NK Cell Plasticity in Cancer


1
Department of Urology, Feinberg School of Medicine, Northwestern University, Chicago, IL 60611, USA
2
Robert Lurie Comprehensive Cancer Center, Chicago, IL 60611, USA
*
Author to whom correspondence should be addressed.
†
Authors contribute equally.
J. Clin. Med. 2019, 8(9), 1492; https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8091492
Submission received: 20 August 2019
/
Revised: 11 September 2019
/
Accepted: 15 September 2019
/
Published: 19 September 2019
(This article belongs to the Special Issue Development and Function of Natural Killer Cells and their Importance in Immunotherapy)
Abstract
:Natural killer (NK) cells are critical immune components in controlling tumor growth and dissemination. Given their innate capacity to eliminate tumor cells without prior sensitization, NK-based therapies for cancer are actively pursued pre-clinically and clinically. However, recent data suggest that tumors could induce functional alterations in NK cells, polarizing them to tumor-promoting phenotypes. The potential functional plasticity of NK cells in the context of tumors could lead to undesirable outcomes of NK-cell based therapies. In this review, we will summarize to-date evidence of tumor-associated NK cell plasticity and provide our insights for future investigations and therapy development.
1. Introduction
Natural killer (NK) cells play important roles in anti-tumor immunosurveillance. Using an array of germline-encoded activating and inhibitory receptors, NK cells are able to detect and kill malignant cells without prior sensitization by sensing the altered surface ligand repertoire on tumor cells [1]. NK cells recognize tumor cells via two major mechanisms [1,2]. One is through the “missing-self” mechanism, whereby NK cells recognize tumor cells that have downregulated self major histocompatibility complex I (MHC I) molecule. In the healthy physiological state, NK cells express an array of inhibitory receptors, such as killer cell inhibitory receptors (KIRs) and CD94/NKG2A complex, that interact with major histocompatibility complex I (MHC I) molecule to deliver an inhibitory signal to NK cells and sustain NK cells in a quiescent state. Tumor cells often lose or have downregulated MHC I expression, which in turn diminishes the inhibitory signals in NK cells and leads to NK cell activation. The second is through the “induced-self” mechanism, whereby NK cell activating receptors engage their ligands “induced” or overexpressed on tumor cells. Ligands for NK cell activating receptors, such as NKG2D, natural cytotoxicity receptors (NCRs), and DNAM-1 (DNAX accessory molecule), are generally absent in normal tissues, but induced on tumor cells in response to various oncogenic stress [1,2]. In the context of therapeutic antibodies, NK cells can also recognize tumor cells through engaging CD16, the receptor for Fc region of certain isotype of antibodies, to mediate antibody-dependent cellular cytotoxicity (ADCC) against tumor cells [1]. NK cells are also an important component in shaping adaptive immune responses through secreting an array of cytokines, such as IFN-γ, TNF-a, GM-CSF, and interleukin (IL)-10, and interacting with dendritic cells [3,4,5,6,7].
A number of clinical studies have demonstrated the significance of NK cells in controlling cancer. A large-scale, 11-year follow up study showed that higher NK activity correlates with lower cancer risk [8]. Early clinical trials of NK adoptive transfer in acute myeloid leukemia (AML) patients have presented a beneficial, although limited role in controlling the disease [9,10]. In the recent IMMUNOSTIM study (NCT00478985), a significantly higher count of CD56dim cytotoxic NK cells was found in non-relapsing as opposed to relapsing chronic myeloid leukemia (CML) patients after imatinib (a tyrosine kinase inhibitor, TKI) discontinuation [11]. In two other independent multi-center clinical trials (DADI trial and EURO-SKI, NCT01596114) in CML patients, a higher rate of disease remission after TKI discontinuation was associated with a higher percentage of activated NK cells after TKI discontinuation [12,13]. In patients with colorectal cancer, gastric cancer, and lung cancer, NK infiltration is associated with better outcomes [14,15,16,17]. High NK cell activity also correlated with better clinical response with increased progression-free survival (PFS) to trastuzumab in metastatic breast cancer patients and delayed progression to castration-resistant diseases in prostate cancer patients [18,19]. A gene signature correlating with increased NK infiltration was associated with improved survival in melanoma patients [6,20]. These studies support the significance of NK cells in controlling cancer. NK cells are thus considered an attractive target for immunotherapies. However, a highly immune suppressive tumor microenvironment can alter NK cell phenotype and function and result in their inability to effectively exert their anti-tumor activity. Moreover, a number of recent studies have presented the potential pro-tumorigenic activities of NK cells, owing to their functional plasticity in the context tumor. With many ongoing endeavors in developing NK cell-based therapies clinically and preclinically, it is critical to understand these various aspects of NK cells. In the following sections, we will summarize current clinical efforts for NK cell-based therapies and recent evidence of tumor-induced NK cell plasticity, with a focus on tumor microenvironment (TME)-induced NK cell phenotypical and functional alternations.
2. Current NK Cell-Based Pre-Clinical and Clinical Studies
NK cell-based therapies have been evaluated in clinical trials [21,22,23]. To date, two main types of NK-based therapies have been pursued: (i) adoptive transfer of NK-92 cell line and cord blood derived-NK cells expanded and activated ex vivo; (ii) use of engineered chimeric antigen receptor (CAR)-NK cells. Immune-stimulatory cytokines, such as IL-2 and IL-15, have been used in the therapeutic settings to potentiate NK cell survival and function. NK cell-based therapies generally presented good safety profiles in clinic. How to achieve desired therapeutic efficacy of NK-cell based therapies is still a challenge. Thus far, the most effective indications for NK cell-based therapies are limited to hematological malignancies. Both pre-clinical and clinical studies have shown that NK cell-based therapies can promote graft-versus-leukemia or graft-versus-tumor effect, without provoking graft-versus-host disease (GvHD) [9,24,25,26]. To improve the therapeutic efficacy of NK cell-based therapies, Ruggeri et al. have pioneered allogenic haploidentical NK cell transfer by selecting patients with KIR ligand mismatch to allow alloreactive NK cell activation. This approach presented to be safe and provoked protection against acute myeloid leukemia (AML) relapse [27]. Miller and colleagues further explored this haploidentical NK cell transfer strategy, along with immune suppressive cyclophosphamide and fludarabine regimens in patients with metastatic melanoma, Hodgkin’s disease, renal cell carcinoma, and poor-prognosis AML [9]. The strategy is generally safe and achieved a good in-patient NK cell expansion. However, efficacy was only presented in AML patients after receiving intensive immunosuppressive regimen of cyclophosphamide and fludarabine, but not in patients receiving low intensity immunosuppressive regimens [9]. Five out of 19 acute myeloid leukemia (AML) patients attained complete remission; however, high doses of interleukin-2 (IL-2) and high conditioning regimens triggered severe hematological as well as non-hematological toxicities in the patients [9]. To address such toxicity issues and to evaluate the suitability of low-intensity immunosuppressive regimens, Rubnitz and group conducted a pilot study (NCT00187096) in children with AML. The study concluded that the low-dose immunosuppressive regimens in patients receiving donor-recipient KIR-HLA mismatched NK cells were safe and feasible [24]. These initial studies generated a great interest in pursuing NK cell-based cancer immunotherapy.
The outcome of clinical trials using NK cell-based therapies for solid tumors has been less optimistic than for hematological malignancies. Although phase I clinical trials demonstrated the safety of adoptively transferred NK cells [28,29,30,31,32], clinical efficacy reported to date is limited. A phase II study (NCT00376805) conducted by Miller and group using adoptively transferred allogenic NK cells to treat patients with recurrent ovarian and breast cancer demonstrated limited efficacy [33]. In this study, 20 patients underwent fludarabine and cyclophosphamide regimen with or without low-dose total body irradiation (TBI) before NK cell infusion. Although donor DNA was detectable after seven days of NK cell infusion in 69% of the patients without TBI and 85% of the patients with TBI, the infused cells did not persist beyond seven days. There was an undesirable expansion of immunosuppressive regulatory T cells (Tregs) in response to low dose IL-2 (10 million unit/dose, 3 times per week, 6 doses in total) that was given in conjunction with NK cell infusion [33]. To avoid IL-2 associated Treg expansion, IL-15 is now being explored as an alternative for NK expansion and activation [34,35]. The clinical efficacy of different NK-based therapies in solid tumors remains to be determined in ongoing phase II clinical trials (e.g., NCT03958097, NCT03007823, NCT02409576, NCT03242603).
The lack of efficacy of primary NK cells was thought in part to be the result of suboptimal expansion of primary NK cells. The development of an optimal protocol to expand primary NK cells ex vivo for therapeutic use remains to be a challenge [36,37,38]. To overcome the challenge, many attempts using the NK cell line, NK-92, or the CD16-engineered NK-92 derivatives, have been made in clinical trials [39,40,41,42,43]. In comparison with primary NK cells, NK-92 can be expanded easily and indefinitely and has been proposed to be an “off-the-shelf” resource for immunotherapy [31,42,44,45]. Despite these benefits, irradiation of NK-92 cells is required prior to infusion into patients in order to prevent the possibility of malignant transformation [46], as the cell line was originated from a non-Hodgkin lymphoma patient [39]. This manipulation may limit the persistence of the infused cells and long-term anti-tumor therapeutic efficacy in patients [47].
Chimeric antigen receptors (CARs) are engineered receptors with an extracellular antigen-recognition domain for specific tumor antigens, connected with intracellular co-stimulatory and signaling domains that trigger lymphocyte activation upon antigen binding. CAR engineered NK-92 cells and primary NK cells have been evaluated in preclinical studies both in hematologic malignancies and solid tumors [48]. A number of clinical trials are underway to evaluate the potential of (CAR)-NK92 and CAR-primary NK cell therapies in refractory CD19+ malignancies, leukemia, and lymphomas, as well as myeloid malignancies in children and adults [48,49]. To overcome the limitations of using NK-92 cells and the challenge of manipulating primary NK cells, human induced pluripotent stem cell (iPSC)-differentiated NK cells have been used to construct iPSC-NK-CAR as an “off-shelf” CAR-NK strategy and tested in preclinical settings. It was shown that iPSC-CAR-NK therapy was significantly more effective than CAR-T therapy in reducing the tumor burden in mice bearing xenografted ovarian tumors [50]. Further, a reduced systemic toxicity, such as weight loss and cytokine release syndrome, was observed in mice receiving iPSC-CAR-NK treatment as compared with CAR-T treatment. This study presented the proof-of-concept that iPSC-NK-CAR can be a potential off-shelf cancer immunotherapeutic approach. Evidently, clinical evaluation of this therapeutic approach is warranted.
3. NK Cells Are Composed of Phenotypically and Functionally Diverse Subsets
Human NK cells are classically divided into two major subsets, CD56dimCD16hi and CD56brightCD16−/lo [51,52,53]. NK cells in human peripheral blood are composed of approximately 90% of CD56dimCD16hi and 10% of CD56brightCD16−/lo [51]. The two subsets of NK cells are thought to differ in maturation status and functions. The CD56dimCD16hi subset is considered mature or terminally differentiated NK cells and predominantly exerts cytolytic function. The CD56bright CD16−/lo NK cell subset is considered immature NK cells being less cytolytic, but has more immune regulatory function through cytokine secretion [51,53]. The CD56-based maturation and functional characteristics of human NK cells were supported by recent characterization based on the surface density of CD27 and CD11b, similar to the maturation characterization of mouse NK cells [54,55]. The increasing levels of maturity of human NK cells were characterized as CD11b−CD27−, CD11b−CD27+, CD11b+ CD27+, and CD11b+ CD27− [56]. By this criterion, it was shown that the majority of CD56bright NK cells were immature CD11b−CD27− and CD27+, whereas the majority of CD56dim NK cells are mature CD11b+CD27− [56]. Functionally, the CD11b+ CD27− subset displayed the highest cytotoxicity, whereas the CD27+ subsets displayed high potential in secreting cytokines. The majority of NK cells in the peripheral blood were shown to be CD11b+ CD27− (>95%) [56].
In healthy individuals, NK cells are widely distributed in the periphery. They not only circulate through the peripheral blood, but are also present in both lymphoid and non-lymphoid organs such as spleen, lymph nodes, liver, lung, and uterus. Contrary to their distribution in the peripheral blood, CD56brightCD16−/lo NK cells are the major subset in many tissues, such as lymph nodes, tonsil, liver, and uterus [52]. It is noteworthy that the CD56bright tissue-resident NK cell subset was shown to be phenotypically and functionally distinct from circulating CD56bright NK cells [51,52,53]. Phenotypically, tissue-resident NK cells express unique sets of surface adhesion molecules, such as CD69, CCR5, CXCR6, CD103, and/or CD49a, to fulfill their retention in respective tissues [51]. For example, decidual NK (dNK), also known as uterine NK (uNK) cells, which accounts for ~70% lymphocyte during early pregnancy, are CD56bright, but uniquely express CD49a and CD9 on their surface [51]. Functionally, dNK cells serve as a critical regulator of pregnancy and are involved in trophoblast invasion and vascular remodeling by secreting various chemokines and pro-angiogenic factors [52,57]. The origin of tissue-resident NK cells is controversial. Some evidence suggests that pre-existing hematopoietic precursors in tissues can differentiate in situ into tissue-resident NK cells [58], while others argue that the circulating NK cells could adapt to the tissue microenvironment and acquire tissue-specific phenotype and functions [59].
4. Enrichment of NK Cell Subsets with Diminished Cytotoxicity in Tumors
Tumor-infiltrating NK cells often have diminished cytotoxic function with distinct phenotypes. Most studies reported that tumor infiltrating NKs are mostly CD56bright. Several studies have shown a significant enrichment of non-cytotoxic CD56brightperforinlow NK cell subsets in lung and breast tumors as compared with the matched normal tissues [60,61]. Consistently, other studies have shown that in breast cancer patients, the subset of poor cytotoxic CD56brightNKG2AhiCD16lowKIRlo NK cells was increased in tumor infiltrates and that the increase correlated with poor disease prognosis [62]. A significant enrichment of the poor cytotoxic CD56brightCD16dim NK cell subset was also found in tumor infiltrates of melanoma and colon cancer [63].
The phenotype or function of NK cells in tumors is generally thought to be shaped by tumor microenvironment (TME) cues. There is evidence suggesting that chemokine milieu in the TME contributes to the accumulation of poor cytotoxic CD56bright subset of NK cells [61]. In neoplastic lung and breast tissues, it was shown that chemokines, such as CXCL2, CX3CL1, CXCL2, CXCL1, and CXCL8, that are specifically attracting the CD56dim NK cell subset, are downregulated, whereas chemokines more specific for attracting the CD56bright NK cell subset, such as CCL5, CCL19, CXCL9, and CXCL10, are upregulated [61]. However, whether the accumulation of the poor cytotoxic NK cells in human tumors is because of TME-induced alterations in NK phenotype, preferential migration of NK cell subsets in response to specific chemokine cues in TME, or differential survival/proliferation ability of the NK subsets in TME, or potential trans-differentiation of NK cells, is not well defined. Nonetheless, these studies demonstrated the complexity of TME in skewing NK cell function.
5. Tumor-Associated Immature NK Cell Phenotype
NK cell function is associated with its maturation status. Tumor infiltrating NK cells often present an immature phenotype. In a B16F10 lung metastasis model, it was showed that impaired NK maturation in mice lacking neonatal Fc receptor associated with reduced tumor control [64]. In patients with hepatocellular carcinoma, accumulation of the immature CD11b−CD27− NK cell subset in tumor infiltrates was shown to correlate with poor clinical outcome [65]. A substantial increase in the CD11b−CD27− NK cells and a concomitant reduction in single and double positive NK populations were observed in the tumor tissues as compared with adjacent non-tumor and control liver tissues [65]. Moreover, the frequency of the CD11b−CD27− NK cell subset correlated with the size of the resected tumors [65]. The CD11b−CD27− NK population was shown to have impaired production of IFNγ, as well as poor cytotoxic potential [65].
Pre-clinical studies suggest tumor secreted soluble mediators can curtail NK cell maturation. Two studies from Richards and group have demonstrated defective NK maturation in the bone marrow of mice bearing tumors of breast, colon, melanoma, and lymphoma [66,67]. In the first study, they found a significant reduction in the mature CD11bhi NK cells in the bone barrow of tumor-bearing mice as compared with non-tumor bearing control mice, suggesting an impact of tumor growth on the maturation status of NK cells [66]. A further study with adoptive transfer of bone-marrow derived immature CD11b− NK cells into tumor bearing mice demonstrated that NK cell maturation was arrested at the CD11blow stage [66]. In the second study, they showed that the tumor growth-associated reduction in NK cell numbers was attributed to the significant reduction in NK cell progenitors (CD122+NK1.1−DX5−CD3−) and common lymphoid progenitors (Lin−CD127+cKit+Sca+) with bone marrow transplant experiments [67]. Although underlying mechanisms associated with these observations were not fully dissected, the findings have evidently demonstrated that tumor-derived soluble factors negatively impact the lymphopoiesis and maturation process of NK cells.
There is evidence that tumors can induce a reversal in the maturation status of NK cells. Using a transgenic spontaneous polyoma middle T antigen (pyMT) breast tumor mouse model, Krneta et al. demonstrated striking differences in maturity and activation markers in intra-tumoral NK cells versus splenic NK cells from the tumor bearing mice [68]. They demonstrated that NK cells from the tumors displayed highly immature CD27lowCD11blow phenotype and low expression of granzyme B and perforin when compared with the splenic NK cells. To further delineate that the tumor microenvironment can directly modulate NK cell maturation and function, they adoptively transferred labeled NKp46+ mature splenic NK cells from C57BL/6 mice into pyMT mice. Intriguingly, after only three days of adoptive transfer, the intra-tumoral NK cells displayed a significant reduction of maturity markers CD27 and DX5 (CD49b) as compared with splenic NK cells. Together, these studies suggest that the tumor microenvironment can re-direct NK cell maturation status and function. Evidently, it is important for further investigation to understand the underlying mediators and pathways.
6. Cancer-Induced Phenotypic Changes in NK
NK cell in cancer patients, whether from tumor infiltrates or peripheral blood, often have an altered surface receptor repertoire that undermines the capacity to recognize and control tumor cells. Downregulation of activating NK receptors (e.g., NCRs, NKG2D, and DNAM-1) represents one of the common mechanisms employed by a wide array of cancer types to evade NK-mediated immunity [69]. In hematopoietic cancers such as AML, patient-derived NK often exhibit downregulation of NCRs (NKp30, NKp44, NKp46), which is correlated with their reduced cytotoxicity ex vivo against autologous leukemic cells [70,71,72]. This phenotype is most likely AML-induced, as the restoration, complete or partial, in the expression level of these receptors was seen in patients with complete remission [71]. The observed reduction in cytotoxicity ex vivo was shown to be a result of impaired target cell recognition owing to downregulation of NCR, rather than any intrinsic defect in the cytolytic capacity of the AML patient derived NK cells [70]. Downregulation of NCR was also shown in patients with solid tumors, such as non-small-cell lung cancer, melanoma, and breast cancer [62,73,74,75,76,77]. In non-small-cell lung carcinoma (NSCLC), for instance, tumor infiltrating NK cells are associated with a distinct surface receptor profile, characterized by lower expression of NKp30, NKp80, NKG2D, DNAM-1, and CD16 [73]. Upregulation of the inhibitory receptor NKG2A is another modulation that impairs NK cell cytotoxicity in cancer patients. It was well demonstrated in patients with breast cancer and liver cancer that tumor infiltrating NK cells have upregulated NKG2A expression [62,78]. It is noteworthy that NCR downregulation not only limits NK cell recognition of tumor cells, but also potentially limits T-cell mediated anti-tumor responses. It has been shown that NK cells can eliminate immature dendritic cells (iDC) through activation of NKp30, which in turn favors the expansion and function of mature DCs [79,80]. Interruption of the interaction of NK with iDC could potentially lead to suboptimal DC maturation and priming of tumor antigen-specific T cells and otherwise iDC-induced tolerance against certain tumor antigen.
Beyond the modifications on NK functional receptors, tumor modification of NK homing or exhaustion status has also been reported. Comparative microarray of tumor infiltrating (TI)-NK versus circulating NK from the same NSCLC patients yielded a unique TI-NK signature that is enriched in genes associated with NK activation, cytotoxicity, and migration [74]. Specifically, the study showed overexpression of CXCR5 and CXCR6 and lower expression of CX3CR1 and S1PR1 (sphingosine 1-phosphate receptor 1, a migration-associated receptor) at the transcript level on the TI-NK as compared with the circulating NK cells [74]. A recent study in endometrial tumor demonstrated severe impairment in the numbers and cytotoxic function of tumor infiltrating NK cells compared with those in the adjacent non-tumor tissues [81]. The same group also identified CD103+ “tumor resident” and CD103− “recruited” NK cell subset in cancer patients. As compared with the CD103− NK cells, the CD103+ NK cells displayed a significant increase in the expression of co-inhibitory markers TIM3 and TIGIT, which have been associated with inept NK cell responses [81,82,83,84,85,86]. NK cells from the patients with lymph node metastatic diseases have upregulated expression of these co-inhibitory molecules as compared with those with only localized diseases, suggesting that tumor-induced modulations affect the phenotypic and functional features of NK cells with disease progression [87].
There is evidence for the presence of phenotypically and functionally unique NK subsets, different from the conventional CD56brightCD16dim and CD56dimCD16bright NK cell phenotype, in cancer patients. Mamessier et al. has described the CD56brightCD16+ and CD56dimCD16− subsets in breast cancer patients and further demonstrated the enrichment of these subsets in relevance to disease progression [88]. They demonstrated that both the NK subsets, CD56brightCD16+ and CD56dimCD16−, had functional immature phenotype of CD27+CD117+ and were highly enriched in the peripheral blood of patients with advanced breast cancer as compared with subjects with benign tumor and localized diseases. Interestingly, they further found that the frequency of CD56brightCD16+ and CD56dimCD16− subsets in mammary tumors reflected the frequency of respective subset population in the peripheral blood [88].
The collective evidence suggests that the impact of tumor microenvironment on NK cell function can be multifaceted. The underlying mechanism whereby tumor microenvironment skews NK cell phenotype and activity is not fully understood, although it has been proposed that various cancer-associated immunosuppressive factors are the key mediators. For instance, reactive oxygen species, TGF-β, PGE2, and IDO1, which are derived from tumor cells or other immunosuppressive cells in the TME, have been shown to contribute to downregulation of NK receptors, including the NCRs and NKG2D [75,89,90,91,92,93,94,95]. On the other hand, it has been shown in vitro that IL-15 can rescue or counteract tumor-induced downregulation of NK receptors [72,96], which supports the current in-development cytokine therapeutic approach in combination with NK cell-based therapy.
7. Tumor-Induced NK Functional Plasticity
Human peripheral blood NKs possess a certain degree of plasticity to acquire an altered functional phenotype [97,98]. In this section, we will present evidence of tumor-induced phenotypical and functional alterations in mature NK cells in mouse models and/or cancer patients (Table 1) and discuss potential mechanisms underlying these conversions.
7.1. Tumor-Induced Conversion of NK to dNK-Like Cells
A subset of NK cells displaying the “dNK-like” phenotype has been shown in the peripheral blood, pleural effusion, and TILs of patients with different types of cancer, including colorectal cancer and lung cancer [63,99,100,103,104]. Phenotypically, these NKs are predominantly CD56brightCD16−/dim and express CD9 and/or CD49a. A number of studies have demonstrated that these “dNK-like” cells secrete pro-angiogenic factors such as VEGF and angiogenin [99,100]. It was shown that supernatants from cultured “dNK-like” cells promoted endothelial cell tube formation in vitro [100,104]. It was also shown that NKs isolated from colorectal cancer patients had upregulated invasion-associated enzymes related to the MMP9-TIMP2/9 axis, a feature similar to dNK in tissue remodeling and angiogenesis [100]. The presence of this population in cancer patients, but not healthy individuals, suggests that the dNK-like phenotype and function is driven by tumor-related factors.
Among many tumor-secreted soluble mediators, TGF-β has been shown to mediate the conversion of classical NK cells into dNK-like cells. Treatment of NK cells from healthy donors with TGF-β1 can induce NK cells to produce elevated levels of VEGF and PIGF (placental growth factor, a member of the VEGF family) [99]. It was shown that upon exposure to TGF-β in vitro, CD16+ peripheral blood NKs could be converted to a CD16− dNK-like phenotype with acquired expression of CD9 and CD49a on their surface [97]. Combining TGF-β with hypoxia and a demethylating agent (5-Aza) further skewed peripheral blood NK cells towards a dNK-like phenotype [98]. The converted CD16−CD9+CD49a+ NK cells displayed reduced cytotoxicity, secreted the pro-angiogenic factor VEGF, and were able to promote trophoblast tumor invasion [98]. With the evidence suggesting that circulating NKs could be recruited to the uterus during early pregnancy and undergo reprogramming in the uterine microenvironment [59], it is plausible to hypothesize that the tumor microenvironment with high levels of TGF-β and hypoxia can induce reprogramming of NK cells to dNK-like phenotype.
7.2. Tumor-Induced Conversion of NK Cell into Type I Innate Lymphoid Cells (ILC1)-Like Phenotype
NK cells and type I innate lymphoid cells (ILC1s) are two members of the group I innate lymphoid cells, grouped together based on their shared capacity to produce IFN-γ [105,106]. In contrast to circulating conventional NK cells, ILC1s consist of tissue-resident subsets and are rarely found in the circulation at steady state [107]. In mice, ILC1s are identified in tissues including spleen, liver, small intestine, salivary gland, and adipose tissues at steady state, and they are also found in tumor tissues [108,109,110,111,112,113,114]. In human, ILC1 subsets are described in tonsil, gut, and tumor tissues [105,115,116,117]. While conventional NK cells are considered to possess dual effector functions of both cytokine secretion and cytotoxicity, ILC1s are often viewed as potent cytokine secretors with poor cytolytic capacity [105,110]. Lineage tracing indicates that conventional NK cells and ILCs arise from distinct progenitors [110,111,118]. In mice, both transcriptional factors T-bet and Eomes are required for conventional NK cell development, whereas only T-bet is required for ILC1 development [119]. Human and mouse ILC1s have been associated with hallmarks of TGF-β imprinting, such as the expression of CD103 [112,115].
Mouse NK cells and ILC1s both express NK1.1 and NKp46 [105,106]. The two populations are often distinguished based on their differential expression of CD49a and Eomes. NK cells are CD49a−Eomes+, whereas ILC1s are CD49a+ Eomes− [101]. At steady state, TRAIL, CD73, CD200R1, and CD61 could serve as additional surface markers for ILC1s [105,120]. The phenotypic distinction between NK cells and ILC1s in pathological conditions is not very well characterized. One study showed that CD49b+ Eomes+ mouse NK cells have increased expression of CD49a and CD61, with markers being associated with ILC1s in murine cytomegalovirus (MCMV)-infected mice [120]. Therefore, discrimination of NK cells and ILC1s solely based on surface markers is insufficient and needs to be combined with other criteria such as Eomes expression and expression of ILC1 signature genes [105,108].
It is well established that NK cells play a critical role in controlling tumor growth and metastasis [121,122,123,124,125]. It is unclear to what extent ILC1s participate in anti-tumor immunosurveillance, despite their phenotypical and functional similarities to NK cells. To date, there is little evidence to support inter-conversion between the NK cells and ILC1s under normal physiological conditions. However, the plasticity between mouse NK and ILC1 in the context of tumors was recently demonstrated [101]. Using a variety of mouse tumor models, Gao et al. showed that tumor-infiltrating NKs (CD49a−CD49b+) could be converted to an intermediate ILC1 (intILC1, CD49a+CD49b+) phenotype characterized by reduced Eomes and increased CD49a expression. Tumor-infiltrating intILC1s lost the ability to control tumor initiation and metastasis, and displayed profound phenotypical and transcriptomic alterations compared with TI-NKs [101]. It was shown that intILC1s are functionally impaired with upregulated expression of immune checkpoint receptors [101]. intILC1s was shown to be pro-tumorigenic. Mechanistically, it was shown that intILC1s secrete pro-angiogenic factors such as platelet-derived growth factor (PDGF)-AB and could promote endothelial tube formation in vitro [101]. In the same study, CXCR6+ ILC1-like cells were described in the peripheral and TILs of patients with gastrointestinal stromal tumors (GIST) [101], although whether these cells are converted from classical NK cells is unknown.
Non-canonical TGF-β signaling has been implicated in promoting the conversion of conventional NK cells to an ILC1-like phenotype. Gao et al. demonstrated that the ILC1-like conversion of NK cells in the mouse models was dependent on non-canonical TGF-β signaling in NK cells, and that NK cells deficient in TGF-β signaling failed the conversion [101]. Cortez et al. further revealed that SMAD4, a central signal transducer of the canonical TGF-β signaling, could paradoxically act as a negative regulator of the conversion by curtailing non-canonical TGF-β signaling [102]. SMAD4-deficient mouse NK cells are hyper-responsive to TGF-β receptor 1 (TGFβ R1)-mediated signals. The SMAD4-deficeint NK cells isolated from the spleen, liver, and small intestine acquire ILC1 characteristic markers, such as CD49a, CD73, and TRAIL. These mice displayed impaired anti-tumor immunity [102]. A recent study by Rautela et al. showed that independent of TGF-β signaling, activin-A (a member of the TGF-β family) signaling could induce a similar ILC1-like conversion (increased CD49a and decreased Eomes expression) in conventional NK cells [126].
Recently, a new subset of regulatory CD3-CD56+ ILC subset in the TILs of patients with high-grade serous tumors has been described [127]. The presence of this subset TIL cultures correlates with a reduction in the time to disease progression. In vitro, this CD3−CD56+ subset was poorly cytotoxic, but suppressed TIL expansion and altered TIL cytokine production. These CD3−CD56+ cells presented a distinct cytokine profile from those of NK cells and other ILCs. It is unclear whether these cells were the result of conversion of NK cells in the tumor microenvironment, or whether they represent a novel, distinct ILC population.
Beyond evidence demonstrating the tumor-induced conversion of NK cells to ILC1-like and dNK-like phenotypes, one study showed that NK cells could be converted to myeloid derived suppressor cells in tumor-bearing mice in a GM-CSF dependent manner [128]. However, to date, no other studies have reported similar results to support this observation.
8. Concluding Remarks
Mature NK cells were considered terminally differentiated NK cells that play a significant role in controlling tumor growth and metastasis. Many NK cell-based therapies with the intention to harness NK cell receptors to enhance NK cell cytolytic function are currently being developed for treating solid tumors. Recent findings indicate that specific tumor environment cues can profoundly impact NK phenotype and function (Figure 1) and NK cell-based therapeutic efficacy, and more profoundly, could potentially convert therapeutic NK cells into tumor-promoting NK cells. Therefore, strategies to co-target tumor microenvironment, such as co-targeting hypoxia pathway, tumor released TGF-β, or non-canonical TGF-β signaling pathways, should be considered in order to achieve desired therapeutic efficacy. Evidently, further investigations to gain an in-depth understanding of the molecular mechanisms that drive NK cell functional plasticity are warranted.
Author Contributions
S.L. and P.D. co-drafted the manuscript. J.D.W. edited the manuscript. J.D.W. conceived the concept.
Funding
NIH-NCI grant 1R01CA208246, 1R01CA204021, and R01CA212409 (to J.D.W.); by the Department of Defense—Prostate Cancer Research Program award W81XWH-15-1-0406 and W81XWH-17-1-0642 (to J.D.W.) and NIH/NCI Prostate Cancer SPORE P50 CA180995.
Conflicts of Interest
The authors declare no conflict of interests.
Abbreviations
| CD | clusters of differentiation; |
| KIR | killer-cell immunoglobulin-like receptor; |
| CD94/NKG2A | natural killer group 2 member A, complexed with CD94; |
| NKG2D | natural killer group 2 member D; |
| NCR | natural cytotoxicity receptor; |
| DNAM-1 | DNAX accessory molecule; |
| IFN-γ | interferon-gamma; |
| TNF-a | tumor necrosis factor-alpha; |
| GM-CSF | granulocyte-macrophage colony-stimulating factor; |
| HLA | human leukocyte antigen; |
| NSCLC | non small cell lung carcinoma; |
| TIM-3 | T-cell immunoglobulin mucin receptor 3; |
| TIGIT | T cell immunoreceptor with Ig and ITIM Domains; |
| LAG-3 | lymphocyte activating 3; |
| CTLA-4 | cytotoxic T-lymphocyte associated protein 4; |
| VEGF | vascular endothelial growth factor; |
| PDGF-AB | platelet-derived growth factor-AB; |
References
- Lanier, L.L. NK cell recognition. Annu. Rev. Immunol. 2005, 23, 225–274. [Google Scholar] [CrossRef] [PubMed]
- Guillerey, C.; Huntington, N.D.; Smyth, M.J. Targeting natural killer cells in cancer immunotherapy. Nat. Immunol. 2016, 17, 1025–1036. [Google Scholar] [CrossRef] [PubMed]
- Souza-Fonseca-Guimaraes, F.; Cursons, J.; Huntington, N.D. The Emergence of Natural Killer Cells as a Major Target in Cancer Immunotherapy. Trends Immunol. 2019, 40, 142–158. [Google Scholar] [CrossRef] [PubMed]
- Bottcher, J.P.; Bonavita, E.; Chakravarty, P.; Blees, H.; Cabeza-Cabrerizo, M.; Sammicheli, S.; Rogers, N.C.; Sahai, E.; Zelenay, S.; Reis e Sousa, C. NK Cells Stimulate Recruitment of cDC1 into the Tumor Microenvironment Promoting Cancer Immune Control. Cell 2018, 172, 1022–1037. [Google Scholar] [CrossRef] [PubMed]
- Walzer, T.; Dalod, M.; Robbins, S.H.; Zitvogel, L.; Vivier, E. Natural-killer cells and dendritic cells: L’union fait la force. Blood 2005, 106, 2252–2258. [Google Scholar] [CrossRef] [PubMed]
- Barry, K.C.; Hsu, J.; Broz, M.L.; Cueto, F.J.; Binnewies, M.; Combes, A.J.; Nelson, A.E.; Loo, K.; Kumar, R.; Rosenblum, M.D.; et al. A natural killer-dendritic cell axis defines checkpoint therapy-responsive tumor microenvironments. Nat. Med. 2018, 24, 1178–1191. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; Ugolini, S. Regulatory natural killer cells: New players in the IL-10 anti-inflammatory response. Cell Host Microbe 2009, 6, 493–495. [Google Scholar] [CrossRef] [PubMed]
- Imai, K.; Matsuyama, S.; Miyake, S.; Suga, K.; Nakachi, K. Natural cytotoxic activity of peripheral-blood lymphocytes and cancer incidence: An 11-year follow-up study of a general population. Lancet 2000, 356, 1795–1799. [Google Scholar] [CrossRef]
- Miller, J.S.; Soignier, Y.; Panoskaltsis-Mortari, A.; McNearney, S.A.; Yun, G.H.; Fautsch, S.K.; McKenna, D.; Le, C.; Defor, T.E.; Burns, L.J.; et al. Successful adoptive transfer and in vivo expansion of human haploidentical NK cells in patients with cancer. Blood 2005, 105, 3051–3057. [Google Scholar] [CrossRef] [Green Version]
- Curti, A.; Ruggeri, L.; D’Addio, A.; Bontadini, A.; Dan, E.; Motta, M.R.; Trabanelli, S.; Giudice, V.; Urbani, E.; Martinelli, G.; et al. Successful transfer of alloreactive haploidentical KIR ligand-mismatched natural killer cells after infusion in elderly high risk acute myeloid leukemia patients. Blood 2011, 118, 3273–3279. [Google Scholar] [CrossRef]
- Rea, D.; Henry, G.; Khaznadar, Z.; Etienne, G.; Guilhot, F.; Nicolini, F.; Guilhot, J.; Rousselot, P.; Huguet, F.; Legros, L.; et al. Natural killer-cell counts are associated with molecular relapse-free survival after imatinib discontinuation in chronic myeloid leukemia: The IMMUNOSTIM study. Haematologica 2017, 102, 1368–1377. [Google Scholar] [CrossRef] [PubMed]
- Imagawa, J.; Tanaka, H.; Okada, M.; Nakamae, H.; Hino, M.; Murai, K.; Ishida, Y.; Kumagai, T.; Sato, S.; Ohashi, K.; et al. Discontinuation of dasatinib in patients with chronic myeloid leukaemia who have maintained deep molecular response for longer than 1 year (DADI trial): A multicentre phase 2 trial. Lancet Haematol. 2015, 2, e528–e535. [Google Scholar] [CrossRef]
- Ilander, M.; Olsson-Stromberg, U.; Schlums, H.; Guilhot, J.; Bruck, O.; Lahteenmaki, H.; Kasanen, T.; Koskenvesa, P.; Soderlund, S.; Hoglund, M.; et al. Increased proportion of mature NK cells is associated with successful imatinib discontinuation in chronic myeloid leukemia. Leukemia 2017, 31, 1108–1116. [Google Scholar] [CrossRef] [PubMed]
- Coca, S.; Perez-Piqueras, J.; Martinez, D.; Colmenarejo, A.; Saez, M.A.; Vallejo, C.; Martos, J.A.; Moreno, M. The prognostic significance of intratumoral natural killer cells in patients with colorectal carcinoma. Cancer 1997, 79, 2320–2328. [Google Scholar] [CrossRef]
- Ishigami, S.; Natsugoe, S.; Tokuda, K.; Nakajo, A.; Che, X.; Iwashige, H.; Aridome, K.; Hokita, S.; Aikou, T. Prognostic value of intratumoral natural killer cells in gastric carcinoma. Cancer 2000, 88, 577–583. [Google Scholar] [CrossRef]
- Takanami, I.; Takeuchi, K.; Giga, M. The prognostic value of natural killer cell infiltration in resected pulmonary adenocarcinoma. J. Thorac. Cardiovasc. Surg. 2001, 121, 1058–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villegas, F.R.; Coca, S.; Villarrubia, V.G.; Jimenez, R.; Chillon, M.J.; Jareno, J.; Zuil, M.; Callol, L. Prognostic significance of tumor infiltrating natural killer cells subset CD57 in patients with squamous cell lung cancer. Lung Cancer 2002, 35, 23–28. [Google Scholar] [CrossRef]
- Beano, A.; Signorino, E.; Evangelista, A.; Brusa, D.; Mistrangelo, M.; Polimeni, M.A.; Spadi, R.; Donadio, M.; Ciuffreda, L.; Matera, L. Correlation between NK function and response to trastuzumab in metastatic breast cancer patients. J. Transl. Med. 2008, 6, 25. [Google Scholar] [CrossRef]
- Pasero, C.; Gravis, G.; Granjeaud, S.; Guerin, M.; Thomassin-Piana, J.; Rocchi, P.; Salem, N.; Walz, J.; Moretta, A.; Olive, D. Highly effective NK cells are associated with good prognosis in patients with metastatic prostate cancer. Oncotarget 2015, 6, 14360–14373. [Google Scholar] [CrossRef]
- Cursons, J.; Souza-Fonseca-Guimaraes, F.; Foroutan, M.; Anderson, A.; Hollande, F.; Hediyeh-Zadeh, S.; Behren, A.; Huntington, N.D.; Davis, M.J. A Gene Signature Predicting Natural Killer Cell Infiltration and Improved Survival in Melanoma Patients. Cancer Immunol. Res. 2019, 7, 1162–1174. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, R.; Wu, H.; Pounds, S.; Inaba, H.; Ribeiro, R.C.; Cullins, D.; Rooney, B.; Bell, T.; Lacayo, N.J.; Heym, K.; et al. A phase II clinical trial of adoptive transfer of haploidentical natural killer cells for consolidation therapy of pediatric acute myeloid leukemia. J. Immunother. Cancer 2019, 7, 81. [Google Scholar] [CrossRef] [PubMed]
- Boyiadzis, M.; Agha, M.; Redner, R.L.; Sehgal, A.; Im, A.; Hou, J.Z.; Farah, R.; Dorritie, K.A.; Raptis, A.; Lim, S.H.; et al. Phase 1 clinical trial of adoptive immunotherapy using “off-the-shelf” activated natural killer cells in patients with refractory and relapsed acute myeloid leukemia. Cytotherapy 2017, 19, 1225–1232. [Google Scholar] [CrossRef] [PubMed]
- Suen, W.C.; Lee, W.Y.; Leung, K.T.; Pan, X.H.; Li, G. Natural Killer Cell-Based Cancer Immunotherapy: A Review on 10 Years Completed Clinical Trials. Cancer Investig. 2018, 36, 431–457. [Google Scholar] [CrossRef] [PubMed]
- Rubnitz, J.E.; Inaba, H.; Ribeiro, R.C.; Pounds, S.; Rooney, B.; Bell, T.; Pui, C.H.; Leung, W. NKAML: A pilot study to determine the safety and feasibility of haploidentical natural killer cell transplantation in childhood acute myeloid leukemia. J. Clin. Oncol. 2010, 28, 955–959. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.A.; Leveson-Gower, D.B.; Gill, S.; Baker, J.; Beilhack, A.; Negrin, R.S. NK cells mediate reduction of GVHD by inhibiting activated, alloreactive T cells while retaining GVT effects. Blood 2010, 115, 4293–4301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, F.; Lin, H.; Gao, S.; Hu, Z.; Zuo, S.; Sun, L.; Jin, C.; Li, W.; Yang, Y. The anti-tumor role of NK cells in vivo pre-activated and re-stimulated by interleukins in acute lymphoblastic leukemia. Oncotarget 2016, 7, 79187–79202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruggeri, L.; Capanni, M.; Urbani, E.; Perruccio, K.; Shlomchik, W.D.; Tosti, A.; Posati, S.; Rogaia, D.; Frassoni, F.; Aversa, F.; et al. Effectiveness of donor natural killer cell alloreactivity in mismatched hematopoietic transplants. Science 2002, 295, 2097–2100. [Google Scholar] [CrossRef]
- Ishikawa, T.; Okayama, T.; Sakamoto, N.; Ideno, M.; Oka, K.; Enoki, T.; Mineno, J.; Yoshida, N.; Katada, K.; Kamada, K.; et al. Phase I clinical trial of adoptive transfer of expanded natural killer cells in combination with IgG1 antibody in patients with gastric or colorectal cancer. Int. J. Cancer 2018, 142, 2599–2609. [Google Scholar] [CrossRef]
- Yang, Y.; Lim, O.; Kim, T.M.; Ahn, Y.O.; Choi, H.; Chung, H.; Min, B.; Her, J.H.; Cho, S.Y.; Keam, B.; et al. Phase I Study of Random Healthy Donor-Derived Allogeneic Natural Killer Cell Therapy in Patients with Malignant Lymphoma or Advanced Solid Tumors. Cancer Immunol. Res. 2016, 4, 215–224. [Google Scholar] [CrossRef]
- Sakamoto, N.; Ishikawa, T.; Kokura, S.; Okayama, T.; Oka, K.; Ideno, M.; Sakai, F.; Kato, A.; Tanabe, M.; Enoki, T.; et al. Phase I clinical trial of autologous NK cell therapy using novel expansion method in patients with advanced digestive cancer. J. Transl. Med. 2015, 13, 277. [Google Scholar] [CrossRef]
- Tonn, T.; Schwabe, D.; Klingemann, H.G.; Becker, S.; Esser, R.; Koehl, U.; Suttorp, M.; Seifried, E.; Ottmann, O.G.; Bug, G. Treatment of patients with advanced cancer with the natural killer cell line NK-92. Cytotherapy 2013, 15, 1563–1570. [Google Scholar] [CrossRef] [PubMed]
- Iliopoulou, E.G.; Kountourakis, P.; Karamouzis, M.V.; Doufexis, D.; Ardavanis, A.; Baxevanis, C.N.; Rigatos, G.; Papamichail, M.; Perez, S.A. A phase I trial of adoptive transfer of allogeneic natural killer cells in patients with advanced non-small cell lung cancer. Cancer Immunol. Immunother. 2010, 59, 1781–1789. [Google Scholar] [CrossRef] [PubMed]
- Geller, M.A.; Cooley, S.; Judson, P.L.; Ghebre, R.; Carson, L.F.; Argenta, P.A.; Jonson, A.L.; Panoskaltsis-Mortari, A.; Curtsinger, J.; McKenna, D.; et al. A phase II study of allogeneic natural killer cell therapy to treat patients with recurrent ovarian and breast cancer. Cytotherapy 2011, 13, 98–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conlon, K.C.; Lugli, E.; Welles, H.C.; Rosenberg, S.A.; Fojo, A.T.; Morris, J.C.; Fleisher, T.A.; Dubois, S.P.; Perera, L.P.; Stewart, D.M.; et al. Redistribution, hyperproliferation, activation of natural killer cells and CD8 T cells, and cytokine production during first-in-human clinical trial of recombinant human interleukin-15 in patients with cancer. J. Clin. Oncol. 2015, 33, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Childs, R.W.; Carlsten, M. Therapeutic approaches to enhance natural killer cell cytotoxicity against cancer: The force awakens. Nat. Rev. Drug Discov. 2015, 14, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Tang, S.Y.; Toh, L.L.; Wang, S. Generation of “Off-the-Shelf” Natural Killer Cells from Peripheral Blood Cell-Derived Induced Pluripotent Stem Cells. Stem Cell Rep. 2017, 9, 1796–1812. [Google Scholar] [CrossRef] [PubMed]
- Fujisaki, H.; Kakuda, H.; Shimasaki, N.; Imai, C.; Ma, J.; Lockey, T.; Eldridge, P.; Leung, W.H.; Campana, D. Expansion of highly cytotoxic human natural killer cells for cancer cell therapy. Cancer Res. 2009, 69, 4010–4017. [Google Scholar] [CrossRef]
- Koehl, U.; Brehm, C.; Huenecke, S.; Zimmermann, S.Y.; Kloess, S.; Bremm, M.; Ullrich, E.; Soerensen, J.; Quaiser, A.; Erben, S.; et al. Clinical grade purification and expansion of NK cell products for an optimized manufacturing protocol. Front. Oncol. 2013, 3, 118. [Google Scholar] [CrossRef]
- Gong, J.H.; Maki, G.; Klingemann, H.G. Characterization of a human cell line (NK-92) with phenotypical and functional characteristics of activated natural killer cells. Leukemia 1994, 8, 652–658. [Google Scholar]
- Tam, Y.K.; Miyagawa, B.; Ho, V.C.; Klingemann, H.G. Immunotherapy of malignant melanoma in a SCID mouse model using the highly cytotoxic natural killer cell line NK-92. J. Hematother. 1999, 8, 281–290. [Google Scholar] [CrossRef]
- Klingemann, H.G.; Wong, E.; Maki, G. A cytotoxic NK-cell line (NK-92) for ex vivo purging of leukemia from blood. Biol. Blood Marrow Transplant. 1996, 2, 68–75. [Google Scholar] [PubMed]
- Klingemann, H.; Boissel, L.; Toneguzzo, F. Natural Killer Cells for Immunotherapy—Advantages of the NK-92 Cell Line over Blood NK Cells. Front. Immunol. 2016, 7, 91. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Oberoi, P.; Oelsner, S.; Waldmann, A.; Lindner, A.; Tonn, T.; Wels, W.S. Chimeric Antigen Receptor-Engineered NK-92 Cells: An Off-the-Shelf Cellular Therapeutic for Targeted Elimination of Cancer Cells and Induction of Protective Antitumor Immunity. Front. Immunol. 2017, 8, 533. [Google Scholar] [CrossRef] [PubMed]
- Hermanson, D.L.; Kaufman, D.S. Utilizing chimeric antigen receptors to direct natural killer cell activity. Front. Immunol. 2015, 6, 195. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Steinherz, P.; Klingemann, H.G.; Dennig, D.; Childs, B.H.; McGuirk, J.; O’Reilly, R.J. Antileukemia activity of a natural killer cell line against human leukemias. Clin. Cancer Res. 1998, 4, 2859–2868. [Google Scholar] [PubMed]
- Rezvani, K.; Rouce, R.H. The Application of Natural Killer Cell Immunotherapy for the Treatment of Cancer. Front. Immunol. 2015, 6, 578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schonfeld, K.; Sahm, C.; Zhang, C.; Naundorf, S.; Brendel, C.; Odendahl, M.; Nowakowska, P.; Bonig, H.; Kohl, U.; Kloess, S.; et al. Selective inhibition of tumor growth by clonal NK cells expressing an ErbB2/HER2-specific chimeric antigen receptor. Mol. Ther. 2015, 23, 330–338. [Google Scholar] [CrossRef]
- Rezvani, K.; Rouce, R.; Liu, E.; Shpall, E. Engineering Natural Killer Cells for Cancer Immunotherapy. Mol. Ther. 2017, 25, 1769–1781. [Google Scholar] [CrossRef]
- Lin, C.; Zhang, J. Reformation in chimeric antigen receptor based cancer immunotherapy: Redirecting natural killer cell. Biochim. Biophys. Acta Rev. Cancer 2018, 1869, 200–215. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hermanson, D.L.; Moriarity, B.S.; Kaufman, D.S. Human iPSC-Derived Natural Killer Cells Engineered with Chimeric Antigen Receptors Enhance Anti-tumor Activity. Cell Stem Cell 2018, 23, 181–192.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freud, A.G.; Mundy-Bosse, B.L.; Yu, J.; Caligiuri, M.A. The Broad Spectrum of Human Natural Killer Cell Diversity. Immunity 2017, 47, 820–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjorkstrom, N.K.; Ljunggren, H.G.; Michaelsson, J. Emerging insights into natural killer cells in human peripheral tissues. Nat. Rev. Immunol. 2016, 16, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Melsen, J.E.; Lugthart, G.; Lankester, A.C.; Schilham, M.W. Human Circulating and Tissue-Resident CD56(bright) Natural Killer Cell Populations. Front. Immunol. 2016, 7, 262. [Google Scholar] [CrossRef] [PubMed]
- Chiossone, L.; Chaix, J.; Fuseri, N.; Roth, C.; Vivier, E.; Walzer, T. Maturation of mouse NK cells is a 4-stage developmental program. Blood 2009, 113, 5488–5496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayakawa, Y.; Smyth, M.J. CD27 dissects mature NK cells into two subsets with distinct responsiveness and migratory capacity. J. Immunol. 2006, 176, 1517–1524. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.; Wang, F.; Sun, R.; Ling, B.; Tian, Z.; Wei, H. CD11b and CD27 reflect distinct population and functional specialization in human natural killer cells. Immunology 2011, 133, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Hanna, J.; Goldman-Wohl, D.; Hamani, Y.; Avraham, I.; Greenfield, C.; Natanson-Yaron, S.; Prus, D.; Cohen-Daniel, L.; Arnon, T.I.; Manaster, I.; et al. Decidual NK cells regulate key developmental processes at the human fetal-maternal interface. Nat. Med. 2006, 12, 1065–1074. [Google Scholar] [CrossRef]
- Vacca, P.; Vitale, C.; Montaldo, E.; Conte, R.; Cantoni, C.; Fulcheri, E.; Darretta, V.; Moretta, L.; Mingari, M.C. CD34+ hematopoietic precursors are present in human decidua and differentiate into natural killer cells upon interaction with stromal cells. Proc. Natl. Acad. Sci. USA 2011, 108, 2402–2407. [Google Scholar] [CrossRef]
- Carlino, C.; Stabile, H.; Morrone, S.; Bulla, R.; Soriani, A.; Agostinis, C.; Bossi, F.; Mocci, C.; Sarazani, F.; Tedesco, F.; et al. Recruitment of circulating NK cells through decidual tissues: A possible mechanism controlling NK cell accumulation in the uterus during early pregnancy. Blood 2008, 111, 3108–3115. [Google Scholar] [CrossRef]
- Carrega, P.; Morandi, B.; Costa, R.; Frumento, G.; Forte, G.; Altavilla, G.; Ratto, G.B.; Mingari, M.C.; Moretta, L.; Ferlazzo, G. Natural killer cells infiltrating human nonsmall-cell lung cancer are enriched in CD56 bright CD16(-) cells and display an impaired capability to kill tumor cells. Cancer 2008, 112, 863–875. [Google Scholar] [CrossRef]
- Carrega, P.; Bonaccorsi, I.; Di Carlo, E.; Morandi, B.; Paul, P.; Rizzello, V.; Cipollone, G.; Navarra, G.; Mingari, M.C.; Moretta, L.; et al. CD56(bright)perforin(low) noncytotoxic human NK cells are abundant in both healthy and neoplastic solid tissues and recirculate to secondary lymphoid organs via afferent lymph. J. Immunol. 2014, 192, 3805–3815. [Google Scholar] [CrossRef] [PubMed]
- Mamessier, E.; Sylvain, A.; Thibult, M.L.; Houvenaeghel, G.; Jacquemier, J.; Castellano, R.; Goncalves, A.; Andre, P.; Romagne, F.; Thibault, G.; et al. Human breast cancer cells enhance self tolerance by promoting evasion from NK cell antitumor immunity. J. Clin. Investig. 2011, 121, 3609–3622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levi, I.; Amsalem, H.; Nissan, A.; Darash-Yahana, M.; Peretz, T.; Mandelboim, O.; Rachmilewitz, J. Characterization of tumor infiltrating natural killer cell subset. Oncotarget 2015, 6, 13835–13843. [Google Scholar] [CrossRef] [PubMed]
- Castaneda, D.C.; Dhommee, C.; Baranek, T.; Dalloneau, E.; Lajoie, L.; Valayer, A.; Arnoult, C.; Demattei, M.V.; Fouquenet, D.; Parent, C.; et al. Lack of FcRn Impairs Natural Killer Cell Development and Functions in the Tumor Microenvironment. Front. Immunol. 2018, 9, 2259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.F.; Yin, W.W.; Xia, Y.; Yi, Y.Y.; He, Q.F.; Wang, X.; Ren, H.; Zhang, D.Z. Liver-infiltrating CD11b(-)CD27(-) NK subsets account for NK-cell dysfunction in patients with hepatocellular carcinoma and are associated with tumor progression. Cell Mol. Immunol. 2017, 14, 819–829. [Google Scholar] [CrossRef]
- Richards, J.O.; Chang, X.; Blaser, B.W.; Caligiuri, M.A.; Zheng, P.; Liu, Y. Tumor growth impedes natural-killer-cell maturation in the bone marrow. Blood 2006, 108, 246–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, J.; McNally, B.; Fang, X.; Caligiuri, M.A.; Zheng, P.; Liu, Y. Tumor growth decreases NK and B cells as well as common lymphoid progenitor. PLoS ONE 2008, 3, e3180. [Google Scholar] [CrossRef]
- Krneta, T.; Gillgrass, A.; Chew, M.; Ashkar, A.A. The breast tumor microenvironment alters the phenotype and function of natural killer cells. Cell Mol. Immunol. 2016, 13, 628–639. [Google Scholar] [CrossRef]
- Chretien, A.S.; Le Roy, A.; Vey, N.; Prebet, T.; Blaise, D.; Fauriat, C.; Olive, D. Cancer-Induced Alterations of NK-Mediated Target Recognition: Current and Investigational Pharmacological Strategies Aiming at Restoring NK-Mediated Anti-Tumor Activity. Front. Immunol. 2014, 5, 122. [Google Scholar] [CrossRef] [Green Version]
- Costello, R.T.; Sivori, S.; Marcenaro, E.; Lafage-Pochitaloff, M.; Mozziconacci, M.J.; Reviron, D.; Gastaut, J.A.; Pende, D.; Olive, D.; Moretta, A. Defective expression and function of natural killer cell-triggering receptors in patients with acute myeloid leukemia. Blood 2002, 99, 3661–3667. [Google Scholar] [CrossRef]
- Fauriat, C.; Just-Landi, S.; Mallet, F.; Arnoulet, C.; Sainty, D.; Olive, D.; Costello, R.T. Deficient expression of NCR in NK cells from acute myeloid leukemia: Evolution during leukemia treatment and impact of leukemia cells in NCRdull phenotype induction. Blood 2007, 109, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Szczepanski, M.J.; Szajnik, M.; Welsh, A.; Foon, K.A.; Whiteside, T.L.; Boyiadzis, M. Interleukin-15 enhances natural killer cell cytotoxicity in patients with acute myeloid leukemia by upregulating the activating NK cell receptors. Cancer Immunol. Immunother. 2010, 59, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Platonova, S.; Cherfils-Vicini, J.; Damotte, D.; Crozet, L.; Vieillard, V.; Validire, P.; Andre, P.; Dieu-Nosjean, M.C.; Alifano, M.; Regnard, J.F.; et al. Profound coordinated alterations of intratumoral NK cell phenotype and function in lung carcinoma. Cancer Res. 2011, 71, 5412–5422. [Google Scholar] [CrossRef] [PubMed]
- Gillard-Bocquet, M.; Caer, C.; Cagnard, N.; Crozet, L.; Perez, M.; Fridman, W.H.; Sautes-Fridman, C.; Cremer, I. Lung tumor microenvironment induces specific gene expression signature in intratumoral NK cells. Front. Immunol. 2013, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Pietra, G.; Manzini, C.; Rivara, S.; Vitale, M.; Cantoni, C.; Petretto, A.; Balsamo, M.; Conte, R.; Benelli, R.; Minghelli, S.; et al. Melanoma cells inhibit natural killer cell function by modulating the expression of activating receptors and cytolytic activity. Cancer Res. 2012, 72, 1407–1415. [Google Scholar] [CrossRef] [PubMed]
- Konjevic, G.; Mirjacic Martinovic, K.; Vuletic, A.; Jovic, V.; Jurisic, V.; Babovic, N.; Spuzic, I. Low expression of CD161 and NKG2D activating NK receptor is associated with impaired NK cell cytotoxicity in metastatic melanoma patients. Clin. Exp. Metastasis 2007, 24, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Velazquez, N.G.; Torres-Ramos, Y.D.; Munoz-Sanchez, J.L.; Espinosa-Godoy, L.; Gomez-Cortes, S.; Moreno, J.; Moreno-Eutimio, M.A. Altered Expression of Natural Cytotoxicity Receptors and NKG2D on Peripheral Blood NK Cell Subsets in Breast Cancer Patients. Transl. Oncol. 2016, 9, 384–391. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Xu, J.; Huang, Q.; Huang, M.; Wen, H.; Zhang, C.; Wang, J.; Song, J.; Zheng, M.; Sun, H.; et al. High NKG2A expression contributes to NK cell exhaustion and predicts a poor prognosis of patients with liver cancer. Oncoimmunology 2017, 6, e1264562. [Google Scholar] [CrossRef]
- Fauriat, C.; Moretta, A.; Olive, D.; Costello, R.T. Defective killing of dendritic cells by autologous natural killer cells from acute myeloid leukemia patients. Blood 2005, 106, 2186–2188. [Google Scholar] [CrossRef]
- Vitale, M.; Della Chiesa, M.; Carlomagno, S.; Pende, D.; Arico, M.; Moretta, L.; Moretta, A. NK-dependent DC maturation is mediated by TNFalpha and IFNgamma released upon engagement of the NKp30 triggering receptor. Blood 2005, 106, 566–571. [Google Scholar] [CrossRef]
- Anderson, A.C.; Joller, N.; Kuchroo, V.K. Lag-3, Tim-3, and TIGIT: Co-Inhibitory Receptors with Specialized Functions in Immune Regulation. Immunity 2016, 44, 989–1004. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, I.P.; Gallois, A.; Jimenez-Baranda, S.; Khan, S.; Anderson, A.C.; Kuchroo, V.K.; Osman, I.; Bhardwaj, N. Reversal of NK-cell exhaustion in advanced melanoma by Tim-3 blockade. Cancer Immunol. Res. 2014, 2, 410–422. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Xia, P.; Du, Y.; Liu, S.; Huang, G.; Chen, J.; Zhang, H.; Hou, N.; Cheng, X.; Zhou, L.; et al. T-cell immunoglobulin and ITIM domain (TIGIT) receptor/poliovirus receptor (PVR) ligand engagement suppresses interferon-gamma production of natural killer cells via beta-arrestin 2-mediated negative signaling. J. Biol. Chem. 2014, 289, 17647–17657. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhang, H.; Li, M.; Hu, D.; Li, C.; Ge, B.; Jin, B.; Fan, Z. Recruitment of Grb2 and SHIP1 by the ITT-like motif of TIGIT suppresses granule polarization and cytotoxicity of NK cells. Cell Death Differ. 2013, 20, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Stanietsky, N.; Rovis, T.L.; Glasner, A.; Seidel, E.; Tsukerman, P.; Yamin, R.; Enk, J.; Jonjic, S.; Mandelboim, O. Mouse TIGIT inhibits NK-cell cytotoxicity upon interaction with PVR. Eur. J. Immunol. 2013, 43, 2138–2150. [Google Scholar] [CrossRef] [PubMed]
- Stanietsky, N.; Simic, H.; Arapovic, J.; Toporik, A.; Levy, O.; Novik, A.; Levine, Z.; Beiman, M.; Dassa, L.; Achdout, H.; et al. The interaction of TIGIT with PVR and PVRL2 inhibits human NK cell cytotoxicity. Proc. Natl. Acad. Sci. USA 2009, 106, 17858–17863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degos, C.; Heinemann, M.; Barrou, J.; Boucherit, N.; Lambaudie, E.; Savina, A.; Gorvel, L.; Olive, D. Endometrial Tumor Microenvironment Alters Human NK Cell Recruitment, and Resident NK Cell Phenotype and Function. Front. Immunol. 2019, 10, 877. [Google Scholar] [CrossRef]
- Mamessier, E.; Pradel, L.C.; Thibult, M.L.; Drevet, C.; Zouine, A.; Jacquemier, J.; Houvenaeghel, G.; Bertucci, F.; Birnbaum, D.; Olive, D. Peripheral blood NK cells from breast cancer patients are tumor-induced composite subsets. J. Immunol. 2013, 190, 2424–2436. [Google Scholar] [CrossRef]
- Castriconi, R.; Cantoni, C.; Della Chiesa, M.; Vitale, M.; Marcenaro, E.; Conte, R.; Biassoni, R.; Bottino, C.; Moretta, L.; Moretta, A. Transforming growth factor beta 1 inhibits expression of NKp30 and NKG2D receptors: Consequences for the NK-mediated killing of dendritic cells. Proc. Natl. Acad. Sci. USA 2003, 100, 4120–4125. [Google Scholar] [CrossRef]
- Lee, J.C.; Lee, K.M.; Kim, D.W.; Heo, D.S. Elevated TGF-beta1 secretion and down-modulation of NKG2D underlies impaired NK cytotoxicity in cancer patients. J. Immunol. 2004, 172, 7335–7340. [Google Scholar] [CrossRef]
- Spaggiari, G.M.; Capobianco, A.; Abdelrazik, H.; Becchetti, F.; Mingari, M.C.; Moretta, L. Mesenchymal stem cells inhibit natural killer-cell proliferation, cytotoxicity, and cytokine production: Role of indoleamine 2,3-dioxygenase and prostaglandin E2. Blood 2008, 111, 1327–1333. [Google Scholar] [CrossRef] [PubMed]
- Della Chiesa, M.; Carlomagno, S.; Frumento, G.; Balsamo, M.; Cantoni, C.; Conte, R.; Moretta, L.; Moretta, A.; Vitale, M. The tryptophan catabolite L-kynurenine inhibits the surface expression of NKp46- and NKG2D-activating receptors and regulates NK-cell function. Blood 2006, 108, 4118–4125. [Google Scholar] [CrossRef] [PubMed]
- Balsamo, M.; Scordamaglia, F.; Pietra, G.; Manzini, C.; Cantoni, C.; Boitano, M.; Queirolo, P.; Vermi, W.; Facchetti, F.; Moretta, A.; et al. Melanoma-associated fibroblasts modulate NK cell phenotype and antitumor cytotoxicity. Proc. Natl. Acad. Sci. USA 2009, 106, 20847–20852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, A.I.; Thoren, F.B.; Brune, M.; Hellstrand, K. NKp46 and NKG2D receptor expression in NK cells with CD56dim and CD56bright phenotype: Regulation by histamine and reactive oxygen species. Br. J. Haematol. 2006, 132, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Han, Y.; Guo, Q.; Zhang, M.; Cao, X. Cancer-expanded myeloid-derived suppressor cells induce anergy of NK cells through membrane-bound TGF-beta 1. J. Immunol. 2009, 182, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Correa, B.; Bergua, J.M.; Pera, A.; Campos, C.; Arcos, M.J.; Banas, H.; Duran, E.; Solana, R.; Tarazona, R. In Vitro Culture with Interleukin-15 Leads to Expression of Activating Receptors and Recovery of Natural Killer Cell Function in Acute Myeloid Leukemia Patients. Front Immunol. 2017, 8, 931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keskin, D.B.; Allan, D.S.; Rybalov, B.; Andzelm, M.M.; Stern, J.N.; Kopcow, H.D.; Koopman, L.A.; Strominger, J.L. TGFbeta promotes conversion of CD16+ peripheral blood NK cells into CD16- NK cells with similarities to decidual NK cells. Proc. Natl. Acad. Sci. USA 2007, 104, 3378–3383. [Google Scholar] [CrossRef]
- Cerdeira, A.S.; Rajakumar, A.; Royle, C.M.; Lo, A.; Husain, Z.; Thadhani, R.I.; Sukhatme, V.P.; Karumanchi, S.A.; Kopcow, H.D. Conversion of peripheral blood NK cells to a decidual NK-like phenotype by a cocktail of defined factors. J. Immunol. 2013, 190, 3939–3948. [Google Scholar] [CrossRef]
- Bruno, A.; Focaccetti, C.; Pagani, A.; Imperatori, A.S.; Spagnoletti, M.; Rotolo, N.; Cantelmo, A.R.; Franzi, F.; Capella, C.; Ferlazzo, G.; et al. The proangiogenic phenotype of natural killer cells in patients with non-small cell lung cancer. Neoplasia 2013, 15, 133–142. [Google Scholar] [CrossRef]
- Bruno, A.; Bassani, B.; D’Urso, D.G.; Pitaku, I.; Cassinotti, E.; Pelosi, G.; Boni, L.; Dominioni, L.; Noonan, D.M.; Mortara, L.; et al. Angiogenin and the MMP9-TIMP2 axis are up-regulated in proangiogenic, decidual NK-like cells from patients with colorectal cancer. FASEB J. 2018, 32, 5365–5377. [Google Scholar] [CrossRef]
- Gao, Y.; Souza-Fonseca-Guimaraes, F.; Bald, T.; Ng, S.S.; Young, A.; Ngiow, S.F.; Rautela, J.; Straube, J.; Waddell, N.; Blake, S.J.; et al. Tumor immunoevasion by the conversion of effector NK cells into type 1 innate lymphoid cells. Nat. Immunol. 2017, 18, 1004–1015. [Google Scholar] [CrossRef] [PubMed]
- Cortez, V.S.; Ulland, T.K.; Cervantes-Barragan, L.; Bando, J.K.; Robinette, M.L.; Wang, Q.; White, A.J.; Gilfillan, S.; Cella, M.; Colonna, M. SMAD4 impedes the conversion of NK cells into ILC1-like cells by curtailing non-canonical TGF-beta signaling. Nat. Immunol. 2017, 18, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Bruno, A.; Ferlazzo, G.; Albini, A.; Noonan, D.M. A think tank of TINK/TANKs: Tumor-infiltrating/tumor-associated natural killer cells in tumor progression and angiogenesis. J. Natl. Cancer Inst. 2014, 106, dju200. [Google Scholar] [CrossRef] [PubMed]
- Bosi, A.; Zanellato, S.; Bassani, B.; Albini, A.; Musco, A.; Cattoni, M.; Desio, M.; Nardecchia, E.; D’Urso, D.G.; Imperatori, A.; et al. Natural Killer Cells from Malignant Pleural Effusion Are Endowed with a Decidual-Like Proangiogenic Polarization. J. Immunol. Res. 2018, 2018, 2438598. [Google Scholar] [CrossRef] [PubMed]
- Colonna, M. Innate Lymphoid Cells: Diversity, Plasticity, and Unique Functions in Immunity. Immunity 2018, 48, 1104–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivier, E.; Artis, D.; Colonna, M.; Diefenbach, A.; Di Santo, J.P.; Eberl, G.; Koyasu, S.; Locksley, R.M.; McKenzie, A.N.J.; Mebius, R.E.; et al. Innate Lymphoid Cells: 10 Years On. Cell 2018, 174, 1054–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasteiger, G.; Fan, X.; Dikiy, S.; Lee, S.Y.; Rudensky, A.Y. Tissue residency of innate lymphoid cells in lymphoid and nonlymphoid organs. Science 2015, 350, 981–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinette, M.L.; Fuchs, A.; Cortez, V.S.; Lee, J.S.; Wang, Y.; Durum, S.K.; Gilfillan, S.; Colonna, M.; Immunological Genome Consortium. Transcriptional programs define molecular characteristics of innate lymphoid cell classes and subsets. Nat. Immunol. 2015, 16, 306–317. [Google Scholar] [CrossRef]
- Sojka, D.K.; Plougastel-Douglas, B.; Yang, L.; Pak-Wittel, M.A.; Artyomov, M.N.; Ivanova, Y.; Zhong, C.; Chase, J.M.; Rothman, P.B.; Yu, J.; et al. Tissue-resident natural killer (NK) cells are cell lineages distinct from thymic and conventional splenic NK cells. eLife 2014, 3, e01659. [Google Scholar] [CrossRef]
- Daussy, C.; Faure, F.; Mayol, K.; Viel, S.; Gasteiger, G.; Charrier, E.; Bienvenu, J.; Henry, T.; Debien, E.; Hasan, U.A.; et al. T-bet and Eomes instruct the development of two distinct natural killer cell lineages in the liver and in the bone marrow. J. Exp. Med. 2014, 211, 563–577. [Google Scholar] [CrossRef]
- Klose, C.S.N.; Flach, M.; Mohle, L.; Rogell, L.; Hoyler, T.; Ebert, K.; Fabiunke, C.; Pfeifer, D.; Sexl, V.; Fonseca-Pereira, D.; et al. Differentiation of type 1 ILCs from a common progenitor to all helper-like innate lymphoid cell lineages. Cell 2014, 157, 340–356. [Google Scholar] [CrossRef] [PubMed]
- Cortez, V.S.; Cervantes-Barragan, L.; Robinette, M.L.; Bando, J.K.; Wang, Y.; Geiger, T.L.; Gilfillan, S.; Fuchs, A.; Vivier, E.; Sun, J.C.; et al. Transforming Growth Factor-beta Signaling Guides the Differentiation of Innate Lymphoid Cells in Salivary Glands. Immunity 2016, 44, 1127–1139. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, T.E.; Rapp, M.; Fan, X.; Weizman, O.E.; Bhardwaj, P.; Adams, N.M.; Walzer, T.; Dannenberg, A.J.; Sun, J.C. Adipose-Resident Group 1 Innate Lymphoid Cells Promote Obesity-Associated Insulin Resistance. Immunity 2016, 45, 428–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dadi, S.; Chhangawala, S.; Whitlock, B.M.; Franklin, R.A.; Luo, C.T.; Oh, S.A.; Toure, A.; Pritykin, Y.; Huse, M.; Leslie, C.S.; et al. Cancer Immunosurveillance by Tissue-Resident Innate Lymphoid Cells and Innate-like T Cells. Cell 2016, 164, 365–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, A.; Vermi, W.; Lee, J.S.; Lonardi, S.; Gilfillan, S.; Newberry, R.D.; Cella, M.; Colonna, M. Intraepithelial type 1 innate lymphoid cells are a unique subset of IL-12- and IL-15-responsive IFN-gamma-producing cells. Immunity 2013, 38, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Bernink, J.H.; Peters, C.P.; Munneke, M.; te Velde, A.A.; Meijer, S.L.; Weijer, K.; Hreggvidsdottir, H.S.; Heinsbroek, S.E.; Legrand, N.; Buskens, C.J.; et al. Human type 1 innate lymphoid cells accumulate in inflamed mucosal tissues. Nat. Immunol. 2013, 14, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Simoni, Y.; Fehlings, M.; Kloverpris, H.N.; McGovern, N.; Koo, S.L.; Loh, C.Y.; Lim, S.; Kurioka, A.; Fergusson, J.R.; Tang, C.L.; et al. Human Innate Lymphoid Cell Subsets Possess Tissue-Type Based Heterogeneity in Phenotype and Frequency. Immunity 2017, 46, 148–161. [Google Scholar] [CrossRef] [PubMed]
- Constantinides, M.G.; McDonald, B.D.; Verhoef, P.A.; Bendelac, A. A committed precursor to innate lymphoid cells. Nature 2014, 508, 397–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, S.M.; Chaix, J.; Rupp, L.J.; Wu, J.; Madera, S.; Sun, J.C.; Lindsten, T.; Reiner, S.L. The transcription factors T-bet and Eomes control key checkpoints of natural killer cell maturation. Immunity 2012, 36, 55–67. [Google Scholar] [CrossRef]
- Weizman, O.E.; Adams, N.M.; Schuster, I.S.; Krishna, C.; Pritykin, Y.; Lau, C.; Degli-Esposti, M.A.; Leslie, C.S.; Sun, J.C.; O’Sullivan, T.E. ILC1 Confer Early Host Protection at Initial Sites of Viral Infection. Cell 2017, 171, 795–808. [Google Scholar] [CrossRef]
- Lopez-Soto, A.; Gonzalez, S.; Smyth, M.J.; Galluzzi, L. Control of Metastasis by NK Cells. Cancer Cell 2017, 32, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Cong, J.; Wang, X.; Zheng, X.; Wang, D.; Fu, B.; Sun, R.; Tian, Z.; Wei, H. Dysfunction of Natural Killer Cells by FBP1-Induced Inhibition of Glycolysis during Lung Cancer Progression. Cell Metab. 2018, 28, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Iguchi-Manaka, A.; Kai, H.; Yamashita, Y.; Shibata, K.; Tahara-Hanaoka, S.; Honda, S.; Yasui, T.; Kikutani, H.; Shibuya, K.; Shibuya, A. Accelerated tumor growth in mice deficient in DNAM-1 receptor. J. Exp. Med. 2008, 205, 2959–2964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerra, N.; Tan, Y.X.; Joncker, N.T.; Choy, A.; Gallardo, F.; Xiong, N.; Knoblaugh, S.; Cado, D.; Greenberg, N.M.; Raulet, D.H. NKG2D-deficient mice are defective in tumor surveillance in models of spontaneous malignancy. Immunity 2008, 28, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Waldhauer, I.; Steinle, A. NK cells and cancer immunosurveillance. Oncogene 2008, 27, 5932–5943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rautela, J.; Dagley, L.F.; de Oliveira, C.C.; Schuster, I.S.; Hediyeh-Zadeh, S.; Delconte, R.B.; Cursons, J.; Hennessy, R.; Hutchinson, D.S.; Harrison, C.; et al. Therapeutic blockade of activin-A improves NK cell function and antitumor immunity. Sci. Signal. 2019, 12. [Google Scholar] [CrossRef] [PubMed]
- Crome, S.Q.; Nguyen, L.T.; Lopez-Verges, S.; Yang, S.Y.; Martin, B.; Yam, J.Y.; Johnson, D.J.; Nie, J.; Pniak, M.; Yen, P.H.; et al. A distinct innate lymphoid cell population regulates tumor-associated T cells. Nat. Med. 2017, 23, 368–375. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.J.; Song, B.; Kim, Y.S.; Kim, E.K.; Lee, J.M.; Lee, G.E.; Kim, J.O.; Kim, Y.J.; Chang, W.S.; Kang, C.Y. Tumor microenvironmental conversion of natural killer cells into myeloid-derived suppressor cells. Cancer Res. 2013, 73, 5669–5681. [Google Scholar] [CrossRef]
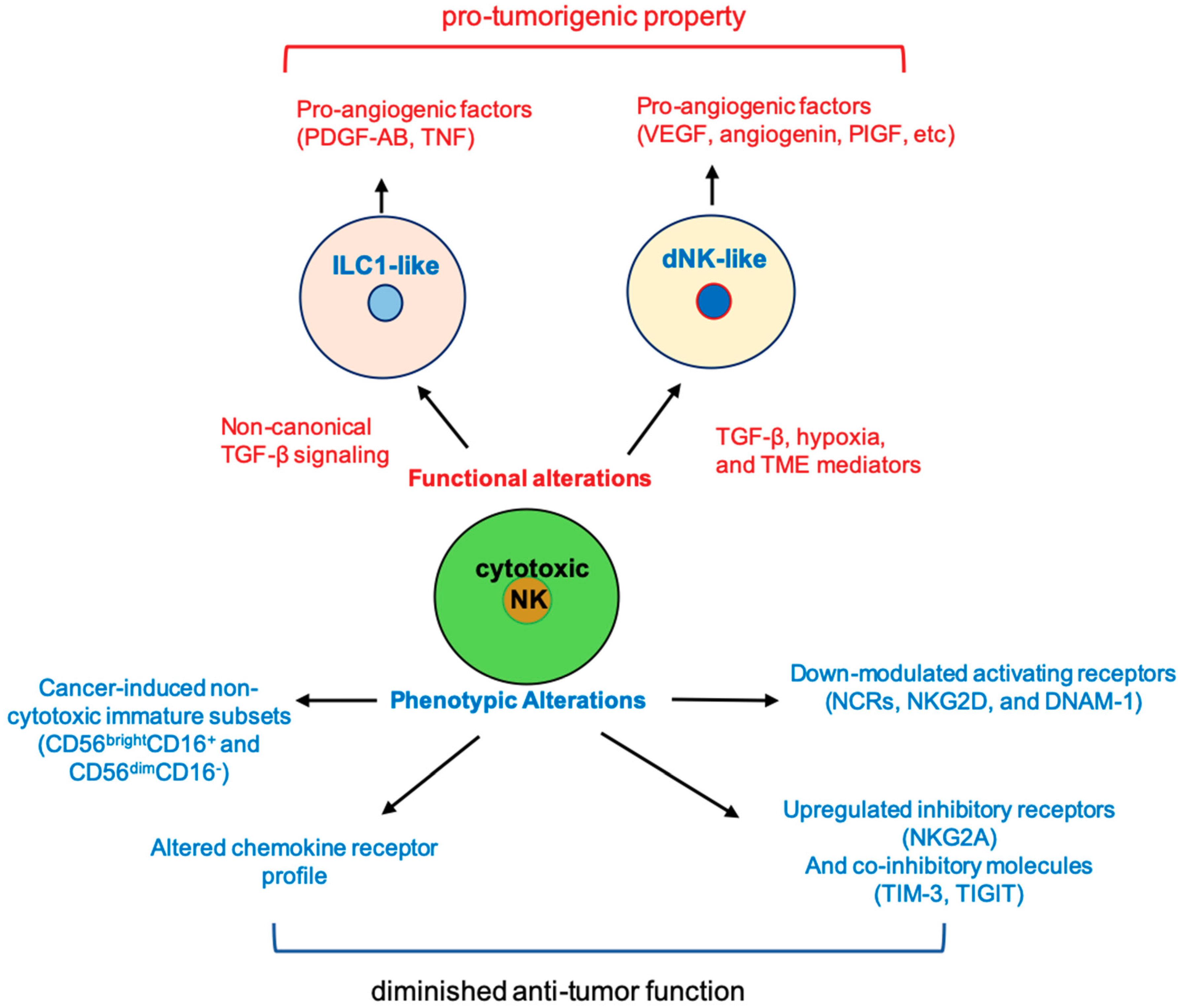
Figure 1.
Cancer-induced phenotypic and functional alterations in natural killer (NK) cells. The function and phenotype of cytotoxic NK cells can be altered by tumor environment cues to become pro-tumorigenic or to lose anti-tumor function. PIGF, placental growth factor; ILC1, type I innate lymphoid cells; TME, tumor microenvironment; dNK, decidual NK; NCR, natural cytotoxicity receptor; DNAM, DNAX accessory molecule.
Figure 1.
Cancer-induced phenotypic and functional alterations in natural killer (NK) cells. The function and phenotype of cytotoxic NK cells can be altered by tumor environment cues to become pro-tumorigenic or to lose anti-tumor function. PIGF, placental growth factor; ILC1, type I innate lymphoid cells; TME, tumor microenvironment; dNK, decidual NK; NCR, natural cytotoxicity receptor; DNAM, DNAX accessory molecule.


{kind=link}
Table 1.
Pro-tumor conversions of natural killers (NKs).
| Type of Conversion | Mouse Models/Human Patients | Phenotype | Functions | Limitations |
|---|---|---|---|---|
| Conventional NK to decidual NK-like phenotype [99,100] | Human patients of non-small cell lung carcinoma and colorectal cancer | ↑↑↑ 1 CD49a and/or CD9 | ↑↑↑ Secretion of pro-angiogenic factors and enzymes associated with tissue remodeling | The actual contribution to tumor progression in vivo is hard to determine |
| NK to ILC1-like phenotype [101,102] | Mouse models of fibrosarcoma, melanoma, and prostate cancer | ↓↓↓ 1 Eomes and CD62L ↑↑↑ CD49a, CD69, DNAM-1 and TRAIL ↑↑↑ LAG-3, CD96, and CTLA-4 | ↑↑↑ Secretion of PDGF-AB and GM-CSF ↓↓↓ Secretion of RANTES | Human relevancy is unclear |
1 ↑↑↑—upregulated; ↓↓↓—downregulated.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, S.; Dhar, P.; Wu, J.D. NK Cell Plasticity in Cancer. J. Clin. Med. 2019, 8, 1492. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8091492
AMA Style
Liu S, Dhar P, Wu JD. NK Cell Plasticity in Cancer. Journal of Clinical Medicine. 2019; 8(9):1492. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8091492
Chicago/Turabian StyleLiu, Sizhe, Payal Dhar, and Jennifer D. Wu. 2019. "NK Cell Plasticity in Cancer" Journal of Clinical Medicine 8, no. 9: 1492. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8091492
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.