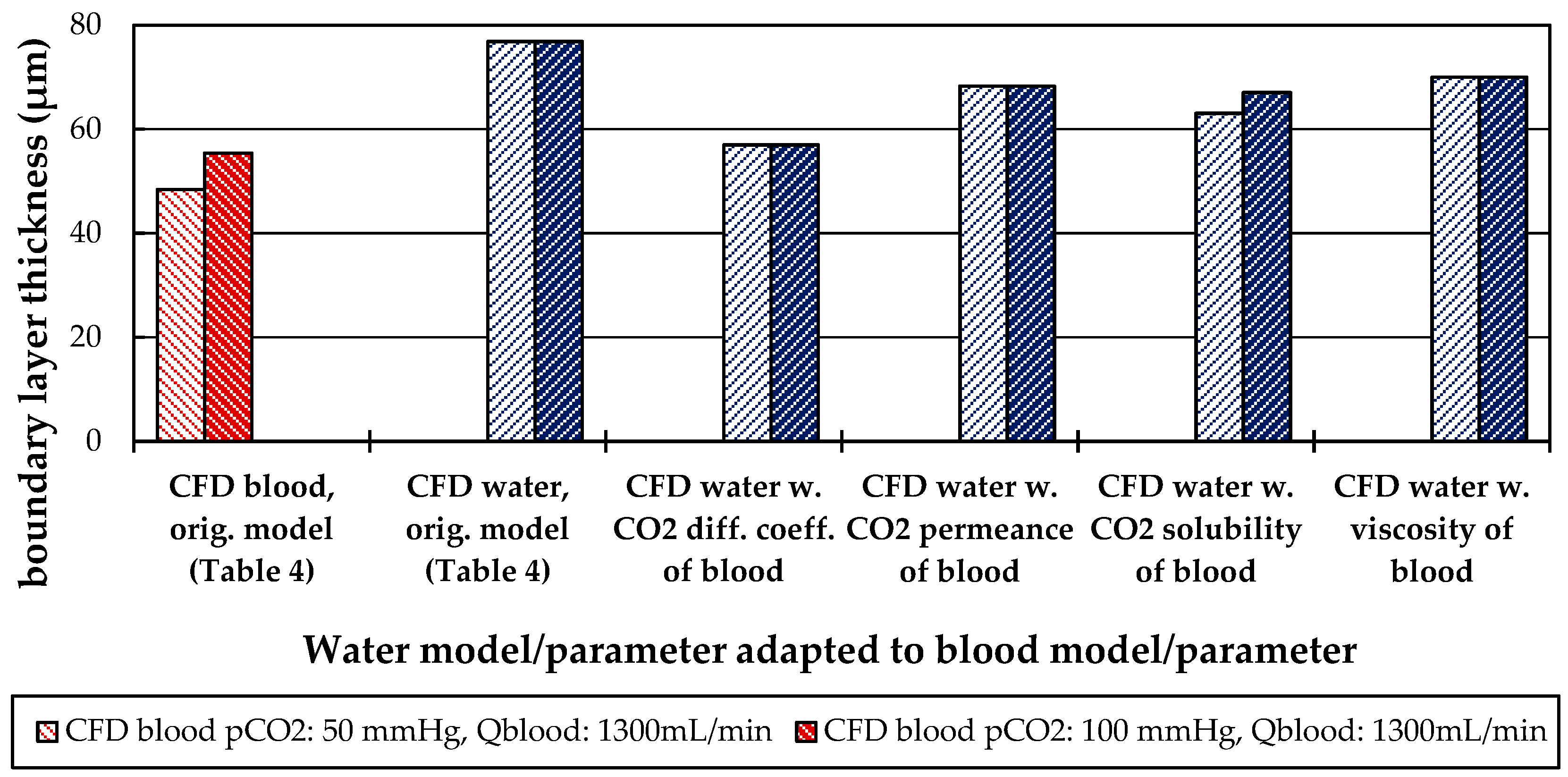
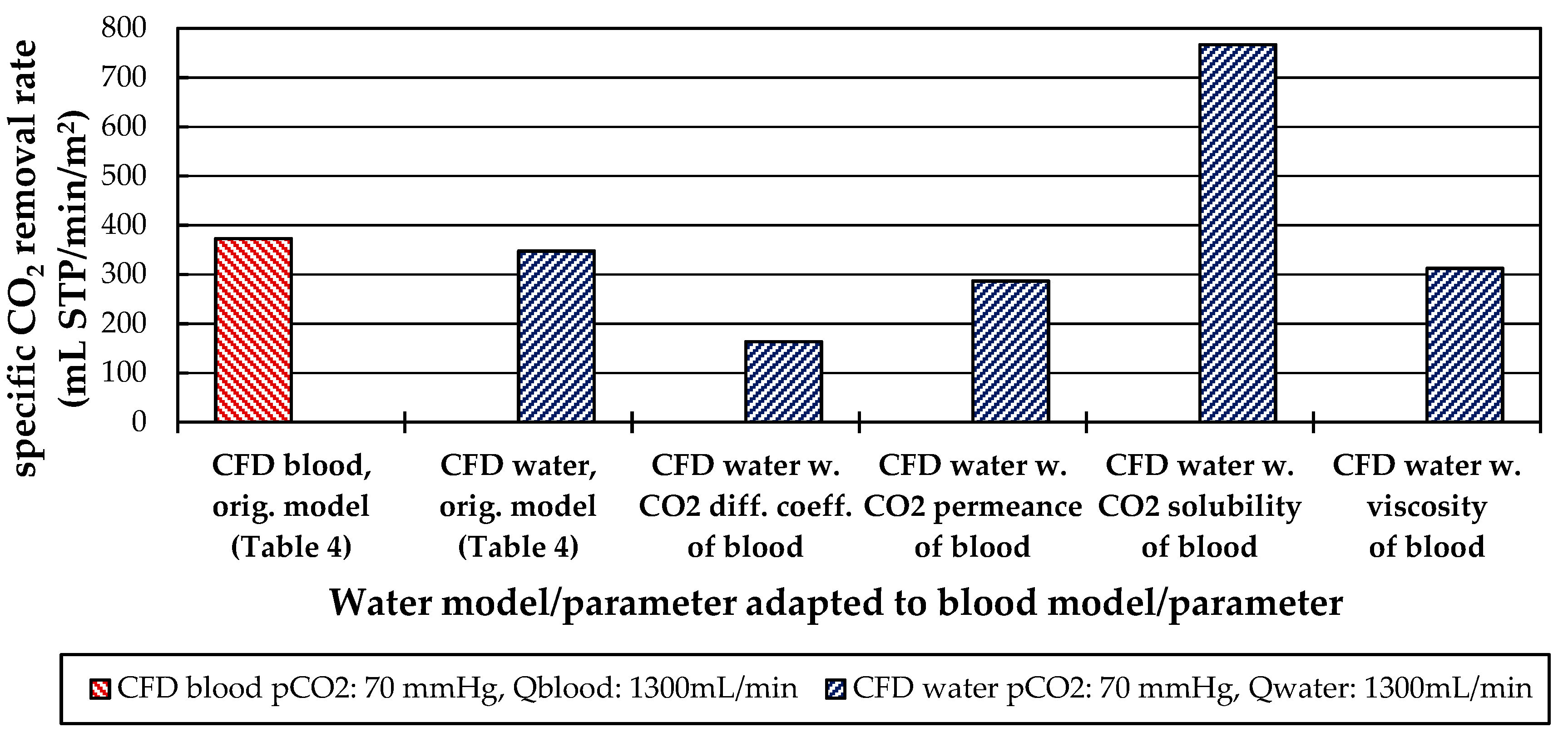
Besides differences in the fluid properties of blood and water, our experimental and simulation data indicate that additionally higher CO2 permeances are available during tests with water, probably due to the absence of blood residues on the membrane surface. The influences of the differing fluid properties (CO2 diffusion, CO2 solubility and viscosity) as well as membrane material properties (CO2 permeance) on CO2 removal were quantified, utilizing the developed CFD models. The difference in CO2 solubility between blood and water has the strongest effect on the CO2 removal rate, followed by the difference in the CO2 diffusion rate, difference in CO2 permeance and difference in viscosity.
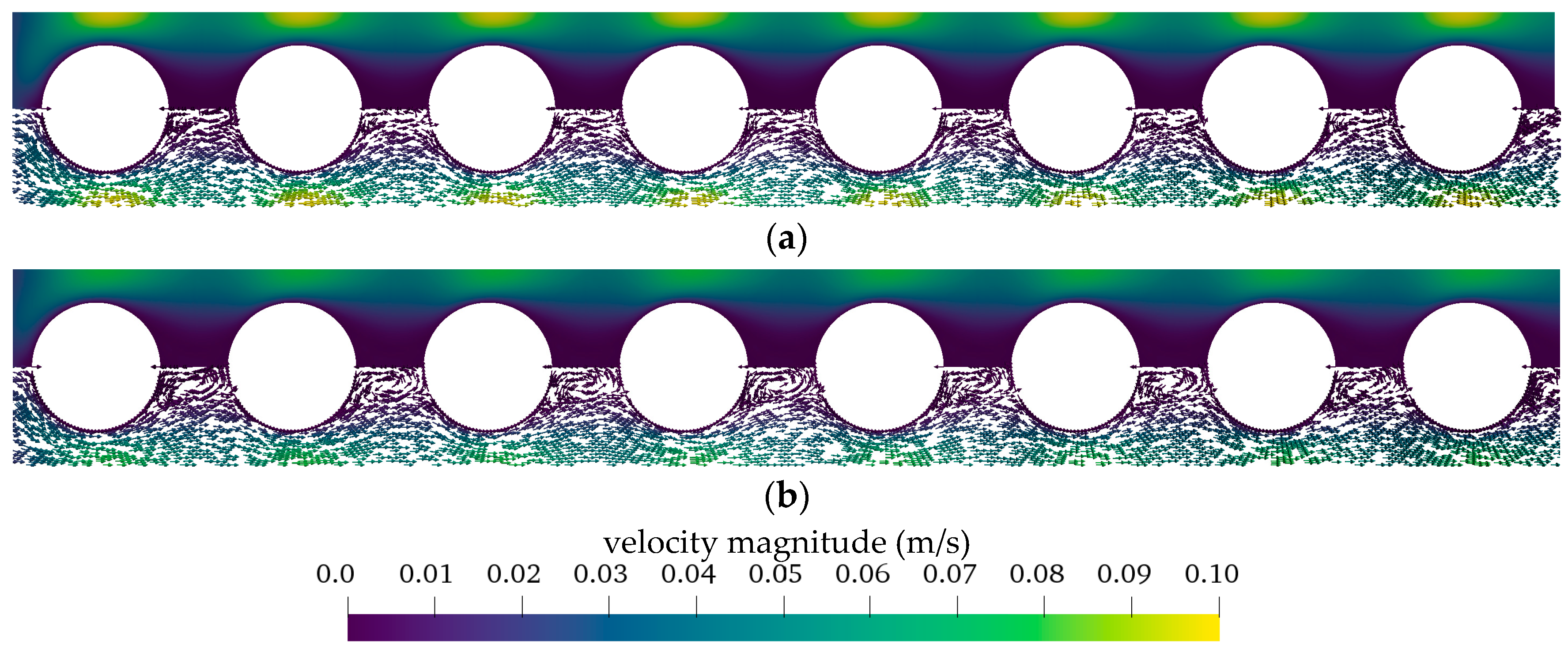
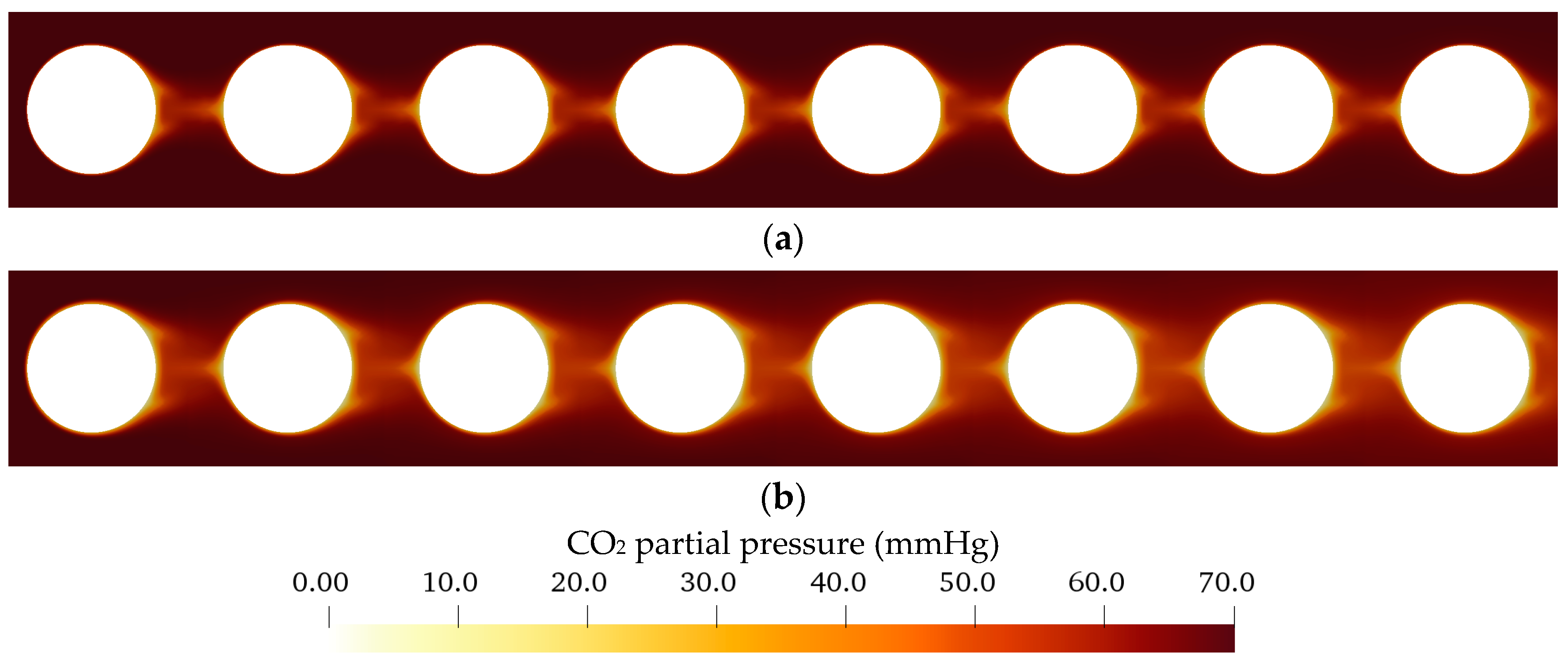
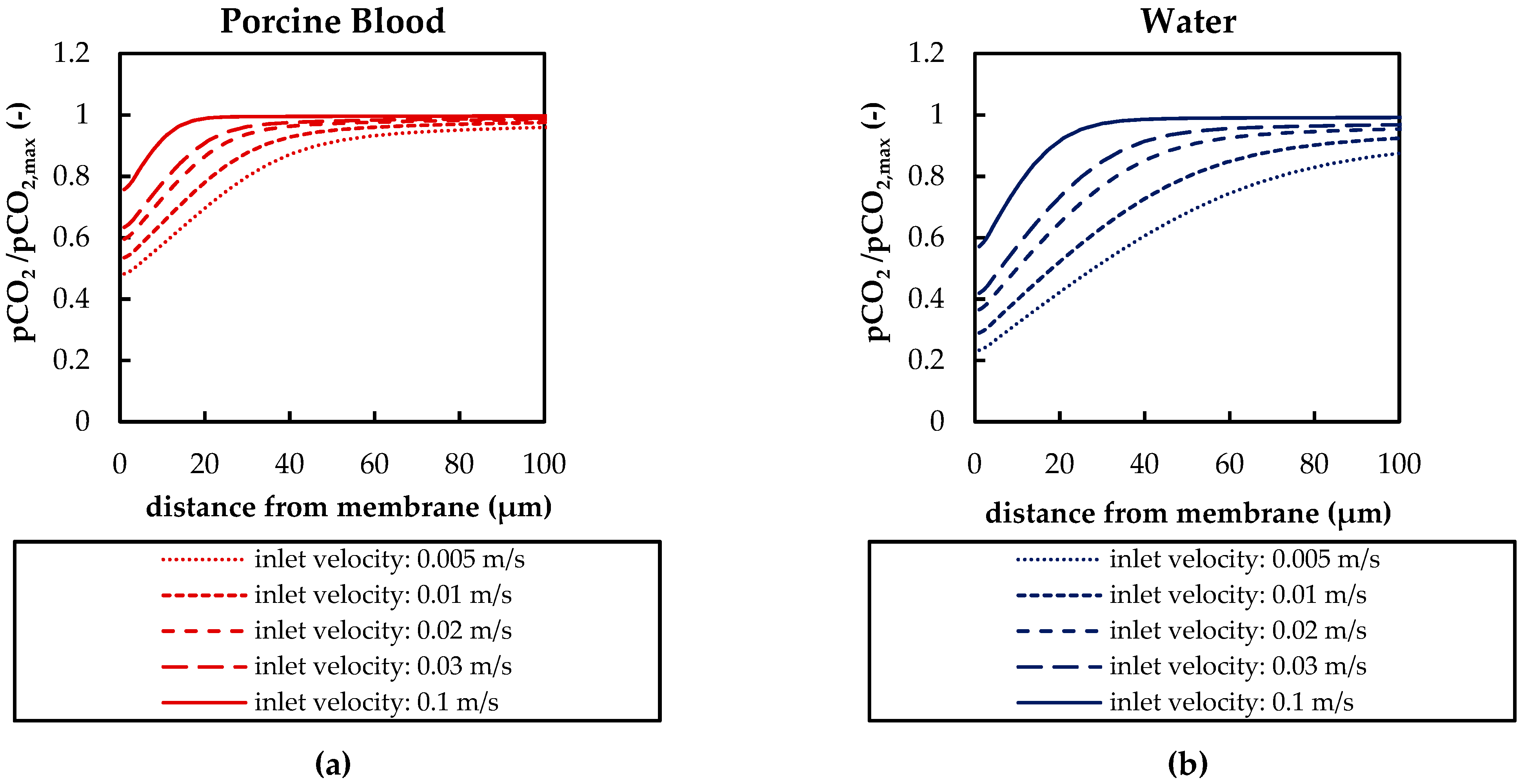
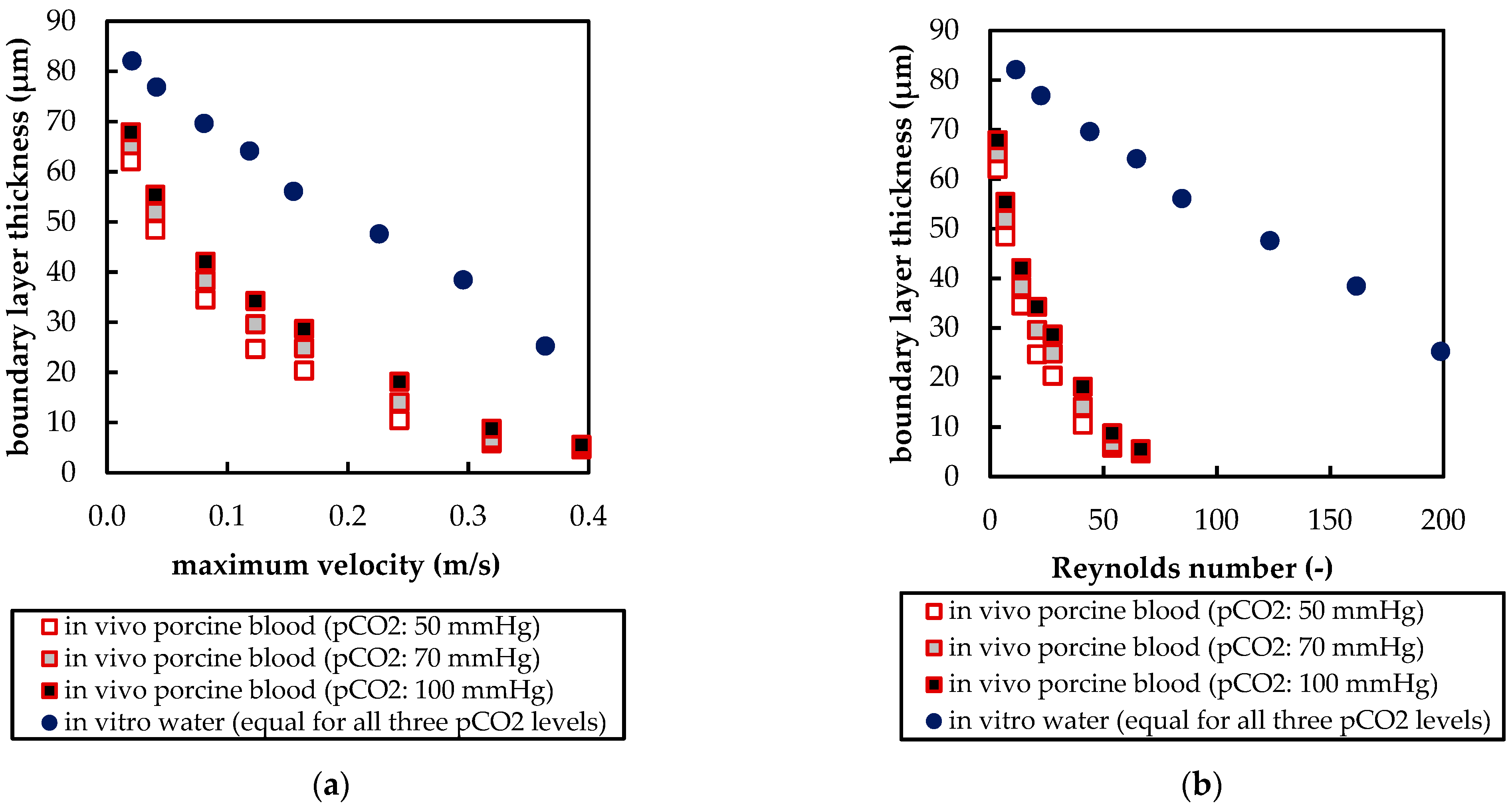
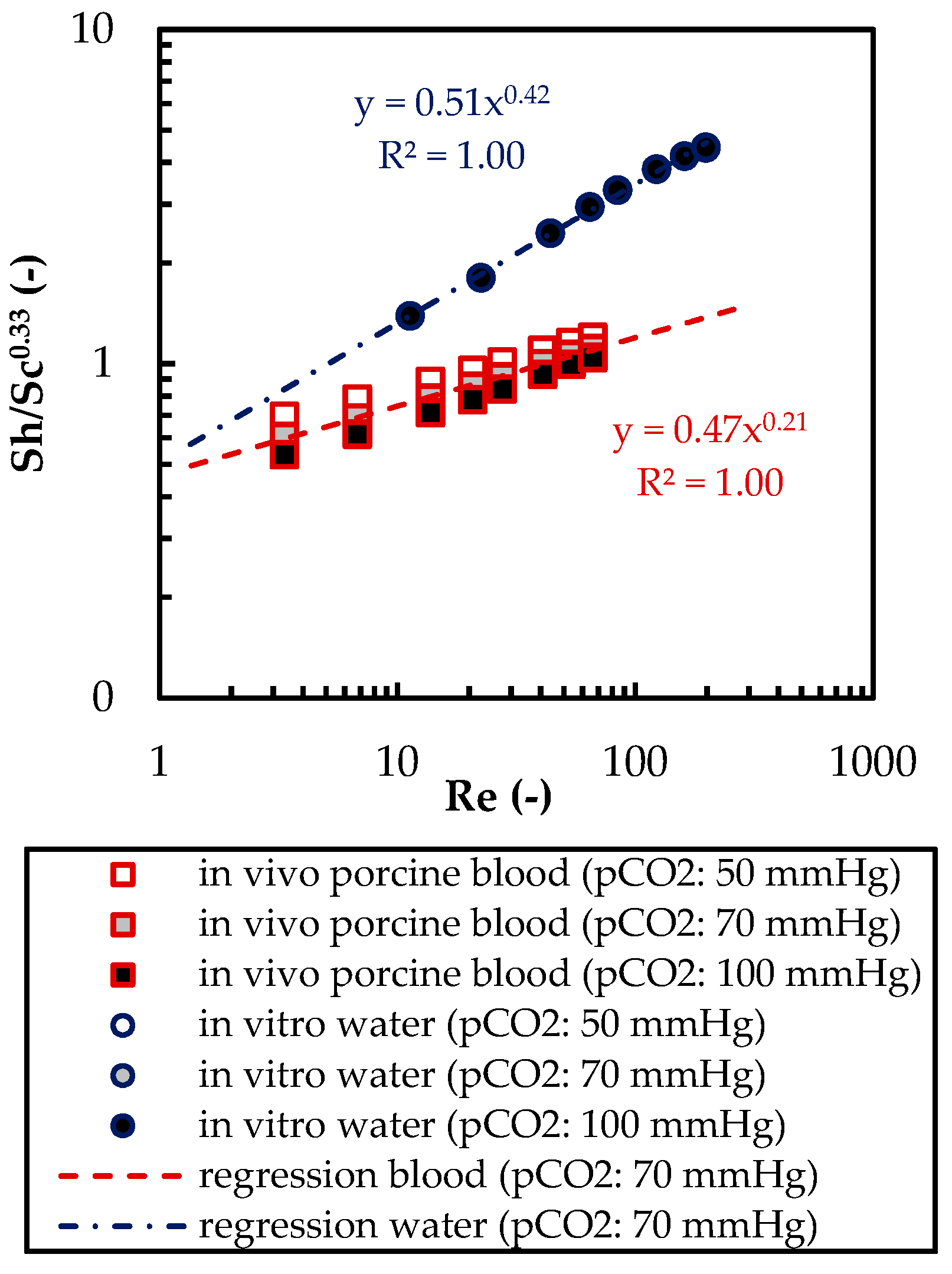
The CFD simulations also allow to resolve and study the main CO2 transport resistance—the diffusional boundary layer attached to the membrane surface. The simulations show that the pCO2 boundary layer in water is, in general, thicker than that in blood. The CFD results indicate that the thicker boundary layer in water can be mainly attributed to the higher diffusion coefficient of CO2 in water (2.38 × 10−9 m2/s) than in blood (6.96 × 10−10 m2/s). Furthermore, the CFD model suggests that the pCO2 boundary layer thickness in blood is dependent on bulk pCO2. This is probably due to the binding of CO2 in multiple components (physically dissolved, bicarbonate and carbaminohemoglobin) and the resulting nonlinear dependency of the CO2 partial pressure from the CO2 concentration (CO2 solubility). Additionally, CFD simulations indicate that the pCO2 boundary layer thicknesses of porcine blood and water are in good agreement when compared at same shear stresses. However, the CO2 removal rates of porcine blood and water deviate stronger at the same shear stresses (>20%) than at the same blood/water flow rates (approx. 10%). Consequently, the boundary layer thickness and CO2 removal rate cannot be matched simultaneously. Differences in the CO2 mass transfer characteristics of the boundary layer can be confirmed when comparing Sherwood correlations established for porcine blood and water.
To conclude, the boundary layers of blood and water behave, in general, differently. Studies of blood oxygenators aiming to investigate the boundary layer or measures for boundary layer reduction should, therefore, rely on blood tests. However, the determination of the total CO2 removal rate of an oxygenator using water as a blood substitute should be possible with reasonable accuracy. Deviation of the CO2 removal rate determined with water to the CO2 removal rate determined with blood should lie within 10%. This level of accuracy could potentially differ for flow conditions other than those studied here (crossflow, Re within the packing approx. 20). Nevertheless, the results of this work provide a basis for future optimization of the CO2 removal performance of oxygenators, using in vitro tests with water.


 ,
, 
























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}