
Circular RNAs Acting as miRNAs’ Sponges and Their Roles in Stem Cells

Abstract
:1. Introduction
2. Biogenesis and Characteristics of circRNAs
2.1. Biogenesis and Types of circRNAs
2.2. Cellular Localization of circRNAs
2.3. Abundance, Stability, Multiplicity and Specificity of circRNAs
3. Function of circRNAs
3.1. Transcriptional Regulation
3.1.1. miRNA Sponges
3.1.2. Regulation of Alternative Splicing
3.2. Translating to Peptide
3.3. Function as Protein Scaffolds, Decoys and Recruiters through Interaction with Proteins
4. Roles of circRNAs in Stem Cells
5. Opportunities and Challenges of circRNA-Based Therapeutic Technology
5.1. CircRNAs Are More Stable Than mRNAs Because of Their Circle Morphology
5.2. CircRNAs Are Tissue Specific and Have Less Side Effects Than mRNAs
5.3. Challenges of circRNA Technology
6. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, H.; Feng, C.; Jin, Y.; Tan, W.; Wei, F. Identificationand characterization of circularRNA involved in mechanical force induced periodontal stem cells. J. Cell. Physiol. 2019, 234, 10166–10177. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.B.; Kjems, J.; Chrstian, K.D. CircularRNA and miR-7 in cancer. Cancer Res. 2013, 73, 5609–5612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Xiao, Q.; Guan, C. The emerging landscape of circular RNA ciRS-7 in cancer. Oncol. Rep. 2015, 33, 2669–2674. [Google Scholar]
- Chen, L. The expanding regulatory mechanisms and cellular functions of circular RNAs. Nat. Rev. Mol. Cell Biol. 2020, 21, 475–490. [Google Scholar] [CrossRef]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef]
- Szabo, L.; Salzman, J. Detecting circular RNAs: Bioinformatic and experimental challenges. Nat. Rev. Genet. 2016, 17, 679–692. [Google Scholar] [CrossRef] [Green Version]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. CircRNA biogenesis competes with pre-mRNA splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Conn, S.J.; Pillman, K.A.; Toubia, J.; Conn, V.M.; Salmanidis, M.; Phillips, C.A.; Roslan, S.; Schreiber, A.W.; Gregory, P.A.; Goodall, G.J. The RNA binding protein quaking regulates formation of circRNAs. Cell 2015, 160, 1125–1134. [Google Scholar] [CrossRef] [Green Version]
- Xiao, M.; Ai, Y.; Wilusz, J.E. Biogenesis and Functions of Circular RNAs Come into Focus. Trends Cell Biol. 2020, 30, 226–240. [Google Scholar] [CrossRef]
- Li, P.; Chen, S.; Chen, H.; Mo, X.; Li, T.; Shao, Y.; Xiao, B.; Guo, J. Using circular RNA as a novel type of biomarker in the screening of gastric cancer. Clin. Chim. Acta 2015, 444, 132–136. [Google Scholar] [CrossRef]
- Qin, M.; Liu, G.; Huo, X.; Tao, X.; Sun, X.; Ge, Z.; Yang, J.; Fan, J.; Liu, L.; Qin, W. Hsa_circ_0001649: A circular RNA and potential novel biomarker for hepatocellular carcinoma. Cancer Biomark. 2016, 16, 161–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, B.; Tian, Z.; Fan, W.; Ni, B. Circular RNA: A novel biomarker and therapeutic target for human cancers. Int. J. Med. Sci. 2019, 16, 292–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, S.; Zhong, Y.; Shang, R.; Zhang, X.; Song, W.; Kjems, J.; Li, H. The emerging landscape of circular RNA in life processes. RNA Biol. 2017, 14, 992–999. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Guo, S.; Li, W.; Yu, P. The circular RNA Cdr1as, via miR-7 and its targets, regulates insulin transcription and secretion in islet cells. Sci. Rep. 2015, 5, 12453. [Google Scholar] [CrossRef] [PubMed]
- Lukiw, W.J. Circular RNA (circRNA) in Alzheimer’s disease (AD). Front. Genet. 2013, 4, 307. [Google Scholar] [CrossRef] [Green Version]
- Akhter, R. Circular RNA and Alzheimer’s Disease. Adv. Exp. Med. Biol. 2018, 1087, 239–243. [Google Scholar]
- Junn, E.; Lee, K.W.; Jeong, B.S.; Chan, T.W.; Im, J.Y.; Mouradian, M.M. Repression of alpha-synuclein expression and toxicity by microRNA-7. Proc. Natl. Acad. Sci. USA 2009, 106, 13052–13057. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, J.; Li, J.; Gui, R.; Nie, X.; Huang, R. CircRNA_014511 affects the radiosensitivity of bone marrow mesenchymal stem cells by binding to miR-29b-2-5p. Bosn. J. Basic Med. Sci. 2019, 19, 155–163. [Google Scholar] [CrossRef]
- Yan, N.; Xu, H.; Zhang, J.; Xu, L.; Zhang, Y.; Zhang, L.; Xu, Y.; Zhang, F. Circular RNA profile indicates circular RNA VRK1 is negatively related with breast cancer stem cells. Oncotarget 2017, 8, 95704–95718. [Google Scholar] [CrossRef] [Green Version]
- Cherubini, A.; Barilani, M.; Rossi, R.L.; Jalal, M.M.K.; Rusconi, F.; Buono, G.; Ragni, E.; Cantarella, G.; Simpson, H.; Péault, B.; et al. FOXP1 circular RNA sustains mesenchymal stem cell identity via microRNA inhibition. Nucleic Acids Res. 2019, 47, 5325–5340. [Google Scholar] [CrossRef] [Green Version]
- Javier Tadeo Granados-Riveron, G.A.-J. Does the linear Sry transcript function as a ceRNA for miR-138 The sense of antisense. F1000Research 2014, 3, 90. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Niu, H.; Li, M.; Zhang, H.; Yang, Z.; Tian, L.; Wu, Z.; Li, D.; Chen, X. Cyclic RNA hsa-circ-000595 regulates apoptosis of aortic smooth muscle cells. Mol. Med. Rep. 2015, 12, 6656–6662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Z.; Lv, M.; Chen, J. Screening differential circular RNA expression profiles reveals the regulatory role of circTCF25-miR-103a-3p/miR-107-CDK6 pathway in bladder carcinoma. Sci. Rep. 2016, 6, 30919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Shi, S.; Zhang, Q.; Dong, H.; Zhang, J. MicroRNA-206 upregulation relieves circTCF25-induced osteosarcoma cell proliferation and migration. J. Cell. Physiol. 2020. ahead of print. [Google Scholar]
- Li, H.; Lan, M.; Liao, X.; Tang, Z.; Yang, C. Circular RNA cir-ITCH Promotes Osteosarcoma Migration and Invasion through cir-ITCH/miR-7/EGFR Pathway. Technol. Cancer Res. Treat. 2020, 19, 1533033819898728. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Ma, K.; Sun, M.; Shi, S. Identification of the tumor-suppressive function of circular RNA ITCH in glioma cells through sponging miR-214 and promoting linear ITCH expression. Am. J. Transl. Res. 2018, 10, 1373–1386. [Google Scholar]
- Shen, F.; Liu, P.; Xu, Z.; Li, N.; Yi, Z.; Tie, X.; Zhang, Y.; Gao, L. CircRNA_001569 promotes cell proliferation through absorbing miR-145 in gastric cancer. J. Biochem. 2019, 165, 27–36. [Google Scholar] [CrossRef]
- Wu, L.; Xia, J.; Yang, J.; Shi, Y.; Xia, H.; Xiang, X.; Yu, X. Circ-ZNF609 promotes migration of colorectal cancer by inhibiting Gli1 expression via microRNA-150. J. BUON 2018, 23, 1343–1349. [Google Scholar]
- Wang, S.; Xue, X.; Wang, R.; Li, X.; Li, Q.; Wang, Y.; Xie, P.; Kang, Y.; Meng, R.; Feng, X. CircZNF609 promotes breast cancer cell growth, migration, and invasion by elevating p70S6K1 via sponging miR-145-5p. Cancer Manag. Res. 2018, 10, 3881–3890. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Liu, Y.; Yang, Y.; Mao, X.M.; Yin, Z.D. CircRNA ZNF609 promotes growth and metastasis of nasopharyngeal carcinoma by competing with microRNA-150-5p. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 2817–2826. [Google Scholar] [PubMed]
- Xiong, Y.; Zhang, J.; Song, C. CircRNA ZNF609 functions as a competitive endogenous RNA to regulate FOXP4 expression by sponging miR-138-5p in renal carcinoma. J. Cell. Physiol. 2019, 234, 10646–10654. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Wei, N.; Shao, G.; Jiang, C.; Zhang, S.; Wang, L. circZNF609 promotes the proliferation and migration of gastric cancer by sponging miR-483-3p and regulating CDK6. OncoTargets Ther. 2019, 12, 8197–8205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Pan, H.M.; Xin, L.; Zhang, Y.; Zhang, W.M.; Cao, P.; Xu, H.W. Circ-ZNF609 promotes carcinogenesis of gastric cancer cells by inhibiting miRNA-145-5p expression. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 9411–9417. [Google Scholar]
- Jin, C.; Zhao, W.; Zhang, Z.; Liu, W. Silencing circular RNA circZNF609 restrains growth, migration and invasion by up-regulating microRNA-186-5p in prostate cancer. Artif. Cells Nanomed. Biotechnol. 2019, 47, 3350–3358. [Google Scholar] [CrossRef] [Green Version]
- Tong, H.; Zhao, K.; Wang, J.; Xu, H.; Xiao, J. CircZNF609/miR-134-5p/BTG-2 axis regulates proliferation and migration of glioma cell. J. Pharm. Pharmacol. 2020, 72, 68–75. [Google Scholar] [CrossRef]
- Li, M.; Li, Y.; Yu, M. CircRNA ZNF609 Knockdown Suppresses Cell Growth via Modulating miR-188/ELF2 Axis in Nasopharyngeal Carcinoma. OncoTargets Ther. 2020, 13, 2399–2409. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Y.; Shen, W.; Wang, C.; Niu, N.; Pu, J. Circular RNA Circ-ZNF609 Promotes Lung Adenocarcinoma Proliferation by Modulating miR-1224-3p/ETV1 Signaling. Cancer Manag. Res. 2020, 12, 2471–2479. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Huang, H.; Jin, L.; Zhang, F.; Zeng, M.; Wei, L.; Tang, S.; Chen, D.; Wang, W. CircZNF609 enhances hepatocellular carcinoma cell proliferation, metastasis, and stemness by activating the Hedgehog pathway through the regulation of miR-15a-5p/15b-5p and GLI2 expressions. Cell Death Dis. 2020, 11, 358. [Google Scholar] [CrossRef]
- Liao, X.; Zhan, W.; Tian, B.; Luo, Y.; Gu, F.; Li, R. Circular RNA ZNF609 Promoted Hepatocellular Carcinoma Progression by Upregulating PAP2C Expression via Sponging miR-342-3p. OncoTargets Ther. 2020, 13, 7773–7783. [Google Scholar] [CrossRef]
- Du, S.; Zhang, P.; Ren, W.; Yang, F.; Du, C. Circ-ZNF609 Accelerates the Radioresistance of Prostate Cancer Cells by Promoting the Glycolytic Metabolism Through miR-501-3p/HK2 Axis. Cancer Manag. Res. 2020, 12, 7487–7499. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Fu, J.; Luan, J.; Qi, H.; Jiao, C.; Ran, M.; Wang, D.; Hao, X.; Zhang, Y.; Kopp, J.B.; et al. CircZNF609 is involved in the pathogenesis of focal segmental glomerulosclerosis by sponging miR-615-5p. Biochem. Biophys. Res. Commun. 2020, 531, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Ge, R.; Gao, G. Anti-antioxidant impacts of circZNF609 silence in HaCaT cells through regulating miR-145. Artif. Cells Nanomed. Biotechnol. 2020, 48, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Luo, Y.; Zhang, Y.; Wei, M.; Zhang, M.; Liu, H.; Su, Z. CircZNF609 aggravates neuropathic pain via miR-22-3p/ENO1 axis in CCI rat models. Gene 2020, 763, 145069. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Liu, F.; Wang, F.; Yang, X.; Guo, W. CircZNF609 promotes cell proliferation, migration, invasion, and glycolysis in nasopharyngeal carcinoma through regulating HRAS via miR-338-3p. Mol. Cell. Biochem. 2021, 476, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Yang, N.; Jiang, X.; Wang, J.; Dong, J.; Gao, Y. FUS-induced circular RNA ZNF609 promotes tumorigenesis and progression via sponging miR-142-3p in lung cancer. J. Cell. Physiol. 2021, 236, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Li, X.; Jia, X.; Geng, L. CircRNA ZNF609 Knockdown Represses the Development of Non-Small Cell Lung Cancer via miR-623/FOXM1 Axis. Cancer Manag. Res. 2021, 13, 1029–1039. [Google Scholar] [CrossRef]
- Du, S.; Li, H.; Lu, F.; Zhang, S.; Tang, J. Circular RNA ZNF609 promotes the malignant progression of glioma by regulating miR-1224-3p/PLK1 signaling. J. Cancer 2021, 12, 3354–3366. [Google Scholar] [CrossRef]
- Wang, J.; Lin, Y.; Jiang, D.H.; Yang, X.; He, X.G. CircRNA ZNF609 promotes angiogenesis in nasopharyngeal carcinoma by regulating miR-145/STMN1 axis. Kaohsiung J. Med. Sci. 2021, 37, 686–698. [Google Scholar] [CrossRef]
- Liu, Q.; Cui, W.; Yang, C.; Du, L.P. Circular RNA ZNF609 drives tumor progression by regulating the miR-138-5p/SIRT7 axis in melanoma. Aging 2021, 13, 19822–19834. [Google Scholar] [CrossRef]
- Gu, Q.; Hou, W.; Shi, L.; Liu, H.; Zhu, Z.; Ye, W. Circular RNA ZNF609 functions as a competing endogenous RNA in regulating E2F transcription factor 6 through competitively binding to microRNA-197-3p to promote the progression of cervical cancer progression. Bioengineered 2021, 12, 927–936. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, L.S.; Okholm, T.L.H.; Venø, M.T.; Kjems, J. Circular RNAs are abundantly expressed and upregulated during human epidermal stem cell differentiation. RNA Biol. 2018, 15, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Zhao, Y.; Chen, Q.; Zhu, S.; Niu, Y.; Ye, Z.; Hu, P.; Chen, D.; Xu, P.; Chen, J.; et al. Hypoxic exosomal HIF-1α-stabilizing circZNF91 promotes chemoresistance of normoxic pancreatic cancer cells via enhancing glycolysis. Oncogene 2021, 40, 5505–5517. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Quanbo, Z.; Su, D.; Luo, Y.; Fu, Z.; Huang, L.; Li, Z.; Jiang, D.; Kong, Y.; Chen, R.; et al. Circular RNA circBFAR promotes the progression of pancreatic ductal adenocarcinoma via the miR-34b-5p/MET/Akt axis. Mol. Cancer 2020, 19, 83. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, B.; Zhao, R.; Li, H.; Wei, B.; Dai, G. Knockdown of circBFAR inhibits proliferation and glycolysis in gastric cancer by sponging miR-513a-3p/hexokinase 2 axis. Biochem. Biophys. Res. Commun. 2021, 560, 80–86. [Google Scholar] [CrossRef]
- Szabo, L.; Morey, R.; Palpant, N.J.; Wang, P.L.; Afari, N.; Jiang, C.; Parast, M.M.; Murry, C.E.; Laurent, L.C.; Salzman, J. Statistically based splicing detection reveals neural enrichment and tissue-specific induction of circular RNA during human fetal development. Genome Biol. 2015, 16, 126. [Google Scholar] [CrossRef] [Green Version]
- Starke, S.; Jost, I.; Rossbach, O.; Schneider, T.; Schreiner, S.; Hung, L.H.; Bindereif, A. Exon circularization requires canonical splice signals. Cell Rep. 2015, 10, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Ragan, C.; Goodall, G.J.; Shirokikh, N.E. Insights into the biogenesis and potential functions of exonic circular RNA. Sci. Rep. 2019, 9, 2048. [Google Scholar] [CrossRef]
- Salzman, J.; Gawad, C.; Wang, P.L.; Lacayo, N.; Brown, P.O. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS ONE 2012, 7, e30733. [Google Scholar] [CrossRef] [Green Version]
- Liang, D.; Wilusz, J.E. Short intronic repeat sequences facilitate circular RNA production. Genes Dev. 2014, 28, 2233–2247. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.O.; Wang, H.B.; Zhang, Y.; Lu, X.; Chen, L.L.; Yang, L. Complementary sequence-mediated exon circularization. Cell 2014, 159, 134–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhdanov, D.D.; Pokrovsky, V.S.; Orlova, E.V.; Orlova, V.S.; Pokrovskaya, M.V.; Aleksandrova, S.S.; Sokolov, N.N. Intracellular Localization of Apoptotic Endonuclease EndoG and Splice-Variants of Telomerase Catalytic Subunit hTERT. Biochemistry 2017, 82, 894–905. [Google Scholar] [CrossRef] [PubMed]
- Zhdanov, D.D.; Vasina, D.A.; Orlova, V.S.; Gotovtseva, V.Y.; Bibikova, M.V.; Pokrovsky, V.S.; Pokrovskaya, M.V.; Aleksandrova, S.S.; Sokolov, N.N. Apoptotic endonuclease EndoG induces alternative splicing of telomerase catalytic subunit hTERT and death of tumor cells. Biomed. Khim. 2016, 62, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Plyasova, A.A.; Zhdanov, D.D. Alternative Splicing of Human Telomerase Reverse Transcriptase (hTERT) and Its Implications in Physiological and Pathological Processes. Biomedicines 2021, 9, 256. [Google Scholar] [CrossRef]
- Zhdanov, D.D.; Vasina, D.A.; Grachev, V.A.; Orlova, E.V.; Orlova, V.S.; Pokrovskaya, M.V.; Alexandrova, S.S.; Sokolov, N.N. Alternative splicing of telomerase catalytic subunit hTERT generated by apoptotic endonuclease EndoG induces human CD4(+) T cell death. Eur. J. Cell Biol. 2017, 96, 653–664. [Google Scholar] [CrossRef]
- Li, X.; Yang, L.; Chen, L.L. The Biogenesis, Functions, and Challenges of Circular RNAs. Mol. Cell 2018, 71, 428–442. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Xie, X.; Zhou, J.; Sheng, M.; Yin, X.; Ko, E.-A.; Zhou, T.; Gu, W. Quantifying circular RNA expression from RNA-seq data using model-based framework. Oxf. Univ. Press 2017, 33, 2131–2139. [Google Scholar] [CrossRef]
- Huang, C.; Liang, D.; Tatomer, D.C.; Wilusz, J.E. A length-dependent evolutionarily conserved pathway controls nuclear export of circular RNAs. Genes Dev. 2018, 32, 639–644. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.O.; Chen, T.; Xiang, J.F.; Yin, Q.F.; Xing, Y.H.; Zhu, S.; Yang, L.; Chen, L.L. Circular intronic long noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Rybak-Wolf, A.; Stottmeister, C.; Glažar, P.; Jens, M.; Pino, N.; Giusti, S.; Hanan, M.; Behm, M.; Bartok, O.; Ashwal-Fluss, R.; et al. Circular RNAs in the Mammalian Brain Are Highly Abundant, Conserved, and Dynamically Expressed. Mol. Cell 2015, 58, 870–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, S.; Zhou, H.; Feng, Z.; Xu, Z.; Tang, Y.; Li, P.; Wu, M. CircRNA: Functions and properties of a novel potential biomarker for cancer. Mol. Cancer 2017, 16, 94. [Google Scholar] [CrossRef] [PubMed]
- Panda, A.C. Circular RNAs Act as miRNA Sponges. Adv. Exp. Med. Biol. 2018, 1087, 67–79. [Google Scholar]
- Lu, Q.; Liu, T.; Feng, H.; Yang, R.; Zhao, X.; Chen, W.; Jiang, B.; Qin, H.; Guo, X.; Liu, M.; et al. Circular RNA circSLC8A1 acts as a sponge of miR-130b/miR-494 in suppressing bladder cancer progression via regulating PTEN. Mol. Cancer 2019, 18, 111. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wu, H.; Wang, Y.; Zhu, S.; Liu, J.; Fang, X.; Chen, H. Circular RNA of cattle casein genes are highly expressed in bovine mammary gland. J. Dairy Sci. 2016, 99, 4750–4760. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Li, Y.; Ge, P.; Zhang, K.; Liu, H.; Jiang, N. QKI-Regulated Alternative Splicing Events in Cervical Cancer: Pivotal Mechanism and Potential Therapeutic Strategy. DNA Cell Biol. 2021, 40, 1261–1277. [Google Scholar] [CrossRef]
- Song, J.; Wang, H.L.; Song, K.H.; Ding, Z.W.; Wang, H.L.; Ma, X.S.; Lu, F.Z.; Xia, X.L.; Wang, Y.W.; Zou, F.; et al. CircularRNA_104670 plays a critical role in intervertebral disc degeneration by functioning as a ceRNA. Exp. Mol. Med. 2018, 50, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Qu, S.; Yang, X.; Li, X.; Wang, J.; Gao, Y.; Shang, R.; Sun, W.; Dou, K.; Li, H. Circular RNA. A new star of noncoding RNAs. Cancer Lett. 2015, 365, 141–148. [Google Scholar] [CrossRef]
- Legnini, I.; Di Timoteo, G.; Rossi, F.; Morlando, M.; Briganti, F.; Sthandier, O.; Fatica, A.; Santini, T.; Andronache, A.; Wade, M.; et al. Circ-ZNF609 Is a Circular RNA that Can Be Translated and Functions in Myogenesis. Mol. Cell 2017, 66, 22–37.e9. [Google Scholar] [CrossRef] [Green Version]
- Pamudurti, N.R.; Bartok, O.; Jens, M.; Ashwal-Fluss, R.; Stottmeister, C.; Ruhe, L.; Hanan, M.; Wyler, E.; Perez-Hernandez, D.; Ramberger, E.; et al. Translation of CircRNAs. Mol. Cell 2017, 66, 9–21.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Chen, L.; Zhou, Y.; Wang, Q.; Zheng, Z.; Xu, B.; Wu, C.; Zhou, Q.; Hu, W.; Jiang, J. A novel protein encoded by a circular RNA circPPP1R12A promotes tumor pathogenesis and metastasis of colon cancer via Hippo-YAP signaling. Mol. Cancer 2019, 18, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Ma, M.; Yang, X.; Zhang, M.; Luo, J.; Zhou, H.; Huang, N.; Xiao, F.; Lai, B.; Lv, W.; et al. Circular HER2 RNA positive triple negative breast cancer is sensitive to Pertuzumab. Mol. Cancer 2020, 19, 142. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Xu, Y.; Zhang, X.; Deng, S.; Yuan, Y.; Luo, X.; Hossain, M.T.; Zhu, X.; Du, K.; Hu, F.; et al. A novel protein AXIN1-295aa encoded by circAXIN1 activates the Wnt/β-catenin signaling pathway to promote gastric cancer progression. Mol. Cancer 2021, 20, 158. [Google Scholar] [CrossRef] [PubMed]
- Ye, F.; Gao, G.; Zou, Y.; Zheng, S.; Zhang, L.; Ou, X.; Xie, X.; Tang, H. circFBXW7 Inhibits Malignant Progression by Sponging miR-197-3p and Encoding a 185-aa Protein in Triple-Negative Breast Cancer. Mol. Ther. Nucleic Acids 2019, 18, 88–98. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Gao, X.; Zhang, M.; Yan, S.; Sun, C.; Xiao, F.; Huang, N.; Yang, X.; Zhao, K.; Zhou, H.; et al. Novel Role of FBXW7 Circular RNA in Repressing Glioma Tumorigenesis. J. Natl. Cancer Inst. 2018, 110, 304–315. [Google Scholar] [CrossRef] [Green Version]
- Pan, Z.; Cai, J.; Lin, J.; Zhou, H.; Peng, J.; Liang, J.; Xia, L.; Yin, Q.; Zou, B.; Zheng, J.; et al. A novel protein encoded by circFNDC3B inhibits tumor progression and EMT through regulating Snail in colon cancer. Mol. Cancer 2020, 19, 71. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.Y.; Cai, Z.R.; Liu, J.; Wang, D.S.; Ju, H.Q.; Xu, R.H. Circular RNA: Metabolism, functions and interactions with proteins. Mol. Cancer 2020, 19, 172. [Google Scholar] [CrossRef]
- Du, W.W.; Fang, L.; Yang, W.; Wu, N.; Awan, F.M.; Yang, Z.; Yang, B.B. Induction of tumor apoptosis through a circular RNA enhancing Foxo3 activity. Cell Death Differ. 2017, 24, 357–370. [Google Scholar] [CrossRef]
- Du, W.W.; Yang, W.; Liu, E.; Yang, Z.; Dhaliwal, P.; Yang, B.B. Foxo3 circular RNA retards cell cycle progression via forming ternary complexes with p21 and CDK2. Nucleic Acids Res. 2016, 44, 2846–2858. [Google Scholar] [CrossRef] [Green Version]
- Du, W.W.; Yang, W.; Chen, Y.; Wu, Z.K.; Foster, F.S.; Yang, Z.; Li, X.; Yang, B.B. Foxo3 circular RNA promotes cardiac senescence by modulating multiple factors associated with stress and senescence responses. Eur. Heart J. 2017, 38, 1402–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, W.W.; Yang, W.; Li, X.; Awan, F.M.; Yang, Z.; Fang, L.; Lyu, J.; Li, F.; Peng, C.; Krylov, S.N.; et al. A circular RNA circ-DNMT1 enhances breast cancer progression by activating autophagy. Oncogene 2018, 37, 5829–5842. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Feng, J.; Lei, L.; Hu, J.; Xia, L.; Wang, J.; Xiang, Y.; Liu, L.; Zhong, S.; Han, L.; et al. Comprehensive characterization of tissue-specific circular RNAs in the human and mouse genomes. Brief. Bioinform. 2017, 18, 984–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misir, S.; Wu, N.; Yang, B.B. Specific expression and functions of circular RNAs. Cell Death Differ. 2022, 29, 481–491. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, Z.; Yu, M.; Yang, G. Roles of circular RNAs in regulating the self-renewal and differentiation of adult stem cells. Differentiation 2020, 113, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Li, X.; Huang, Y.; Jia, L.; Li, W. The Circular RNA landscape of periodontal ligament stem cells during osteogenesis. J. Periodontol. 2017, 88, 906–914. [Google Scholar] [CrossRef]
- Sun, B.; Shi, L.; Shi, Q.; Jiang, Y.; Su, Z.; Yang, X.; Zhang, Y. Circular RNAs are abundantly expressed and upregulated during repair of the damaged endometrium by Wharton’s jelly-derived mesenchymal stem cells. Stem Cell Res. Ther. 2018, 9, 314. [Google Scholar] [CrossRef]
- Sun, X.; Jin, Y.; Liang, Q.; Tang, J.; Chen, J.; Yu, Q.; Li, F.; Li, Y.; Wu, J.; Wu, S. Altered expression of circular RNAs in human placental chorionic plate-derived mesenchymal stem cells pretreated with hypoxia. J. Clin. Lab. Anal. 2019, 33, e22825. [Google Scholar] [CrossRef] [Green Version]
- Lei, W.; Feng, T.; Fang, X.; Yu, Y.; Yang, J.; Zhao, Z.A.; Liu, J.; Shen, Z.; Deng, W.; Hu, S. Signature of circular RNAs in human induced pluripotent stem cells and derived cardiomyocytes. Stem Cell Res. Ther. 2018, 9, 56. [Google Scholar] [CrossRef]
- Chen, M.; Yang, Y.; Zeng, J.; Deng, Z.; Wu, B. circRNA Expression Profile in Dental Pulp Stem Cells during Odontogenic Differentiation. Stem Cells Int. 2020, 2020, 5405931. [Google Scholar] [CrossRef]
- Nicolet, B.P.; Engels, S.; Aglialoro, F.; van den Akker, E.; von Lindern, M.; Wolkers, M.C. Circular RNA expression in human hematopoietic cells is widespread and cell-type specific. Nucleic Acids Res. 2018, 46, 8168–8180. [Google Scholar] [CrossRef]
- Yu, C.Y.; Li, T.C.; Wu, Y.Y.; Yeh, C.H.; Chiang, W.; Chuang, C.Y.; Kuo, H.C. The circular RNA circBIRC6 participates in the molecular circuitry controlling human pluripotency. Nat. Commun. 2017, 8, 1149. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Wang, Y.; Dai, X.; Zhou, T.; Chen, J.; Tao, B.; Zhang, J.; Cao, F. The Tumor-Suppressive Human Circular RNA CircITCH Sponges miR-330-5p to Ameliorate Doxorubicin-Induc.ced Cardiotoxicity Through Upregulating SIRT6, Survivin, and SERCA2a. Circ. Res. 2020, 127, e108–e125. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Zhu, S.; Chen, J.; Wang, J.; Rong, Q.; Chen, S. Hsa_circRNA_33287 promotes the osteogenic differentiation of maxillary sinus membrane stem cells via miR-214-3p/Runx3. Biomed. Pharmacother. Biomed. Pharmacother. 2019, 109, 1709–1717. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Xiang, G.; Yu, D.; Yang, G.; He, W.; Yang, S.; Zhou, G.; Liu, A. Hsa_circ_0002468 Regulates the Neuronal Differentiation of SH-SY5Y Cells by Modulating the MiR-561/E2F8 Axis. Med. Sci. Monit. 2019, 25, 2511–2519. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Huan, Y.; Li, X.; Li, J.; Lv, G. RUNX3 derived hsa_circ_0005752 accelerates the osteogenic differentiation of adipose- derived stem cells via the miR-496/MDM2-p53 pathway. Regen Ther. 2021, 18, 430–440. [Google Scholar] [CrossRef]
- Tao, S.Y.; Liu, J.; Shi, D.; Peng, M.; Li, C.; Wu, S. Cancer stem cell-specific expression profiles reveal emerging bladder cancer biomarkers and identify circRNA_1038.809 as an important regulator in bladder cancer. Aging 2020, 12, 17. [Google Scholar] [CrossRef]
- Ouyang, Z.; Tan, T.; Zhang, X.; Wan, J.; Zhou, Y.; Jiang, G.; Yang, D.; Guo, X.; Liu, T. CircRNA hsa_circ_0074834 promotes the osteogenesis-angiogenesis coupling process in bone mesenchymal stem cells (BMSCs) by acting as a ceRNA for miR-942-5p. Cell Death Dis. 2019, 10, 932. [Google Scholar] [CrossRef]
- Guo, Z.Y.; Zhao, L.Y.; Ji, S.H.; Long, T.; Huang, Y.L.; Ju, R.; Tang, W.; Tian, W.D.; Long, J. CircRNA-23525 regulates osteogenic diferentiation of adipose-derived mesenchymal stem cells via miR-30a-3p. Cell Tissue Res. 2021, 383, 13. [Google Scholar] [CrossRef]
- Han, W.; Tao, X.; Weng, T.; Chen, L. Circular RNA PVT1 inhibits tendon stem/progenitor cell senescence by sponging microRNA-199a-5p. Toxicol. Vitro 2021, 79, 105297. [Google Scholar] [CrossRef]
- Chia, W.; Liu, J.; Huang, Y.G.; Zhang, C. A circular RNA derived from DAB1 promotes cell proliferation and osteogenic differentiation of BMSCs via RBPJ/DAB1 axis. Cell Death Dis. 2020, 11, 372. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.B.; Li, P.B.; Guo, S.F.; Yang, Q.S.; Chen, Z.X.; Wang, D.; Shi, S.B. circRNA_0006393 promotes osteogenesis in glucocorticoid-induced osteoporosis by sponging miR-145-5p and upregulating FOXO1. Mol. Med. Rep. 2019, 20, 2851–2858. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Jia, L.; Zheng, Y. circRNA Expression Profiles in Human Bone Marrow Stem Cells Undergoing Osteoblast Differentiation. Stem Cell Rev. Rep. 2019, 15, 126–138. [Google Scholar] [CrossRef]
- He, W.; Shi, X.; Guo, Z.; Wang, H.; Kang, M.; Lv, Z. Circ_0019693 promotes osteogenic differentiation of bone marrow mesenchymal stem cell and enhances osteogenesis-coupled angiogenesis via regulating microRNA-942-5p-targeted purkinje cell protein 4 in the development of osteoporosis. Bioengineered 2022, 13, 2181–2193. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Xie, H.; Gui, R.; Nie, X.; Shan, D.; Huang, R.; Deng, H.; Zhang, J. CircRNA-011235 Counteracts The Deleterious Effect of Irradiation Treatment on Bone Mesenchymal Stem Cells by Regulating the miR-741-3p/CDK6 Pathway. Cell J. 2022, 24, 15–21. [Google Scholar] [PubMed]
- Lu, Y.; Liu, Y.K.; Wan, F.Y.; Shi, S.; Tao, R. CircSmg5 stimulates the osteogenic differentiation of bone marrow mesenchymal stem cells by targeting the miR-194-5p/Fzd6 axis and beta-catenin signaling. Environ. Toxicol. 2022, 37, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Gu, D.; Xu, B.; Yang, C.; Wang, L. Circular RNA circ_0005835 promotes promoted neural stem cells proliferation and differentiate to neuron and inhibits inflammatory cytokines levels through miR-576-3p in Alzheimer’s disease. Environ. Sci. Pollut. Res. Int. 2022, 29, 35934–35943. [Google Scholar] [CrossRef]
- Tian, T.; Li, F.; Chen, R.; Wang, Z.; Su, X.; Yang, C. Therapeutic Potential of Exosomes Derived From circRNA_0002113 Lacking Mesenchymal Stem Cells in Myocardial Infarction. Front. Cell Dev. Biol. 2021, 9, 779524. [Google Scholar] [CrossRef]
- Ye, Y.; Ke, Y.; Liu, L.; Xiao, T.; Yu, J. CircRNA FAT1 Regulates Osteoblastic Differentiation of Periodontal Ligament Stem Cells via miR-4781-3p/SMAD5 Pathway. Stem Cells Int. 2021, 2021, 5177488. [Google Scholar] [CrossRef]
- Qi, W.; Chen, J.; Cheng, X.; Huang, J.; Xiang, T.; Li, Q.; Long, H.; Zhu, B. Targeting the Wnt-Regulatory Protein CTNNBIP1 by microRNA-214 Enhances the Stemness and Self-Renewal of Cancer Stem-Like Cells in Lung Adenocarcinomas. Stem Cells 2015, 33, 3423–3436. [Google Scholar] [CrossRef]
- Zhang, X.L.; Xu, L.L.; Wang, F. Hsa_circ_0020397 regulates colorectal cancer cell viability, apoptosis and invasion by promoting the expression of the miR-138 targets TERT and PD-L1. Cell Biol. Int. 2017, 41, 1056–1064. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; Li, G.; Liu, H.; Li, T.; Liu, J.; Zhao, Q.; Wang, C. Comprehensive Circular RNA Profiling Reveals That hsa_circ_0005075, a New Circular RNA Biomarker, Is Involved in Hepatocellular Crcinoma Development. Medicine 2016, 95, e3811. [Google Scholar] [CrossRef] [PubMed]
- Xiao, T.; Xue, J.; Shi, M.; Chen, C.; Luo, F.; Xu, H.; Chen, X.; Sun, B.; Sun, Q.; Yang, Q.; et al. Circ008913, via miR-889 regulation of DAB2IP/ZEB1, is involved in the arsenite-induced acquisition of CSC-like properties by human keratinocytes in carcinogenesis. Metallomics 2018, 10, 1328–1338. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhang, Y.; Niu, M.; Shi, Y.; Liu, H.; Yang, D.; Li, F.; Lu, Y.; Bo, Y.; Zhang, R.; et al. Whole-Transcriptome Analysis of CD133+CD144+ Cancer Stem Cells Derived from Human Laryngeal Squamous Cell Carcinoma Cells. Cell. Physiol. Biochem. 2018, 47, 1696–1710. [Google Scholar] [CrossRef]
- Chen, X.; Chen, R.X.; Wei, W.S.; Li, Y.H.; Feng, Z.H.; Tan, L.; Chen, J.W.; Yuan, G.J.; Chen, S.L.; Guo, S.J.; et al. PRMT5 Circular RNA Promotes Metastasis of Urothelial Carcinoma of the Bladder through Sponging miR-30c to Induce Epithelial-Mesenchymal Transition. Clin. Cancer Res. 2018, 24, 6319–6330. [Google Scholar] [CrossRef] [Green Version]
- Jakobi, T.; Siede, D.; Eschenbach, J.; Heumüller, A.W.; Busch, M.; Nietsch, R.; Meder, B.; Most, P.; Dimmeler, S.; Backs JKatus, H.A.; et al. Deep Characterization of Circular RNAs from Human Cardiovascular Cell Models and Cardiac Tissue. Cells 2020, 9, 1616. [Google Scholar] [CrossRef]
- Li, X.; Zheng, Y.; Huang, Y.; Zhang, Y.; Jia, L.; Li, W. Circular RNA CDR1as regulates osteoblastic differentiation of periodontal ligament stem cells via the miR-7/GDF5/SMAD and p38 MAPK signaling pathway. Stem Cell Res. Ther. 2018, 9, 232. [Google Scholar] [CrossRef]
- Shen, W.; Sun, B.; Zhou, C.; Ming, W.; Zhang, S.; Wu, X. CircFOXP1/FOXP1 promotes osteogenic differentiation in adipose-derived mesenchymal stem cells and bone regeneration in osteoporosis via miR-33a-5p. J. Cell Mol. Med. 2020, 24, 12513–12524. [Google Scholar] [CrossRef]
- Zhang, J.; Luo, C.J.; Xiong, X.Q.; Li, J.; Tang, S.H.; Sun, L.; Su, Q. MiR-21-5p-expressing bone marrow mesenchymal stem cells alleviate myocardial ischemia/reperfusion injury by regulating the circRNA_0031672/miR-21-5p/programmed cell death protein 4 pathway. J. Geriatr. Cardiol. 2021, 18, 1029–1043. [Google Scholar]
- Ma, L.; Wei, J.; Zeng, Y.; Liu, J.; Xiao, E.; Kang, Y. Mesenchymal stem cell-originated exosomal circDIDO1 suppresses hepatic stellate cell activation by miR-141-3p/PTEN/AKT pathway in human liver fibrosis. Drug Deliv. 2022, 29, 440–453. [Google Scholar] [CrossRef]
- Shen, W.; Zhao, X.; Li, S. Exosomes Derived from ADSCs Attenuate Sepsis-Induced Lung Injury by Delivery of Circ-Fryl and Regulation of the miR-490-3p/SIRT3 Pathway. Inflammation 2022, 45, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Zhou, N.; Wang, Z.; Li, G.; Kou, Y.; Yu, S.; Feng, Y.; Chen, L.; Yang, J.; Tian, F. circGprc5a Promoted Bladder Oncogenesis and Metastasis through Gprc5a-Targeting Peptide. Mol. Ther. Nucleic Acids 2018, 13, 633–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Liu, X.; Xu, L.; Wang, Y.; Wang, S.; Li, Q.; Huang, Y.; Liu, T. Long non-coding RNA BACE1-AS is a novel target for anisomycin-mediated suppression of ovarian cancer stem cell proliferation and invasion. Oncol. Rep. 2016, 35, 1916–1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Z.; Chen, T.; Yao, Q.; Zheng, L.; Zhang, Z.; Wang, J.; Hu, Z.; Cui, H.; Han, Y.; Han, X.; et al. The circular RNA ciRS-7 promotes APP and BACE1 degradation in an NF-κB-dependent manner. FEBS J. 2017, 284, 1096–1109. [Google Scholar] [CrossRef] [PubMed]
- Zhan, W.; Liao, X.; Wang, Y.; Li, L.; Li, J.; Chen, Z.; Tian, T.; He, J. circCTIC1 promotes the self-renewal of colon TICs through BPTF-dependent c-Myc expression. Carcinogenesis 2019, 40, 560–568. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.J.; Zheng, B.; Luo, G.J.; Ma, X.K.; Lu, X.Y.; Lin, X.M.; Yang, S.; Zhao, Q.; Wu, T.; Li, Z.X.; et al. Circular RNAs negatively regulate cancer stem cells by physically binding FMRP against CCAR1 complex in hepatocellular carcinoma. Theranostics 2019, 9, 3526–3540. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.J.; Li, J.; Yang, G.; Yi, C.J.; Zhang, D.; Yu, F.; Ma, Z. Circular RNAs in stem cells: From basic research to clinical implications. Biosci. Rep. 2022, 42, BSR20212510. [Google Scholar] [CrossRef]
- Qu, L.; Yi, Z.; Shen, Y.; Lin, L.; Chen, F.; Xu, Y.; Wu, Z.; Tang, H.; Zhang, X.; Tian, F.; et al. Circular RNA vaccines against SARS-CoV-2 and emerging variants. Cell 2022, in press. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Name | Organism | miRNAs It Binds | Function | References |
|---|---|---|---|---|
| ciRs-7/CDR1as | Homo sapien; Mus musculus | miR-7 | Prevent downregulation of target gene | [2,3] |
| circSry | Homo sapien; Mus musculus | miR-138 | Involve in cancer pathogenesis; Regulate hypoxia-induced apoptosis in cardiac myocytes | [21,22] |
| Hsa-circ-000595 | Homo sapiens | miR-19a | Decreased apoptosis in human aortic smooth muscle cell | [23] |
| circTCF25 | Homo sapiens | miR-107; miR103a-3p; miR-206 | Upregulation of CDK6 in urinary bladder carcinoma tissue Osteosarcoma cell proliferation and migration | [24,25] |
| circ-ITCH | Homo sapiens | miR-7; miR-214 | Osteosarcoma Migration and Invasion promoting linear ITCH expression | [26,27] |
| circRNA_001569 | Homo sapiens | miR-145 | Cell proliferation regulation | [28] |
| circZNF609 | Homo sapiens | miR-150; miR-145-5p; miR-150-5p; miR-138-5p; miR-483-3p; miR-186-5p; miR-134-5p; miR-188; miR-1224-3p; miR-342-3p; miR-501-3p; miR-615-5p; miR-22-3p; miR-338-3p; miR-142-3p; miR-623; miR-1224-3p; miR-197-3p | Cancer cell growth, migration and invasion | [29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51] |
| circZNF91 | Homo sapiens | miR-23b-3p; | Chemoresistance of normoxic pancreatic cancer cells by enhancing glycolysis; Epidermal stem cell differentiation | [52,53] |
| circBFAR | Homo sapiens | miR-34b-5p; miR-513a-3p | Progression of pancreatic ductal adenocarcinoma Proliferation and glycolysis in gastric cancer | [54,55] |
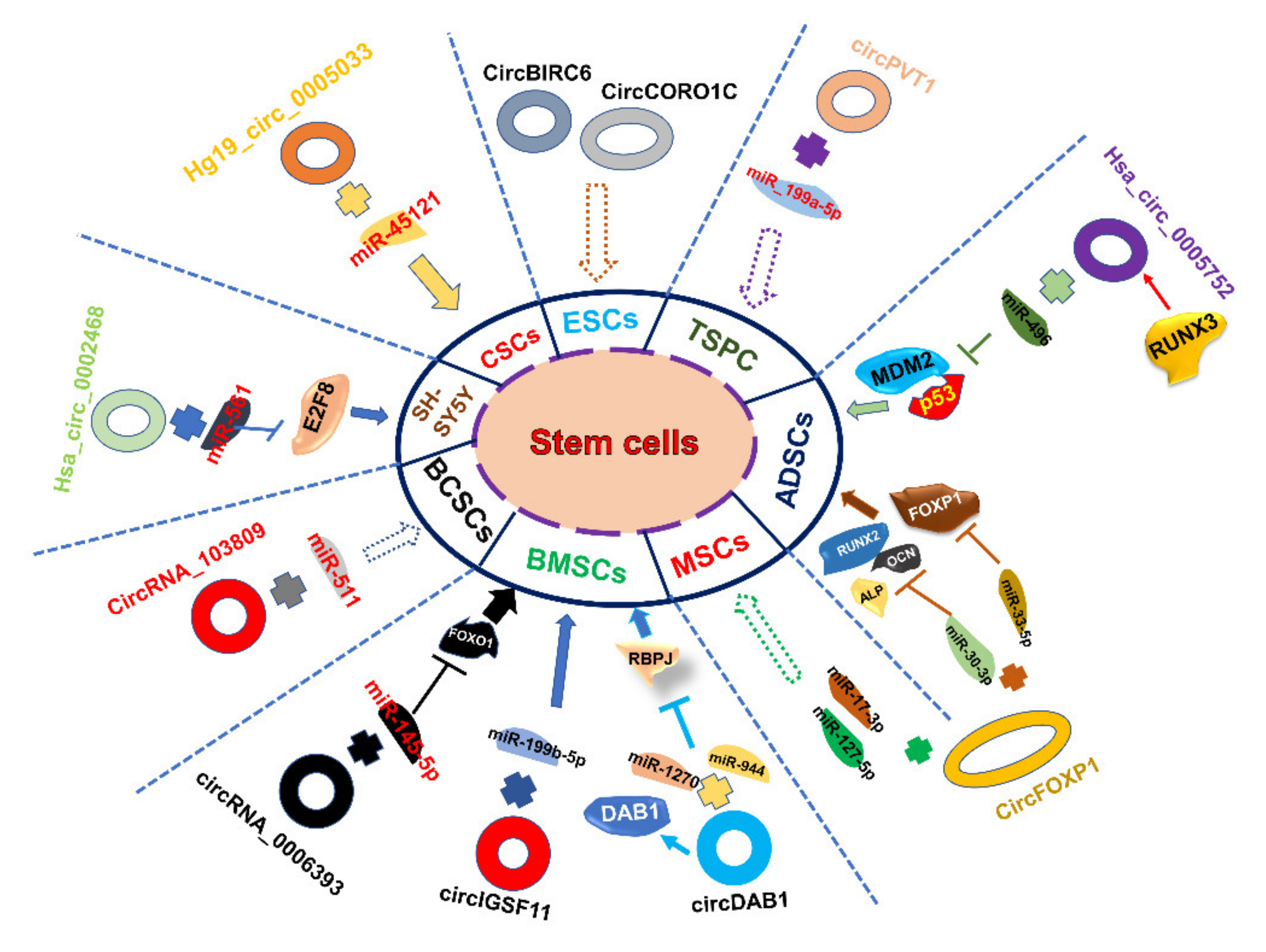
| Stem Cell Types | CircRNA | Role in Stem Cells | miRNAs It Binds | References |
|---|---|---|---|---|
| Pluripotent stem cells | circBIRC6; circCORO1C | Regulates the differentiation and pluripotency of hESCs | miR-34a, miR-145 | [102] |
| circSLC8A1 | Overexpression of circSLC8A1 is related to heart disease from the study of hiPS cell-derived cardiomyocytes | miR-133a | [99] | |
| CircITCH | Downregulated CircITCH in doxorubicin-treated hiPS cell-derived cardiomyocytes can alleviate DOX-induced cardiomyocyte injury | miR-330-5p | [103] | |
| Somatic stem cells | circRNA-33287 | Promotes osteogenic differentiation of maxillary sinus membrane stem cells | miR-214-3p | [104] |
| hsa_circ_0002468 | Regulates SH-SY5Y differentiation and proliferation | miR-561 | [105] | |
| hsa_circ_0005752 | Derived from RUNX3, promotes the osteogenic differentiation of adipose-derived stem cells through release the limit of MDM2-p53 | miR-496 | [106] | |
| circFOXP1 | Accelerates differentiation and proliferation of MSCs | miR-17–3p and miR-127–5p | [20] | |
| circRNA_103809 | The highest expressed circRNAs identified in BCSCs, promotes the self-renewal, migration and invasion capabilities of bladder cancer | miR-511 | [107] | |
| hsa_circ_0074834 | As a ceRNA in bone mesenchymal stem cells (BMSCs) | miR-942-5p | [108] | |
| circRNA-23525 | Regulates osteogenic differentiation of adipose-derived mesenchymal stem cells | miR-30a-3p | [109] | |
| circPVT1 | Inhibits tendon stem/progenitor cell (TSPC) senescence | miR-199a-5p | [110] | |
| circDAB1 | Derived from DAB1 gene to upregulate RBPJ through sponging miRNAs, and upregulates the host gene DAB1 in BMSCs | miR-1270 and miR-944 | [111] | |
| circ—0006393 | Promotes osteogenesis in human BMSCs by inducing the FOXO1 gene | miR-145-5p | [112] | |
| circIGSF11 | Inhibits the osteogenic differentiation of hBMSCs | miR-199b-5p | [113] | |
| circ-0019693 | Promotes osteogenic differentiation of BMSCs by inducing PCP4 gene | miR-942-5p | [114] | |
| circ-011235 | Counteracts the harm of irradiation treatment on BMSCs through the miR-741-3p/CDK6 pathway | miR-741-3p | [115] | |
| circSmg5 | Accelerates the osteogenic differentiation of BMSCs through miR-194-5p/Fzd6 and beta-catenin signaling | miR-194-5p | [116] | |
| circ-0005835 | Inhibits NSC proliferation and differentiate to neuron | miR-576-3p | [117] | |
| circ-0002113 | Lacking MSCs suppress myocardial infarction through regulate RUNX1 | miR-188-3p | [118] | |
| CircFAT1 | Regulates osteogenic differentiation of periodontal ligament stem cells (PDLSCs) by regulating SMAD5 | miR-4781-3p | [119] | |
| Cancer stem cells | circ-ITCH | Promotes the self-renewal and stemness of CSCs by repressing the expression of CTNNBIP1 | miR-214 | [120] |
| hsa_circ_0020397 | Promotes malignant proliferation of liver CSCs | miR-138 | [121] | |
| hsa_circ_0005075 | Promotes proliferation and differentiation of breast CSCs and a new biomarker of BCSCs | miR-93 | [122] | |
| Circ-008913 | Regulates acquisition of CSC-like properties and neoplastic capacity of arsenite-transformed HaCaT cells | miR-889 | [123] | |
| Hg19_circ_0005033 | Promotes proliferation, migration, invasion and chemotherapy resistance of CD133+ CD44+ laryngeal CSCs | miR-45121 | [124] | |
| CircPRMT5 | Facilitates UCB cell’s EMT and/or aggressiveness | miR30-c | [125] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, J.; Joseph, S.; Xia, M.; Teng, F.; Chen, X.; Huang, R.; Zhai, L.; Deng, W. Circular RNAs Acting as miRNAs’ Sponges and Their Roles in Stem Cells. J. Clin. Med. 2022, 11, 2909. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm11102909
Xiao J, Joseph S, Xia M, Teng F, Chen X, Huang R, Zhai L, Deng W. Circular RNAs Acting as miRNAs’ Sponges and Their Roles in Stem Cells. Journal of Clinical Medicine. 2022; 11(10):2909. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm11102909
Chicago/Turabian StyleXiao, Juan, Shija Joseph, Mengwei Xia, Feng Teng, Xuejiao Chen, Rufeng Huang, Lihong Zhai, and Wenbin Deng. 2022. "Circular RNAs Acting as miRNAs’ Sponges and Their Roles in Stem Cells" Journal of Clinical Medicine 11, no. 10: 2909. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm11102909