Indexes of Radicle are Sensitive and Effective for Assessing Copper and Zinc Tolerance in Germinating Seeds of Suaeda salsa


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Chemicals
2.3. Germination Experiment
2.4. Seedling Incubation
2.5. Data Analysis
3. Results
3.1. Germination
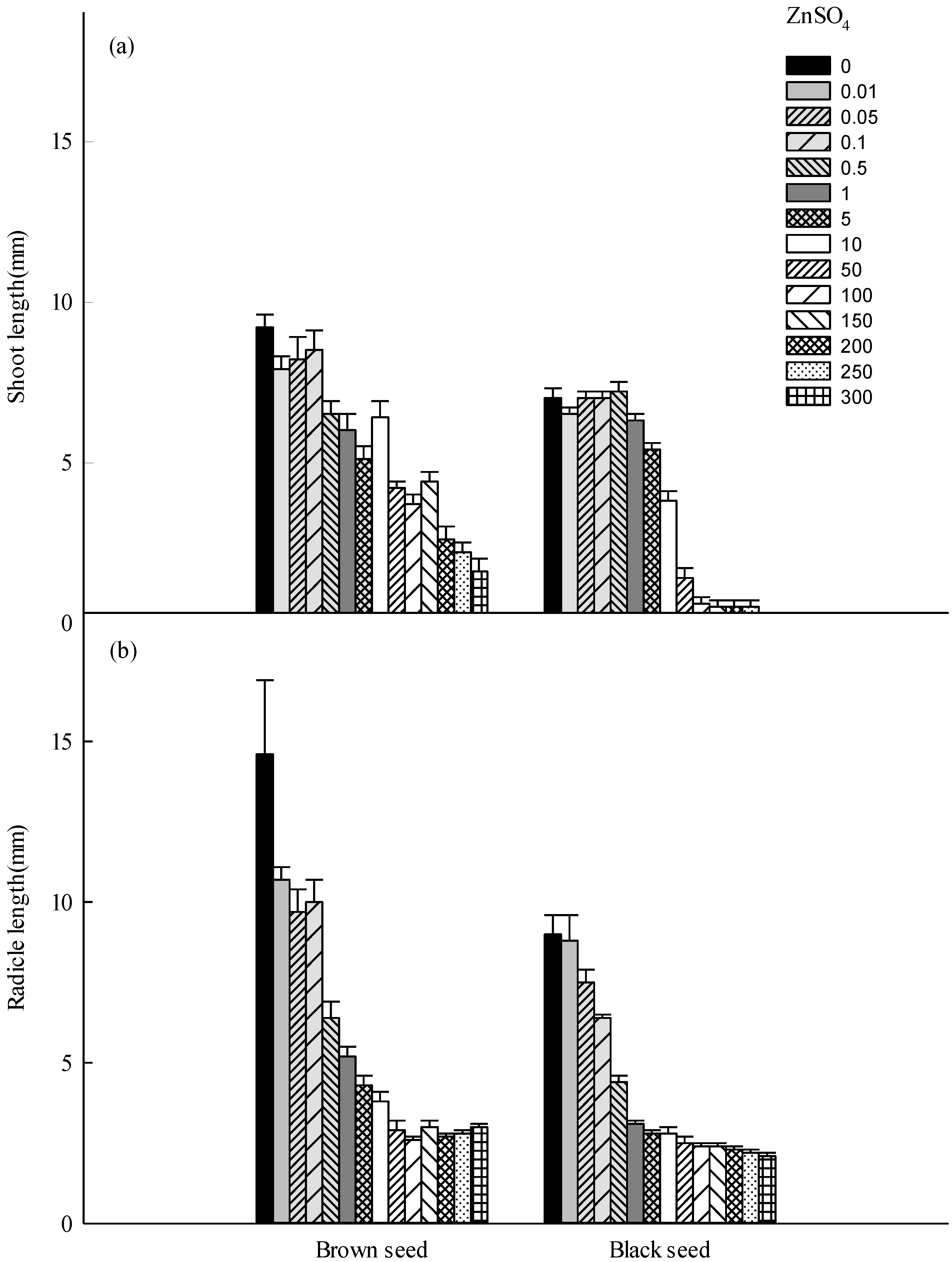
3.2. Seedling Growth
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hussain, S.; Khaliq, A.; Noor, M.A.; Tanveer, M.; Husssin, H.A.; Hussain, S.; Shah, T.; Mehmood, T. Metal toxicity and nitrogen metabolism in plants: An overview. In Carbon and Nitrogen Cycling in Soil; Datta, R., Meena, R.S., Pathan, S.I., Ceccherini, M.T., Eds.; Springer: Singapore, 2020; pp. 221–248. [Google Scholar]
- Muthusaravanan, S.; Sivarajasekar, N.; Vivek, J.S.; Paramasivan, T.; Naushad, M.; Prakashmaran, J.; Gayathri, V.; AI-Duaij, O.K. Phytoremediation of heavy metals: Mechanisms, methods and enhancements. Environ. Chem. Lett. 2018, 16, 1339–1359. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals-Concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef]
- Ghori, N.H.; Ghori, T.; Hayat, M.Q.; Imadi, S.R.; Gul, A.; Altay, V.; Ozturk, M. Heavy metal stress and responses in plants. Int. J. Environ. Sci. Technol. 2019, 16, 1807–1828. [Google Scholar] [CrossRef]
- He, J.; Ji, Z.X.; Wang, Q.Z.; Liu, C.F.; Zhou, Y.B. Effect of Cu and Pb pollution on the growth and antioxidant enzyme activity of Suaeda heteroptera. Ecol. Eng. 2016, 87, 102–109. [Google Scholar] [CrossRef]
- Nedjimi, B. Germination characteristics of Peganum harmala L. (Nitrariaceae) subjected to heavy metals: Implications for the use in polluted dryland restoration. Int. J. Environ. Sci. Technol. 2020, 17, 2113–2122. [Google Scholar] [CrossRef]
- Shang, C.L.; Wang, L.; Tian, C.Y.; Song, J. Heavy metal tolerance and potential for remediation of heavy metal-contaminated saline soils for the euhalophyte Suaeda salsa. Plant Signal. Behav. 2020. [Google Scholar] [CrossRef]
- Wang, H.L.; Tian, C.Y.; Jiang, L.; Wang, L. Remediation of heavy metals contaminated saline soils: A halophyte choice? Environ. Sci. Technol. 2014, 48, 21–22. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Maggio, A. Functional biology of halophytes in the phytoremediation of heavy metal contaminated soils. Environ. Exp. Bot. 2015, 111, 135–146. [Google Scholar] [CrossRef]
- Gutterman, Y. Survival Strategies of Annual Desert Plants; Springer: Berlin, Germany, 2002. [Google Scholar]
- Ye, N.; Li, H.; Zhu, G.; Liu, R.; Xu, W.; Jing, Y.; Peng, X.; Zhang, J. Copper suppresses abscisic acid catabolism and catalase activity, and inhibits seed germination of rice. Plant Cell Physiol. 2014, 55, 2008–2016. [Google Scholar] [CrossRef] [Green Version]
- Pokorska-Niewiada, K.; Rajkowska-Myśliwiec, M.; Protasowicki, M. Acute lethal toxicity of heavy metals to the seeds of plants of high importance to humans. Bull. Environ. Contam. Toxicol. 2018, 101, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Khan, M.A.; Yamaguchi, S.; Kamiya, Y. Effects of heavy metals on seed germination and early seedling growth of Arabidopsis thaliana. Plant Growth Regul. 2005, 46, 45–50. [Google Scholar] [CrossRef]
- Guterres, J.; Rossato, L.; Doley, D.; Pudmenzky, A.; Bee, C.; Cobena, V. Assessing germination characteristics of Australian native plant species in metal/metalloid solution. J. Hazard. Mater. 2019, 364, 173–181. [Google Scholar] [CrossRef]
- Yáñez-Espinosa, L.; Briones-Gallardo, R.; Flores, J.; Álvarez del Castillo, E. Effect of heavy metals on seed germination and seedling development of Nama aff. stenophylla collected on the slope of a mine tailing dump. Int. J. Phytoremediation 2020. [Google Scholar] [CrossRef]
- Nanda, R.; Agrawal, V. Elucidation of zinc and copper induced oxidative stress, DNA damage and activation of defence system during seed germination in Cassia angustifolia Vahl. Environ. Exp. Bot. 2016, 125, 31–41. [Google Scholar] [CrossRef]
- Liu, X.; Shen, X.; Lai, Y.; Ji, K.; Sun, H.; Wang, Y.; Hou, C.; Zou, N.; Wan, J.; Yu, J. Toxicological proteomic responses of halophyte Suaeda salsa to lead and zinc. Ecotoxicol. Environ. Saf. 2016, 134, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Marschner, P. Marschner’s Mineral Nutrition of High Plants, 3rd ed.; Science Press: Beijing, China, 2013. [Google Scholar]
- Lange, B.; van der Ent, A.; Baker, A.J.M.; Echevarria, G.; Mahy, G.; Malaisse, F.; Meerts, P.; Pourret, O.; Verbruggen, N.; Faucon, M.P. Copper and cobalt accumulation in plants: A critical assessment of the current state of knowledge. New Phytol. 2016, 213, 537–551. [Google Scholar] [CrossRef] [PubMed]
- Balafrej, H.; Bogusz, D.; Triqui, Z.-E.A.; Guedira, A.; Bendaou, N.; Smouni, A.; Fahr, M. Zinc hyperaccumulation in plants: A review. Plants 2020, 9, 562. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Wang, B. Using euhalophytes to understand salt tolerance and to develop saline agriculture: Suaeda salsa as a promising model. Ann. Bot. 2015, 115, 541–553. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.L.; Song, N.N. Salinity-induced alterations in plant growth, antioxidant enzyme activities, and lead transportation and accumulation in Suaeda salsa: Implications for phytoremediation. Ecotoxicology 2019, 28, 520–527. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, L.; Zhang, K.; Tian, C.Y. Copper-induced similar changes in growth and physiological responses of plants grown from dimorphic seeds of Suaeda salsa. Pak. J. Bot. 2018, 50, 871–877. [Google Scholar]
- Song, J.; Fan, H.; Zhao, Y.; Jia, Y.; Du, X.; Wang, B. Effect of salinity on germination, seedling emergence, seedling growth and ion accumulation of a euhalophyte Suaeda salsa in an interidal zone and on saline inland. Aquat. Bot. 2008, 88, 331–337. [Google Scholar] [CrossRef]
- Liu, S.; Yang, C.; Xie, W.; Xia, C.; Fan, P. The effects of cadmium on germination and seedling growth of Suaeda salsa. Procedia Environ. Sci. 2012, 16, 293–298. [Google Scholar] [CrossRef] [Green Version]
- Wan, Z.; Gao, T.; Zhou, Y.; Wang, Y.; Chang, G.; Ju, T.; Yang, Y.; Zhang, Q. Seed germination, bud growth and heavy-metal accumulation of Suaeda salsa. Chin. J. Biotechnol. 2020, 36, 493–507, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Ajmal, K.M.; Gul, B.; Weber, D.J. Germination of dimorphic seeds of Suaeda moquinii under high salinity stress. Aust. J. Bot. 2001, 49, 185–192. [Google Scholar]
- Zhang, K.; Yao, L.; Zhang, Y.; Tao, J. Achene heteromorphism in Bidens pilosa (Asteraceae): Differences in germination and possible adaptive significance. AoB Plants 2019, 11, plz026. [Google Scholar] [CrossRef] [Green Version]
- Nisar, F.; Gul, B.; Khan, M.A.; Hameed, A. Heteromorphic seeds of coastal halophytes Arthrocnemum macrostachyum and A. indicum display differential patterns of hydrogen peroxide accumulation, lipid peroxidation and antioxidant activities under increasing salinity. Plant Physiol. Biochem. 2019, 144, 58–63. [Google Scholar] [CrossRef]
- Payamani, R.; Nosratti, I.; Amerian, M. Variations in the germination characteristics in response to environmental factors between the hairy and spiny seeds of hedge parsley (Torilis arvensis Huds.). Weed Biol. Manag. 2018, 18, 176–183. [Google Scholar] [CrossRef]
- Redondo-Gómez, S.; Mateos-Naranjo, E.; Cambrollé, J.; Luque, T.; Figueroa, M.E.; Davy, A.J. Carry-over of differential salt tolerance in plants grown from dimorphic seeds of Suaeda splendens. Ann. Bot. 2008, 102, 103–112. [Google Scholar] [CrossRef]
- van der Ent, A.; Baker, A.J.M.; Reeves, R.D.; Pollard, A.J.; Schat, H. Hyperaccumulators of metal and metalloid trace elements: Facts and fiction. Plant Soil 2013, 362, 319–334. [Google Scholar] [CrossRef]
- Boi, M.E.; Porceddu, M.; Cappai, G.; De Giudici, G.; Bacchetta, G. Effects of zinc and lead on seed germination of Helichrysum microphyllum subsp. tyrrhenicum, a metal-tolerant plant. Int. J. Environ. Sci. Technol. 2020, 17, 1917–1928. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Factor | B (SE) | β | p-Value |
|---|---|---|---|---|
| Germination percentage | Metal type | 8.607 (1.561) | 0.259 | <0.001 |
| Seed type | −10.036 (1.561) | −0.302 | <0.001 | |
| Solution concentration | −0.097 (0.008) | −0.598 | <0.001 | |
| Germination index | Metal type | 8.471 (1.41) | 0.199 | <0.001 |
| Seed type | −29.421 (1.41) | −0.692 | <0.001 | |
| Solution concentration | −0.102 (0.007) | −0.489 | <0.001 | |
| Radicle length | Metal type | 1.137 (0.209) | 0.143 | <0.001 |
| Seed type | −1.651 (0.209) | −0.208 | <0.001 | |
| Solution concentration | −0.016 (0.001) | −0.404 | <0.001 | |
| Shoot length | Metal type | 1.102 (0.14) | 0.158 | <0.001 |
| Seed type | −1.394 (0.14) | −0.201 | <0.001 | |
| Solution concentration | −0.024 (0.001) | −0.695 | <0.001 |
| Concentration (mM) | CuSO4 | ZnSO4 | ||
|---|---|---|---|---|
| Brown Seed | Black Seed | Brown Seed | Black Seed | |
| 0 | 100.0 ± 0.0 Aa | 74.0 ± 2.6 Bab | 100.0 ± 0.0 Aa | 82.0 ± 2.6 Ba |
| 0.01 | 100.0 ± 0.0 Aa | 71.0 ± 1.9B Abc | 99.0 ± 1.0 Aa | 72.0 ± 5.4 Ba |
| 0.05 | 96.0 ± 1.6 Aab | 77.0 ± 4.4 Bab | 100.0 ± 0.0 Aa | 68.0 ± 2.3 Ba |
| 0.1 | 99.0 ± 1.0 Aab | 73.0 ± 3.4 Bab | 98.0 ± 1.2 Aa | 69.0 ± 5.7 Ba |
| 0.5 | 100.0 ± 0.0 Aa | 83.0 ± 2.5 Ba | 99.0 ± 1.0 Aa | 81.0 ± 1.9 Ba |
| 1 | 98.0 ± 1.2 Aab | 73.0 ± 6.6 Bab | 99.0 ± 1.0 Aa | 80.0 ± 5.2 Ba |
| 5 | 81.0 ± 5.7 Ac | 70.0 ± 4.8 Aabc | 99.0 ± 1.0 Aa | 76.0 ± 0.0 Ba |
| 10 | 85.0 ± 3.8 Abc | 71.0 ± 4.1 Babc | 98.0 ± 1.2 Aa | 81.0 ± 5.5 Aa |
| 50 | 73.0 ± 4.4 Acd | 66.0 ± 5.3 Aabc | 78.0 ± 2.0 Ab | 77.0 ± 3.0 Aa |
| 100 | 61.0 ± 1.0 Ade | 57.0 ± 5.7 Abc | 70.0 ± 3.8 Abc | 82.0 ± 5.0 Aa |
| 150 | 55.0 ± 1.0 Be | 63.0 ± 1.0 Aabc | 77.0 ± 4.7 Ab | 82.0 ± 2.6 Aa |
| 200 | 55.0 ± 2.5 Ae | 51.0 ± 3.4 Ac | 69.0 ± 3.4 Abc | 79.0 ± 5.3 Aa |
| 250 | 63.0 ± 3.0 Ade | 64.0 ± 3.7 Aabc | 66.0 ± 4.2 Abc | 77.0 ± 1.9 Aa |
| 300 | 38.0 ± 4.8 Bf | 52.0 ± 2.3 Ac | 54.0 ± 10.5 Ac | 78.0 ± 4.8 Aa |
| Concentration(mM) | CuSO4 | ZnSO4 | ||
|---|---|---|---|---|
| Brown Seed | Black Seed | Brown Seed | Black Seed | |
| 0 | 97.6 ± 0.4 Aa | 49.0 ± 2.4B Abcd | 97.4 ± 0.2 Aa | 60.9 ± 2.2 Ba |
| 0.01 | 97.8 ± 0.1 Aa | 51.0 ± 1.0B Abc | 96.5 ± 1.1 Aa | 52.6 ± 4.3 Ba |
| 0.05 | 93.5 ± 1.2 A Ab | 56.2 ± 2.8 Bab | 98.0 ± 0.4 Aa | 50.5 ± 2.4 Ba |
| 0.1 | 96.0 ± 0.7 Aa | 50.3 ± 2.3 Babc | 95.8 ± 1.0 Aa | 51.1 ± 4.1 Ba |
| 0.5 | 98.2 ± 0.1 Aa | 61.1 ± 2.5 Ba | 96.0 ± 1.1 Aa | 58.7 ± 1.6 Ba |
| 1 | 96.5 ± 1.1 Aa | 51.9 ± 5.2B Ab | 96.9 ± 1.0 Aa | 58.2 ± 4.8 Ba |
| 5 | 78.1 ± 5.2 Ac | 49.8 ± 4.6B Abc | 97.2 ± 1.1 Aa | 56.9 ± 0.9 Ba |
| 10 | 81.2 ± 3.9 Abc | 47.5 ± 3.9B Abcd | 94.9 ± 1.5 Aa | 61.0 ± 3.9 Ba |
| 50 | 70.5 ± 4.1 Acd | 44.2 ± 3.7 Bbcde | 76.4 ± 2.2 Ab | 53.0 ± 3.0 Ba |
| 100 | 56.7 ± 1 Ae | 35.4 ± 5.5 Bbcde | 68.2 ± 3.9 Abc | 57.2 ± 3.9 Aa |
| 150 | 52.0 ± 0.7 Ae | 41.3 ± 0.5 Bde | 74.5 ± 4.1 Ab | 57.5 ± 2.0 Ba |
| 200 | 52.6 ± 2.3 Ae | 33.1 ± 1.8 Bde | 66.9 ± 3.5 Abc | 50.5 ± 2.9 Ba |
| 250 | 59.0 ± 2.7 Ade | 42.6 ± 2.2 Bbcde | 63.9 ± 3.9 Abc | 50.7 ± 1.3 Ba |
| 300 | 36.2 ± 4.3 Af | 30.9 ± 1.2 Ae | 52.1 ± 10.0 Ac | 53.9 ± 2.6 Aa |
| Concentration (mM) | CuSO4 | ZnSO4 | ||
|---|---|---|---|---|
| Brown Seed | Black Seed | Brown Seed | Black Seed | |
| 0 | 100.00 | 100.00 | 100.00 | 100.00 |
| 0.01 | 80.33 | 81.08 | 73.40 | 97.68 |
| 0.05 | 40.53 | 63.80 | 66.88 | 83.18 |
| 0.1 | 30.65 | 45.70 | 68.61 | 70.8 |
| 0.5 | 30.66 | 34.71 | 43.82 | 48.47 |
| 1 | 24.40 | 32.26 | 35.95 | 34.83 |
| 5 | 23.77 | 29.59 | 29.53 | 30.62 |
| 10 | 15.35 | 27.62 | 26.05 | 30.73 |
| 50 | 19.25 | 18.93 | 19.68 | 27.82 |
| 100 | 18.73 | 18.79 | 17.79 | 26.07 |
| 150 | 19.52 | 16.23 | 20.62 | 26.60 |
| 200 | 18.56 | 14.04 | 18.79 | 26.06 |
| 250 | 17.91 | 14.95 | 18.91 | 23.94 |
| 300 | 19.21 | 16.32 | 20.76 | 23.03 |
| Concentration (mM) | CuSO4 | ZnSO4 | ||
|---|---|---|---|---|
| Brown Seed | Black Seed | Brown Seed | Black Seed | |
| 0 | 100.00 | 100.00 | 100.00 | 100.00 |
| 0.01 | 110.38 | 96.15 | 86.03 | 95.08 |
| 0.05 | 95.48 | 91.51 | 88.74 | 99.15 |
| 0.1 | 93.58 | 102.76 | 92.82 | 100.06 |
| 0.5 | 72.58 | 87.00 | 71.07 | 102.76 |
| 1 | 58.40 | 81.77 | 65.50 | 90.03 |
| 5 | 37.51 | 33.47 | 54.97 | 76.59 |
| 10 | 33.28 | 7.61 | 69.47 | 53.57 |
| 50 | 21.58 | 0.00 | 45.73 | 19.57 |
| 100 | 15.96 | 0.00 | 40.69 | 8.16 |
| 150 | 0.00 | 0.00 | 47.44 | 7.73 |
| 200 | 0.00 | 0.00 | 28.13 | 7.55 |
| 250 | 0.00 | 0.00 | 23.56 | 7.42 |
| 300 | 0.00 | 0.00 | 17.39 | 0.00 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Jiang, L.; Tanveer, M.; Ma, J.; Zhao, Z.; Wang, L. Indexes of Radicle are Sensitive and Effective for Assessing Copper and Zinc Tolerance in Germinating Seeds of Suaeda salsa. Agriculture 2020, 10, 445. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10100445
Zhang H, Jiang L, Tanveer M, Ma J, Zhao Z, Wang L. Indexes of Radicle are Sensitive and Effective for Assessing Copper and Zinc Tolerance in Germinating Seeds of Suaeda salsa. Agriculture. 2020; 10(10):445. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10100445
Chicago/Turabian StyleZhang, Hao, Li Jiang, Mohsin Tanveer, Jinbiao Ma, Zhenyong Zhao, and Lei Wang. 2020. "Indexes of Radicle are Sensitive and Effective for Assessing Copper and Zinc Tolerance in Germinating Seeds of Suaeda salsa" Agriculture 10, no. 10: 445. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10100445