Microbiota Characterization of Agricultural Green Waste-Based Suppressive Composts Using Omics and Classic Approaches

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Composting Process and Sampling
2.2. Chemical and Biological Compost Characterization
2.3. Microbial DNA Isolation, Amplification and Sequencing
2.4. Computational Study
2.5. Statistical Analysis
3. Results
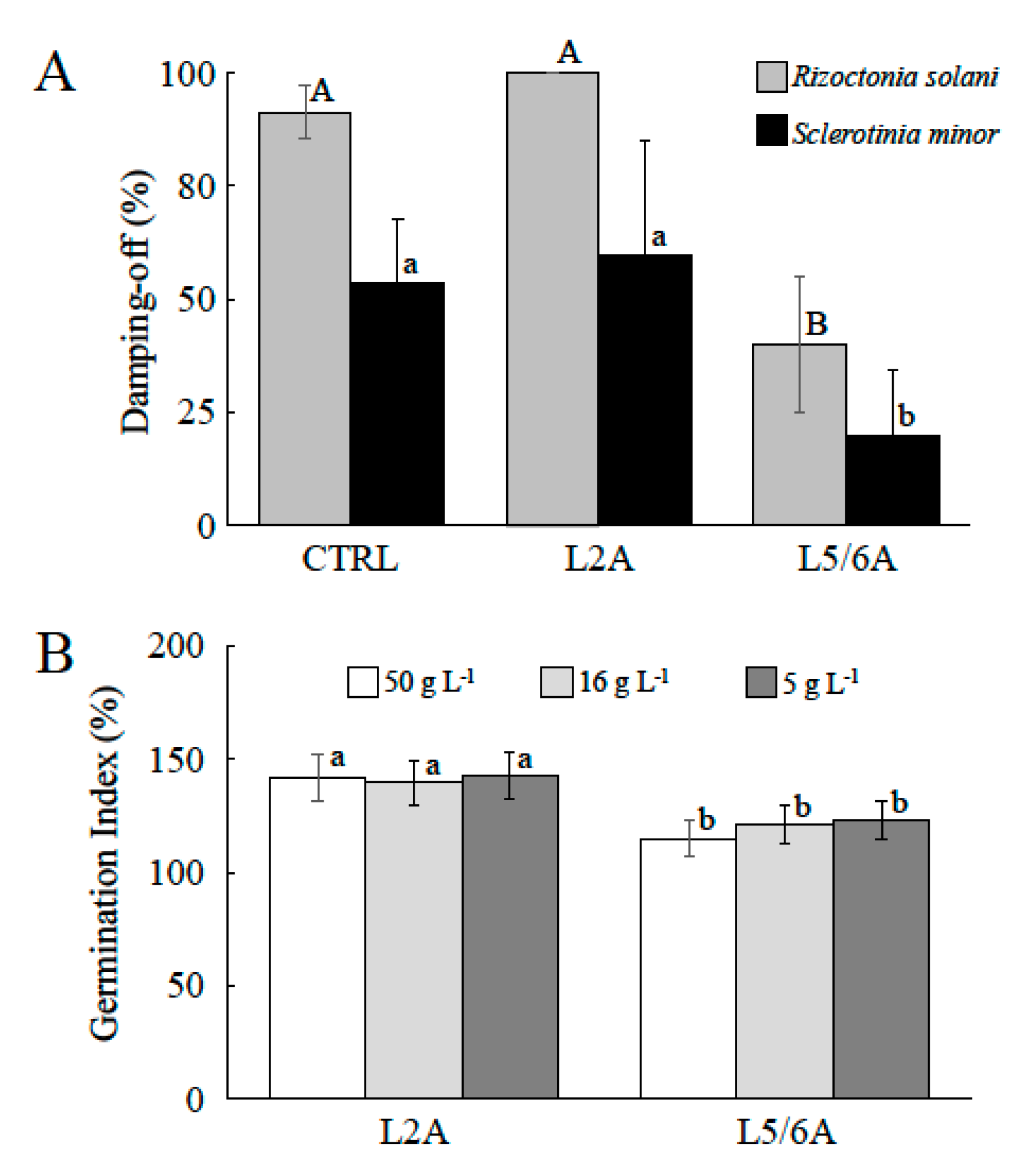
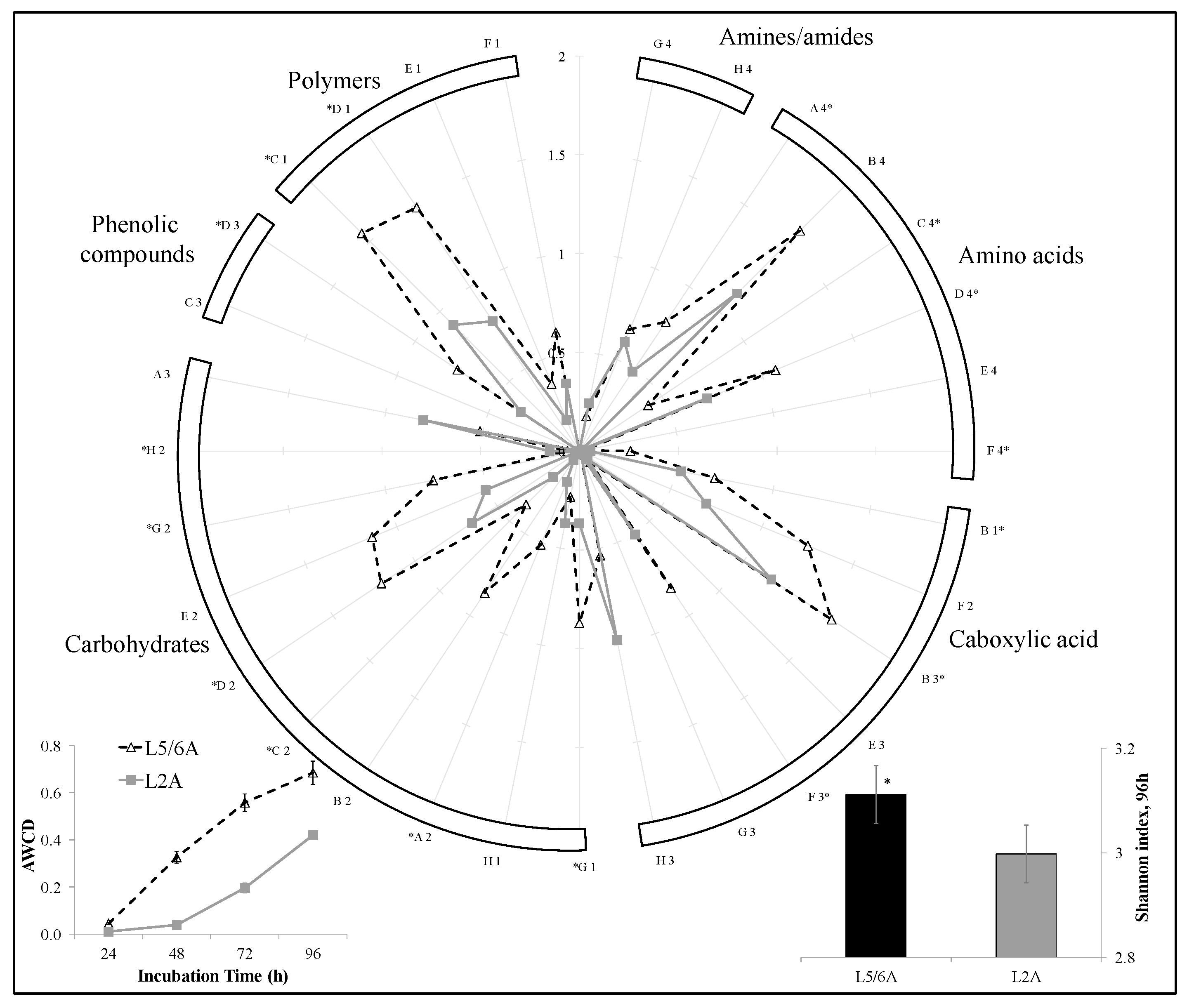
3.1. Compost Chemical and Biological Properties
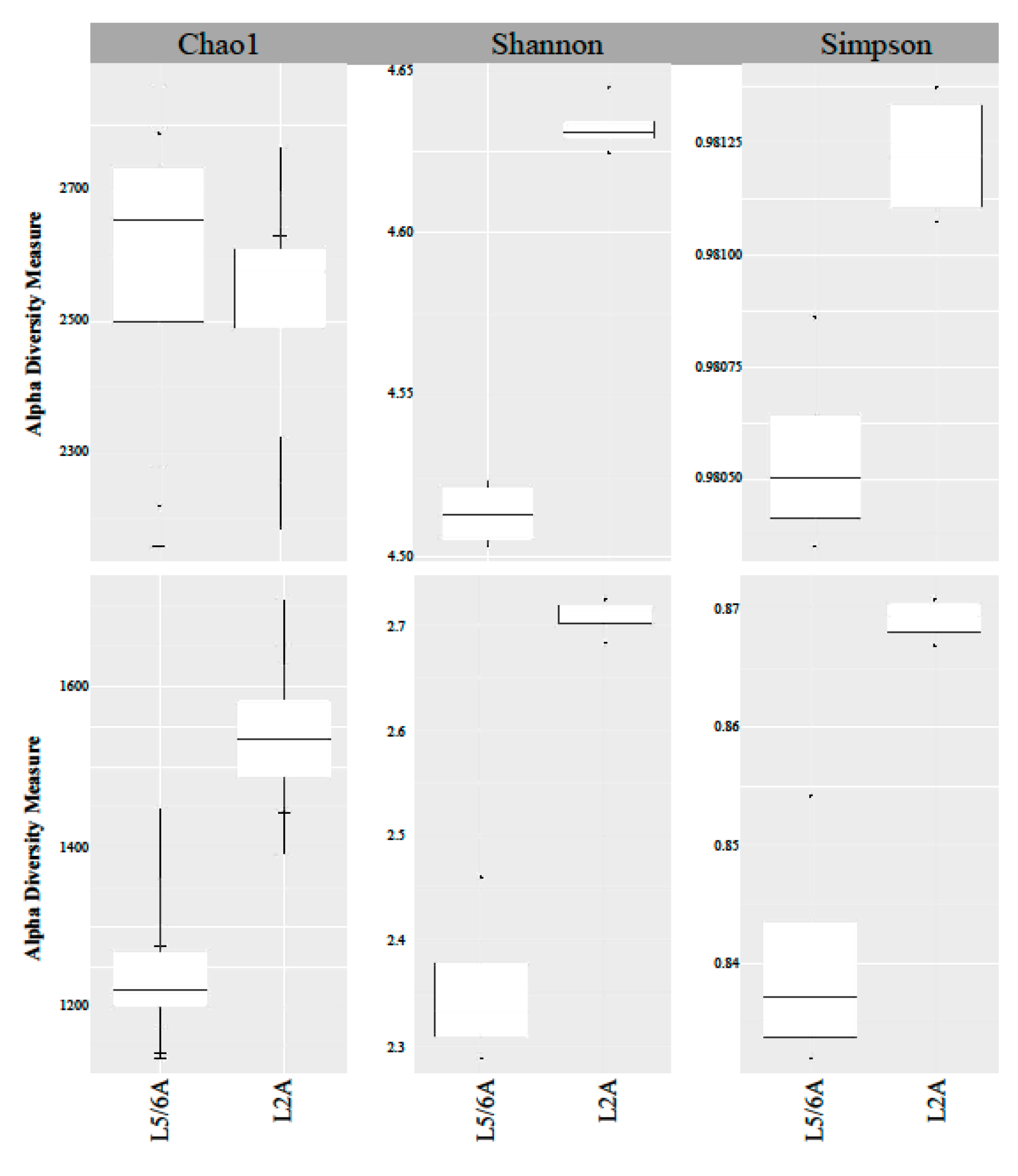
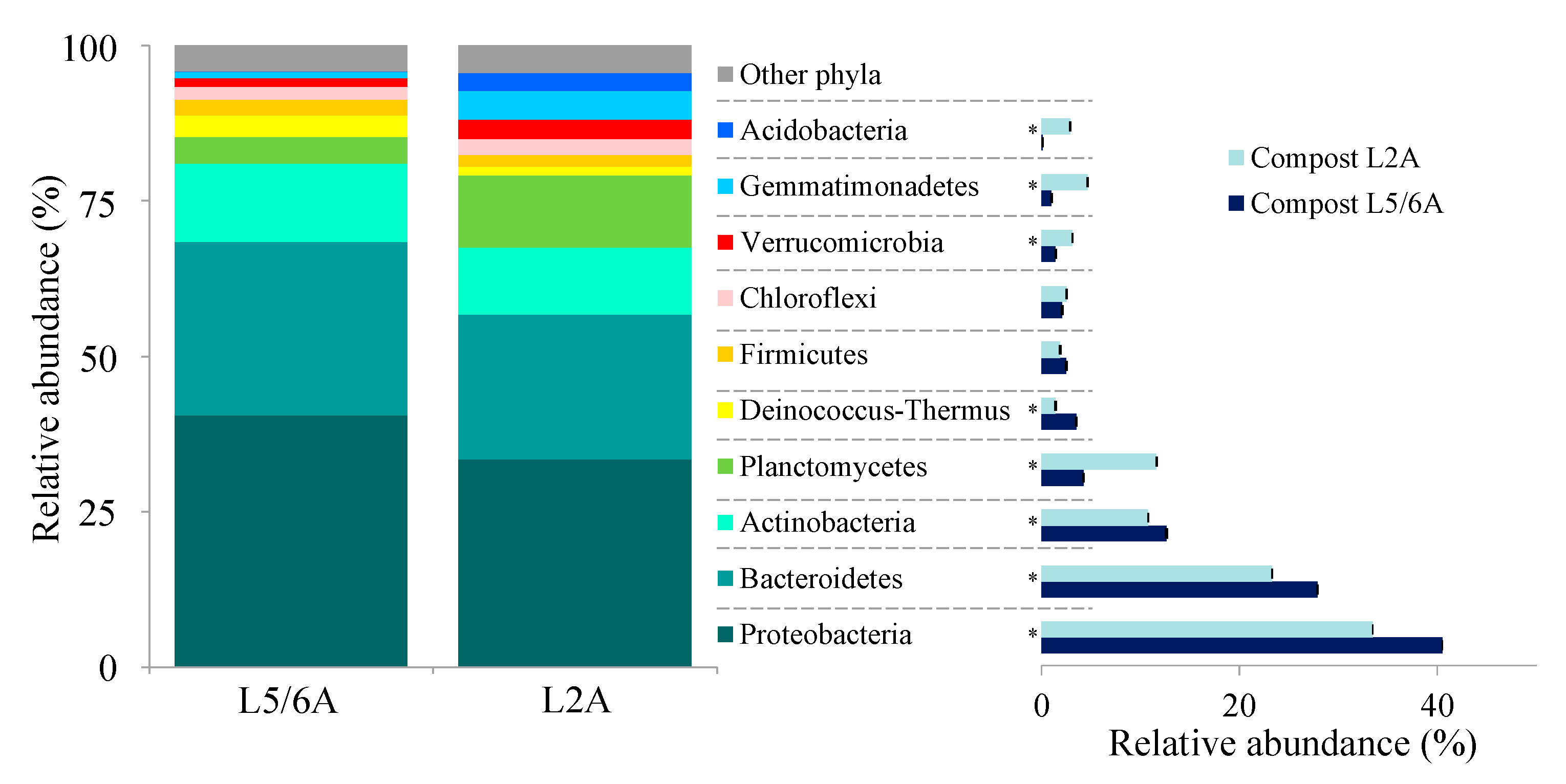
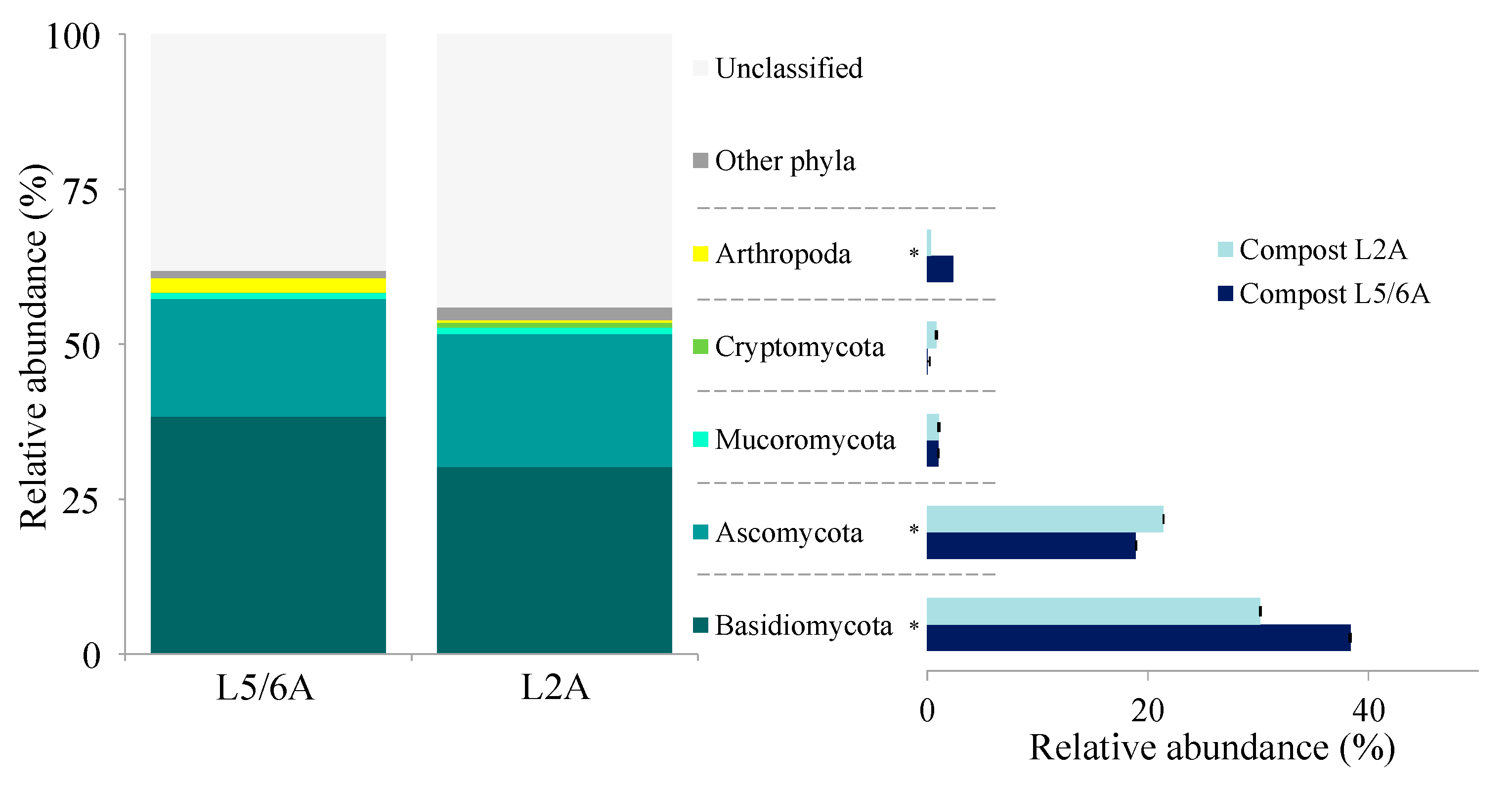
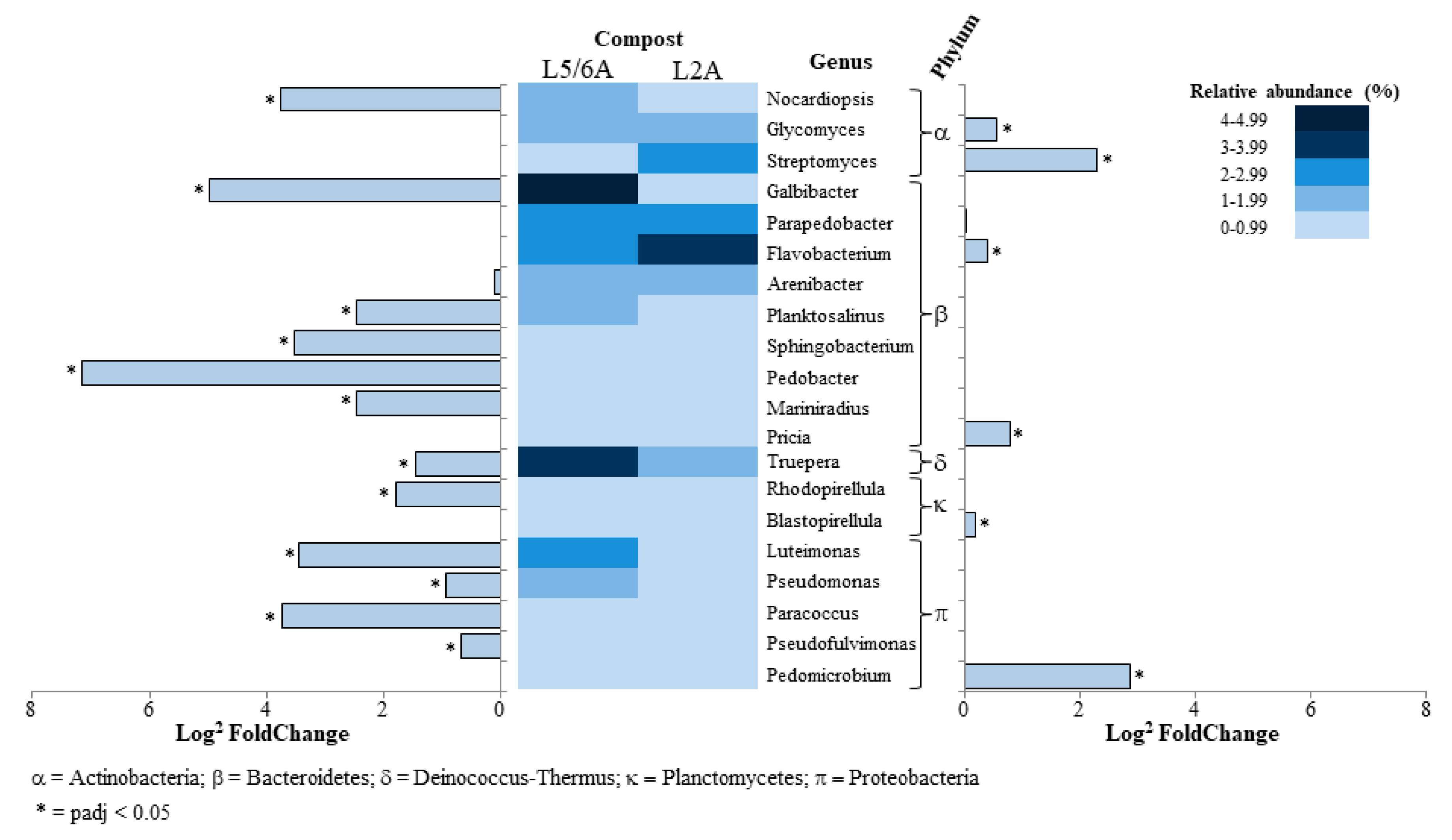
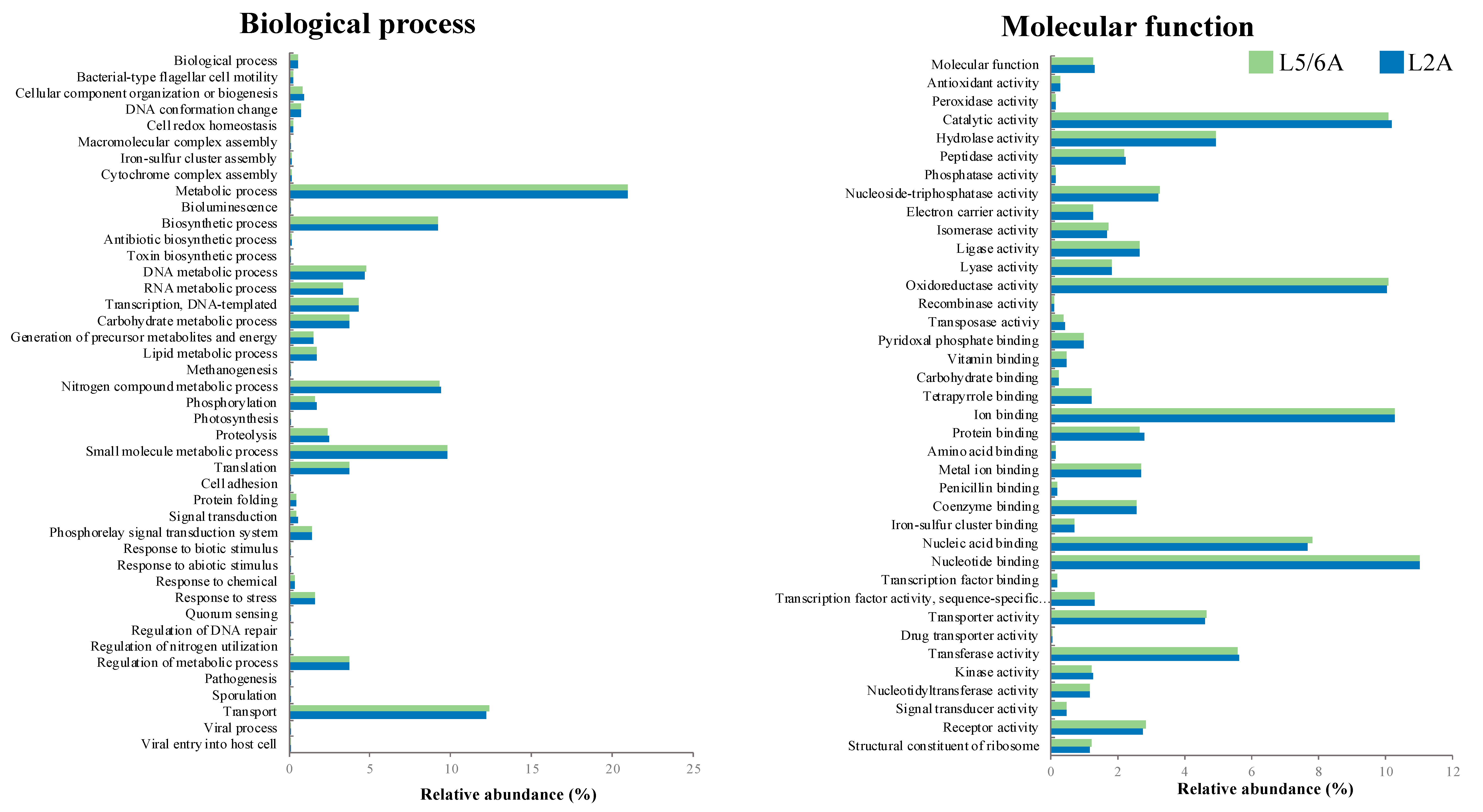
3.2. Compost Microbial Composition
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pane, C.; Piccolo, A.; Spaccini, R.; Celano, G.; Villecco, D.; Zaccardelli, M. Agricultural waste-based composts exhibiting suppressivity to diseases caused by the phytopathogenic soil-borne fungi Rhizoctonia solani and Sclerotinia minor. Appl. Soil Ecol. 2013, 65, 43–51. [Google Scholar] [CrossRef]
- Brito, L.M.; Mourão, I.; Coutinho, J.; Smith, S.R. Simple technologies for on-farm composting of cattle slurry solid fraction. Waste Manag. 2012, 32, 1332–1340. [Google Scholar] [CrossRef]
- Mehta, C.M.; Palni, U.; Franke-Whittle, I.H.; Sharma, A.K. Compost: Its role, mechanism and impact on reducing soil-borne plant diseases. Waste Manag. 2014, 34, 607–622. [Google Scholar] [CrossRef] [PubMed]
- Bonanomi, G.; Antignani, V.; Pane, C.; Scala, F. Suppression of soilborne fungal diseases with organic amendments. J. Plant Pathol. 2007, 14, 311–324. [Google Scholar]
- Pane, C.; Villecco, D.; Campanile, F.; Zaccardelli, M. Novel strains of Bacillus, isolated from compost and compost-amended soils, as biological control agents against soil-borne phytopathogenic fungi. Biocontrol Sci. Technol. 2012, 22, 1373–1388. [Google Scholar] [CrossRef]
- Zhang, W.; Dick, W.A.; Hoitink, H.A.J. Compost-induced systemic acquired resistance in cucumber to pythium root rot and anthracnoce. Phytopathology 1996, 86, 1066–1070. [Google Scholar] [CrossRef]
- Loffredo, E.; Senesi, N. In vitro and in vivo assessment of the potential of compost and its humic acid fraction to protect ornamental plants from soil-borne pathogenic fungi. Sci. Hortic. 2009, 122, 432–439. [Google Scholar] [CrossRef]
- Neher, D.A.; Weicht, T.R.; Bates, S.T.; Leff, J.W.; Fierer, N. Changes in Bacterial and Fungal Communities across Compost Recipes, Preparation Methods, and Composting Times. PLoS ONE 2013, 8, e79512. [Google Scholar] [CrossRef] [Green Version]
- Morales-Rodríguez, C.; Palo, C.; Palo, E.; Rodríguez-Molina, M.C. Control of Phytophthora nicotianae with Mefenoxam, Fresh Brassica Tissues, and Brassica Pellets. Plant Dis. 2014, 98, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Blaya, J.; Lloret, E.; Ros, M.; Pascual, J.A. Identification of predictor parameters to determine agro-industrial compost suppressiveness against Fusarium oxysporum and Phytophthora capsici diseases in muskmelon and pepper seedlings. J. Sci. Food Agric. 2015, 95, 1482–1490. [Google Scholar] [CrossRef]
- Blaya, J.; Lacasa, C.; Lacasa, A.; Martínez, V.; Santísima-Trinidad, A.B.; Pascual, J.A.; Ros, M. Characterization of Phytophthora nicotianae isolates in southeast Spain and their detection and quantification through a real-time TaqMan PCR. J. Sci. Food Agric. 2015, 95, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Sparks, D.L. Methods of Soil Analysis Part 3–Chemical Methods. Soil Sci. Soc. Am. Book Ser. 1996, 5, 1018–1020. [Google Scholar]
- Erhart, E.; Burian, K.; Hartl, W.; Stich, K. Suppression of Pythium ultimum by Biowaste Composts in Relation to Compost Microbial Biomass, Activity and Content of Phenolic Compounds. J. Phytopathol. 1999, 147, 299–305. [Google Scholar] [CrossRef]
- Bonanomi, G.; Sicurezza, M.G.; Caporaso, S.; Esposito, A.; Mazzoleni, S. Phytotoxicity dynamics of decaying plant materials. New Phytol. 2006, 169, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Pane, C.; Spaccini, R.; Piccolo, A.; Scala, F.; Bonanomi, G. Compost amendments enhance peat suppressiveness to Pythium ultimum, Rhizoctonia solani and Sclerotinia minor. Biol. Control 2011, 56, 115–124. [Google Scholar] [CrossRef]
- Veeken, A.H.M.; Blok, W.J.; Curci, F.; Coenen, G.C.M.; Termorshuizen, A.J.; Hamelers, H.V.M. Improving quality of composted biowaste to enhance disease suppressiveness of compost-amended, peat-based potting mixes. Soil Biol. Biochem. 2005, 37, 2131–2140. [Google Scholar] [CrossRef]
- Tiquia, S.M.; Tam, N.F.Y. Elimination of phytotoxicity during co-composting of spent pig-manure sawdust litter and pig sludge. Bioresour. Technol. 1998, 65, 43–49. [Google Scholar] [CrossRef]
- Bartelt-Ryser, J.; Joshi, J.; Schmid, B.; Brandl, H.; Balser, T. Soil feedbacks of plant diversity on soil microbial communities and subsequent plant growth. Perspect. Plant Ecol. Evol. Syst. 2005, 7, 27–49. [Google Scholar] [CrossRef]
- Fulthorpe, R.R.; Allen, D.G. Evaluation of Biolog MT plates for aromatic and chloroaromatic substrate utilization tests. Can. J. Microbiol. 1994, 40, 1067–1071. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics; Academic Press Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Mitchell, A.L.; Scheremetjew, M.; Denise, H.; Potter, S.; Tarkowska, A.; Qureshi, M.; Salazar, G.A.; Pesseat, S.; Boland, M.A.; Hunter, F.M.I.; et al. EBI Metagenomics in 2017: Enriching the analysis of microbial communities, from sequence reads to assemblies. Nucleic Acids Res. 2018, 46, D726–D735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. 2019. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 30 December 2019).
- Nurk, S.; Meleshko, D.; Korobeynikov, A.; Pevzner, P.A. metaSPAdes: A new versatile metagenomic assembler. Genome Res. 2017, 27, 824–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, D.D.; Froula, J.; Egan, R.; Wang, Z. MetaBAT, an efficient tool for accurately reconstructing single genomes from complex microbial communities. PeerJ 2015, 3, e1165. [Google Scholar] [CrossRef] [Green Version]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- Bowers, R.M.; Kyrpides, N.C.; Stepanauskas, R.; Harmon-Smith, M.; Doud, D.; Reddy, T.B.K.; Schulz, F.; Jarett, J.; Rivers, A.R.; Eloe-Fadrosh, E.A.; et al. Minimum information about a single amplified genome (MISAG) and a metagenome-assembled genome (MIMAG) of bacteria and archaea. Nat. Biotechnol. 2017, 35, 725–731. [Google Scholar] [CrossRef] [Green Version]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Bonanomi, G.; Antignani, V.; Capodilupo, M.; Scala, F. Identifying the characteristics of organic soil amendments that suppress soilborne plant diseases. Soil Biol. Biochem. 2010, 42, 136–144. [Google Scholar] [CrossRef]
- Hadar, Y. Suppressive compost: When plant pathology met microbial ecology. Phytoparasitica 2011, 39, 311–314. [Google Scholar] [CrossRef] [Green Version]
- Blaya, J.; Marhuenda, F.C.; Pascual, J.A.; Ros, M. Microbiota Characterization of Compost Using Omics Approaches Opens New Perspectives for Phytophthora Root Rot Control. PLoS ONE 2016, 11, e0158048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, G.S.; Sivasithamparam, K. Antagonism of fungi and actinomycetes isolated from composted eucalyptus bark to Phytophthora drechsleri in a steamed and non-steamed composted eucalyptus bark-amended container medium. Soil Biol. Biochem. 1995, 27, 243–246. [Google Scholar] [CrossRef]
- López-González, J.A.; Suárez-Estrella, F.; Vargas-García, M.C.; López, M.J.; Jurado, M.M.; Moreno, J. Dynamics of bacterial microbiota during lignocellulosic waste composting: Studies upon its structure, functionality and biodiversity. Bioresour. Technol. 2015, 175, 406–416. [Google Scholar] [CrossRef]
- Takaku, H.; Kodaira, S.; Kimoto, A.; Nashimoto, M.; Takagi, M. Microbial communities in the garbage composting with rice hull as an amendment revealed by culture-dependent and -independent approaches. J. Biosci. Bioeng. 2006, 101, 42–50. [Google Scholar] [CrossRef]
- Eichorst, S.A.; Varanasi, P.; Stavila, V.; Zemla, M.; Auer, M.; Singh, S.; Simmons, B.A.; Singer, S.W. Community dynamics of cellulose-adapted thermophilic bacterial consortia. Environ. Microbiol. 2013, 15, 2573–2587. [Google Scholar] [CrossRef]
- Eichorst, S.A.; Joshua, C.; Sathitsuksanoh, N.; Singh, S.; Simmons, B.A.; Singer, S.W. Substrate-Specific Development of Thermophilic Bacterial Consortia by Using Chemically Pretreated Switchgrass. Appl. Environ. Microbiol. 2014, 80, 7423–7432. [Google Scholar] [CrossRef] [Green Version]
- Kolton, M.; Erlacher, A.; Berg, G.; Cytryn, E. The Flavobacterium Genus in the Plant Holobiont: Ecological, Physiological, and Applicative Insights. In Microbial Models: From Environmental to Industrial Sustainability; Castro-Sowinski, S., Ed.; Microorganisms for Sustainability; Springer: Singapore, 2016; pp. 189–207. ISBN 978-981-10-2555-6. [Google Scholar]
- Kamil, F.H.; Saeed, E.E.; El-Tarabily, K.A.; AbuQamar, S.F. Biological Control of Mango Dieback Disease Caused by Lasiodiplodia theobromae Using Streptomycete and Non-streptomycete Actinobacteria in the United Arab Emirates. Front. Microbiol. 2018, 9, 829. [Google Scholar] [CrossRef]
- Vurukonda, S.S.K.P.; Giovanardi, D.; Stefani, E. Plant Growth Promoting and Biocontrol Activity of Streptomyces spp. as Endophytes. Int. J. Mol. Sci. 2018, 19, 952. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, A.H.; Desoukey, S.Y.; Fouad, M.A.; Kamel, M.S.; Gulder, T.A.M.; Abdelmohsen, U.R. Natural Product Potential of the Genus Nocardiopsis. Mar. Drugs 2018, 16, 147. [Google Scholar] [CrossRef]
- Ganeshan, G.; Kumar, A.M. Pseudomonas fluorescens, a potential bacterial antagonist to control plant diseases. J. Plant Interact. 2005, 1, 123–134. [Google Scholar] [CrossRef]
- Jain, A.; Das, S. Insight into the Interaction between Plants and Associated Fluorescent Pseudomonas spp. Available online: https://www.hindawi.com/journals/ija/2016/4269010/ (accessed on 5 September 2019).
- Qin, S.; Xing, K.; Jiang, J.-H.; Xu, L.-H.; Li, W.-J. Biodiversity, bioactive natural products and biotechnological potential of plant-associated endophytic actinobacteria. Appl. Microbiol. Biotechnol. 2011, 89, 457–473. [Google Scholar] [CrossRef] [PubMed]
- Dochhil, H.; Dkhar, M.; Barman, D. Seed Germination Enhancing Activity of Endophytic Streptomyces Isolated from Indigenous Ethno-Medicinal. Int. J. Pharm. Biol. Sci. 2013, 4, 256–262. [Google Scholar]
- Manter, D.K.; Delgado, J.A.; Holm, D.G.; Stong, R.A. Pyrosequencing Reveals a Highly Diverse and Cultivar-Specific Bacterial Endophyte Community in Potato Roots. Microb. Ecol. 2010, 60, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Sang, M.K.; Kim, K.D. The volatile-producing Flavobacterium johnsoniae strain GSE09 shows biocontrol activity against Phytophthora capsici in pepper. J. Appl. Microbiol. 2012, 113, 383–398. [Google Scholar] [CrossRef] [PubMed]
- Tsavkelova, E.A.; Cherdyntseva, T.A.; Botina, S.G.; Netrusov, A.I. Bacteria associated with orchid roots and microbial production of auxin. Microbiol. Res. 2007, 162, 69–76. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Raw Materials | Species | L5/6A (%) | L2A (%) |
|---|---|---|---|
| Rocket salad | Eruca sativa | 58 | 8 |
| Endive | Cichorium endivia | 9 | 48 |
| Lettuce | Lactuca sativa | 2 | 25 |
| Fennel | Foeniculum vulgare | 14 | - |
| Mandarin orange | Citrus reticulata | 6 | - |
| Broccoli | Brassica oleracea | 2 | - |
| Pumpkin | Cucurbita pepo | - | 1 |
| Basil | Ocimum basilicum | - | 11 |
| Wood scraps | 9 | 7 | |
| Total | 100 | 100 |
| Compost | ||||
|---|---|---|---|---|
| L5/6A | L2A | |||
| pH | 6.95 | ±0.07 | 9.02 | ±0.02 |
| EC | 1950 | ±25 | 1514 | ±31 |
| Total nitrogen | 1.32 | ±0.07 | 1.68 | ±0.22 |
| Type | Substrates | Number |
|---|---|---|
| Amines/amides | Phenyletyl-amine | G4 |
| Amines/amides | Putrescine | H4 |
| Amino acids | L-Arginine | A4 |
| Amino acids | L-Asparagine | B4 |
| Amino acids | L-Phenylalanine | C4 |
| Amino acids | L-Serine | D4 |
| Amino acids | L-Threonine | E4 |
| Amino acids | Glycil-L-Glutamic Acid | F4 |
| Caboxylic acid | Pyruvic Acid Methyl Ester | B1 |
| Caboxylic acid | D-Galacturonic Acid | B3 |
| Caboxylic acid | γ-Hydroxybutyric Acid | E3 |
| Caboxylic acid | D-Glucosaminic Acid | F2 |
| Caboxylic acid | Itaconic Acid | F3 |
| Caboxylic acid | α-Ketobutyric Acid | G3 |
| Caboxylic acid | D-Malic Acid | H3 |
| Carbohydrates | β-Methyl-D Glucoside | A2 |
| Carbohydrates | D-Galactonic Acid γ-Lactone | A3 |
| Carbohydrates | D-Xylose | B2 |
| Carbohydrates | i-Erythritol | C2 |
| Carbohydrates | D-Mannitol | D2 |
| Carbohydrates | N-Acetil-D-glucosamine | E2 |
| Carbohydrates | D-Cellobiose | G1 |
| Carbohydrates | Glucose-1-Phosphate | G2 |
| Carbohydrates | α-D-Lactose | H1 |
| Carbohydrates | D,L-α-Glycerol Phosphate | H2 |
| Phenolic compounds | 2-Hidroxy Benzoic Acid | C3 |
| Phenolic compounds | 4-Hydroxy Benzoic Acid | D3 |
| Polymers | Tween 40 | C1 |
| Polymers | Tween 80 | D1 |
| Polymers | α-Cyclodextrine | E1 |
| Polymers | Glycogen | F1 |
| Sample | Amplicon | Raw Reads | High Quality Reads | Total Number of Reads Assigned to the Reference Database | Assigned OTUs | |
|---|---|---|---|---|---|---|
| L5/6A | 16S | 2,578,978 | 1,938,089 | 1,934,201 | (99.80%) | 2884 |
| L2A | 16S | 1,868,462 | 1,400,736 | 1,397,531 | (99.77%) | 2880 |
| L5/6A | 18S | 1,891,223 | 1,757,231 | 1,748,980 | (99.53%) | 1375 |
| L2A | 18S | 1,866,587 | 1,692,091 | 1,677,857 | (99.16%) | 1696 |
| L5/6A | ||||
|---|---|---|---|---|
| MAG Id | CMPL | CNTM | #UM | Taxonomy |
| MAG.1 | 74.45 | 2.2 | 29 | k__Bacteria |
| MAG.10 | 62.28 | 1.3 | 33 | k__Bacteria;p__Bacteroidetes;c__Flavobacteriia;o__Flavobacteriales;f__Flavobacteriaceae |
| MAG.11 | 98.62 | 1.13 | 43 | k__Bacteria;p__Bacteroidetes;c__Flavobacteriia;o__Flavobacteriales;f__Flavobacteriaceae |
| MAG.13 | 93.23 | 7.54 | 43 | k__Bacteria;p__Proteobacteria;c__Gammaproteobacteria;o__Xanthomonadales;f__Xanthomonadaceae;g__Luteimonas |
| MAG.14 | 83.26 | 5.79 | 42 | k__Bacteria;p__Deinococcus-Thermus;c__Deinococci;o__Deinococcales;f__Deinococcaceae |
| MAG.15 | 60.55 | 9.12 | 11 | k__Bacteria;p__Proteobacteria;c__Alphaproteobacteria;o__Rhodobacterales;f__Rhodobacteraceae;g__Paracoccus |
| MAG.17 | 85.16 | 7.57 | 41 | k__Bacteria;p__Bacteroidetes;c__Sphingobacteriia;o__Sphingobacteriales;f__Sphingobacteriaceae;g__Sphingobacterium |
| MAG.18 | 99.14 | 1.19 | 43 | k__Bacteria;p__Proteobacteria;c__Alphaproteobacteria;o__Sphingomonadales |
| MAG.2 | 57.23 | 5.17 | 35 | k__Bacteria;p__Bacteroidetes;c__Cytophagia;o__Cytophagales;f__Cyclobacteriaceae |
| MAG.4 | 66.22 | 6.43 | 17 | k__Bacteria;p__Bacteroidetes;c__Sphingobacteriia;o__Sphingobacteriales;f__Sphingobacteriaceae;g__Sphingobacterium |
| MAG.6 | 53.38 | 4.7 | 24 | k__Bacteria;p__Actinobacteria;c__Actinobacteria;o__Actinomycetales;f__Promicromonosporaceae |
| MAG.8 | 89.47 | 6.36 | 42 | k__Bacteria;p__Bacteroidetes;c__Flavobacteriia;o__Flavobacteriales;f__Flavobacteriaceae |
| L2A | ||||
| MAG Id | CMPL | CNTM | #UM | Taxonomy |
| MAG.10 | 74.89 | 2.62 | 35 | k__Bacteria;p__Bacteroidetes;c__Flavobacteriia;o__Flavobacteriales;f__Flavobacteriaceae |
| MAG.11 | 97.88 | 1.19 | 43 | k__Bacteria;p__Proteobacteria;c__Alphaproteobacteria;o__Sphingomonadales |
| MAG.12 | 98.81 | 4.88 | 40 | k__Bacteria;p__Bacteroidetes;c__Sphingobacteriia;o__Sphingobacteriales;f__Sphingobacteriaceae;g__Sphingobacterium |
| MAG.14 | 85.16 | 3.58 | 39 | k__Bacteria |
| MAG.18 | 66.51 | 2.28 | 36 | k__Bacteria;p__Deinococcus-Thermus;c__Deinococci;o__Deinococcales;f__Deinococcaceae |
| MAG.2 | 87.8 | 3.62 | 27 | k__Bacteria;p__Bacteroidetes;c__Flavobacteriia;o__Flavobacteriales;f__Flavobacteriaceae |
| MAG.4 | 80.52 | 4.45 | 39 | k__Bacteria;p__Proteobacteria;c__Gammaproteobacteria;o__Xanthomonadales;f__Xanthomonadaceae;g__Luteimonas |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scotti, R.; Mitchell, A.L.; Pane, C.; Finn, R.D.; Zaccardelli, M. Microbiota Characterization of Agricultural Green Waste-Based Suppressive Composts Using Omics and Classic Approaches. Agriculture 2020, 10, 61. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10030061
Scotti R, Mitchell AL, Pane C, Finn RD, Zaccardelli M. Microbiota Characterization of Agricultural Green Waste-Based Suppressive Composts Using Omics and Classic Approaches. Agriculture. 2020; 10(3):61. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10030061
Chicago/Turabian StyleScotti, Riccardo, Alex L. Mitchell, Catello Pane, Rob D. Finn, and Massimo Zaccardelli. 2020. "Microbiota Characterization of Agricultural Green Waste-Based Suppressive Composts Using Omics and Classic Approaches" Agriculture 10, no. 3: 61. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10030061