Phytocannabinoids—A Green Approach toward Non-Alcoholic Fatty Liver Disease Treatment

 , , and
, , and
Abstract
:1. Introduction
2. Endocannabinoid System (ECS)
3. Cannabinoid System in Non-Alcoholic Fatty Liver Disease (NAFLD) Development and Progression—The Role of CB1R and CB2R and PPARs
4. Phytocannabinoids
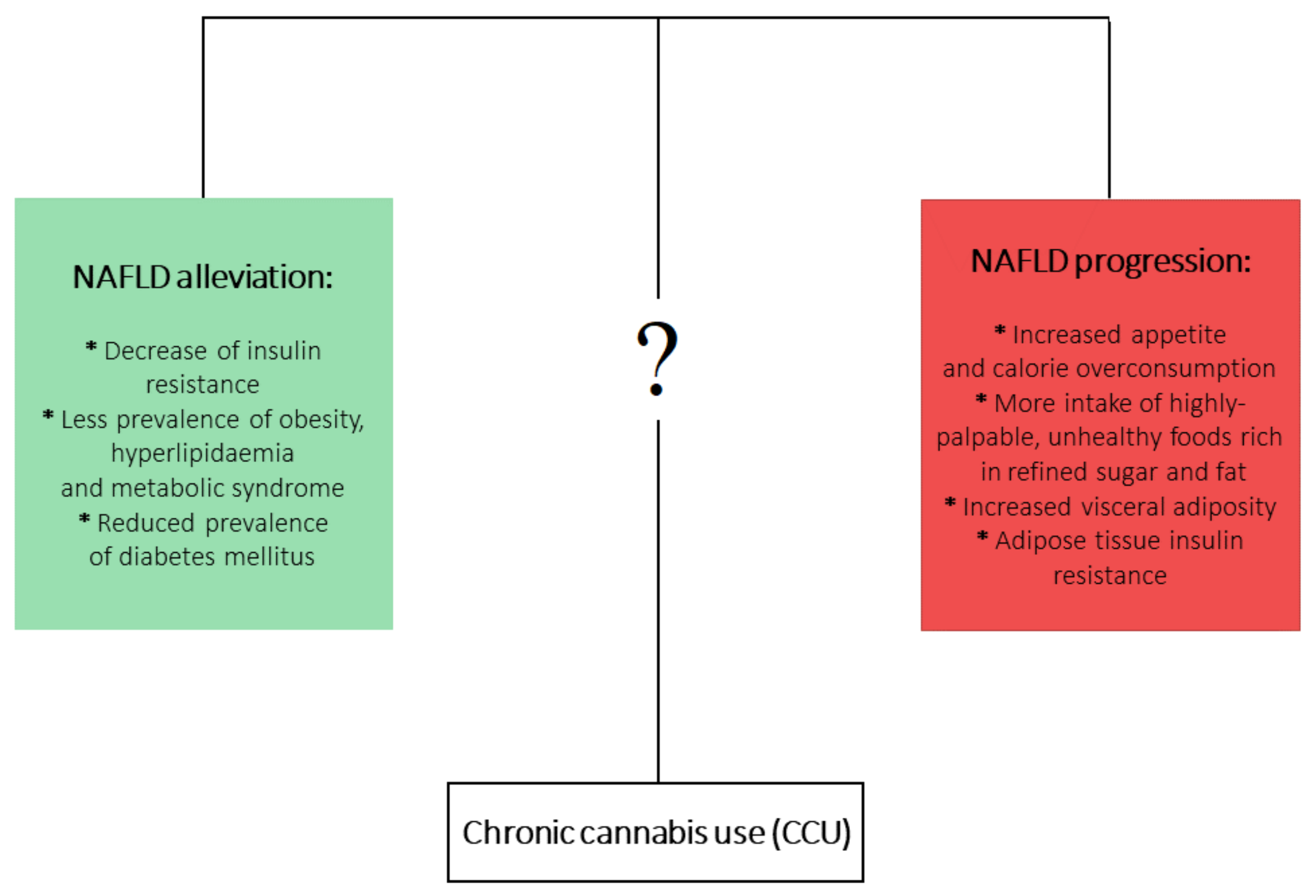
5. Effects of Prolonged Cannabis Use in the Context of NAFLD and its Comorbidities
6. THC and NAFLD
7. CBD and NAFLD
8. THCV and NAFLD
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kanwar, P.; Kowdley, K.V. The Metabolic Syndrome and Its Influence on Nonalcoholic Steatohepatitis. Clin. Liver Dis. 2016, 20, 225–243. [Google Scholar] [CrossRef] [PubMed]
- Chalasani, N.; Younossi, Z.; Lavine, J.E.; Diehl, A.M.; Brunt, E.M.; Cusi, K.; Charlton, M.; Sanyal, A.J. The diagnosis and management of non-alcoholic fatty liver disease: Practice Guideline by the American Association for the Study of Liver Diseases, American College of Gastroenterology, and the American Gastroenterological Association. Hepatology 2012, 55, 2005–2023. [Google Scholar] [CrossRef] [PubMed]
- Lonardo, A.; Bellentani, S.; Argo, C.K.; Ballestri, S.; Byrne, C.D.; Caldwell, S.H.; Cortez-Pinto, H.; Grieco, A.; Machado, M.V.; Miele, L.; et al. Epidemiological modifiers of non-alcoholic fatty liver disease: Focus on high-risk groups. Dig. Liver Dis. 2015, 47, 997–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilg, H.; Moschen, A.R. Evolution of inflammation in nonalcoholic fatty liver disease: The multiple parallel hits hypothesis. Hepatology 2010, 52, 1836–1846. [Google Scholar] [CrossRef]
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD). Metabolism 2016, 65, 1038–1048. [Google Scholar] [CrossRef]
- Cristino, L.; Bisogno, T.; Di Marzo, V. Cannabinoids and the expanded endocannabinoid system in neurological disorders. Nat. Rev. Neurol. 2020, 16, 9–29. [Google Scholar] [CrossRef]
- Maccarrone, M.; Bab, I.; Bíró, T.; Cabral, G.A.; Dey, S.K.; Di Marzo, V.; Konje, J.C.; Kunos, G.; Mechoulam, R.; Pacher, P.; et al. Endocannabinoid signaling at the periphery: 50 years after THC. Trends Pharmacol. Sci. 2015, 36, 277–296. [Google Scholar] [CrossRef] [Green Version]
- Baggelaar, M.P.; Maccarrone, M.; van der Stelt, M. 2-Arachidonoylglycerol: A signaling lipid with manifold actions in the brain. Prog. Lipid Res. 2018, 71, 1–17. [Google Scholar] [CrossRef]
- Horne, E.A.; Stella, N. The ins and outs of endocannabinoid signaling in healthy and diseased brain. Future Lipidol. 2008, 3, 435–452. [Google Scholar] [CrossRef]
- Siegmund, S.V.; Qian, T.; De Minicis, S.; Harvey-White, J.; Kunos, G.; Vinod, K.Y.; Hungund, B.; Schwabe, R.F. The endocannabinoid 2-arachidonoyl glycerol induces death of hepatic stellate cells via mitochondrial reactive oxygen species. FASEB J. 2007, 21, 2798–2806. [Google Scholar] [CrossRef]
- Urquhart, P.; Nicolaou, A.; Woodward, D.F. Endocannabinoids and their oxygenation by cyclo-oxygenases, lipoxygenases and other oxygenases. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2015, 1851, 366–376. [Google Scholar] [CrossRef] [PubMed]
- De Bus, I.; Witkamp, R.; Zuilhof, H.; Albada, B.; Balvers, M. The role of n-3 PUFA-derived fatty acid derivatives and their oxygenated metabolites in the modulation of inflammation. Prostaglandins Other Lipid Mediat. 2019, 144, 106351. [Google Scholar] [CrossRef] [PubMed]
- Herkenham, M.; Lynn, A.B.; Little, M.D.; Johnson, M.R.; Melvin, L.S.; De Costa, B.R.; Rice, K.C. Cannabinoid receptor localization in brain. Proc. Natl. Acad. Sci. USA 1990, 87, 1932–1936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, S.; Kumar, U. Cannabinoid receptors and the endocannabinoid system: Signaling and function in the central nervous system. Int. J. Mol. Sci. 2018, 19, 833. [Google Scholar]
- Pacher, P.; Mechoulam, R. Is lipid signaling through cannabinoid 2 receptors part of a protective system? Prog. Lipid Res. 2011, 50, 193–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wold, E.A.; Chen, J.; Cunningham, K.A.; Zhou, J. Allosteric Modulation of Class A GPCRs: Targets, Agents, and Emerging Concepts. J. Med. Chem. 2019, 62, 88–127. [Google Scholar] [CrossRef]
- Ye, L.; Cao, Z.; Wang, W.; Zhou, N. New Insights in Cannabinoid Receptor Structure and Signaling. Curr. Mol. Pharmacol. 2019, 12, 239–248. [Google Scholar] [CrossRef]
- Morales, P.; Reggio, P.H. GPCRs and Insulin Receptor Signaling in Conversation: Novel Avenues for Drug Discovery. Cannabis Cannabinoid Res. 2017, 19, 1436–1444. [Google Scholar]
- Bazwinsky-Wutschke, I.; Zipprich, A.; Dehghani, F. Endocannabinoid System in Hepatic Glucose Metabolism, Fatty Liver Disease, and Cirrhosis. Int. J. Mol. Sci. 2019, 20, 2516. [Google Scholar] [CrossRef] [Green Version]
- Lipina, C.; Walsh, S.K.; Mitchell, S.E.; Speakman, J.R.; Wainwright, C.L.; Hundal, H.S. GPR55 deficiency is associated with increased adiposity and impaired insulin signaling in peripheral metabolic tissues. FASEB J. 2019, 33, 1299–1312. [Google Scholar] [CrossRef] [Green Version]
- Balenga, N.A.B.; Aflaki, E.; Kargl, J.; Platzer, W.; Schröder, R.; Blättermann, S.; Kostenis, E.; Brown, A.J.; Heinemann, A.; Waldhoer, M. GPR55 regulates cannabinoid 2 receptor-mediated responses in human neutrophils. Cell Res. 2011, 21, 1452–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simcocks, A.C.; O’Keefe, L.; Jenkin, K.A.; Mathai, M.L.; Hryciw, D.H.; McAinch, A.J. A potential role for GPR55 in the regulation of energy homeostasis. Drug Discov. Today 2014, 19, 1145–1151. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.W.; Kim, H.S.; Im, J.H.; Kim, J.W.; Jun, D.W.; Lim, S.C.; Lee, K.; Choi, J.M.; Kim, S.K.; Kang, K.W. GPR119: A promising target for nonalcoholic fatty liver disease. FASEB J. 2016, 30, 324–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajaraman, G.; Simcocks, A.; Hryciw, D.H.; Hutchinson, D.S.; McAinch, A.J. G protein coupled receptor 18: A potential role for endocannabinoid signaling in metabolic dysfunction. Mol. Nutr. Food Res. 2016, 60, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.W.; Kim, H.S.; Choi, Y.W.; Kim, Y.M.; Kang, K.W. Therapeutic application of GPR119 ligands in metabolic disorders. Diabetes Obes. Metab. 2018, 20, 257–269. [Google Scholar] [CrossRef]
- Reggio, P.H.; Shore, D.M. The therapeutic potential of orphan GPCRs, GPR35 and GPR55. Front. Pharmacol. 2015, 6, 69. [Google Scholar]
- Tam, J.; Liu, J.; Mukhopadhyay, B.; Cinar, R.; Godlewski, G.; Kunos, G. Endocannabinoids in liver disease. Hepatology 2011, 53, 346–355. [Google Scholar] [CrossRef] [Green Version]
- González-Mariscal, I.; Krzysik-Walker, S.M.; Doyle, M.E.; Liu, Q.R.; Cimbro, R.; Santa-Cruz Calvo, S.; Ghosh, S.; Cieala, A.; Moaddel, R.; Carlson, O.D.; et al. Human CB1 Receptor Isoforms, present in Hepatocytes and β-cells, are Involved in Regulating Metabolism. Sci. Rep. 2016, 6, 33302. [Google Scholar] [CrossRef] [Green Version]
- Turcotte, C.; Blanchet, M.R.; Laviolette, M.; Flamand, N. The CB2 receptor and its role as a regulator of inflammation. Cell. Mol. Life Sci. 2016, 73, 4449–4470. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.M.; Kim, T.H.; Kim, A.; Koo, J.H.; Joo, M.S.; Kim, S.G. Liver X Receptor α–Induced Cannabinoid Receptor 2 Inhibits Ubiquitin-Specific Peptidase 4 Through miR-27b, Protecting Hepatocytes From TGF-β. Hepatol. Commun. 2019, 3, 1373–1387. [Google Scholar] [CrossRef] [Green Version]
- Busquets-Garcia, A.; Bains, J.; Marsicano, G. CB 1 Receptor Signaling in the Brain: Extracting Specificity from Ubiquity. Neuropsychopharmacology 2018, 43, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Bazwinsky-Wutschke, I.; Zipprich, A.; Dehghani, F. Daytime-Dependent changes of cannabinoid receptor type 1 and type 2 expression in rat liver. Int. J. Mol. Sci. 2017, 18, 1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaughn, L.K.; Denning, G.; Stuhr, K.L.; De Wit, H.; Hill, M.N.; Hillard, C.J. Endocannabinoid signalling: Has it got rhythm? Br. J. Pharmacol. 2010, 160, 530–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, M.D.; Sakurai, T.; Rainero, I.; Maj, M.C.; Kukkonen, J.P. Orexin receptor multimerization versus functional interactions: Neuropharmacological implications for opioid and cannabinoid signalling and pharmacogenetics. Pharmaceuticals 2017, 10, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, N.; Onaivi, E.S. Endocannabinoid System Components: Overview and Tissue Distribution. In Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Wang, M.; Meng, N.; Chang, Y.; Tang, W. Endocannabinoids signaling: Molecular mechanisms of liver regulation and diseases. Front. Biosci. Landmark 2016, 21, 1488–1501. [Google Scholar]
- Ruiz de Azua, I.; Lutz, B. Multiple endocannabinoid-mediated mechanisms in the regulation of energy homeostasis in brain and peripheral tissues. Cell. Mol. Life Sci. 2019, 76, 1341–1363. [Google Scholar] [CrossRef]
- Agudo, J.; Martin, M.; Roca, C.; Molas, M.; Bura, A.S.; Zimmer, A.; Bosch, F.; Maldonado, R. Deficiency of CB2 cannabinoid receptor in mice improves insulin sensitivity but increases food intake and obesity with age. Diabetologia 2010, 53, 2629–2640. [Google Scholar] [CrossRef] [Green Version]
- Kirkham, T.C. Endocannabinoids in the regulation of appetite and body weight. Behav. Pharmacol. 2005, 16, 297–313. [Google Scholar] [CrossRef]
- Miranda, R.A.; De Almeida, M.M.; Da Rocha, C.P.D.; de Brito Fassarella, L.; De Souza, L.L.; De Souza, A.F.P.; De Andrade, C.B.V.; Fortunato, R.S.; Pazos-Moura, C.C.; Trevenzoli, I.H. Maternal high-fat diet consumption induces sex-dependent alterations of the endocannabinoid system and redox homeostasis in liver of adult rat offspring. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Osei-Hyiaman, D.; DePetrillo, M.; Pacher, P.; Liu, J.; Radaeva, S.; Bátkai, S.; Harvey-White, J.; Mackie, K.; Offertáler, L.; Wang, L.; et al. Endocannabinoid activation at hepatic CB 1 receptors stimulates fatty acid synthesis and contributes to diet-induced obesity. J. Clin. Investig. 2005, 115, 1298–1305. [Google Scholar] [CrossRef] [Green Version]
- De Gottardi, A.; Spahr, L.; Ravier-Dall’Antonia, F.; Hadengue, A. Cannabinoid receptor 1 and 2 agonists increase lipid accumulation in hepatocytes. Liver Int. 2010, 30, 1482–1489. [Google Scholar] [CrossRef] [PubMed]
- Dibba, P.; Li, A.; Cholankeril, G.; Iqbal, U.; Gadiparthi, C.; Khan, M.; Kim, D.; Ahmed, A. Mechanistic Potential and Therapeutic Implications of Cannabinoids in Nonalcoholic Fatty Liver Disease. Medicines 2018, 5, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Cinar, R.; Xiong, K.; Godlewski, G.; Jourdan, T.; Lin, Y.; Ntambi, J.M.; Kunos, G. Monounsaturated fatty acids generated via stearoyl CoA desaturase-1 are endogenous inhibitors of fatty acid amide hydrolase. Proc. Natl. Acad. Sci. USA 2013, 110, 18832–18837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osei-Hyiaman, D.; Liu, J.; Zhou, L.; Godlewski, G.; Harvey-White, J.; Jeong, W.I.; Bátkai, S.; Marsicano, G.; Lutz, B.; Buettner, C.; et al. Hepatic CB1 receptor is required for development of diet-induced steatosis, dyslipidemia, and insulin and leptin resistance in mice. J. Clin. Investig. 2008, 118, 3160–3169. [Google Scholar] [CrossRef] [PubMed]
- Tam, J.; Godlewski, G.; Earley, B.J.; Zhou, L.; Jourdan, T.; Szanda, G.; Cinar, R.; Kunos, G. Role of adiponectin in the metabolic effects of cannabinoid type 1 receptor blockade in mice with diet-induced obesity. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E457–E468. [Google Scholar] [CrossRef]
- Irungbam, K.; Churin, Y.; Matono, T.; Weglage, J.; Ocker, M.; Glebe, D.; Hardt, M.; Koeppel, A.; Roderfeld, M.; Roeb, E. Cannabinoid receptor 1 knockout alleviates hepatic steatosis by downregulating perilipin 2. Lab. Investig. 2019, 100, 454–465. [Google Scholar] [CrossRef]
- Vernon, G.; Baranova, A.; Younossi, Z.M. Systematic review: The epidemiology and natural history of non-alcoholic fatty liver disease and non-alcoholic steatohepatitis in adults. Aliment. Pharmacol. Ther. 2011, 34, 274–285. [Google Scholar] [CrossRef]
- Mu, W.; Xuefang, C.; Liu, Y.; Qianzhou, L.; Gaolin, L.; Jigang, Z.; Xiaoyu, L. Potential nexus of non-alcoholic fatty liver disease and type 2 diabetes mellitus: Insulin resistance between hepatic and peripheral tissues. Front. Pharmacol. 2019, 9, 1566. [Google Scholar] [CrossRef] [Green Version]
- Sabio, G.; Das, M.; Mora, A.; Zhang, Z.; Jun, J.Y.; Hwi, J.K.; Barrett, T.; Kim, J.K.; Davis, R.J. A stress signaling pathway in adipose tissue regulates hepatic insulin resistance. Science 2008, 322, 1539–1543. [Google Scholar] [CrossRef] [Green Version]
- O’Hare, J.D.; Zieliński, E.; Cheng, B.; Scherer, T.; Buettner, C. Central endocannabinoid signaling regulates hepatic glucose production and systemic lipolysis. Diabetes 2011, 60, 1055–1062. [Google Scholar] [CrossRef] [Green Version]
- Jourdan, T.; Godlewski, G.; Kunos, G. Endocannabinoid regulation of β-cell functions: Implications for glycaemic control and diabetes. Diabetes Obes. Metab. 2016, 18, 549–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chanda, D.; Kim, D.K.; Li, T.; Kim, Y.H.; Koo, S.H.; Lee, C.H.; Chiang, J.Y.L.; Choi, H.S. Cannabinoid Receptor Type 1 (CB1R) signaling regulates hepatic gluconeogenesis via induction of endoplasmic reticulum-bound transcription factor cAMP-responsive element-binding protein H (CREBH) in primary hepatocytes. J. Biol. Chem. 2011, 286, 27971–27979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Zhou, L.; Xiong, K.; Godlewski, G.; Mukhopadhyay, B.; Tam, J.; Yin, S.; Gao, P.; Shan, X.; Pickel, J.; et al. Hepatic cannabinoid receptor-1 mediates diet-induced insulin resistance via inhibition of insulin signaling and clearance in mice. Gastroenterology 2012, 142, 1218–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bermudez-Silva, F.J.; Sanchez-Vera, I.; Suárez, J.; Serrano, A.; Fuentes, E.; Juan-Pico, P.; Nadal, A.; Rodríguez de Fonseca, F. Role of cannabinoid CB2 receptors in glucose homeostasis in rats. Eur. J. Pharmacol. 2007, 565, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.; Bellini, G.; Luongo, L.; Manzo, I.; Tolone, S.; Tortora, C.; Bernardo, M.E.; Grandone, A.; Conforti, A.; Docimo, L.; et al. Cannabinoid receptor 2 as antiobesity target: Inflammation, fat storage, and browning modulation. J. Clin. Endocrinol. Metab. 2016, 101, 3469–3478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deveaux, V.; Cadoudal, T.; Ichigotani, Y.; Teixeira-Clerc, F.; Louvet, A.; Manin, S.; Van Nhieu, J.T.; Belot, M.P.; Zimmer, A.; Even, P.; et al. Cannabinoid CB2 receptor potentiates obesity-associated inflammation, insulin resistance and hepatic steatosis. PLoS ONE 2009, 4, e5844. [Google Scholar] [CrossRef]
- Sáez, T.; Salsoso, R.; Leiva, A.; Toledo, F.; de Vos, P.; Faas, M.; Sobrevia, L. Human umbilical vein endothelium-derived exosomes play a role in foetoplacental endothelial dysfunction in gestational diabetes mellitus. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 499–508. [Google Scholar] [CrossRef]
- Atwood, B.K.; MacKie, K. CB 2: A cannabinoid receptor with an identity crisis. Br. J. Pharmacol. 2010, 160, 467–479. [Google Scholar] [CrossRef] [Green Version]
- Mendez-Sanchez, N.; Zamora-Valdes, D.; Pichardo-Bahena, R.; Barredo-Prieto, B.; Ponciano-Rodriguez, G.; Bermejo-Martínez, L.; Chavez-Tapia, N.C.; Baptista-González, H.A.; Uribe, M. Endocannabinoid receptor CB2 in nonalcoholic fatty liver disease. Liver Int. 2007, 27, 215–219. [Google Scholar] [CrossRef]
- Auguet, T.; Berlanga, A.; Guiu-Jurado, E.; Terra, X.; Martinez, S.; Aguilar, C.; Filiu, E.; Alibalic, A.; Sabench, F.; Hernández, M.; et al. Endocannabinoid receptors gene expression in morbidly obese women with nonalcoholic fatty liver disease. Biomed Res. Int. 2014, 2014, 502542. [Google Scholar] [CrossRef]
- Dibba, P.; Li, A.; Cholankeril, G.; Iqbal, U.; Gadiparthi, C.; Khan, M.; Kim, D.; Ahmed, A. The Role of Cannabinoids in the Setting of Cirrhosis. Medicines 2018, 5, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorgačević, B.; Vučević, D.; Vesković, M.; Mladenović, D.; Vukićević, D.; Vukićević, R.J.; Todorović, V.; Radosavljević, T. The effect of cannabinoid receptor 1 blockade on adipokine and proinflammatory cytokine concentration in adipose and hepatic tissue in mice with nonalcoholic fatty liver disease. Can. J. Physiol. Pharmacol. 2019, 97, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Baldassarre, M.; Giannone, F.A.; Napoli, L.; Tovoli, A.; Ricci, C.S.; Tufoni, M.; Caraceni, P. The endocannabinoid system in advanced liver cirrhosis: Pathophysiological implication and future perspectives. Liver Int. 2013, 33, 1298–1308. [Google Scholar] [CrossRef] [PubMed]
- Julien, B.; Grenard, P.; Teixeira-Clerc, F.; Van Nhieu, J.T.; Li, L.; Karsak, M.; Zimmer, A.; Mallat, A.; Lotersztajn, S. Antifibrogenic role of the cannabinoid receptor CB2 in the liver. Gastroenterology 2005, 128, 742–755. [Google Scholar] [CrossRef]
- Guillot, A.; Hamdaoui, N.; Bizy, A.; Zoltani, K.; Souktani, R.; Zafrani, E.S.; Mallat, A.; Lotersztajn, S.; Lafdil, F. Cannabinoid receptor 2 counteracts interleukin-17-induced immune and fibrogenic responses in mouse liver. Hepatology 2014, 59, 296–306. [Google Scholar] [CrossRef]
- Muñoz-Luque, J.; Ros, J.; Fernández-Varo, G.; Tugues, S.; Morales-Ruiz, M.; Alvarez, C.E.; Friedman, S.L.; Arroyo, V.; Jiménez, W. Regression of fibrosis after chronic stimulation of cannabinoid CB2 receptor in cirrhotic rats. J. Pharmacol. Exp. Ther. 2008, 324, 475–483. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, S.E. Cannabinoid activation of peroxisome proliferator-activated receptors: An update and review of the physiological relevance. Wiley Interdiscip. Rev. Membr. Transp. Signal. 2013, 2, 17–25. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, S.E. An update on PPAR activation by cannabinoids. Br. J. Pharmacol. 2016, 173, 1899–1910. [Google Scholar] [CrossRef] [Green Version]
- Jin, D.; Dai, K.; Xie, Z.; Chen, J. Secondary Metabolites Profiled in Cannabis Inflorescences, Leaves, Stem Barks, and Roots for Medicinal Purposes. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Aizpurua-Olaizola, O.; Soydaner, U.; Öztürk, E.; Schibano, D.; Simsir, Y.; Navarro, P.; Etxebarria, N.; Usobiaga, A. Evolution of the Cannabinoid and Terpene Content during the Growth of Cannabis sativa Plants from Different Chemotypes. J. Nat. Prod. 2016, 79, 324–331. [Google Scholar] [CrossRef]
- Wu, J. Cannabis, cannabinoid receptors, and endocannabinoid system: Yesterday, today, and tomorrow. Acta Pharmacol. Sin. 2019, 40, 297–299. [Google Scholar] [CrossRef] [PubMed]
- Palomares, B.; Ruiz-Pino, F.; Navarrete, C.; Velasco, I.; Sánchez-Garrido, M.A.; Jimenez-Jimenez, C.; Pavicic, C.; Vazquez, M.J.; Appendino, G.; Bellido, M.L.; et al. Tetrahydrocannabinolic acid A (THCA-A) reduces adiposity and prevents metabolic disease caused by diet-induced obesity. Sci. Rep. 2019, 171, 113693. [Google Scholar] [CrossRef] [PubMed]
- Franco, R.; Rivas-Santisteban, R.; Reyes-Resina, I.; Casanovas, M.; Pérez-Olives, C.; Ferreiro-Vera, C.; Navarro, G.; Sánchez de Medina, V.; Nadal, X. Pharmacological potential of varinic-, minor-, and acidic phytocannabinoids. Pharmacol. Res. 2020, 158, 104801. [Google Scholar] [CrossRef] [PubMed]
- McPartland, J.M.; Duncan, M.; Di Marzo, V.; Pertwee, R.G. Are cannabidiol and Δ9-tetrahydrocannabivarin negative modulators of the endocannabinoid system? A systematic review. Br. J. Pharmacol. 2015, 172, 737–753. [Google Scholar] [CrossRef] [Green Version]
- Goyal, H.; Rahman, M.R.; Perisetti, A.; Shah, N.; Chhabra, R. Cannabis in liver disorders: A friend or a foe? Eur. J. Gastroenterol. Hepatol. 2018, 30, 1283–1290. [Google Scholar] [CrossRef]
- Laprairie, R.B.; Bagher, A.M.; Kelly, M.E.M.; Denovan-Wright, E.M. Cannabidiol is a negative allosteric modulator of the cannabinoid CB1 receptor. Br. J. Pharmacol. 2015, 172, 4790–4805. [Google Scholar] [CrossRef] [Green Version]
- Pertwee, R.G. The diverse CB 1 and CB 2 receptor pharmacology of three plant cannabinoids: Δ 9-tetrahydrocannabinol, cannabidiol and Δ 9-tetrahydrocannabivarin. Br. J. Pharmacol. 2008, 153, 199–215. [Google Scholar] [CrossRef] [Green Version]
- Morales, P.; Hurst, D.P.; Reggio, P.H. Molecular Targets of the Phytocannabinoids: A Complex Picture. In Progress in the Chemistry of Organic Natural Products; Springer: Cham, Switzerland, 2017; Volume 103, pp. 103–131. [Google Scholar]
- Borgelt, L.M.; Franson, K.L.; Nussbaum, A.M.; Wang, G.S. The pharmacologic and clinical effects of medical cannabis. Pharmacotherapy 2013, 33, 195–209. [Google Scholar] [CrossRef]
- Product Monograph. Eliquis®. 2010. ISBN 1866234234. Available online: https://www.pfizer.ca/sites/default/files/201910/ELIQUIS_PM_229267_07Oct2019_Marketed_E.pdf (accessed on 20 November 2020).
- U.S. Food and Drug Administration. Cesamet (nabilone) Capsules. Nda 18-677/S-011; 2006. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2006/018677s011lbl.pdf (accessed on 10 November 2020).
- U.S. Food and Drug Administration. Marinol (Dronabinol) Technical Sheet. Nda 18-651/s-021; 2004. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2005/018651s021lbl.pdf (accessed on 10 November 2020).
- Greenwich Biosciences Inc. Full prescribing information of EPIDIOLEX. Prescr. Inf. Leafl.; 2018. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2018/210365lbl.pdf (accessed on 10 November 2020).
- GW Pharma, L. Part III: Consumer Information Sativex®. Prod. Monogr.; 2015. Available online: https://pdf.hres.ca/dpd_pm/00016162.PDF (accessed on 10 November 2020).
- Kirkham, T.C. Cannabinoids and appetite: Food craving and food pleasure. Int. Rev. Psychiatry 2009, 21, 163–171. [Google Scholar] [CrossRef]
- Foltin, R.W.; Fischman, M.W.; Byrne, M.F. Effects of smoked marijuana on food intake and body weight of humans living in a residential laboratory. Appetite 1988, 11, 1–14. [Google Scholar] [CrossRef]
- Rodondi, N.; Pletcher, M.J.; Liu, K.; Hulley, S.B.; Sidney, S. Marijuana Use, Diet, Body Mass Index, and Cardiovascular Risk Factors (from the CARDIA Study). Am. J. Cardiol. 2006, 98, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Rajavashisth, T.B.; Shaheen, M.; Norris, K.C.; Pan, D.; Sinha, S.K.; Ortega, J.; Friedman, T.C. Decreased prevalence of diabetes in marijuana users: Cross-sectional data from the National Health and Nutrition Examination Survey (NHANES) III. BMJ Open 2012, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smit, E.; Crespo, C.J. Dietary intake and nutritional status of US adult marijuana users: Results from the Third National Health and Nutrition Examination Survey. Public Health Nutr. 2001, 4, 781–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penner, E.A.; Buettner, H.; Mittleman, M.A. The impact of marijuana use on glucose, insulin, and insulin resistance among US adults. Am. J. Med. 2013, 126, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Carrieri, M.P. Cannabis use and reduced risk of insulin-resistance in HIV-HCV infected patients: A longitudinal analysis (ANRS HEPAVIH CO-13). Clin. Infect. Dis. 2015, 61, 40–48. [Google Scholar] [CrossRef]
- Le Strat, Y.; Le Foll, B. Obesity and cannabis use: Results from 2 representative national surveys. Am. J. Epidemiol. 2011, 174, 929–933. [Google Scholar] [CrossRef] [Green Version]
- Vidot, D.C.; Prado, G.; Hlaing, W.W.M.; Florez, H.J.; Arheart, K.L.; Messiah, S.E. Metabolic Syndrome Among Marijuana Users in the United States: An Analysis of National Health and Nutrition Examination Survey Data. Am. J. Med. 2016, 129, 173–179. [Google Scholar] [CrossRef] [Green Version]
- Muniyappa, R.; Sable, S.; Ouwerkerk, R.; Mari, A.; Gharib, A.M.; Walter, M.; Courville, A.; Hall, G.; Chen, K.Y.; Volkow, N.D.; et al. Metabolic effects of chronic cannabis smoking. Diabetes Care 2013, 36, 2415–2422. [Google Scholar] [CrossRef] [Green Version]
- Sansone, R.A.; Sansone, L.A. Marijuana and body weight. Innov. Clin. Neurosci. 2014, 11, 50–54. [Google Scholar]
- Hayatbakhsh, M.R.; O’Callaghan, M.J.; Mamun, A.A.; Williams, G.M.; Clavarino, A.; Najman, J.M. Cannabis use and obesity and young adults. Am. J. Drug Alcohol Abuse 2010, 36, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Kim, W.; Kwak, M.S.; Chung, G.E.; Yim, J.Y.; Ahmed, A. Inverse association of marijuana use with nonalcoholic fatty liver disease among adults in the United States. PLoS ONE 2017, 12, e0186702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vázquez-Bourgon, J.; Ortiz-García de la Foz, V.; Suarez-Pereira, I.; Iruzubieta, P.; Arias-Loste, M.T.; Setién-Suero, E.; Ayesa-Arriola, R.; Gómez-Revuelta, M.; Crespo, J.; Crespo Facorro, B. Cannabis consumption and non-alcoholic fatty liver disease. A three years longitudinal study in first episode non-affective psychosis patients. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 95, 109677. [Google Scholar] [CrossRef] [PubMed]
- Adejumo, A.C.; Alliu, S.; Ajayi, T.O.; Adejumo, K.L.; Adegbala, O.M.; Onyeakusi, N.E.; Akinjero, A.M.; Durojaiye, M.; Bukong, T.N. Cannabis use is associated with reduced prevalence of non-alcoholic fatty liver disease: A cross-sectional study. PLoS ONE 2017, 12, e0176416. [Google Scholar] [CrossRef]
- Thomas, A.; Baillie, G.L.; Phillips, A.M.; Razdan, R.K.; Ross, R.A.; Pertwee, R.G. Cannabidiol displays unexpectedly high potency as an antagonist of CB 1 and CB 2 receptor agonists in vitro. Br. J. Pharmacol. 2007, 150, 613–623. [Google Scholar] [CrossRef] [Green Version]
- Thomas, A.; Stevenson, L.A.; Wease, K.N.; Price, M.R.; Baillie, G.; Ross, R.A.; Pertwee, R.G. Evidence that the plant cannabinoid Δ 9- tetrahydrocannabivarin is a cannabinoid CB 1 and CB 2 receptor antagonist. Br. J. Pharmacol. 2005, 146, 917–926. [Google Scholar] [CrossRef] [Green Version]
- Younossi, Z.; Anstee, Q.M.; Marietti, M.; Hardy, T.; Henry, L.; Eslam, M.; George, J.; Bugianesi, E. Global burden of NAFLD and NASH: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 11. [Google Scholar] [CrossRef]
- Braunersreuther, V.; Viviani, G.L.; Mach, F.; Montecucco, F. Role of cytokines and chemokines in non-alcoholic fatty liver disease. World J. Gastroenterol. 2012, 18, 727–735. [Google Scholar] [CrossRef]
- Huang, Y.; Wan, T.; Pang, N.; Zhou, Y.; Jiang, X.; Li, B.; Gu, Y.; Huang, Y.; Ye, X.; Lian, H.; et al. Cannabidiol protects livers against nonalcoholic steatohepatitis induced by high-fat high cholesterol diet via regulating NF-κB and NLRP3 inflammasome pathway. J. Cell. Physiol. 2019, 234, 21224–21234. [Google Scholar] [CrossRef]
- Ribeiro, A.; Ferraz-De-Paula, V.; Pinheiro, M.L.; Vitoretti, L.B.; Mariano-Souza, D.P.; Quinteiro-Filho, W.M.; Akamine, A.T.; Almeida, V.I.; Quevedo, J.; Dal-Pizzol, F.; et al. Cannabidiol, a non-psychotropic plant-derived cannabinoid, decreases inflammation in a murine model of acute lung injury: Role for the adenosine A 2A receptor. Eur. J. Pharmacol. 2012, 678, 78–85. [Google Scholar] [CrossRef]
- Hirvonen, J.; Goodwin, R.S.; Li, C.T.; Terry, G.E.; Zoghbi, S.S.; Morse, C.; Pike, V.W.; Volkow, N.D.; Huestis, M.A.; Innis, R.B. Reversible and regionally selective downregulation of brain cannabinoid CB 1 receptors in chronic daily cannabis smokers. Mol. Psychiatry 2012, 17, 642–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mechoulam, R. Plant cannabinoids: A neglected pharmacological treasure trove. Br. J. Pharmacol. 2005, 146, 913–915. [Google Scholar] [CrossRef] [PubMed]
- Dibba, P.; Li, A.A.; Cholankeril, G.; Ali Khan, M.; Kim, D.; Ahmed, A. Potential Mechanisms Influencing the Inverse Relationship Between Cannabis and Nonalcoholic Fatty Liver Disease: A Commentary. Nutr. Metab. Insights 2019, 12, 117863881984748. [Google Scholar] [CrossRef] [PubMed]
- Mridha, A.R.; Wree, A.; Robertson, A.A.B.; Yeh, M.M.; Johnson, C.D.; Van Rooyen, D.M.; Haczeyni, F.; Teoh, N.C.H.; Savard, C.; Ioannou, G.N.; et al. NLRP3 inflammasome blockade reduces liver inflammation and fibrosis in experimental NASH in mice. J. Hepatol. 2017, 66, 1037–1046. [Google Scholar] [CrossRef]
- Koppel, B.S.; Brust, J.C.M.; Fife, T.; Bronstein, J.; Youssof, S.; Gronseth, G.; Gloss, D. Systematic review: Efficacy and safety of medical marijuana in selected neurologic disorders: Report of the Guideline Development Subcommittee of the American Academy of Neurology. Neurology 2014, 82, 1556–1563. [Google Scholar] [CrossRef]
- Maroon Joseph, B.J. Review of the neurological benefits of phytocannabinoids. Surg. Neurol. Int. 2018, 9, 91. [Google Scholar] [CrossRef]
- Wiley, J.L.; Burston, J.J.; Leggett, D.C.; Alekseeva, O.O.; Razdan, R.K.; Mahadevan, A.; Martin, B.R. CB 1 cannabinoid receptor-mediated modulation of food intake in mice. Br. J. Pharmacol. 2005, 145, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Koch, J.E.; Matthews, S.M. Δ9-tetrahydrocannabinol stimulates palatable food intake in lewis rats: Effects of peripheral and central administration. Nutr. Neurosci. 2001, 4, 179–187. [Google Scholar] [CrossRef]
- McIntosh, A.L.; Martin, G.G.; Huang, H.; Landrock, D.; Kier, A.B.; Schroeder, F. Δ9-Tetrahydrocannabinol induces endocannabinoid accumulation in mouse hepatocytes: Antagonism by Fabp1 gene ablation. J. Lipid Res. 2018, 59, 646–657. [Google Scholar] [CrossRef] [Green Version]
- Farrimond, J.A.; Hill, A.J.; Whalley, B.J.; Williams, C.M. Cannabis constituents modulate δ9- tetrahydrocannabinol induced hyperphagia in rats. Psychopharmacology 2010, 210, 97–106. [Google Scholar] [CrossRef]
- Laun, A.S.; Shrader, S.H.; Brown, K.J.; Song, Z.H. GPR3, GPR6, and GPR12 as novel molecular targets: Their biological functions and interaction with cannabidiol. Acta Pharmacol. Sin. 2019, 40, 300–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, C.M. A Review of Human Studies Assessing Cannabidiol’s (CBD) Therapeutic Actions and Potential. J. Clin. Pharmacol. 2019, 59, 923–934. [Google Scholar] [CrossRef] [PubMed]
- Viudez-Martínez, A.; García-Gutiérrez, M.S.; Medrano-Relinque, J.; Navarrón, C.M.; Navarrete, F.; Manzanares, J. Cannabidiol does not display drug abuse potential in mice behavior. Acta Pharmacol. Sin. 2019, 40, 358–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iffland, K.; Grotenhermen, F. An Update on Safety and Side Effects of Cannabidiol: A Review of Clinical Data and Relevant Animal Studies. Cannabis Cannabinoid Res. 2017, 2, 139–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewing, L.E.; Skinner, C.M.; Quick, C.M.; Kennon-McGill, S.; McGill, M.R.; Walker, L.A.; ElSohly, M.A.; Gurley, B.J.; Koturbash, I. Hepatotoxicity of a Cannabidiol-rich cannabis extract in the mouse model. Molecules 2019, 24, 1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Rozenfeld, R.; Wu, D.; Devi, L.A.; Zhang, Z.; Cederbaum, A. Cannabidiol protects liver from binge alcohol-induced steatosis by mechanisms including inhibition of oxidative stress and increase in autophagy. Free Radic. Biol. Med. 2014, 68, 260–267. [Google Scholar] [CrossRef] [Green Version]
- Costa, B.; Colleoni, M.; Conti, S.; Parolaro, D.; Franke, C.; Trovato, A.E.; Giagnoni, G. Oral anti-inflammatory activity of cannabidiol, a non-psychoactive constituent of cannabis, in acute carrageenan-induced inflammation in the rat paw. Naunyn Schmiedebergs Arch. Pharmacol. 2004, 369, 294–299. [Google Scholar] [CrossRef]
- Vilela, L.R.; Gomides, L.F.; David, B.A.; Antunes, M.M.; Diniz, A.B.; De Araújo Moreira, F.; Menezes, G.B. Cannabidiol rescues acute hepatic toxicity and seizure induced by cocaine. Mediators Inflamm. 2015, 2015, 523418. [Google Scholar] [CrossRef]
- Silvestri, C.; Paris, D.; Martella, A.; Melck, D.; Guadagnino, I.; Cawthorne, M.; Motta, A.; Di Marzo, V. Two non-psychoactive cannabinoids reduce intracellular lipid levels and inhibit hepatosteatosis. J. Hepatol. 2015, 62, 1382–1390. [Google Scholar] [CrossRef]
- Mukhopadhyay, P.; Rajesh, M.; Horváth, B.; Bátkai, S.; Park, O.; Tanchian, G.; Gao, R.Y.; Patel, V.; Wink, D.A.; Liaudet, L.; et al. Cannabidiol protects against hepatic ischemia/reperfusion injury by attenuating inflammatory signaling and response, oxidative/nitrative stress, and cell death. Free Radic. Biol. Med. 2011, 50, 1368–1381. [Google Scholar] [CrossRef] [Green Version]
- Czaja, A.J. Hepatic inflammation and progressive liver fibrosis in chronic liver disease. World J. Gastroenterol. 2014, 20, 2515–2532. [Google Scholar] [CrossRef] [PubMed]
- GW Research Ltd. Study to Evaluate the Effect of GWP42003 on Liver Fat Levels in Participants With Fatty Liver Disease; GW Research Ltd.: London, UK, 2014. [Google Scholar]
- GW Research Ltd. GWMD1092—GWP42003: GWP42004 Together Plus Alone in Type II Diabetes; GW Research Ltd.: London, UK, 2014. [Google Scholar]
- Jadoon, K.A.; Ratcliffe, S.H.; Barrett, D.A.; Thomas, E.L.; Stott, C.; Bell, J.D.; O’Sullivan, S.E.; Tan, G.D. Efficacy and safety of cannabidiol and tetrahydrocannabivarin on glycemic and lipid parameters in patients with type 2 diabetes: A randomized, double-blind, placebo-controlled, parallel group pilot study. Diabetes Care 2016, 39, 1777–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millar, S.A.; Stone, N.L.; Bellman, Z.D.; Yates, A.S.; England, T.J.; O’Sullivan, S.E. A systematic review of cannabidiol dosing in clinical populations. Br. J. Clin. Pharmacol. 2019, 85, 1888–1900. [Google Scholar] [CrossRef] [PubMed]
- Tudurí, E.; Imbernon, M.; Hernández-Bautista, R.J.; Tojo, M.; Fernø, J.; Diéguez, C.; Nogueiras, R. GPR55: A new promising target for metabolism? J. Mol. Endocrinol. 2009, 58, R191–R202. [Google Scholar] [CrossRef] [PubMed]
- Bjursell, M.; Ryberg, E.; Wu, T.; Greasley, P.J.; Bohlooly-Y, M.; Hjorth, S. Deletion of Gpr55 results in subtle effects on energy metabolism, motor activity and thermal pain sensation. PLoS ONE 2016, 11, e0167965. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.S.; Chen, H.; Sun, H.; Zhu, J.; Jew, C.P.; Wager-Miller, J.; Straiker, A.; Spencer, C.; Bradshaw, H.; Mackie, K.; et al. GPR55, a G-Protein Coupled Receptor for Lysophosphatidylinositol, Plays a Role in Motor Coordination. PLoS ONE 2013, 8, e60314. [Google Scholar] [CrossRef] [Green Version]
- Englund, A.; Atakan, Z.; Kralj, A.; Tunstall, N.; Murray, R.; Morrison, P. The effect of five day dosing with THCV on THC-induced cognitive, psychological and physiological effects in healthy male human volunteers: A placebo-controlled, double-blind, crossover pilot trial. J. Psychopharmacol. 2016, 30, 140–151. [Google Scholar] [CrossRef]
- Wargent, E.T.; Zaibi, M.S.; Silvestri, C.; Hislop, D.C.; Stocker, C.J.; Stott, C.G.; Guy, G.W.; Duncan, M.; Di Marzo, V.; Cawthorne, M.A. The cannabinoid Δ9-tetrahydrocannabivarin (THCV) ameliorates insulin sensitivity in two mouse models of obesity. Nutr. Diabetes 2013, 3, e68. [Google Scholar] [CrossRef]
- Rzepa, E.; Tudge, L.; McCabe, C. The CB1 neutral antagonist tetrahydrocannabivarin reduces default mode network and increases executive control network resting state functional connectivity in healthy volunteers. Int. J. Neuropsychopharmacol. 2016, 19, 1–7. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Feature | THC | CBD | Reference |
|---|---|---|---|
| Interaction with receptors: a) CB1R b) CB2R | + (partial agonist) - (weak antagonist) | - (negative allosteric modulator) - (inverse agonist) | [77,78,79] |
| Psychoactive effect | Yes | No | [80] |
| Appetite stimulation | Yes | No | [80] |
| Cardiovascular stimulation (inducing tachycardia and hypertension) | Yes | No | [80] |
| Anticonvulsant effect | Yes | No | [80] |
| Therapeutic indications approved by the FDA 1 | Anorexia associated with weight loss in AIDS 3 patients Nausea and vomiting associated with anticancer chemotherapy Multiple sclerosis spasticity | Lennox-Gastaut syndrome Dravet syndrome | [81,82,83,84] |
| Formulations available on US pharmaceutical market | Nabilone (trade name Cesamet) synthetic THC analog aviable as oral capsule Dronabinol (trade name Marinol)—synthetic formulation of the main THC constituent enantiomer found in Cannabis: [(−)-trans-Δ9-tetrahydrocannabinol] as an oily resin in capsules | Epidiolex®—pharmaceutical formulation of CBD as an oral solution | [82,83,84] |
| Combination drugs available on US 2 pharmaceutical market | Nabiximol (trade name Sativex)—oral spray standardized in composition, formulation, and dose, delivering of 2.7 mg THC and 2.5 mg CBD per dose. | [85] | |
| Main Outcome | Research Model | Participants | Method | Reference |
|---|---|---|---|---|
| ↓ prevalence of NAFLD | Population-based case-control study | 5,950,391 Cannabis users from the HCUP-NIS 7 database. | Presence of NAFLD diagnosis among patients. | [110] |
| ↓ prevalence of NAFLD | Cross-sectional study | 22,366 adult participants of US NHANES 8 and NHANES III surveys. Healthy adults (>20 years) with no history of excessive alcohol abuse, viral hepatitis or pregnancy. Age, gender, and ethnicity-adjusted analysis. Current and past users of Cannabis compared to non-users, independent of metabolic risk factors | Hepatic steatosis examination: Serum alanine aminotransferase (ALT) Liver ultrasonography | [97] |
| No difference in intrahepatic fat content and OGTT 3. ↑ % of abdominal visceral fat | Cross-sectional, case-control study | 60 patients of JH-BPRU 9 in Baltimore and CRC-NIH 10 in Bethesda Cannabis smokers (n = 30) and control subjects (n = 30) matched for age, sex, ethnicity, and BMI 11. | Abdominal and intrahepatic fat content were assesed by MRI and MRS 4 scan. Insulin-sensitivity was evaluated with OGTT. | [82] |
| ↓ prevalence of metabolic syndrome | Cross-sectional study | 8478 adult participants of NHANES survey. Current and past Cannabis users | Prevalence of metabolic syndrome examined by biochemical markers, BP 14 and waist circumference | [81] |
| ↓ fasting insulin levels ↓ prevalence of IR 1 ↓ waist circumference | Cross-sectional study | 4657 participants of NHANES survey. Current and past Cannabis users | Prevalence of insulin resistance by HOMA-IR 5 calculation. Assesment of waist circumference. | [78] |
| ↓ prevalence of obesity | Cross-sectional study | 52,375 participants of NESARC 12 and NCS-R 13 surveys of US nationally representative samples. Current and past Cannabis users | Prevalence of obesity examined by BMI calculation. | [80] |
| ↓ prevalence of NAFLD | Prospective cohort study | 838 HIV-HCV 15 coinfected patients, adjusted for BMI, hazardous alcohol consumption and current or lifetime use of lamivudine/zidovudine. Current and past Cannabis users | NAFLD examination using liver ultrasonography | [99] |
| ↓ prevalence of IR | Follow-up study | 703 HIV-HCV coinfected patients Current and past Cannabis users | Prevalence of insulin resistance by HOMA-IR calculation. | [79] |
| ↓ prevalence of NAFLD↓ overtime increment of steatosis score No difference in fibrosis score | Follow-up study | 390 patients with diagnosed non-affective psychosis. Current and past Cannabis users | NAFLD and NASH examination using clinical FLI 6 score. | [86] |
| ↓ prevalence of T2DM 2 in Cannabis users | Cross-sectional study | 10,896 adult participants of NHANES III survey. Current and past Cannabis users | Prevalence of T2DM defined based on self-report or abnormal glycaemic parameters | [76] |
| No increase in prevalence or progression of hepatic fibrosis | Meta-analysis | 5,973,595 participants of 9 studies including patients with NAFLD/HCV/HCV-HIV coinfection Current and past Cannabis users | Prevalence and progression of hepatic fibrosis among Cannabis users | [100] |
| Main Outcome | Research Model | Dose of CBD | Reference |
|---|---|---|---|
| Reduced TAG levels in HHL5 1 cells, independently of CB1R or TRPV1R action Reduced TAG content of larvae yolk sacs Reduced liver TAG 8 content in ob/ob mice | HHL5 cell culture Zebrafish 2 embryos and larvae Female ob/ob mice | 10 μM of CBD for 3 days 5 μM of CBD for 3 days 3 mg/kg for 4 weeks | [125] |
| CBD alleviated lipid accumulation and steatohepatitis induced by HFC 5 diet (↓ of serum ALT and TAG concentration, ↓ of hepatic TAG content in group treated with HFC+CBD versus HFC-only) | Male C57BL/6J mice fed with HFC diet and normal diet as control | 5 mg/kg of CBD for 8 weeks | [92] |
| CBD attenuated markers of I/R-induced hepatic injury: ALT 6, AST 7, proinflammatory chemokine, cytokine, and adhesion molecule expression, NF-κB 4 activation, cell necrosis and inflammatory cell infiltration. | Male C57BL/6J mice CB2 −/− mice exposed for hepatic I/R 3 | 3 and 10 mg/kg of CBD | [123] |
| Dose-dependent decrease in rats weight gain | Male Wistar rats | 2.5 and 5 mg/kg/day for 14 days | [124] |
| Main Outcome | Research Model | Participants and CBD Dose | Method | Reference |
|---|---|---|---|---|
| No significant changes in visceral adiposity or liver TAG assessed by MRI/MRS after treatment. | Randomized, double-blind, placebo-controlled, parallel group pilot study | 62 subjects with T2DM treated with 200 mg CBD daily or placebo for 13 weeks | MRI/MRS 1 scan to examine the liver fat content at the beginning and at end of treatment visit | [126] |
| Mean liver TAG levels did not significantly (p > 0.05) differ between the CBD and placebo groups. | Randomized, partially blind, placebo-controlled, dose-ranging phase 2 clinical study | 25 participants with NAFLD treated with 200/400/800 mg of CBD daily or placebo for 8 weeks | MRI/MRS 1 scan to examine the liver fat content at the baseline and at end of treatment visit | [125] |
| Mean % of liver fat did not change significantly (p > 0.05) after treatment. | Randomized, double blind, placebo controlled, parallel group, phase 2 clinical study | 13 participants with T2DM treated with 100 mg CBD daily or placebo for 13 weeks | MRI/MRS 1 scan to examine the liver fat content at the baseline and at end of treatment | [105] |
| Main Outcome | Research Model | Dose of THCV | Reference |
|---|---|---|---|
| Reduction of body mass, an increase of energy expenditure, improvement of glucose tolerance, and amelioration of insulin resistance. No significant effect on liver TAG content. Increase of energy expenditure, significant reduction of liver TAG content. | -DIO 1 mice -ob/ob mice | 12.5 mg/kg of THCV for 30 days 12.5 mg/kg of THCV for 30 days | [129] |
| Reduced TAG levels in HHL5 cells, independently of CB1R or TRPV1R action. Reduced TAG content of larvae yolk sacs | -HHL5 2 cell culture -Zebrafish 3 embryos and larvae | 10 μM of THCV for 3 days 10 μM for 3 days | [125] |
| Decrease in body weight, hypophagia | Male C57 BL6 mice | 3 mg/kg of THCV for 2 days | [131] |
| Main Outcome | Research Model | Participants | Method | Reference |
|---|---|---|---|---|
| Mean % of liver fat did not change significantly (p > 0.05) after treatment | Randomized, double blind, placebo controlled, parallel group, phase 2 clinical study | 13 participants with T2DM treated with 5 mg of THCV daily or placebo for 13 weeks | MRI/MRS 1 scan to examine the liver fat content at the baseline and at end-of-treatment | [105] |
| No significant changes in liver TAG concentration or visceral adiposity in any of the treatment groups. | Randomized, double-blind, placebo-controlled, parallel group pilot study | 62 subjects with T2DM treated with 5 mg of THCV daily or placebo for 13 weeks | MRI/MRS scan to examine the liver fat content at the beginning and at end-of-treatment visit | [126] |
| Increase of neural responding to rewarding and aversive stimuli | Double-blind clinical study | 20 healthy participants received 10 mg of THCV daily or placebo | fMRI 2 scan to examine neural response to rewarding and aversive food stimuli | [130] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berk, K.; Bzdega, W.; Konstantynowicz-Nowicka, K.; Charytoniuk, T.; Zywno, H.; Chabowski, A. Phytocannabinoids—A Green Approach toward Non-Alcoholic Fatty Liver Disease Treatment. J. Clin. Med. 2021, 10, 393. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm10030393
Berk K, Bzdega W, Konstantynowicz-Nowicka K, Charytoniuk T, Zywno H, Chabowski A. Phytocannabinoids—A Green Approach toward Non-Alcoholic Fatty Liver Disease Treatment. Journal of Clinical Medicine. 2021; 10(3):393. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm10030393
Chicago/Turabian StyleBerk, Klaudia, Wiktor Bzdega, Karolina Konstantynowicz-Nowicka, Tomasz Charytoniuk, Hubert Zywno, and Adrian Chabowski. 2021. "Phytocannabinoids—A Green Approach toward Non-Alcoholic Fatty Liver Disease Treatment" Journal of Clinical Medicine 10, no. 3: 393. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm10030393