
The Development of Peritoneal Metastasis from Gastric Cancer and Rationale of Treatment According to the Mechanism

 ,
,
Abstract
:1. Introduction
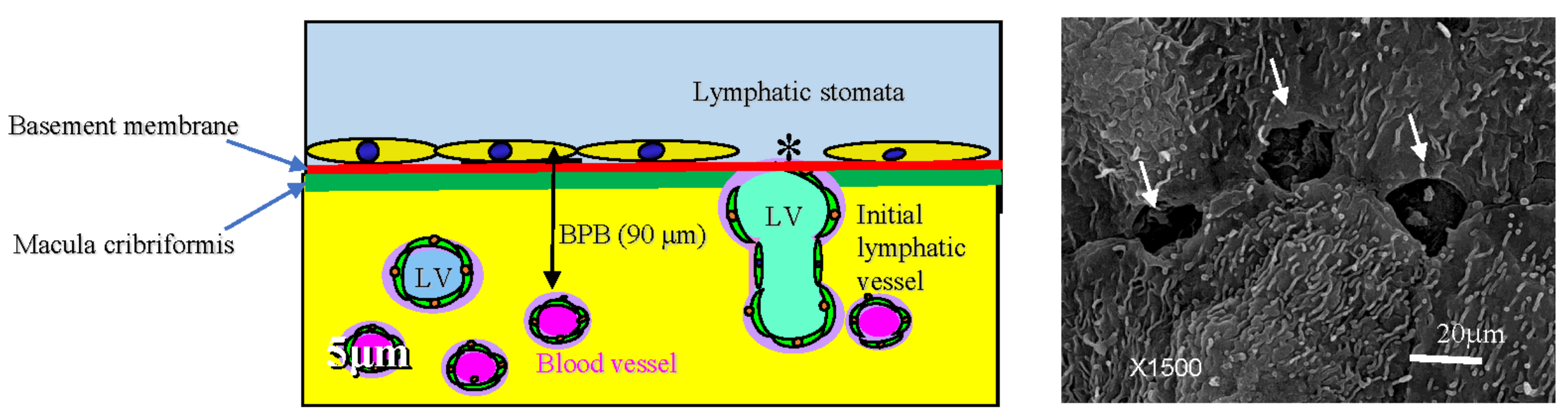
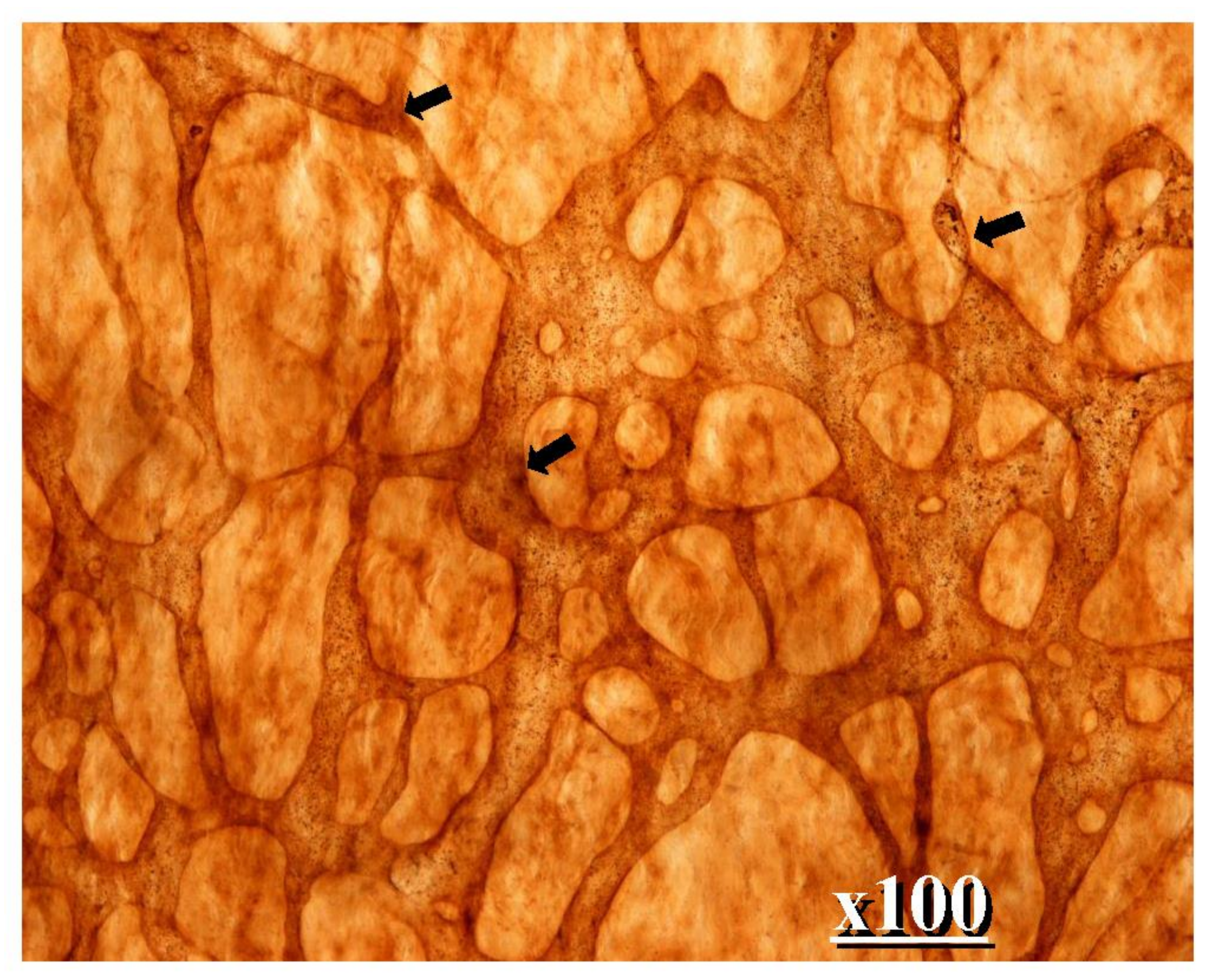
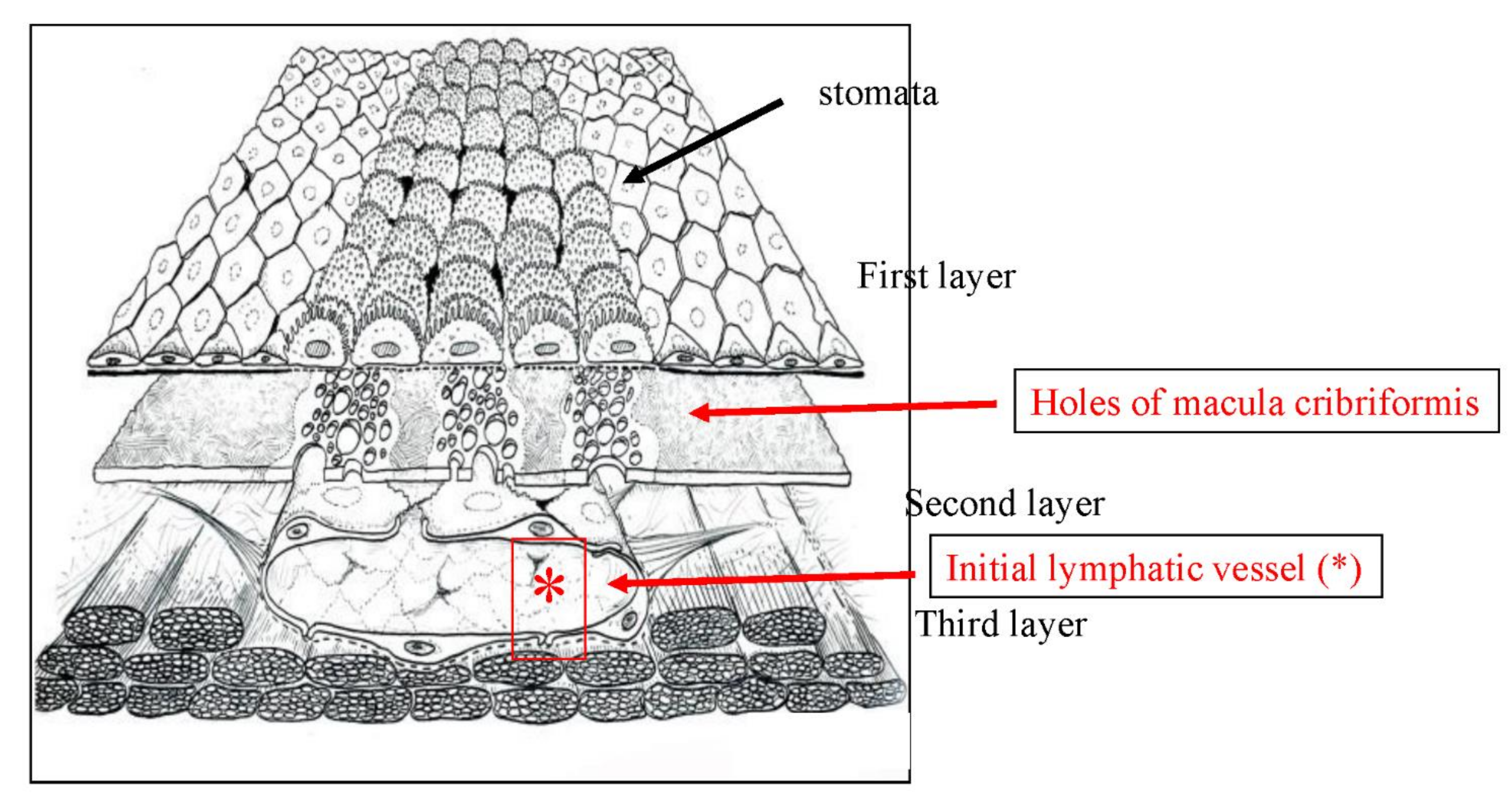
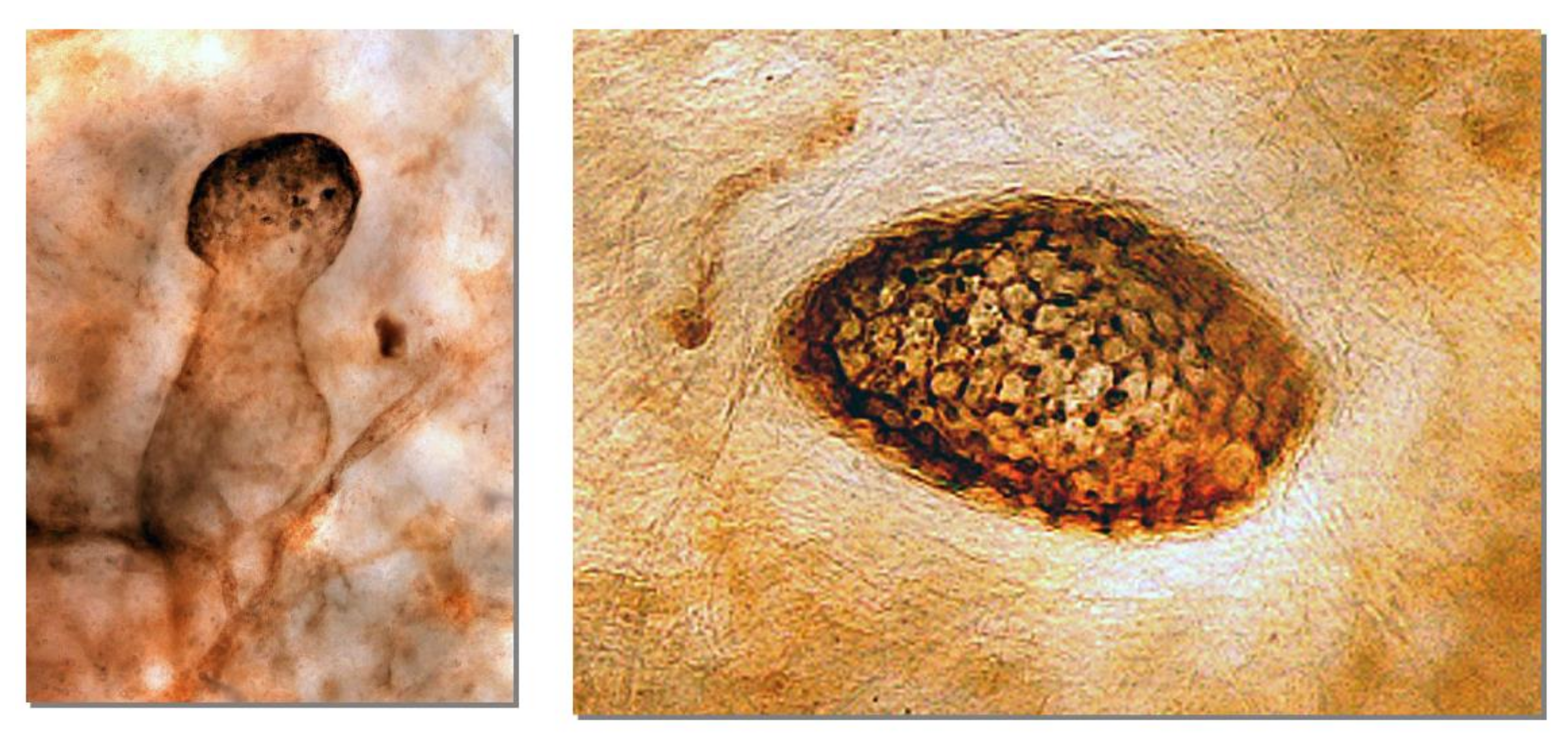
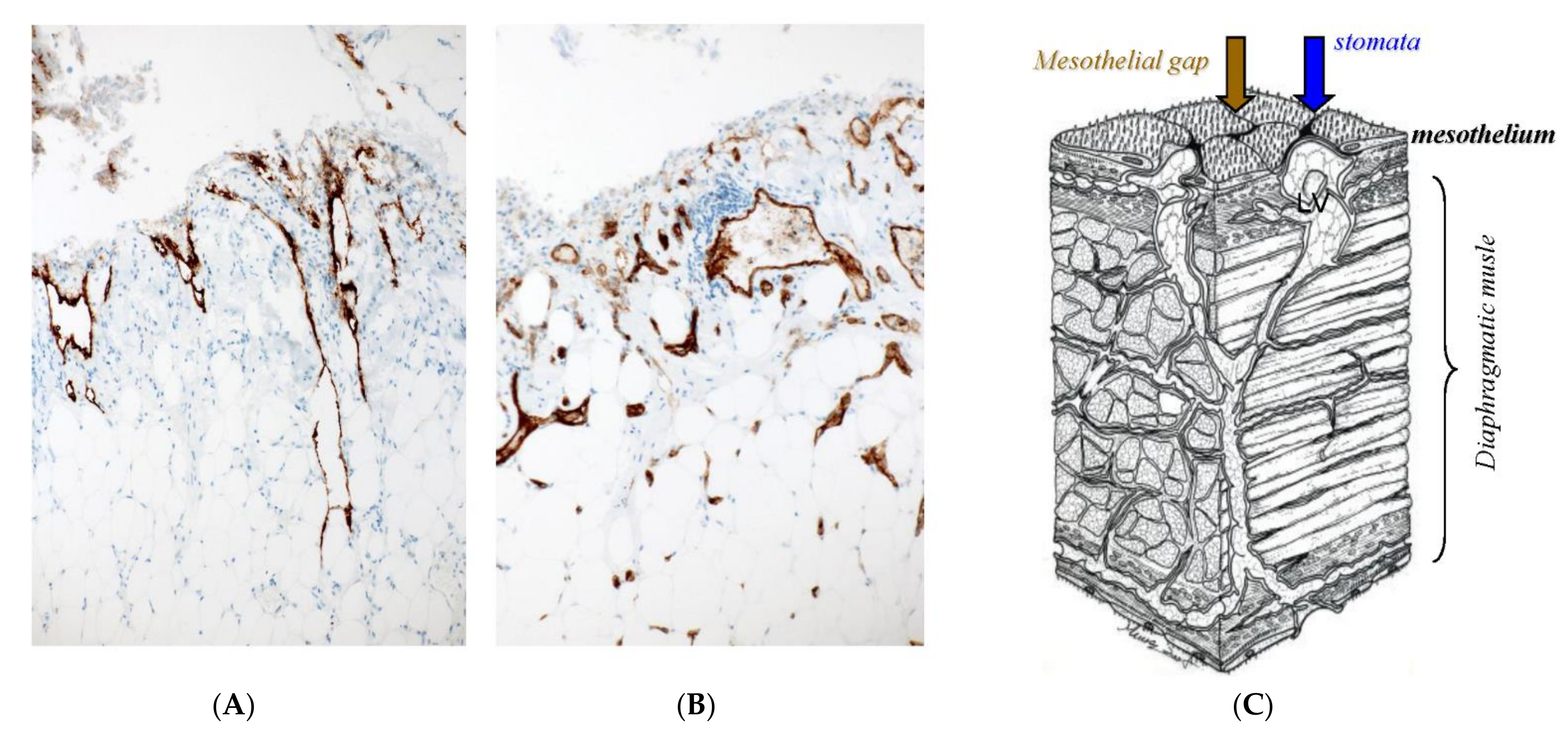
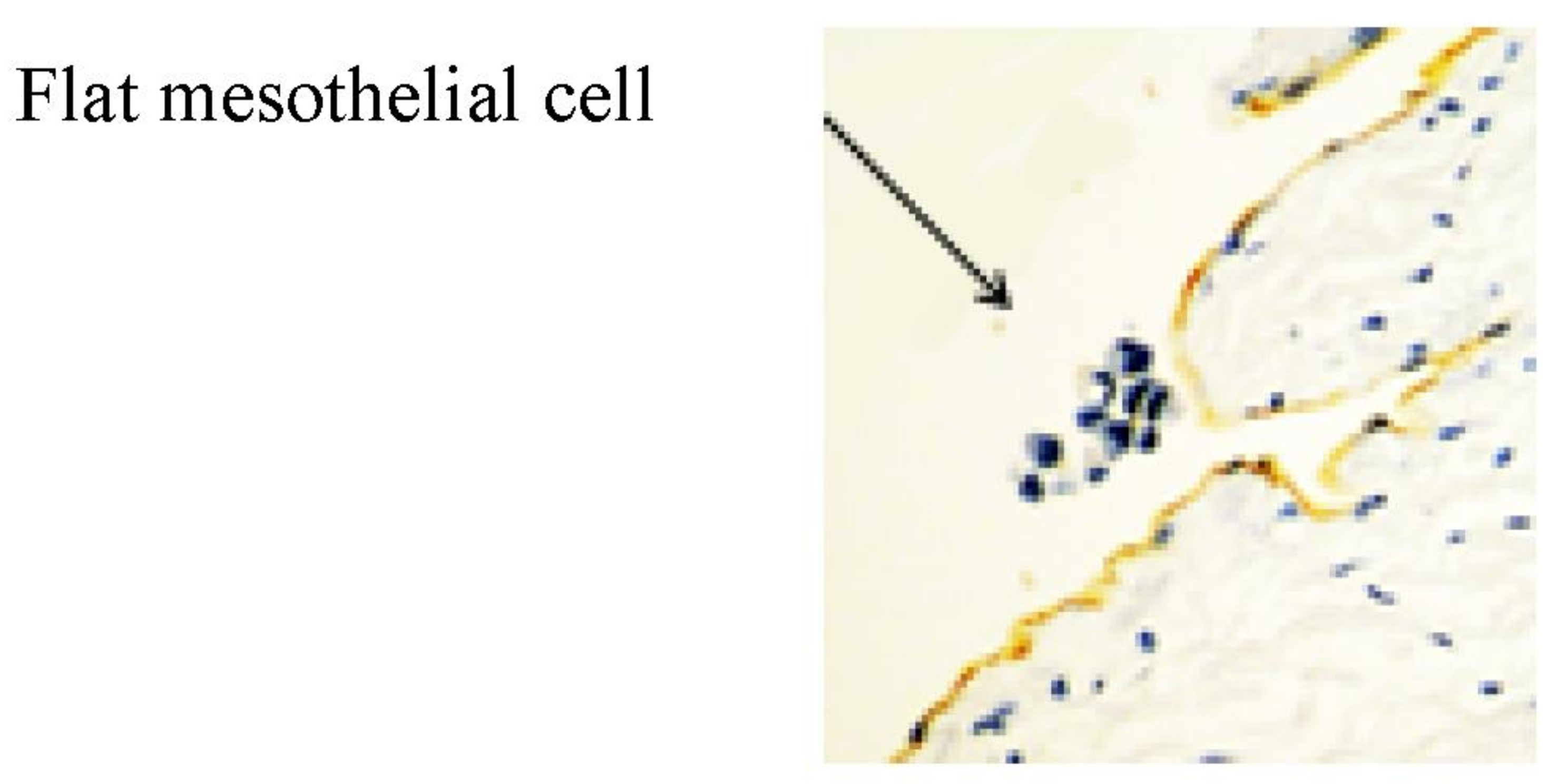
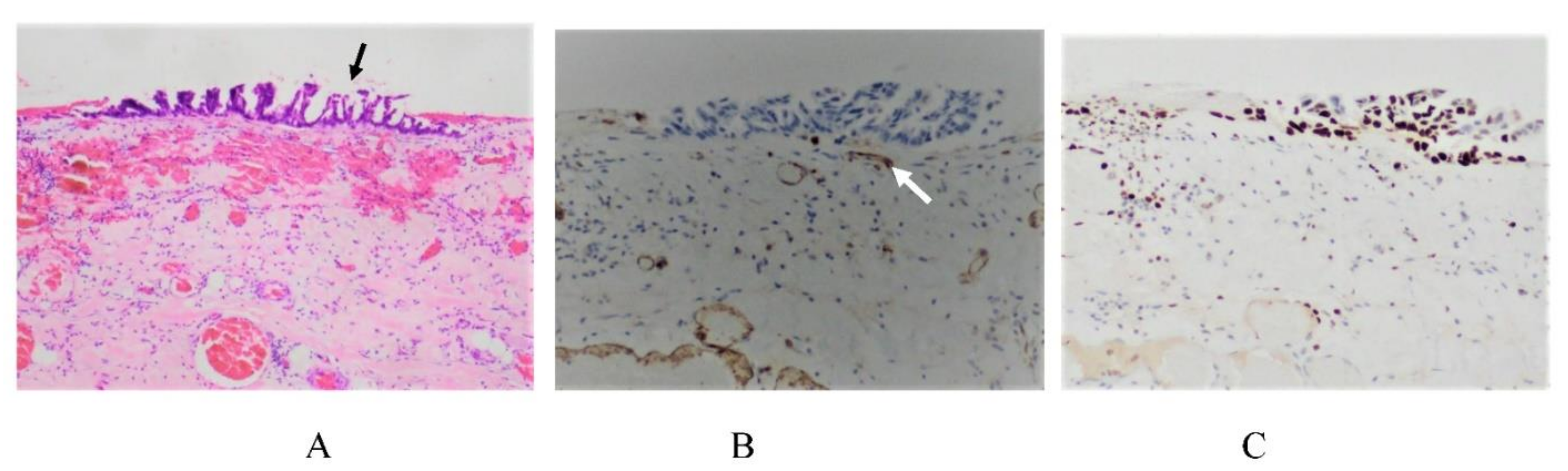
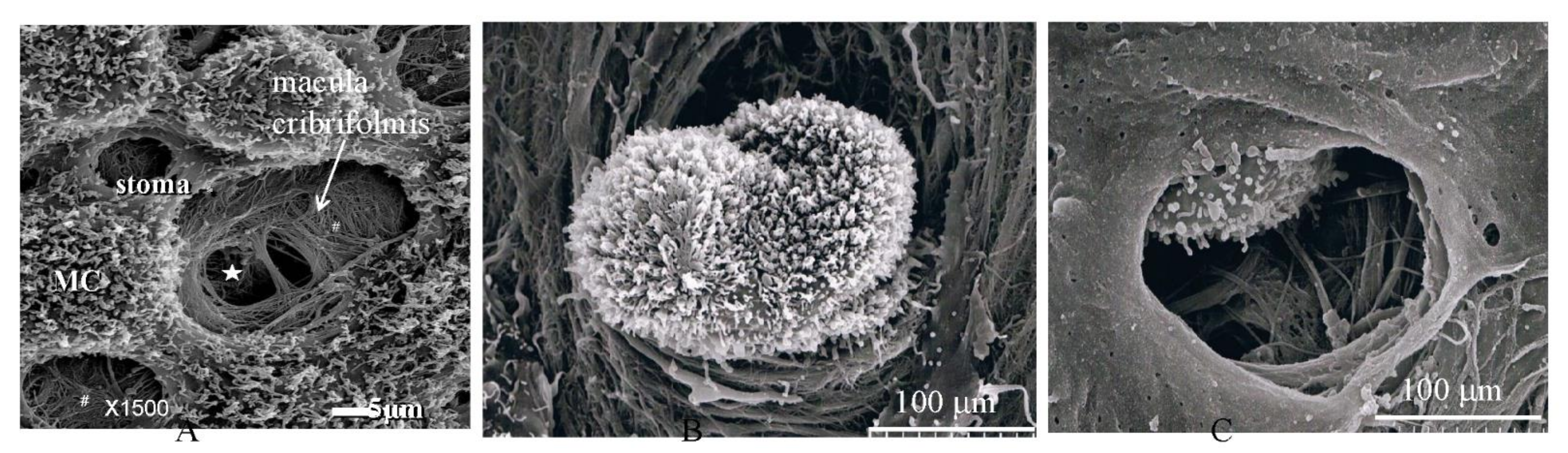
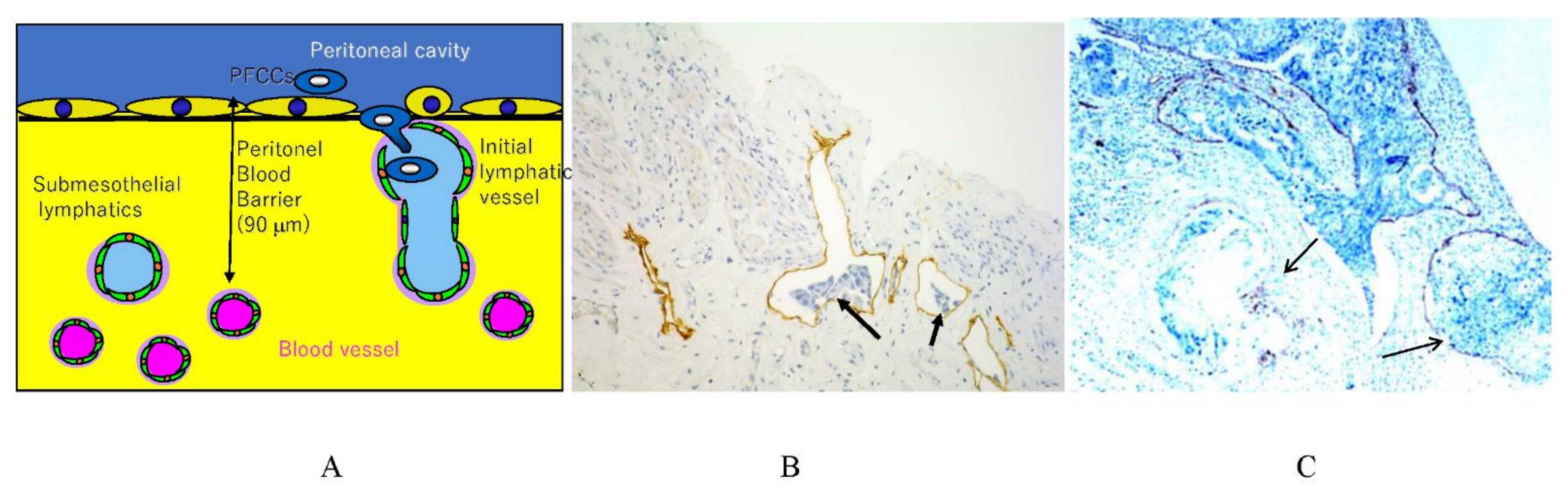
2. Normal Structure of Peritoneum
3. Mechanisms of the Formation of PM from GC
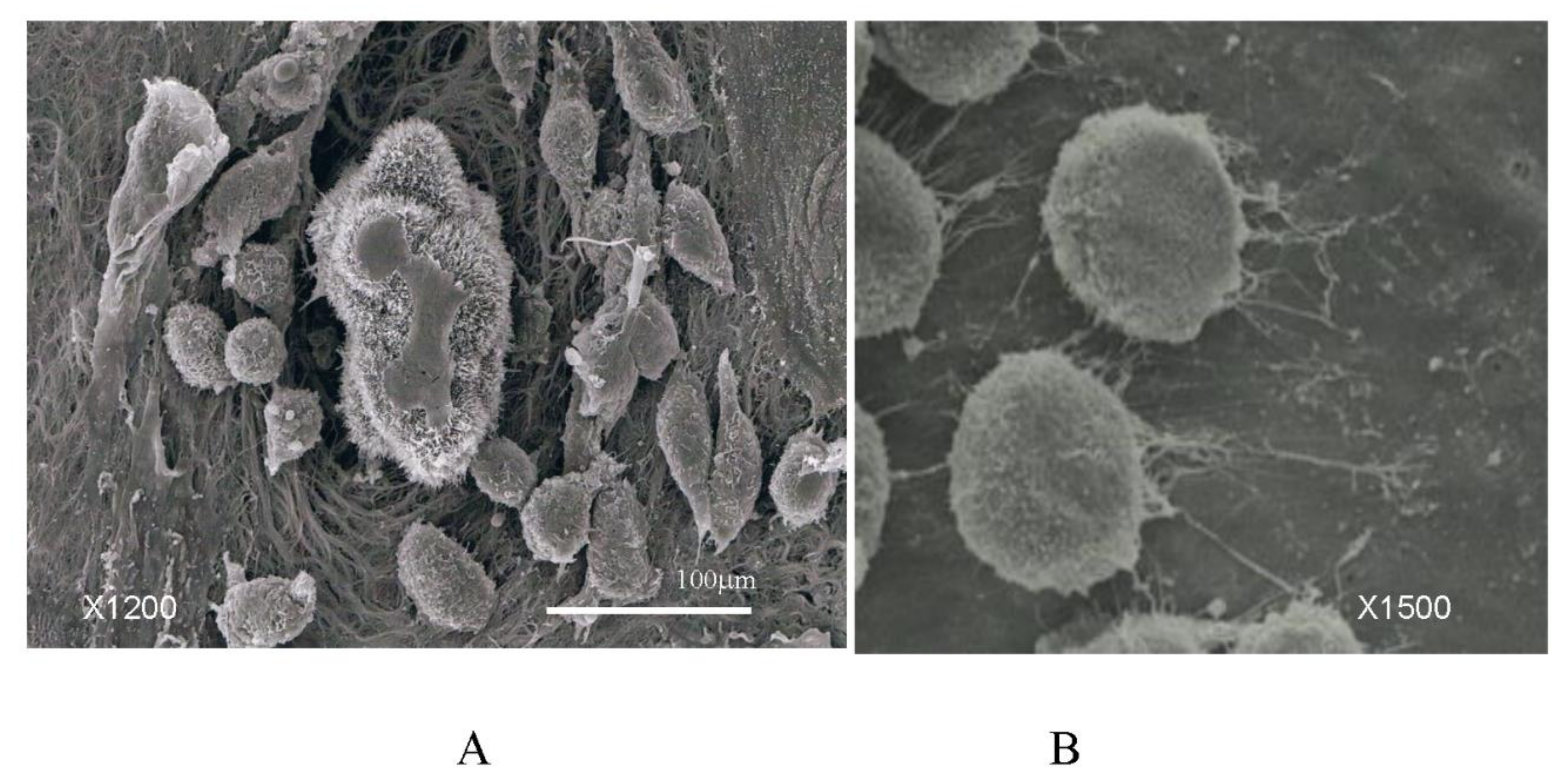
3.1. Mechanisms of the Cell Spillage into the Peritoneal Cavity
3.2. Adhesion of PFCCs to the Distant Peritoneum
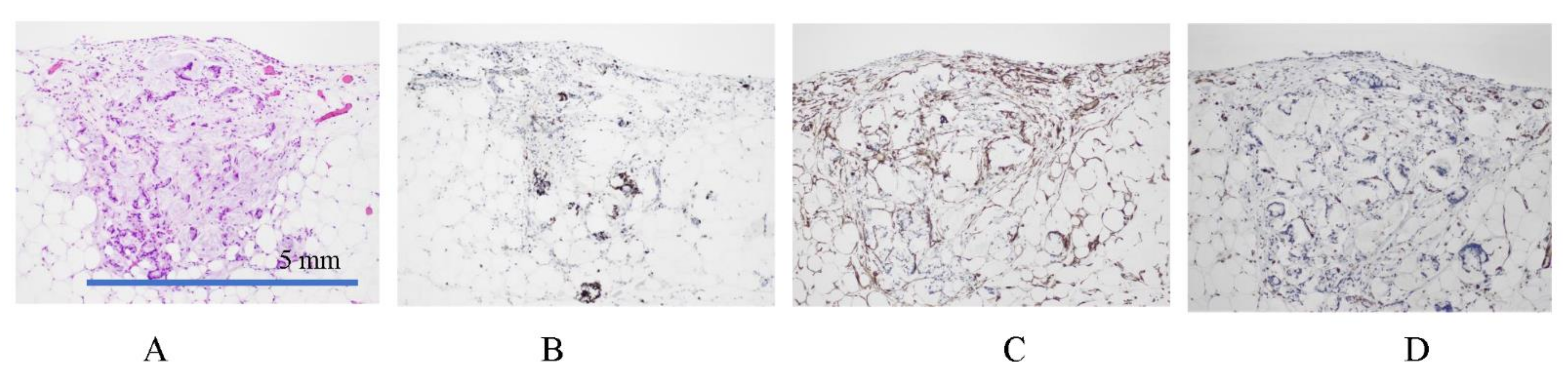
3.3. Mechanisms of Trans-Mesothelial Metastasis
3.4. Mechanisms of Superficial Growing Metastasis
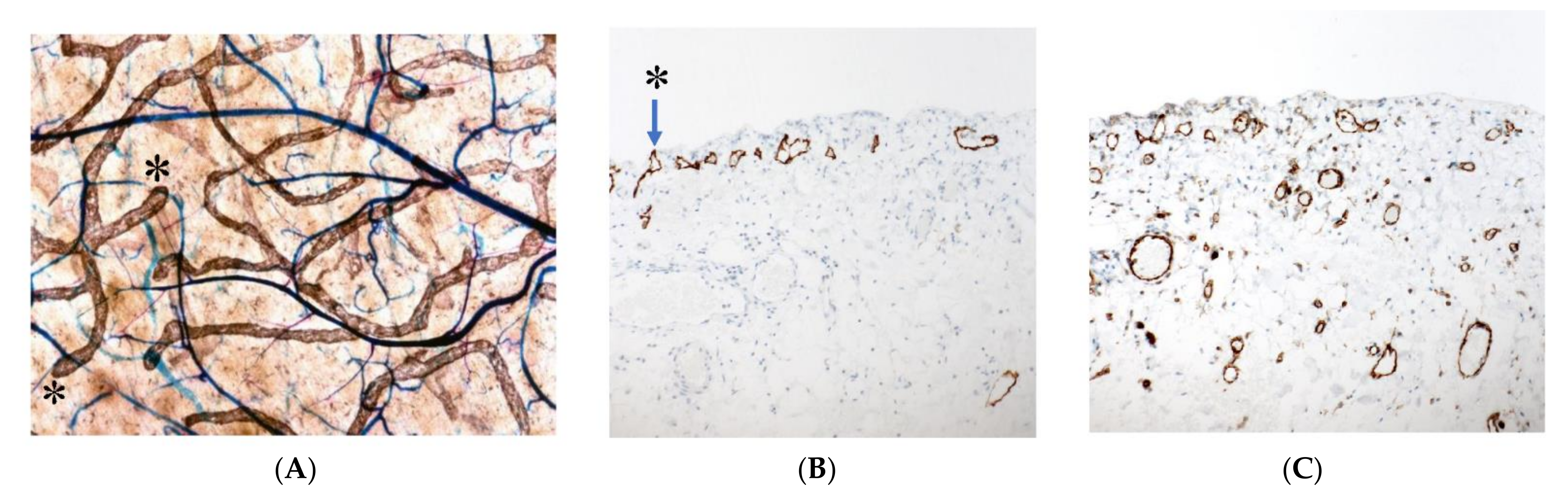
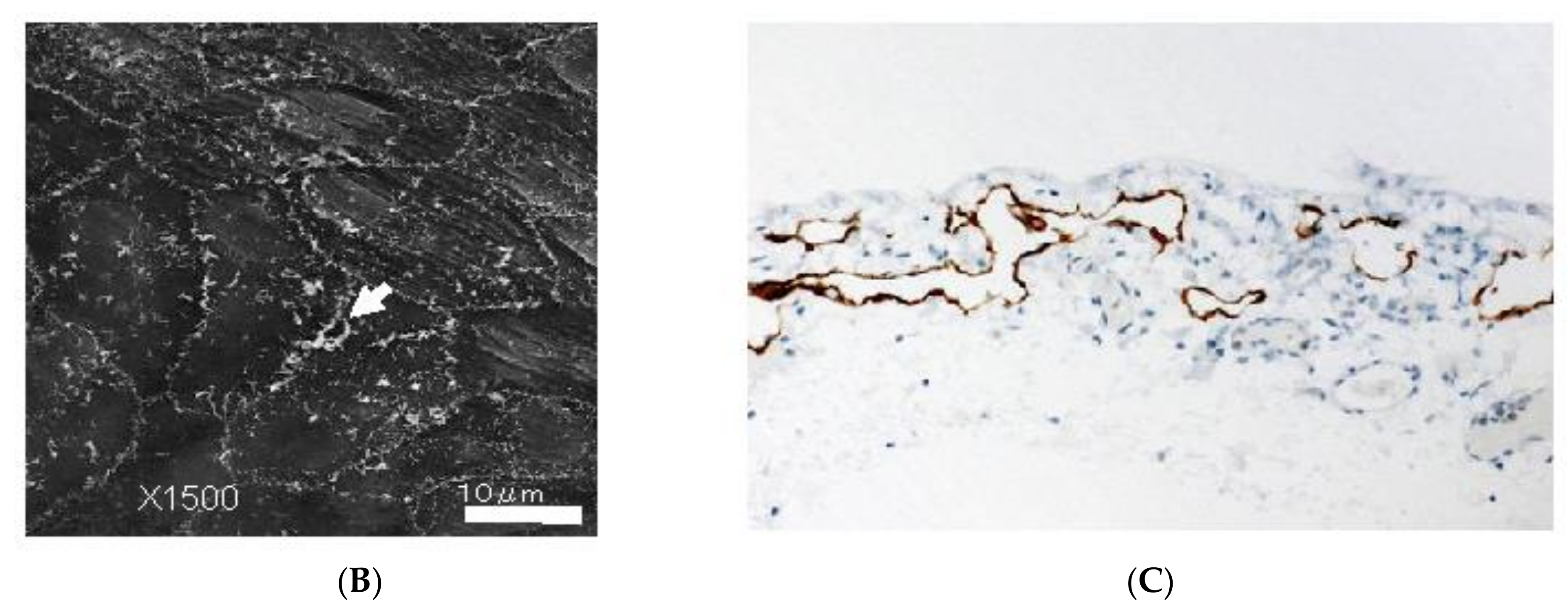
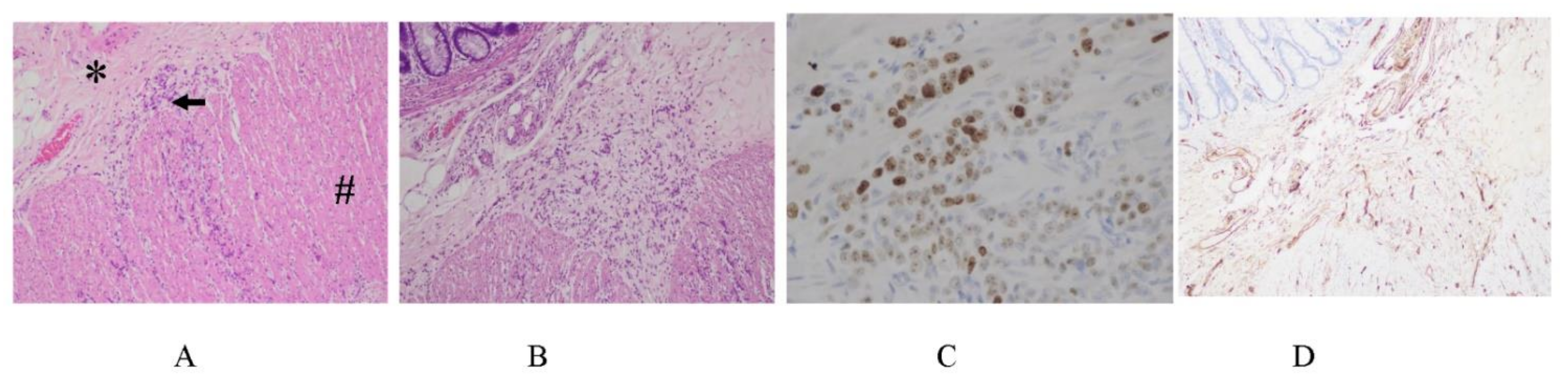
3.5. Mechnisms of Trans-Lymphatic Metastasis
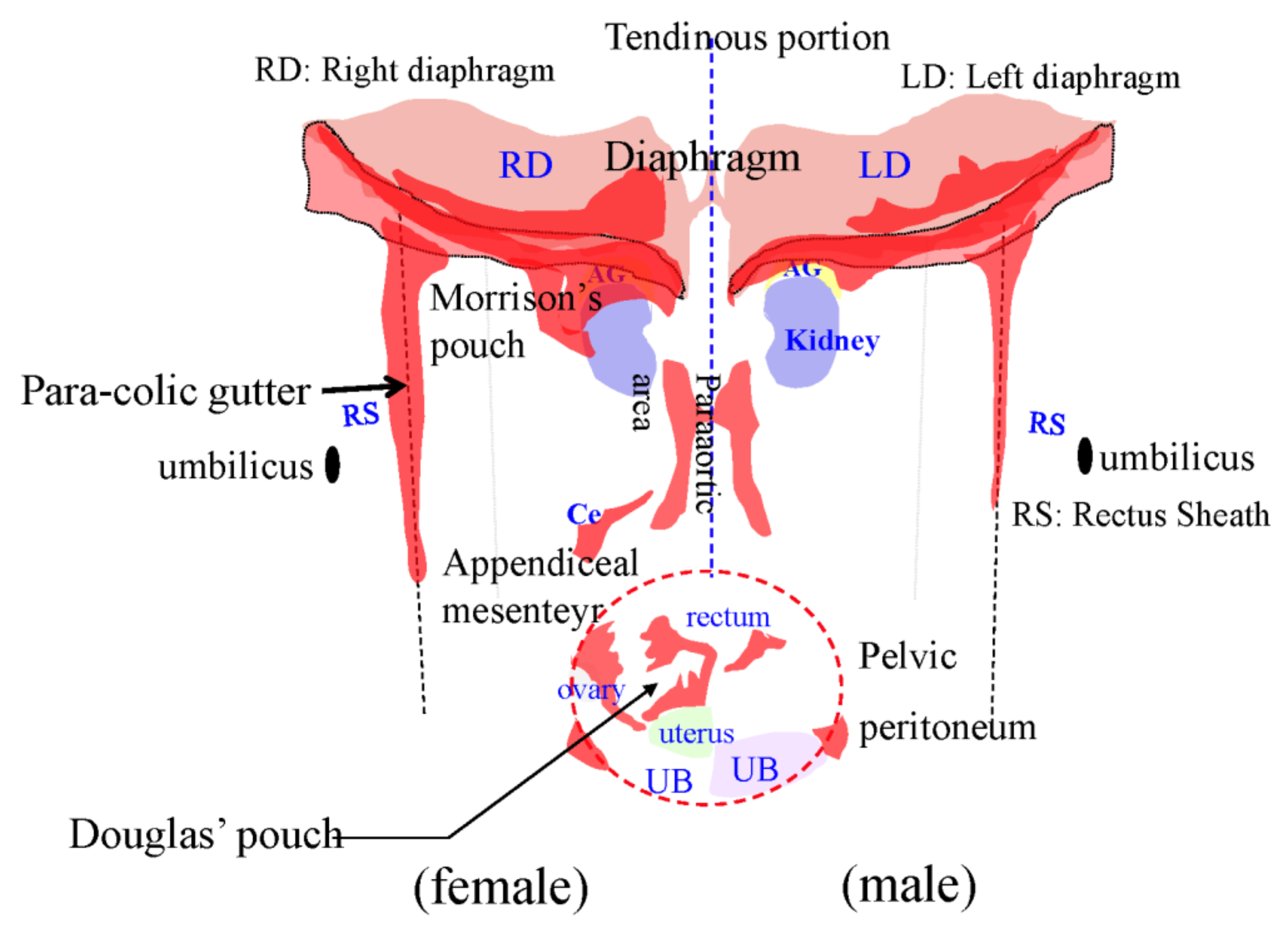
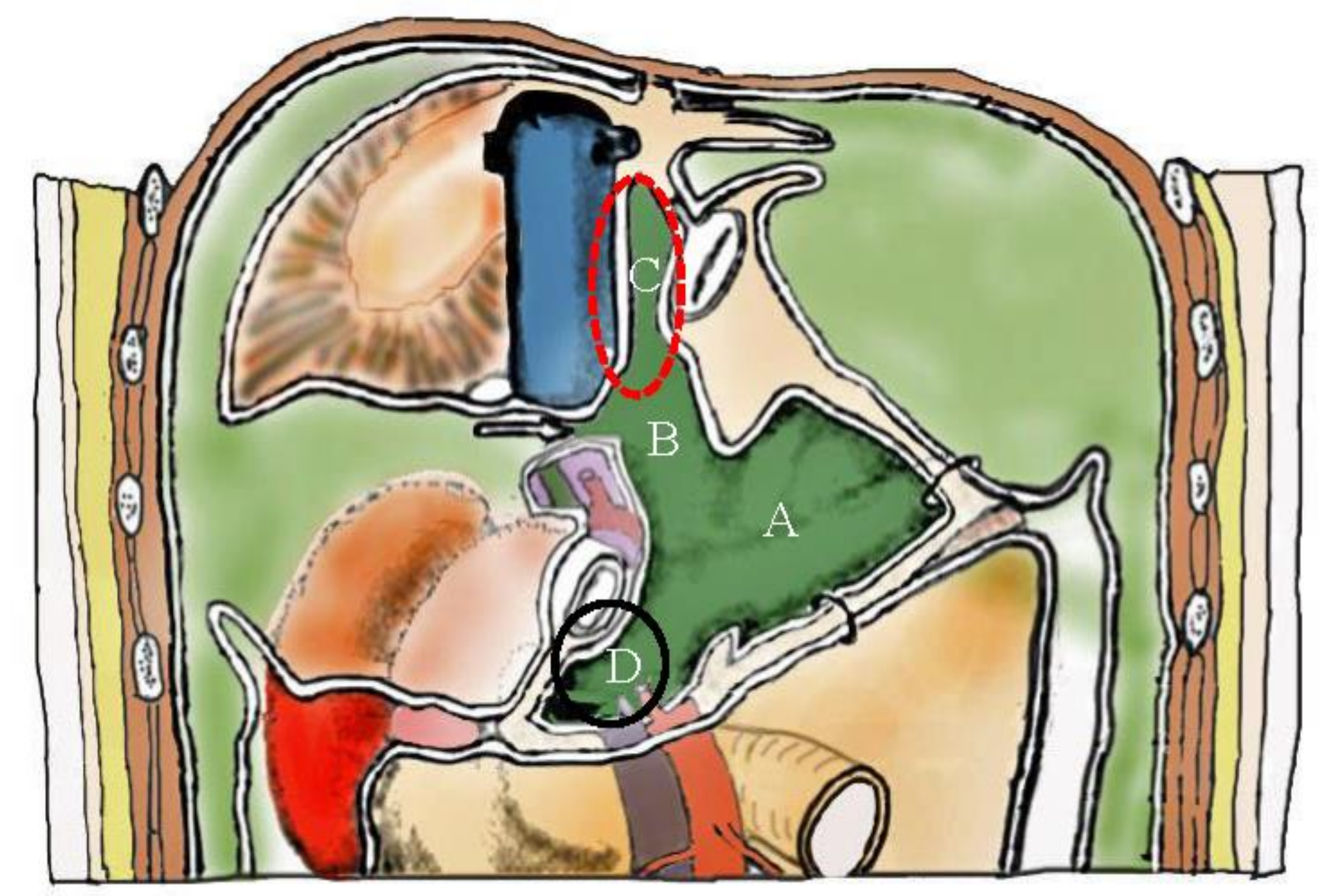
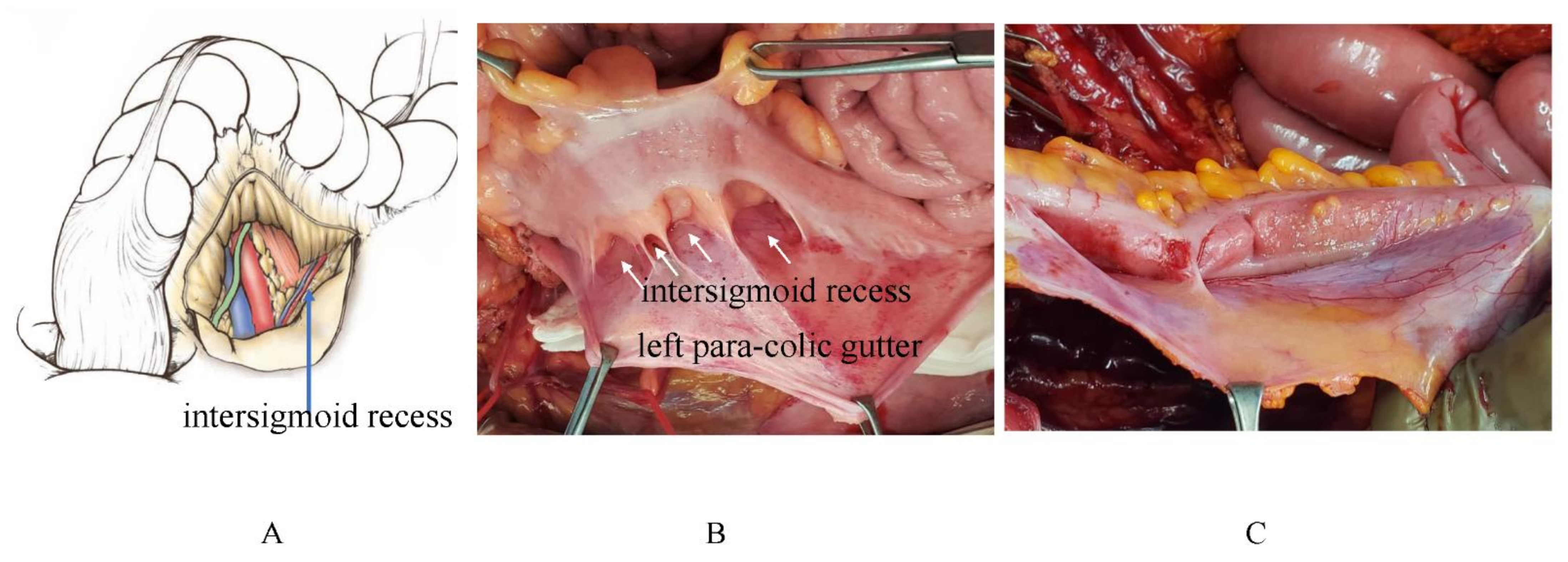
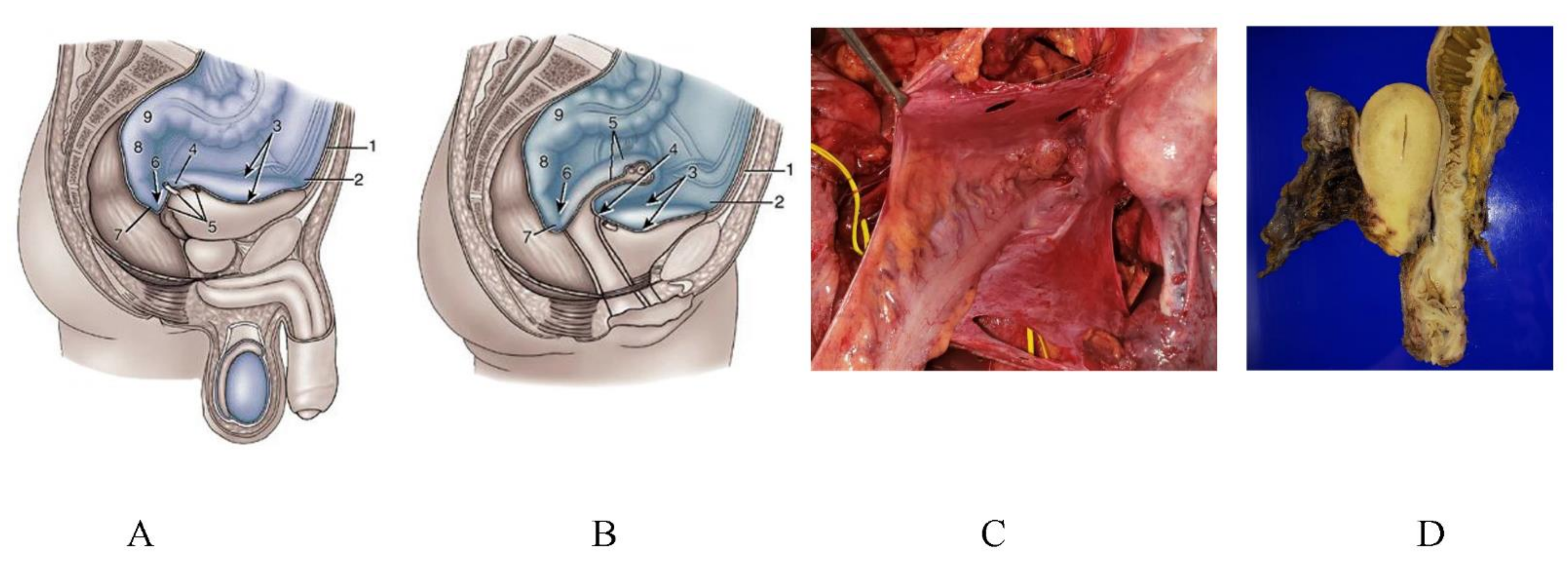
3.6. Pocket-Like Structures in the Peritoneal Cavity, Where PFCCs Are Trapped
4. Selection of the Route of Chemotherapy with Special Reference to the Mechanisms of PM formation
5. Effects of Neoadjuvant Intraperitoneal and Systemic Chemotherapy (NIPS) on Lymph Node Metastasis
6. Effects of NIPS on PM from GC
7. Effects of HIPEC on PM from GC
8. Future Perspectives
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sadeghi, B.; Arvieux, C.; Glehen, O.; Beaujard, A.C.; Rivoire, M.; Baulieux, J.; Fontaumard, E.; Brachet, A.; Caillot, J.L.; Faure, J.L.; et al. Peritoneal carcinomatosis from non gynecologic malignancies: Results of the EVOCAPE a multicentric prospective study. Cancer 2000, 88, 358–363. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, J.G.; Tian, L. Issues on peritoneal metastasis of gastric cancer: An update. World J. Surg. Oncol. 2019, 17, 215. [Google Scholar] [CrossRef] [Green Version]
- Thomassen, I.; van Gestel, Y.R.; van Ramshorst, B.; Luyer, M.D.; Bosscha, K.; Nienhuijs, S.W.; Lemmens, V.E.; de Hingh, I.H. Peritoneal carcinomatosis of gastric origin: A population based study on incidence, survival and risk factors. Int. J. Cancer 2014, 134, 622–628. [Google Scholar] [CrossRef] [PubMed]
- Sant, M.; Allemani, C.; Santaquilani, M.; Knijn, A.; Marchesi, F.; Capocaccia, R.; EUROCARE Working Group. EUOCARE-4. Survival of cancer patients diagnosed in 1995–1999. Results and commentary. Eur. J. Cancer. 2009, 45, 931–991. [Google Scholar] [CrossRef]
- Sugarbaker, P.H. Building on a consensus. J. Surg. Oncol. 2008, 98, 215–216. [Google Scholar] [CrossRef]
- Glehen, O.; Yonemura, Y.; Sugarbaker, P.H. Prevention and Treatment of Peritoneal Metastases From Gastric Cancer. In Cytoreductive Surgery & Periopertaive Chemotherapy For Peritoneal Surface Malignancy; Textbook and Video Atlas; Sugarbaker, P.H.P., Ed.; Cine-Med Publishing, Inc.: North Woodburry, CT, USA, 2013; Chapter 4; pp. 79–89. [Google Scholar]
- Yonemura, Y.; Endou, Y.; Shinbo, M.; Sasaki, T.; Hirano, M.; Mizumoto, A.; Matsuda, T.; Takao, N.; Ichinose, M.; Mizuno, M.; et al. Safety and efficacy of bidirectional chemotherapy for treatment of patients with peritoneal dissemination from gastric cancer: Selection for cytoreductive surgery. J. Surg. Oncol. 2009, 15, 311–316. [Google Scholar] [CrossRef]
- Yonemura, Y.; Bandou, E.; Sawa, T. A new treatment by neoadjuvant intraperitoneal- systemic chemotherapy and peritonectomy for peritoneal dissemination from gastric cancer. Eur. J. Surg. Oncol. 2006, 32, 661–665. [Google Scholar] [CrossRef]
- Gehen, O.; Gilly, F.N.; Cotte, E. Hyperthermic intraperitoneal chemotherapy in advanced gastric cancer: The end of skepticism. Ann. Surg. Oncol. 2011, 18, 1524–1526. [Google Scholar] [CrossRef] [PubMed]
- Yonemura, Y.; Canbay, E.; Endou, Y.; Ishibashi, H.; Mizumoto, A.; Miura, M.; Li, Y.; Liu, Y.; Takeshita, K.; Ichinose, M.; et al. Peritoneal cancer treatment. Expert Opin. 2014, 15, 623–636. [Google Scholar] [CrossRef] [PubMed]
- Yonemura, Y.; Prabhu, A.; Sako, S.; Ishibashi, H.; Mizumoto, A.; Takao, N.; Ichinose, M.; Motoi, S.; Liu, Y.; Nishihara, K.; et al. Long term survival after cytoreductive surgery combined with perioperative chemotherapy in gastric cancer patients with peritoneal metastasis. Cancers 2020, 12, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yonemura, Y. (Ed.) Comprehensive Treatment for Peritoneal Surface Malignancy with an Intent of Cure; Japanese/Asian School of Peritoneal Surface Oncology: Osaka, Japan, 2019; pp. 39–56. ISBN 978-4-9906097-5-7. [Google Scholar]
- Shimotsuma, M.; Kawata, M.; Hagiwara ATakahashi, T. Milky spots in the human greater omentum. Acta Anat. 1989, 136, 211–216. [Google Scholar] [CrossRef]
- Yonemura, Y.; Ishibashi, H.; Mizumoto, A.; Nishihara, K.; Liu, Y.; Wakama, S.; Sako, S.; Takao, N.; Ichinose, M.; Motoi, S.-I.; et al. Mechanisms of peritoneal metastasis formation. In Pathology of Peritoneal Metastasis; Glehen, O., Bhatt, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1–26. ISBN 978. [Google Scholar]
- Tsujimoto, H.; Takahashi, T.; Hagiwara, A.; Shimotsuma, M.; Sakakura, C.; Osaki, K.; Sasaki, S.; Shirasu, M.; Sakakibara, T.; Ohyama, T.; et al. Stockton Press Site-specific implantation in the milky spots of malignant cells in peritoneal dissemination: Immunohistochemical observation in mice inoculated intraperitoneally with bromodeoxyuridine-labelled cells. Brit. J. Cancer 1995, 7L, 468f–472f. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Shimoda, H.; Ji, R.; Miura, M. Lymphangiogenesis and expression of specific molecules as lymphatic endothelial cell markers. Anat. Sci. Int. 2006, 81, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Beelen, R.H.; Fluitsma, D.M.; Hoefsmit, E.C.M. The cellular composition of omental milky spots and the ultrastructure of milky spot macrophage and reticulum cells. J. Reticuloendothel. Soc. 1980, 28, 585–599. [Google Scholar] [PubMed]
- Jayne, D. Molecular Biology of Peritoneal Carcinomatosis. Cancer Treat. Res. 2007, 134, 21–31. [Google Scholar]
- Sugarbaker, P.H. Observation of concerning cancer spread within the peritoneal cavity and concept asupporting an ordered pathophysiology. In Peritoneal Carcinomatosis. A Multidisciplinary Approach; Sugarbaker, P.H., Ed.; Kluwar Academic Publisher: Boston, MA, USA; Dordrecht, The Netherlands; London, UK, 1997; pp. 79–100. [Google Scholar]
- Schneeberger, E.E.; Lynch, R.D. The tight junction: A multifunctional complex. Am. J. Physiol. Cell Physiol. 2004, 286, C1213–C1228. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Felinski, E.A.; Antonetti, D.A. Occludin phosphorylation and ubiquitination regulation tight junction trafficking and endothelial growth factor-induced permeability. J. Biol. Chem. 2009, 284, 21036–21046. [Google Scholar] [CrossRef] [Green Version]
- Kuo, W.-L.; Lee, L.-Y.; Wu, C.-M.; Wang, C.-C.; Yu, J.-S.; Liang, Y.; Lo, C.-H.; Huang, K.-H.; Hwang, T.-L. Differential expression of claudin-4 between intestinal and diffuse-type gastric cancer. Oncol. Rep. 2005, 16, 729–734. [Google Scholar] [CrossRef]
- Ohtani, S.; Terashima, M.; Satoh, J.; Soeta, N.; Saze, Z.; Kashimura, S.; Ohsuka, F.; Hoshino, Y.; Kogure, M.; Gotoh, M. Expression of tight-junction-associated proteins in human gastric cancer. Downregulation of claudin-4 correlates with tumor aggressiveness and survival. Gastric Cancer 2009, 12, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Shimoyama, Y.; Hirohashi, S. Expression of E- and P-cadherin in gastric carcinoma. Cancer Res. 1991, 51, 2185–2192. [Google Scholar] [PubMed]
- Yasui, W.; Kuniyasu, H.; Akama, Y.; Kitahara, K.; Nagafuchi, A.; Ishihara, S.; Tsukita, S.; Tahara, E. Expression of E-cadherin and -catenins in human gastric carcinoma: Correlation with histology and tumor progression. Oncol. Rep. 1995, 2, 111–117. [Google Scholar] [PubMed]
- Yasui, W.; Sano, T.; Nishimura, K.; Kitadai, Y.; Ji, Z.-Q.; Yokozaki, H.; Ito, H.; Tahara, E. Expression of P-cadherin in gastric carcinomas and its reduction in tumor progression. Int. J. Cancer 1993, 54, 49–52. [Google Scholar] [CrossRef]
- Gooding, J.M.; Yap, K.L.; Ikura, M. The cadherin-catenin complex as a focal point of cell adhesion and signaling: New insights from three-dimensional structures. BioEssays 2004, 26, 497–511. [Google Scholar] [CrossRef] [PubMed]
- Halbleib, J.M.; Nelson, W.J. Cadherins in development: Cell adhesion, sorting, and tissue morphogenesis. Genes Dev. 2006, 20, 3199–3214. [Google Scholar] [CrossRef] [Green Version]
- Van Marck, V.; Stove, C.; Bossche, K.V.D.; Paredes, J.; Haeghen, Y.V.; Bracke, M. P-cadherin promotes cell-cell adhesion and counteracts invasion in human melanoma. Cancer Res. 2005, 65, 8774–8783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dangerfield, J.; Larbi, K.Y.; Huang, M.T.; Dewar, A.; Nourshargh, S. PECAM-1 (CD31) homophilic interaction up-regulates alpha 6 beta 1 on transmigrated neutrophils in vivo and plays a functional role in the ability of alpha 6 integrins to mediate leukocyte migration through the perivascular basement membrane. J. Exp. Med. 2002, 196, 1201–1211. [Google Scholar] [CrossRef] [PubMed]
- Hirano, H.; Screaton, G.R.; Bell, M.V.; Jackson, D.G.; Bell, J.J.; Hodes, R.J. CD44 isoform expression mediated by alternative splicing: Tissue-specific regulation in mice. Int. Immunol. 1994, 6, 49–59. [Google Scholar] [CrossRef]
- Harada, N.; Mizoi, T.; Kinouchi, M.; Hoshi, K.; Ishii, S.; Shiiba, K.I.; Sasaki, I.; Matsuno, S. Introduction of antisense CD44 CDNA down- regulates expression of overall CD44 isoforms and inhibits tumor growth and metastasis in highly metastatic colon cancer carcinoma cells. Int. J. Cancer 2001, 91, 67–75. [Google Scholar] [CrossRef]
- Nakamori, S.; Furukawa, H.; Hiratsuka, M.; Iwanaga, T.; Imaoka, S.; Ishikawa, O.; Kabuto, T.; Sasaki, Y.; Kameyama, M.; Ishiguro, S.; et al. Expression of carbohydrate antigen sialyl Le(a): A new functional prognostic factor in gastric cancer. J. Clin. Oncol. 1997, 15, 816–825. [Google Scholar] [CrossRef]
- Sanusi, A.A.; Zweers, M.M.; Weening, J.J.; De Waart, D.R.; Struijk, D.G.; Krediet, R.T. Expression of cancer antigen by peritoneal mesothelial cells is not influenced by duration of peritoneal dialysis. Perit. Dial. Int. 2001, 21, 495–500. [Google Scholar] [CrossRef]
- Scholler, N.; Urban, N. CA125 in ovarian cancer. Biomark. Med. 2007, 1, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Hassan, R.; Ho, M. Mesothelin targeted immunotherapy. Eur. J. Cancer 2008, 44, 46–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arita, T.; Ichikawa, D.; Konishi, H.; Komatsu, S.; Shiozaki, A.; Ogino, S.; Fujita, Y.; Hiramoto, H.; Hamada, J.; Shoda, K.; et al. Tumor exosome-mediated promotion of adhesion to mesothelial cells in gastric cancer cells. Oncotarget 2021, 7, 56855–56863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedli, A.; Fischer, E.; Novak-Hofer, I.; Cohrs, S.; Ballmer-Hofer, K.; Schubiger, P.A.; Schibli, R.; Grünberg, J. The soluble form of the cancer-associated L1 cell adhesion molecules is a pro-angiogenic factor. Int. J. Biochem. Cell Biol. 2009, 41, 1572–1580. [Google Scholar] [CrossRef] [PubMed]
- Bird, S.D. Mesothelial primary cilia of peritoneal other serosal surface. Cell Biol. Int. 2004, 28, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Yao, V.; Plattell, C.; Hall, J.C. Peritoneal mesothelial cells inflammatory related cytokines. N. Z. J. Surg. 2004, 74, 997–1002. [Google Scholar] [CrossRef]
- Akedo, H.; Shinkai, K.; Mukai, M.; Mori, Y.; Tateishi, R.; Tanaka, K.; Yamamoto, R.; Morishita, T. Interaction of rat ascites hepatoma cells with cultured mesothelial cell layers: A model for tumor invasion. Cancer Res. 1986, 46, 2416–2422. [Google Scholar]
- Jayne, D.G.; O’leary, R.; Gill, A.; Hick, A.; Guillou, P.J. A three-dimensional in- vitro model for the study of peritoneal tumor metastasis. Clin. Exp. Metastasis 1999, 17, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Lanfrancone, L.; Boraschi, D.; Ghiara, P.; Falini, B.; Grignani, F.; Peri, G.; Mantovani, A.; Pelicci, P.G. Human peritoneal mesothelial cells produce many cytokines (granulocyte colony-stimulating factor (CSF), granulocyte- monocyte-CSF, macrophage-CSF, interleukin-1, and IL-6 and are activated and stimulated to grow by IL-1. Blood 1992, 80, 2835–2842. [Google Scholar] [CrossRef] [Green Version]
- Hight-Warburton, W.; Parsons, M. Regulation of cell migration by α4 and α9 integrins. Biochem. J. 2019, 476, 705–718. [Google Scholar] [CrossRef] [PubMed]
- Yonemura, Y.; Endou, Y.; Yamaguchi, T.; Nojima, N.; Kawamura, T.; Fujimura, T.; Obata, T.; Kim, B.; Miyazaki, I.; Sasaki, T. Roles of VLA-2 and VLA-3 on the formation of peritoneal dissemination in gastric cancer. Int. J. Oncol. 1996, 8, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, T.; Endo, Y.; Yonemura, Y.; Nojima, N.; Fujita, H.; Fujimura, T.; Obata, T.; Yamaguchi, T.; Sasaki, T. Significance of integrin alpha2/beta1 in peritoneal dissemination of a human gastric cancer xenograft model. Int. J. Oncol. 2001, 18, 809–815. [Google Scholar] [PubMed]
- Takatsuki, H.; Komatsu, S.; Sano, R.; Takada, Y.; Tsuji, T. Adhesion of gastric carcinoma cells in perioneum mediated by α3β1 integrin (VLA-3). Cancer Res. 2004, 64, 6065–6070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemler, M.E.; Elices, M.J.; Chan, B.M.; Zetter, B.; Matsuura, N.; Takada, Y. Multiple ligand binding functions for VLA-2 (α 2 β 1) and VLA-3 (α 3 β 1) in the integrin family. Cell Differ. Dev. 1990, 1232, 229–238. [Google Scholar] [CrossRef]
- Diaz-Flores, L.; Gutierrez, R.; Garcia, M.P.; Sáez, F.J.; Valladares, F.; Madrid, J.F. CD34+ stromal cells/fibroblasts/fibrocysts/telocytes as a tissue reserve and a principal source of mesenchymal cells. Location, morphology, function and role of pathology. Histol. Pathol. 2016, 29, 831–870. [Google Scholar]
- Pan, Y.; Bi, F.; Liu, N.A.; Xue, Y.; Yao, X.; Zheng, Y.; Fan, D. Expression of seven main Rho family members in gastric carcinoma. Biochem. Biophys. Res. Commun. 2004, 315, 686–691. [Google Scholar] [CrossRef]
- Selinfreund, R.H.; Barger, S.W.; Welsh, M.J.; Van Eldik, L.J. Antisense inhibition of glial S100 beta production results in alteration in cell morphology, cytoskeletal organization, and cell proliferation. J. Cell Biol. 1990, 111, 2021–2028. [Google Scholar] [CrossRef]
- Kim, E.J.; Helman, D.M. Characterization of the metastasis-associated protein, S100A4. Roles of calcium binding and dimerization in cellular localization and interaction with myosin. J. Biol. Chem. 2003, 278, 30063–30073. [Google Scholar] [CrossRef] [Green Version]
- Yonemura, Y.; Endou, Y.; Kimura, K.; Fushida, S.; Bandou, E.; Taniguchi, K.; Kinoshita, K.; Ninomiya, I.; Sugiyama, K.; Heizmann, C.W.; et al. Inverse expression of S100A4 and E-cadherin is associated with metastatic potential in gastric cancer. Clin. Cancer Res. 2000, 6, 4234–4242. [Google Scholar]
- Moriyama-Kita, M.; Endo, Y.; Yonemura, Y.; Heizmann, C.W.; Miyamori, H.; Sato, H.; Yamamoto, E.; Sasaki, T. S100A4 regulates E-cadherin expression in oral squamous cell carcinoma. Cancer Lett. 2005, 230, 211–218. [Google Scholar] [CrossRef]
- Warn, R.; Harvey, P.; Warn, A.; Foley-Comer, A.; Heldin, P.; Versnel, M.; Arakaki, N.; Daikuhara, Y.; Laurent, G.J.; Herrick, S.; et al. HGF/SF induces mesothelial cell migration and proliferation by autocrine paracrine pathways. Exp. Cell Res. 2001, 267, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Yoshioka, K.; Akedo, H.; Uehata, M.; Ishizaki, T.; Narumiya, S. An essential part for rho-associated linease in the transcellular invasion of tumor cells. Nat. Med. 1999, 5, 221–225. [Google Scholar] [CrossRef]
- Taniguchi, K.; Yonemura, Y.; Nojima, N.; Hirono, Y.; Fushida, S.; Fujimura, T.; Miwa, K.; Endo, Y.; Yamamoto, H.; Watanabe, H. The relation between the growth patterns of gastric carcinoma and the expression of hepatocyte growth factor receptor (c-met). Autocrine motility factor receptor and urokinase-type plasminogen receptor. Cancer 1998, 82, 2112–2122. [Google Scholar] [CrossRef]
- Ito, H.; Yonemura, Y.; Fujita, H.; Tsuchihara, K.; Kawamura, T.; Nojima, N.; Fujimura, T.; Nose, H.; Endo, Y.; Sasaki, T. Prognostic relevance of urokinase-type plasminogen activator system and plasminogen activator PAI-1 and PAI-2 in gastric cancer. Virchows Arch. 1996, 427, 487–496. [Google Scholar] [CrossRef]
- Poincloux, R.; Lizarraga, F.; Chavrier, P. Matrix invasion by tumor cells: A focus on MT1- MMP trafficking to invadopodia. J. Cell Sci. 2009, 122, 3015–3024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, H.; Takino, T.; Okada, Y.; Cao, J.; Shinagawa, A.; Yamamoto, E.; Seiki, M. A matrix metalloproteinase expressed on the surface of invasive tumor cells. Nature 1994, 370, 61–65. [Google Scholar] [CrossRef]
- Itoh, Y.; Ito, N.; Nagase, H.; Seiki, M. The second dimer interface of MT1-MMP, the transmembrane domain, is essential for proMMP-2 activation on the cell surface. J. Biol. Chem. 2008, 283, 13053–13062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, M.; Yasui, W.; Nakayama, H.; Yokozaki, H.; Ito, H.; Tahara, E. Reduced levels of transforming growth factor- beta type I receptor in human gastric carcinomas. Jpn. J. Cancer Res. 1992, 83, 86–92. [Google Scholar] [CrossRef]
- Stetler-Stevenson, W.G.; Aznavoorian, S.; Liotta, L.A. Tumor cell interaction with the extracellular matrix during invasion and metastasis. Ann. Rev. Cell Biol. 1993, 9, 541–573. [Google Scholar] [CrossRef] [PubMed]
- Bando, E.; Yonemura, Y.; Endou, Y.; Sasaki, T.; Taniguchi, K.; Fujita, H.; Fushida, S.; Fujimura, T.; Nishimura, G.; Miwa, K.; et al. Immunohistochemical study of MT1-MMP tissue status in gastric carcinoma and correlation with survival analyzed by univariate and multivariate analysis. Oncol. Rep. 1998, 5, 1483–1488. [Google Scholar] [PubMed]
- Takeno, A.; Takemasa, I.; Doki, Y.; Yamasaki, M.; Miyata, H.; Takiguchi, S.; Fujiwara, Y.; Matsubara, K.; Monden, M. Integrative approach for differentially overexpressed genes in gastric cancers by combining large-scale expression profiling and network analysis. Br. J. Cancer 2008, 21, 1307–1315. [Google Scholar] [CrossRef] [Green Version]
- Yonemura, Y.; Endou, Y.; Fujita, H.; Fushida, S.; Bandou, E.; Taniguchi, K.; Miwa, K.; Sugiyama, K.; Sasaki, T. Role of MMP-7 on the formation of peritoneal dissemination in gastric cancer. Gastric Cancer 2000, 3, 63–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yonemura, Y.; Endo, Y.; Fujita, H.; Kimura, K.; Sugiyama, K.; Momiyama, N.; Shimada, H.; Sasaki, T. Inhibition of peritoneal dissemination in human gastric cancer by MMP-7-specific antisense oligonucleotide. J. Exp. Clin. Cancer Res. 2001, 20, 205–212. [Google Scholar]
- Elnemr, A.; Yonemura, Y.; Bandou, E.; Kinoshita, K.; Kawamura, T.; Takahashi, S.; Tochiori, S.; Endou, Y.; Sasaki, T. Expression of collagenase-3 (matrix metalloproteinase-13) in human gastric cancer. Gastric Cancer 2003, 6, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Del Casar Lizcano, J.M.; Vizoso Pineito, F.; Gonzalez Sanchez, L.O. Expression and clinical significance of collagenase-3 (MMP-13) in gastric cancer. Gastroenterol. Hepatol. 2003, 26, 1–7. [Google Scholar] [CrossRef]
- Casey, R.C.; Skubitz, A.P. CD44 and beta1 integrins mediate ovarian carcinoma cell migration toward extracellular matrix proteins. Clin. Exp. Metastasis 2000, 18, 67–75. [Google Scholar] [CrossRef]
- Moghaddam, S.M.; Amini, A.; Morris, D.L.; Pourgholami, M.H. Significance of vascular endothelial growth factor in growth and peritoneal dissemination of ovarian cancer. Cancer Metastasis Rev. 2012, 31, 143–162. [Google Scholar] [CrossRef] [Green Version]
- Du, S.; Yang, Z.; Lu, X.; Yousuf, S.; Zhao, M.; Li, W.; Miao, J.; Wang, X.; Yu, H.; Zhu, X.; et al. Anoikis resistant gastric cancer cells promote angiogenesis and peritoneal metastasis through C/EBPb-mediated PDGFB autocrine and paracrine signaling. Oncogene 2021, 40, 5764–5779. [Google Scholar] [CrossRef]
- Ke, Q.; Costa, M. Hypoxia-inducible factor-1 (HIF-1). Mol. Pharmacol. 2006, 70, 1469–1480. [Google Scholar] [CrossRef] [PubMed]
- Thurston, Q. Role of angiopoietins and Tie receptor tyrosin kinase in angiogenesis and lymphangiogenesis. Cell Tissue Res. 2003, 314, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.K. Barrier to drug delivery in solid tumors. Sci. Am. 1994, 7, 42–49. [Google Scholar] [CrossRef]
- Shimotsuma, M.; Takahashi, T.; Kawata, M.; Dux, K. Cellular subset of the milky spots in the human greater omentum. Cell Tissue Res. 1991, 264, 599–601. [Google Scholar] [CrossRef]
- Van der Speeten, K. Pharmacokinetics and pharmacodynamics of peritoneal cancer chemotherapy in peritoneal surface malignancy. Cancer J. 2009, 15, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Fushida, S.; Kinoshita, J.; Yagi, Y.; Funaki, H.; Kinami, S.; Ninomiya, I.; Fujimura, T.; Nishimura, G.; Kayahara, M.; Ohta, T. Dual anticancer effects of weekly intraperitoneal docetaxel in treatment of gastric cancer patients with peritoneal carcinomatosis: A feasibility pharmacokinetic study. Oncol. Rep. 2008, 19, 1305–1310. [Google Scholar] [PubMed] [Green Version]
- Yonemura, Y. Pharmacokinetics of docetaxel during hyperthermic intraperitoneal chemotherapy for peritoneal metastasis. Gan Kagaku Ryoho 2014, 41, 2496–2499. [Google Scholar]
- Yonemura, Y. Pharmacokinetics during hyperthermo chemo-perfusion. In Comprehensive Treatment for Peritoneal Surface Malignancy with an Intent of Cure; Yonemura, Y., Ed.; NPO to Support Treatment of Peritoneal Surface Malignancy; NPO: Hilversum, The Netherlands, 2019; pp. 17–20. ISBN 978-4-9906097-2-6. [Google Scholar]
- Los, G. penetration of carboplatin and cisplatininto rat peritoneal nodules after intraperitoneal chemotherapy. Cancer Chmother. Pharmacol. 1991, 28, 159–165. [Google Scholar] [CrossRef]
- Hao, Y.; Liu, Y.; Ishibashi, H.; Wakama, S.; Nishino, E.; Yonemura, Y. Downstaging of lymph node metastasis after neoadjuvant intraperitoneal and systemic chemotherapy in gastric carcinoma with peritoneal metastasis. Eur. J. Surg. Oncol. 2019, 45, 1493–1497. [Google Scholar] [CrossRef]
- Ishigami, H.; Fujiwara, Y.; Fukushima, R.; Nashimoto, A.; Yabusaki, H.; Imano, M.; Imamoto, H.; Kodera, Y.; Uenosono, Y.; Amagai, K.; et al. Phase III Trial Comparing Intraperitoneal and Intravenous Paclitaxel Plus S-1 Versus Cisplatin Plus S-1 in Patients with Gastric Cancer with Peritoneal Metastasis: PHOENIX-GC Trial. J. Clin. Oncol. 2018, 36, 1922–1929. [Google Scholar] [CrossRef] [PubMed]
- Yonemura, Y.; Canbay, E.; Fujita, T.; Sako, S.; Wakama, S.; Ishibashi, H.; Hirano, M.; Mizumoto, A.; Takeshita, K.; Takao, N.; et al. Effects of neoadjuvant laparoscopic hyperthermic intraperitoneal chemoperfusion and intraperitoneal/systemic chemotherapy on peritoneal metastasis from gastric cancer. J. Peritoneum Other Serosal Surf. 2017, 24, 478–485. Available online: http://www.jperitoneum.org/index.php/joper/article/view/60 (accessed on 31 July 2017).
- Brandl, A.; Yonemura, Y.; Glehen, O.; Sugarbaker, P.; Rau, B. Long term survival in patients with peritoneal metastasised gastric cancer treated with cytoreductive surgery and HIPEC: A multi- institutional cohort from PSOGI. Eur. J. Surg. Oncol. 2021, 47, 172–180. [Google Scholar] [CrossRef]
- Yang, X.-J.; Huang, C.-Q.; Suo, T.; Mei, L.-J.; Yang, G.-L.; Cheng, F.-L.; Zhou, Y.-F.; Xiong, B.; Yonemura, Y.; Li, Y. Cytoreductive surgery and hyperthermic intraperitoneal chemotherapy improves survival of patients with peritoneal carcinomatosis from gastric cancer: Final results of a phase III randomized clinical trial. Ann. Surg. Oncol. 2011, 18, 1575–1581. [Google Scholar] [CrossRef] [Green Version]
- Thadi, A.; Khalili, M.; Morano, W.F.; Richard, S.D.; Katz, S.C.; Bowne, W.B. Erratum: Thadi A.; et al. Early investigations and recent advances in intraperitoneal immunotherapy for peritoneal metastasis. Vaccines 2019, 7, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knödler, M.; Körfer, J.; Kunzmann, V.; Trojan, J.; Daum, S.; Schenk, M.; Kullmann, F.; Schroll, S.; Behringer, D.; Stahl, M.; et al. Randomised phase II trial to investigate catumaxomab (anti-EpCAM anti-CD3) for treatment of peritoneal carcinomatosis in patients with gastric cancer. Br. J. Cancer 2018, 119, 296–302. [Google Scholar] [CrossRef]
- Hagi, T.; Kurokawa, Y.; Kawabata, R.; Omori, T.; Matsuyama, J.; Fujitani, K.; Hirao, M.; Akamaru, Y.; Takahashi, T.; Yamasaki, M.; et al. Multicentre biomarker cohort study on the efficacy of nivolumab treatment forgastric cancer. Br. J. Cancer 2020, 123, 965–972. [Google Scholar] [CrossRef]
- Ishikawa, M.; Iwasa, S.; Nagashima, K.; Aoki, M.; Imazeki, H.; Hirano, H.; Shoji, H.; Honma, Y.; Okita, N.; Takashima, A.; et al. Retrospective comparison of nab-paclitaxel plus ramucirumab and paclitaxel plus ramucirumab as second-line treatment for advanced gastric cancer focusing on peritoneal metastasis. Investig. New Drugs 2020, 38, 533–540. [Google Scholar] [CrossRef]
- Alyami, M.; Hübner, M.; Grass, F.; Bakrin, N.; Villeneuve, L.; Laplace, N.; Passot, G.; Glehen, O.; Kepenekian, V. Pressurised intraperitoneal aerosol chemotherapy: Rationale, evidence, and potential indications. Lancet Oncol. 2019, 20, e368–e377. [Google Scholar] [CrossRef]
- Zhou, W.; Dai, S.; Zhu, H.; Song, Z.; Cai, Y.; Lee, J.B.; Li, Z.; Hu, X.; Fang, B.; He, C.; et al. Telomerase-specific oncolytic adenovirus expressing TRAIL suppresses peritoneal dissemination of gastric cancer. Gene Ther. 2017, 24, 199–207. [Google Scholar] [CrossRef] [PubMed]































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drugs | MW | pAUC/sAUC | Penetratoin Distance | MTD | Thermal Enhancement |
|---|---|---|---|---|---|
| Doxorubicin | 380 | 230 | 4–6 cell layer | 15 mg/m2 | Yes |
| Melphalan | 305 | 93 | NA | 70 mg/m2 | Marked |
| Mitomycin C | 334 | 32.5 | NA | 35 mg/m2 | Yes |
| Cisplatin | 300 | 7.8 | 1–2 mm | 300 mg/m2 | Yes |
| Gemcitabine | 299 | 500 | NA | 1000 mg/m2 | at 48 h |
| Miroxantron | 517 | 115–255 | 5–6 cell layer | 28 mg/m2 | Yes |
| Oxaliplatin | 387 | 16 | 1–2 mm | 460 mg/m2 | Yes |
| Etoposide | 568 | 63 | NA | 200 mg/m2 | Yes |
| Irrinotecan | 677 | NA | NA | NA | No |
| Paclitaxel | 853 | 10,000 | 80 cell layer | 120–180 mg/m2 | No |
| Docetaxel | 861 | 552 | 1.4 mm | 156 mg/m2 | Yes |
| 5-FU | 130 | 250 | 0.2 mm | 650 mg/m2 × 5 days | Yes |
| carboplatin | 371 | 10 | 0.5 mm | 300 mg/m2 | Yes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yonemura, Y.; Ishibashi, H.; Mizumoto, A.; Tukiyama, G.; Liu, Y.; Wakama, S.; Sako, S.; Takao, N.; Kitai, T.; Katayama, K.; et al. The Development of Peritoneal Metastasis from Gastric Cancer and Rationale of Treatment According to the Mechanism. J. Clin. Med. 2022, 11, 458. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm11020458
Yonemura Y, Ishibashi H, Mizumoto A, Tukiyama G, Liu Y, Wakama S, Sako S, Takao N, Kitai T, Katayama K, et al. The Development of Peritoneal Metastasis from Gastric Cancer and Rationale of Treatment According to the Mechanism. Journal of Clinical Medicine. 2022; 11(2):458. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm11020458
Chicago/Turabian StyleYonemura, Yutaka, Haruaki Ishibashi, Akiyoshi Mizumoto, Gorou Tukiyama, Yang Liu, Satoshi Wakama, Shouzou Sako, Nobuyuki Takao, Toshiyuki Kitai, Kanji Katayama, and et al. 2022. "The Development of Peritoneal Metastasis from Gastric Cancer and Rationale of Treatment According to the Mechanism" Journal of Clinical Medicine 11, no. 2: 458. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm11020458