
Single-Cell RNA Transcriptomics Reveals the State of Hepatic Lymphatic Endothelial Cells in Hepatitis B Virus-Related Acute-on-Chronic Liver Failure

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Collection of Human Liver Samples
2.2. Liver Tissue Dissociation and Preparation
2.3. scRNA-seq and Primary Analyses of Raw Read Data
2.4. Quality Control, Dimension Reduction, and Clustering
2.5. Analyses of Signaling-Pathway Enrichment Using the Kyoto Encyclopedia of Genes and Genomes (KEGG) Database
2.6. Ligand/Receptor Analyses
2.7. Measurement of Plasma Alanine Aminotransferase (ALT), Aspartate Aminotransferase (AST), Total Bilirubin (TBil), and International Normalized Ratio (INR)
2.8. Immunohistochemical (IHC) Staining
2.9. Isolation of Intrahepatic Monocyte/Macrophages
2.10. Cell Immunofluorescence Staining
2.11. H&E Staining
2.12. Masson Staining
2.13. Acquisition and Quantification of Images
2.14. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.15. Culture of Human LyECs and SPP1 Treatment
2.16. Flow Cytometry
2.17. Tube Formation Assay for LyECs
2.18. Statistical Analysis
2.19. Ethics Statement
3. Results
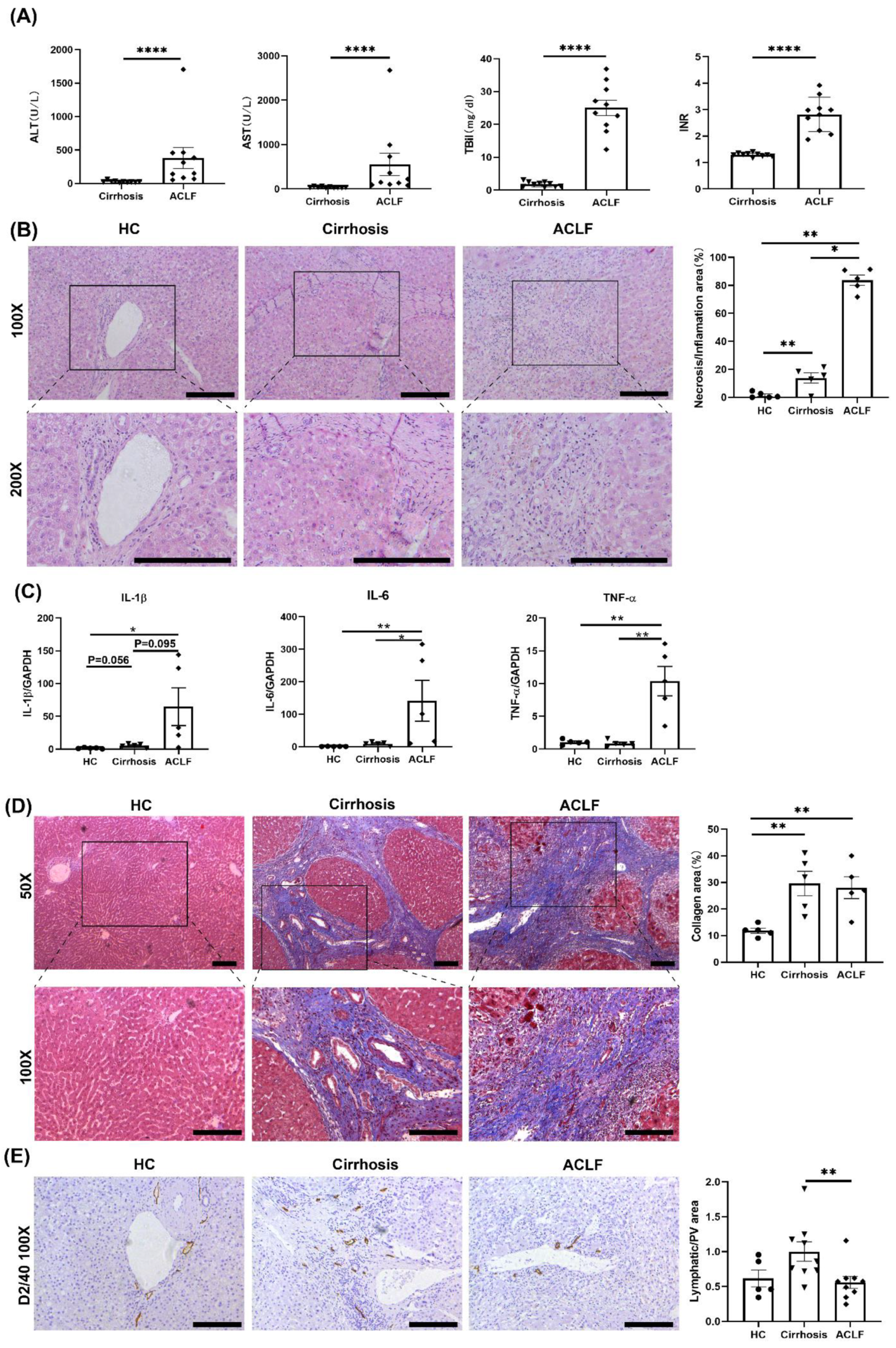
3.1. ACLF Patients Exhibited More Severe Liver Damage and Inflammation and Fewer LVs than Cirrhosis Patients
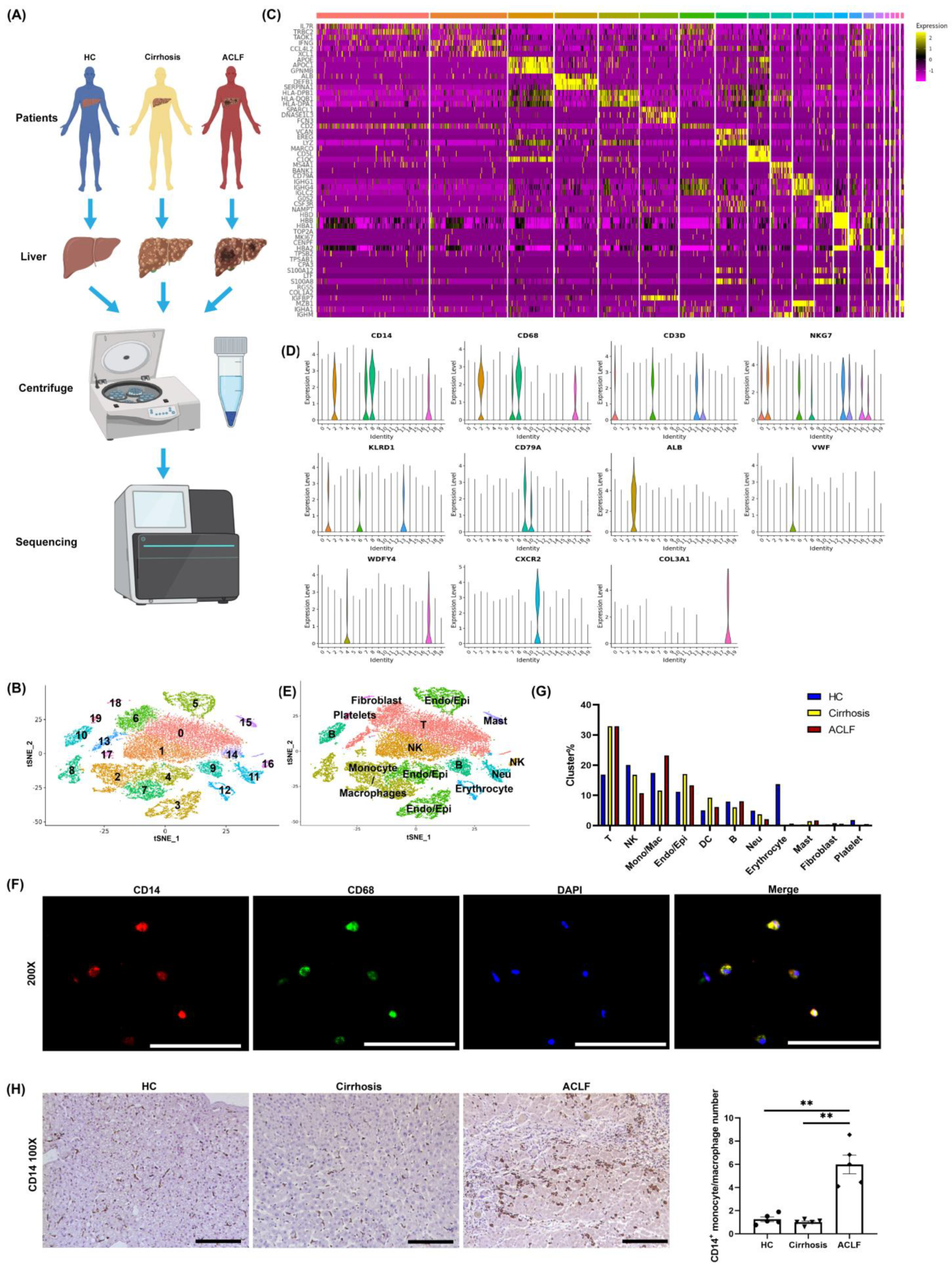
3.2. scRNA-seq Revealed the Infiltration of Many Monocyte/Macrophages into the Liver of ACLF Patients
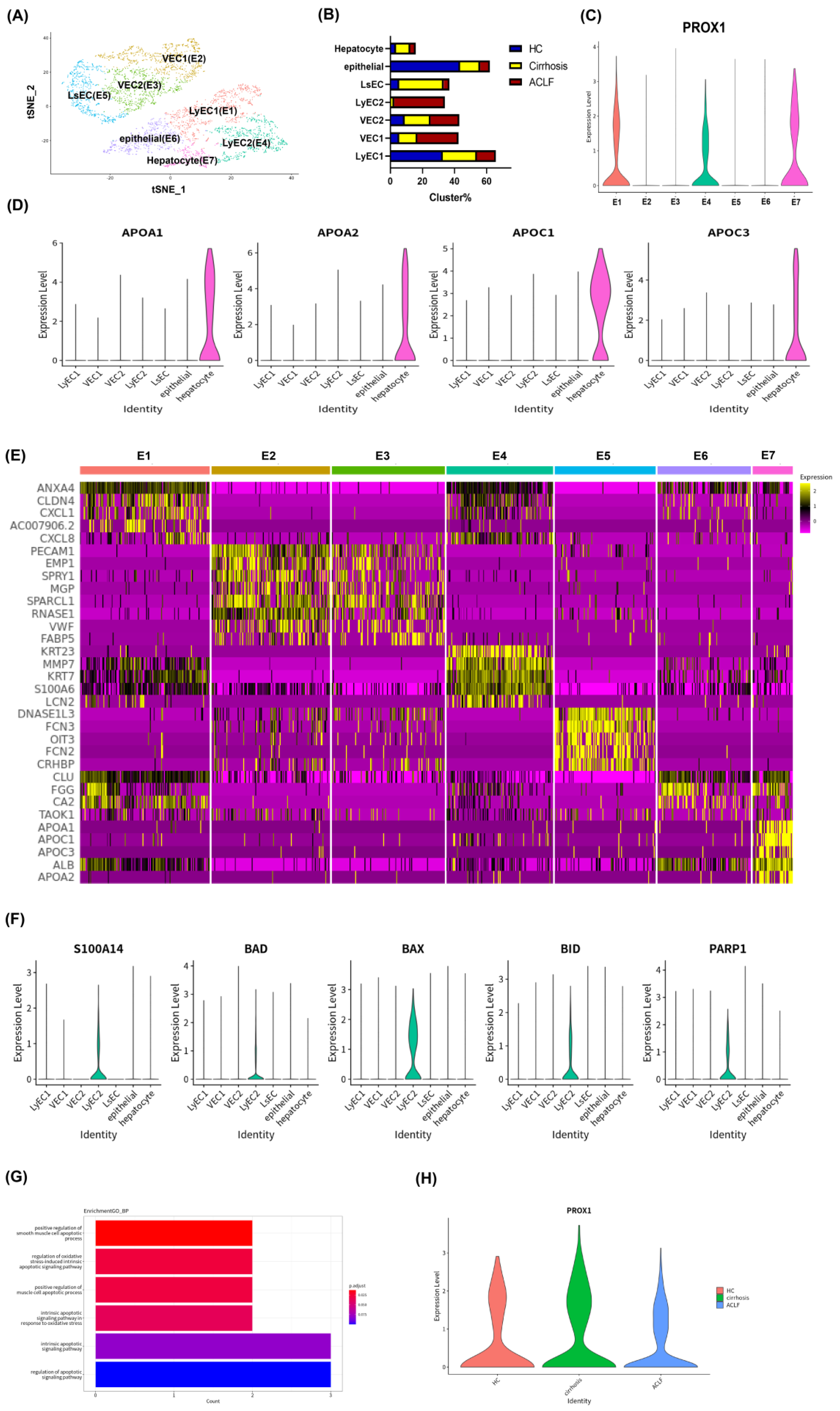
3.3. scRNA-seq Revealed Apoptosis and Dysfunction of Hepatic LyECs in ACLF Patients
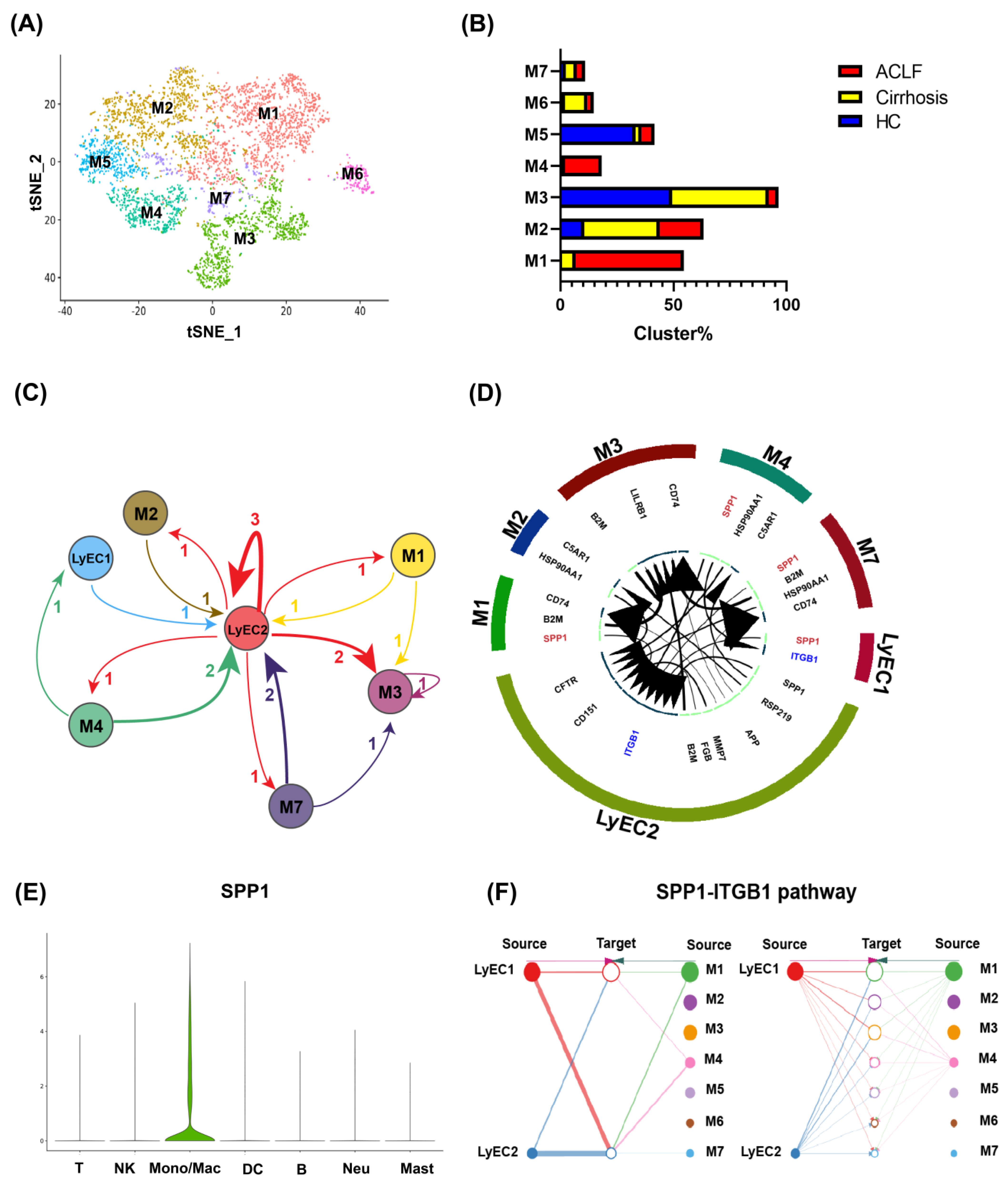
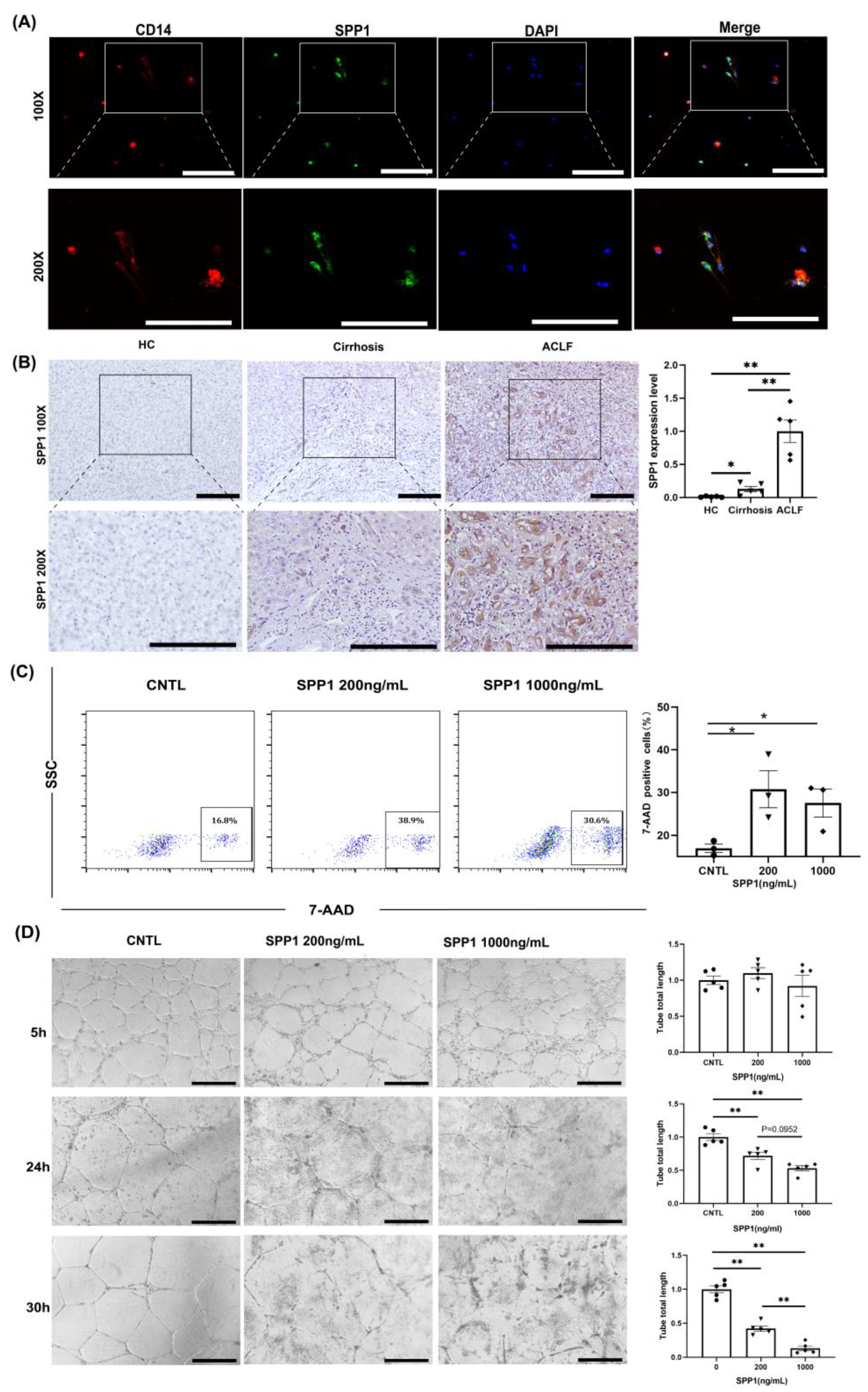
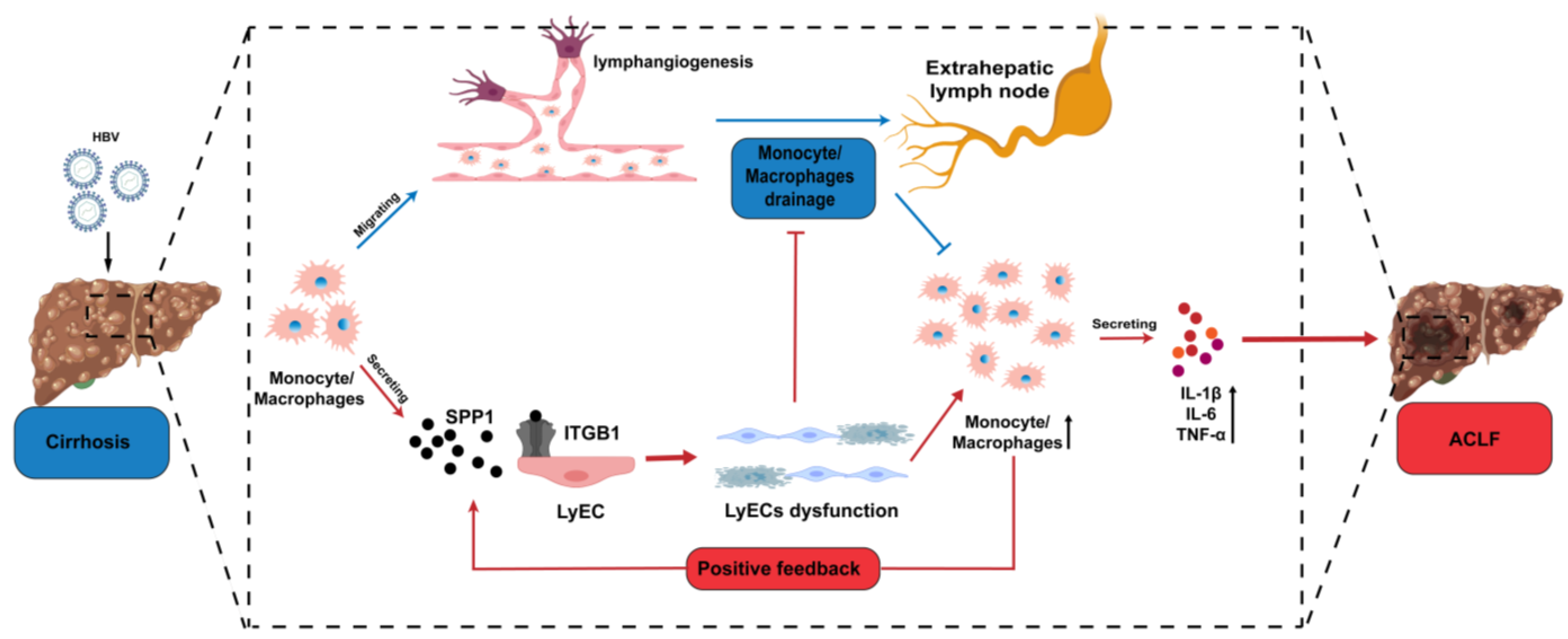
3.4. Infiltrating Monocyte/Macrophages Secreted SPP1 to Induce LyECs Apoptosis and Dysfunction in the Liver of ACLF Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alam, A.; Suen, K.C.; Ma, D. Acute-on-chronic liver failure: Recent update. J. Biomed. Res. 2017, 31, 283–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreau, R.; Jalan, R.; Gines, P.; Pavesi, M.; Angeli, P.; Cordoba, J.; Durand, F.; Gustot, T.; Saliba, F.; Domenicali, M.; et al. Acute-on-Chronic Liver Failure Is a Distinct Syndrome That Develops in Patients with Acute Decompensation of Cirrhosis. Gastroenterology 2013, 144, 1426–1437.e9. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, V.; Moreau, R.; Kamath, P.S.; Jalan, R.; Gines, P.; Nevens, F.; Fernández, J.; To, U.; García-Tsao, G.; Schnabl, B.; et al. Acute-on-chronic liver failure in cirrhosis. Nat. Rev. Dis. Primers 2016, 2, 16041. [Google Scholar] [CrossRef] [Green Version]
- Arroyo, V.; Moreau, R.; Jalan, R. Acute-on-Chronic Liver Failure. N Engl. J. Med. 2020, 382, 2137–2145. [Google Scholar] [CrossRef]
- Sarin, S.K.; Kumar, A.; Almeida, J.A.; Chawla, Y.K.; Fan, S.T.; Garg, H.; de Silva, H.J.; Hamid, S.S.; Jalan, R.; Komolmit, P.; et al. Acute-on-chronic liver failure: Consensus recommendations of the Asian Pacific Association for the study of the liver (APASL). Hepatol. Int. 2008, 3, 269–282. [Google Scholar] [CrossRef]
- You, S.; Rong, Y.; Zhu, B.; Zhang, A.; Zang, H.; Liu, H.; Li, D.; Wan, Z.; Xin, S. Changing etiology of liver failure in 3,916 patients from northern China: A 10-year survey. Hepatol. Int. 2013, 7, 714–720. [Google Scholar] [CrossRef]
- Choudhury, A.; Sharma, M.; Maiwal, R.; Jain, P.; Mahtab, M.; Chawla, Y.; Tan, S.; Ning, Q.; Devarbhavi, H.; Amarapurkar, D.; et al. The Decision for Liver Transplant in Acute on Chronic Liver Failure (ACLF)-First Week is the Crucial Period-Analysis of the Apasl ACLF Research Consortium (AARC) Prospective Data of 1021 Patients. J. Hepatol. 2016, 64, S151–S152. [Google Scholar] [CrossRef]
- Iwakiri, Y. The lymphatic system: A new frontier in hepatology. Hepatology 2016, 64, 706–707. [Google Scholar] [CrossRef] [Green Version]
- Ohtani, O.; Ohtani, Y. Lymph Circulation in the Liver. Anat. Rec. 2008, 291, 643–652. [Google Scholar] [CrossRef]
- Tanaka, M.; Iwakiri, Y. The Hepatic Lymphatic Vascular System: Structure, Function, Markers, and Lymphangiogenesis. Cell. Mol. Gastroenterol. Hepatol. 2016, 2, 733–749. [Google Scholar] [CrossRef] [Green Version]
- Serhan, C.N.; Savill, J. Resolution of inflammation: The beginning programs the end. Nat. Immunol. 2005, 6, 1191–1197. [Google Scholar] [CrossRef] [PubMed]
- Kataru, R.P.; Jung, K.; Jang, C.; Yang, H.; Schwendener, R.A.; Baik, J.E. Critical role of CD11b+ macrophages and VEGF in inflammatory lymphangiogenesis, antigen clearance, and inflammation resolution. Blood 2009, 113, 5650–5659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alitalo, K. The lymphatic vasculature in disease. Nat. Med. 2011, 17, 1371–1380. [Google Scholar] [CrossRef]
- Ribera, J.; Pauta, M.; Melgar-Lesmes, P.; Tugues, S.; Fernández-Varo, G.; Held, K.F.; Soria, G.; Tudela, R.; Planas, A.M.; Fernández-Hernando, C.; et al. Increased nitric oxide production in lymphatic endothelial cells causes impairment of lymphatic drainage in cirrhotic rats. Gut 2012, 62, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Heymann, F.; Tacke, F. Immunology in the liver—From homeostasis to disease. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 88–110. [Google Scholar] [CrossRef] [PubMed]
- Kubes, P.; Mehal, W.Z. Sterile Inflammation in the Liver. Gastroenterology 2012, 143, 1158–1172. [Google Scholar] [CrossRef]
- Mancardi, S.; Vecile, E.; Dusetti, N.; Calvo, E.; Stanta, G.; Burrone, O.R.; Dobrina, A. Evidence of CXC, CC and C chemokine production by lymphatic endothelial cells. Immunology 2003, 108, 523–530. [Google Scholar] [CrossRef]
- Ji, R.-C. Macrophages are important mediators of either tumor- or inflammation-induced lymphangiogenesis. Cell. Mol. Life Sci. 2012, 69, 897–914. [Google Scholar] [CrossRef]
- Triantafyllou, E.; Woollard, K.J.; McPhail, M.; Antoniades, C.G.; Possamai, L.A. The Role of Monocytes and Macrophages in Acute and Acute-on-Chronic Liver Failure. Front. Immunol. 2018, 9, 2948. [Google Scholar] [CrossRef]
- Haque, A.; Engel, J.; Teichmann, S.A.; Lönnberg, T. A practical guide to single-cell RNA-sequencing for biomedical research and clinical applications. Genome Med. 2017, 9, 75. [Google Scholar] [CrossRef]
- Arazi, A.; Rao, D.A.; Berthier, C.C.; Davidson, A.; Liu, Y.; Hoover, P.J.; Chicoine, A.; Eisenhaure, T.M.; Jonsson, A.H.; Li, S.; et al. The immune cell landscape in kidneys of patients with lupus nephritis. Nat. Immunol. 2019, 20, 902–914. [Google Scholar] [CrossRef] [PubMed]
- Su, T.; Yang, Y.; Lai, S.; Jeong, J.; Jung, Y.; McConnell, M.; Utsumi, T.; Iwakiri, Y. Single-Cell Transcriptomics Reveals Zone-Specific Alterations of Liver Sinusoidal Endothelial Cells in Cirrhosis. Cell. Mol. Gastroenterol. Hepatol. 2020, 11, 1139–1161. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Li, J.; Shao, L.; Xin, J.; Jiang, L.; Zhou, Q.; Shi, D.; Jiang, J.; Sun, S.; Jin, L.; et al. Development of diagnostic criteria and a prognostic score for hepatitis B virus-related acute-on-chronic liver failure. Gut 2017, 67, 2181–2191. [Google Scholar] [CrossRef] [PubMed]
- Kegel, V.; Deharde, D.; Pfeiffer, E.; Zeilinger, K.; Seehofer, D.; Damm, G. Protocol for Isolation of Primary Human Hepatocytes and Corresponding Major Populations of Non-parenchymal Liver Cells. J. Vis. Exp. 2016, 109, e53069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; He, Y.; Luo, N.; Patel, S.J.; Han, Y.; Gao, R.; Modak, M.; Carotta, S.; Haslinger, C.; Kind, D.; et al. Landscape and Dynamics of Single Immune Cells in Hepatocellular Carcinoma. Cell 2019, 179, 829–845.e20. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Fan, J.; He, Y.; Xiong, A.; Yu, J.; Li, Y.; Zhang, Y.; Zhao, W.; Zhou, F.; Li, W.; et al. Single-cell profiling of tumor heterogeneity and the microenvironment in advanced non-small cell lung cancer. Nat. Commun. 2021, 12, 1–11. [Google Scholar] [CrossRef]
- Tang, R.; Meng, T.; Lin, W.; Shen, C.; Ooi, J.D.; Eggenhuizen, P.J.; Jin, P.; Ding, X.; Chen, J.; Tang, Y.; et al. A Partial Picture of the Single-Cell Transcriptomics of Human IgA Nephropathy. Front. Immunol. 2021, 12, 645988. [Google Scholar] [CrossRef]
- Dura, B.; Choi, J.; Zhang, K.; Damsky, W.; Thakral, D.; Bosenberg, M. scFTD-seq: Freeze-thaw lysis based, portable approach toward highly distributed single-cell 3’ mRNA profiling. Nucleic Acids Res. 2019, 47, e16. [Google Scholar] [CrossRef] [Green Version]
- Satija, R.; Farrell, J.; Gennert, D.; Schier, A.F.; Regev, A. Spatial reconstruction of single-cell gene expression data. Nat. Biotechnol. 2015, 33, 495–502. [Google Scholar] [CrossRef] [Green Version]
- Shi, W.; Wang, Y.; Zhang, C.; Jin, H.; Zeng, Z.; Wei, L.; Tian, Y.; Zhang, D.; Sun, G. Isolation and purification of immune cells from the liver. Int. Immunopharmacol. 2020, 85, 106632. [Google Scholar] [CrossRef]
- Kubota, Y.; Kleinman, H.K.; Martin, G.R.; Lawley, T.J. Role of laminin and basement membrane in the morphological differentiation of human endothelial cells into capillary-like structures. J. Cell Biol. 1988, 107, 1589–1598. [Google Scholar] [CrossRef] [PubMed]
- Arnaoutova, I.; George, J.; Kleinman, H.K.; Benton, G. The endothelial cell tube formation assay on basement membrane turns 20: State of the science and the art. Angiogenesis 2009, 12, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Arnaoutova, I.; Kleinman, H.K. In vitro angiogenesis: Endothelial cell tube formation on gelled basement membrane extract. Nat. Protoc. 2010, 5, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Wigle, J.T.; Oliver, G. Prox1 Function Is Required for the Development of the Murine Lymphatic System. Cell 1999, 98, 769–778. [Google Scholar] [CrossRef] [Green Version]
- Wigle, J.; Harvey, N.; Detmar, M.; Lagutina, I.; Grosveld, G.; Gunn, M.; Jackson, D.G.; Oliver, G. An essential role for Prox1 in the induction of the lymphatic endothelial cell phenotype. EMBO J. 2002, 21, 1505–1513. [Google Scholar] [CrossRef] [PubMed]
- Wilting, J.; Papoutsi, M.; Christ, B.; Nicolaides, K.H.; von Kaisenberg, C.S.; Borges, J.; Stark, G.B.; Alitalo, K.; Tomarev, S.I.; Niemeyer, C.; et al. The transcription factor Prox1 is a marker for lymphatic endothelial cells in normal and diseased human tissues. FASEB J. 2002, 16, 1271–1273. [Google Scholar] [CrossRef] [PubMed]
- Takeda, A.; Hollmén, M.; Dermadi, D.; Pan, J.; Brulois, K.F.; Kaukonen, R.; Lönnberg, T.; Boström, P.; Koskivuo, I.; Irjala, H.; et al. Single-Cell Survey of Human Lymphatics Unveils Marked Endothelial Cell Heterogeneity and Mechanisms of Homing for Neutrophils. Immunity 2019, 51, 561–572.e5. [Google Scholar] [CrossRef]
- Burchill, M.A.; Finlon, J.M.; Goldberg, A.R.; Gillen, A.E.; Dahms, P.A.; McMahan, R.H.; Tye, A.; Winter, A.B.; Reisz, J.A.; Bohrnsen, E.; et al. Oxidized Low-Density Lipoprotein Drives Dysfunction of the Liver Lymphatic System. Cell. Mol. Gastroenterol. Hepatol. 2020, 11, 573–595. [Google Scholar] [CrossRef]
- André, F.; Corazao-Rozas, P.; Idziorek, T.; Quesnel, B.; Kluza, J.; Marchetti, P. GILZ overexpression attenuates endoplasmic reticulum stress-mediated cell death via the activation of mitochondrial oxidative phosphorylation. Biochem. Biophys. Res. Commun. 2016, 478, 513–520. [Google Scholar] [CrossRef]
- Du, W.; Amarachintha, S.; Wilson, A.F.; Pang, Q. SCO2 Mediates Oxidative Stress-Induced Glycolysis to Oxidative Phosphorylation Switch in Hematopoietic Stem Cells. Stem Cells 2015, 34, 960–971. [Google Scholar] [CrossRef] [Green Version]
- Jin, Q.; Chen, H.; Luo, A.; Ding, F.; Liu, Z. S100A14 Stimulates Cell Proliferation and Induces Cell Apoptosis at Different Concentrations via Receptor for Advanced Glycation End Products (RAGE). PLoS ONE 2011, 6, e19375. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Cao, J.; Chen, Z. Mining prognostic markers of Asian hepatocellular carcinoma patients based on the apoptosis-related genes. BMC Cancer 2021, 21, 175. [Google Scholar] [CrossRef] [PubMed]
- Honarpisheh, M.; Desai, J.; Marschner, J.A.; Weidenbusch, M.; Lech, M.; Vielhauer, V.; Anders, H.-J.; Mulay, S.R. Regulated necrosis-related molecule mRNA expression in humans and mice and in murine acute tissue injury and systemic autoimmunity leading to progressive organ damage, and progressive fibrosis. Biosci. Rep. 2016, 36, e00425. [Google Scholar] [CrossRef] [PubMed]
- Katoonizadeh, A.; Laleman, W.; Verslype, C.; Wilmer, A.; Maleux, G.; Roskams, T.; Nevens, F. Early features of acute-on-chronic alcoholic liver failure: A prospective cohort study. Gut 2010, 59, 1561–1569. [Google Scholar] [CrossRef] [PubMed]
- Clària, J.; Stauber, R.E.; Coenraad, M.J.; Moreau, R.; Jalan, R.; Pavesi, M.; Amorós, A.; Titos, E.; Alcaraz-Quiles, J.; Oettl, K.; et al. Systemic inflammation in decompensated cirrhosis: Characterization and role in acute-on-chronic liver failure. Hepatology 2016, 64, 1249–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trebicka, J.; Amoros, A.; Pitarch, C.; Titos, E.; Alcaraz-Quiles, J.; Schierwagen, R.; Deulofeu, C.; Fernandez-Gomez, J.; Piano, S.; Caraceni, P.; et al. Addressing Profiles of Systemic Inflammation Across the Different Clinical Phenotypes of Acutely Decompensated Cirrhosis. Front. Immunol. 2019, 10, 476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajaj, J.; Moreau, R.; Kamath, P.; Vargas, H.; Arroyo, V.; Reddy, K. Acute-on-Chronic Liver Failure: Getting Ready for Prime Time? Hepatology 2018, 68, 1621–1632. [Google Scholar] [CrossRef] [Green Version]
- Casulleras, M.; Zhang, I.W.; López-Vicario, C.; Clària, J. Leukocytes, Systemic Inflammation and Immunopathology in Acute-on-Chronic Liver Failure. Cells 2020, 9, 2632. [Google Scholar] [CrossRef]
- Liu, F.; Duan, X.; Wan, Z.; Zang, H.; You, S.; Yang, R.; Liu, H.; Li, D.; Li, J.; Zhang, Y.; et al. Lower number and decreased function of natural killer cells in hepatitis B virus related acute-on-chronic liver failure. Clin. Res. Hepatol. Gastroenterol. 2016, 40, 605–613. [Google Scholar] [CrossRef]
- Weiss, E.; de la Grange, P.; Defaye, M.; Lozano, J.J.; Aguilar, F.; Hegde, P.; Jolly, A.; Moga, L.; Sukriti, S.; Agarwal, B.; et al. Characterization of Blood Immune Cells in Patients With Decompensated Cirrhosis Including ACLF. Front. Immunol. 2021, 11, 619039. [Google Scholar] [CrossRef]
- Yamauchi, Y.; Michitaka, K.; Onji, M. Morphometric Analysis of Lymphatic and Blood Vessels in Human Chronic Viral Liver Diseases. Am. J. Pathol. 1998, 153, 1131–1137. [Google Scholar] [CrossRef] [Green Version]
- Tugues, S.; Morales-Ruiz, M.; Fern´ández-Varo, G.; Ros, J.; Arteta, D.; Muñoz, J.; Arroyo, V.; Rodés, J.; Jiménez, W. Microarray Analysis of Endothelial Differentially Expressed Genes in Liver of Cirrhotic Rats. Gastroenterology 2005, 129, 1686–1695. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Iwakiri, Y. Lymphatics in the liver. Curr. Opin. Immunol. 2018, 53, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Oliver, G. Lymphatic Endothelial Cell Plasticity in Development and Disease. Physiology 2017, 32, 444–452. [Google Scholar] [CrossRef]
- Iida, T.; Wagatsuma, K.; Hirayama, D.; Nakase, H. Is Osteopontin a Friend or Foe of Cell Apoptosis in Inflammatory Gastrointestinal and Liver Diseases? Int. J. Mol. Sci. 2017, 19, 7. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Lu, J.; Ye, C.; Lin, L.; Zheng, S.; Zhang, H. Serum osteopontin is a predictor of prognosis for HBV-associated acute-on-chronic liver failure. Biomed Rep. 2018, 8, 166–171. [Google Scholar] [CrossRef] [Green Version]
- Zhou, N.; Wang, K.; Fang, S.; Zhao, X.; Huang, T.; Chen, H.; Yan, F.; Tang, Y.; Zhou, H.; Zhu, J. Discovery of a Potential Plasma Protein Biomarker Panel for Acute-on-Chronic Liver Failure Induced by Hepatitis B Virus. Front. Physiol. 2017, 8, 1009. [Google Scholar] [CrossRef] [Green Version]
- Leung, K. (64)Cu-1,4,7-Triazacyclononane,1-glutaric acid-4,7-acetic acid-cyclo(Arg-Gly-Asp-d-Tyr-Lys). In Molecular Imaging and Contrast Agent Database (MICAD); National Center for Biotechnology Information: Bethesda, MD, USA, 2004. [Google Scholar]
- Guo, Q.; Furuta, K.; Lucien, F.; Sanchez, L.H.G.; Hirsova, P.; Krishnan, A.; Kabashima, A.; Pavelko, K.D.; Madden, B.; Alhuwaish, H.; et al. Integrin β1-enriched extracellular vesicles mediate monocyte adhesion and promote liver inflammation in murine NASH. J. Hepatol. 2019, 71, 1193–1205. [Google Scholar] [CrossRef]
- Lamort, A.-S.; Giopanou, I.; Psallidas, I.; Stathopoulos, G.T. Osteopontin as a Link between Inflammation and Cancer: The Thorax in the Spotlight. Cells 2019, 8, 815. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Chen, W.; Athavale, D.; Ge, X.; Desert, R.; Das, S.; Han, H.; Nieto, N. Osteopontin Takes Center Stage in Chronic Liver Disease. Hepatology 2020, 73, 1594–1608. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, P.; Li, H.; Zhou, C.; Liu, K.; Peng, B.; She, X.; Cheng, K.; Liu, H.; Ming, Y. Single-Cell RNA Transcriptomics Reveals the State of Hepatic Lymphatic Endothelial Cells in Hepatitis B Virus-Related Acute-on-Chronic Liver Failure. J. Clin. Med. 2022, 11, 2910. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm11102910
Zhang P, Li H, Zhou C, Liu K, Peng B, She X, Cheng K, Liu H, Ming Y. Single-Cell RNA Transcriptomics Reveals the State of Hepatic Lymphatic Endothelial Cells in Hepatitis B Virus-Related Acute-on-Chronic Liver Failure. Journal of Clinical Medicine. 2022; 11(10):2910. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm11102910
Chicago/Turabian StyleZhang, Pengpeng, Hao Li, Chen Zhou, Kai Liu, Bo Peng, Xingguo She, Ke Cheng, Hong Liu, and Yingzi Ming. 2022. "Single-Cell RNA Transcriptomics Reveals the State of Hepatic Lymphatic Endothelial Cells in Hepatitis B Virus-Related Acute-on-Chronic Liver Failure" Journal of Clinical Medicine 11, no. 10: 2910. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm11102910