Secondary Metabolites Produced by Macrophomina phaseolina Isolated from Eucalyptus globulus

 , , ,
, , ,  , , , , and
, , , , and 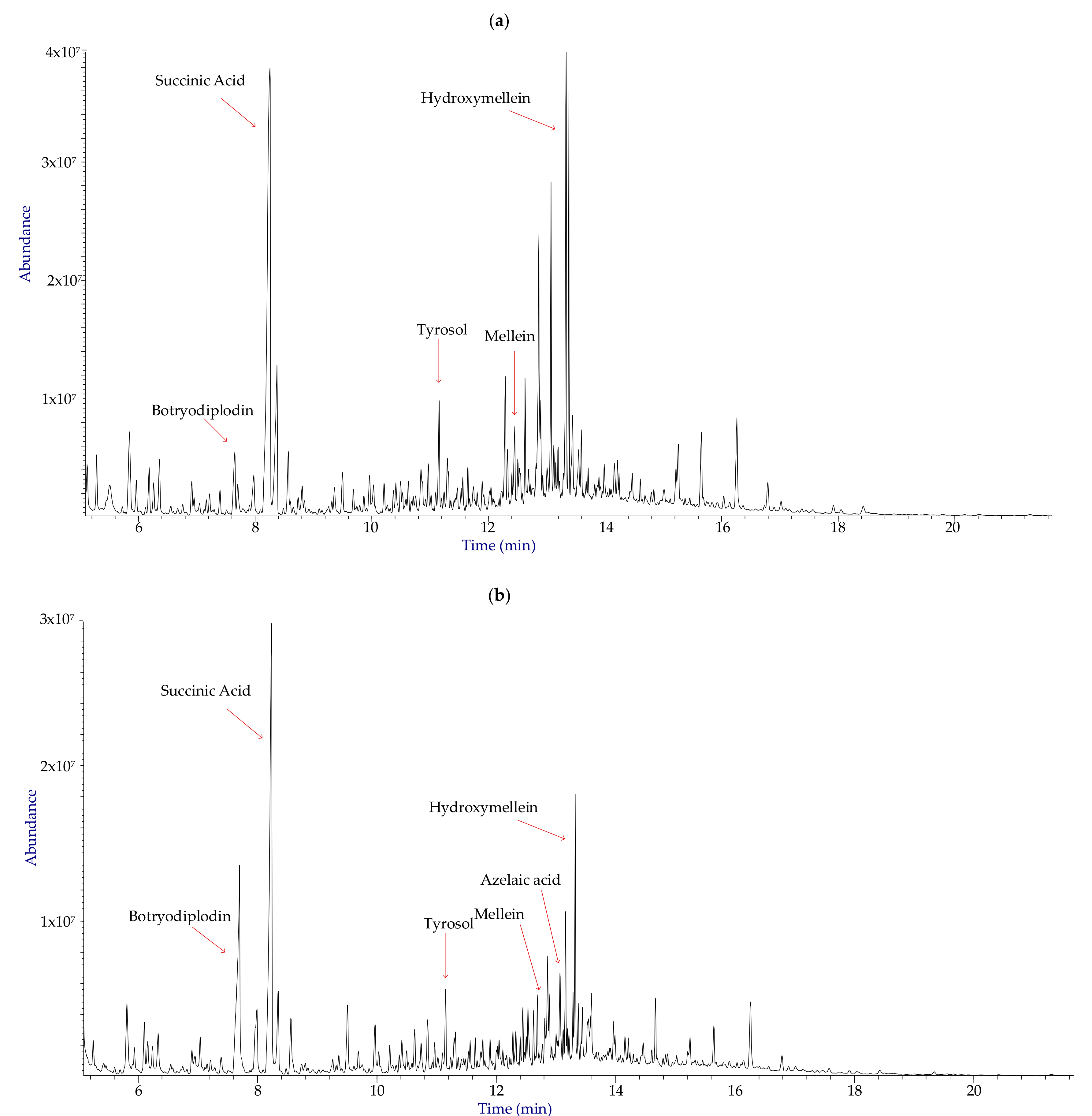{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Experimental Procedures
2.2. Fungal Isolate and Growth Conditions
2.3. Extraction and Purification of Fungal Metabolites
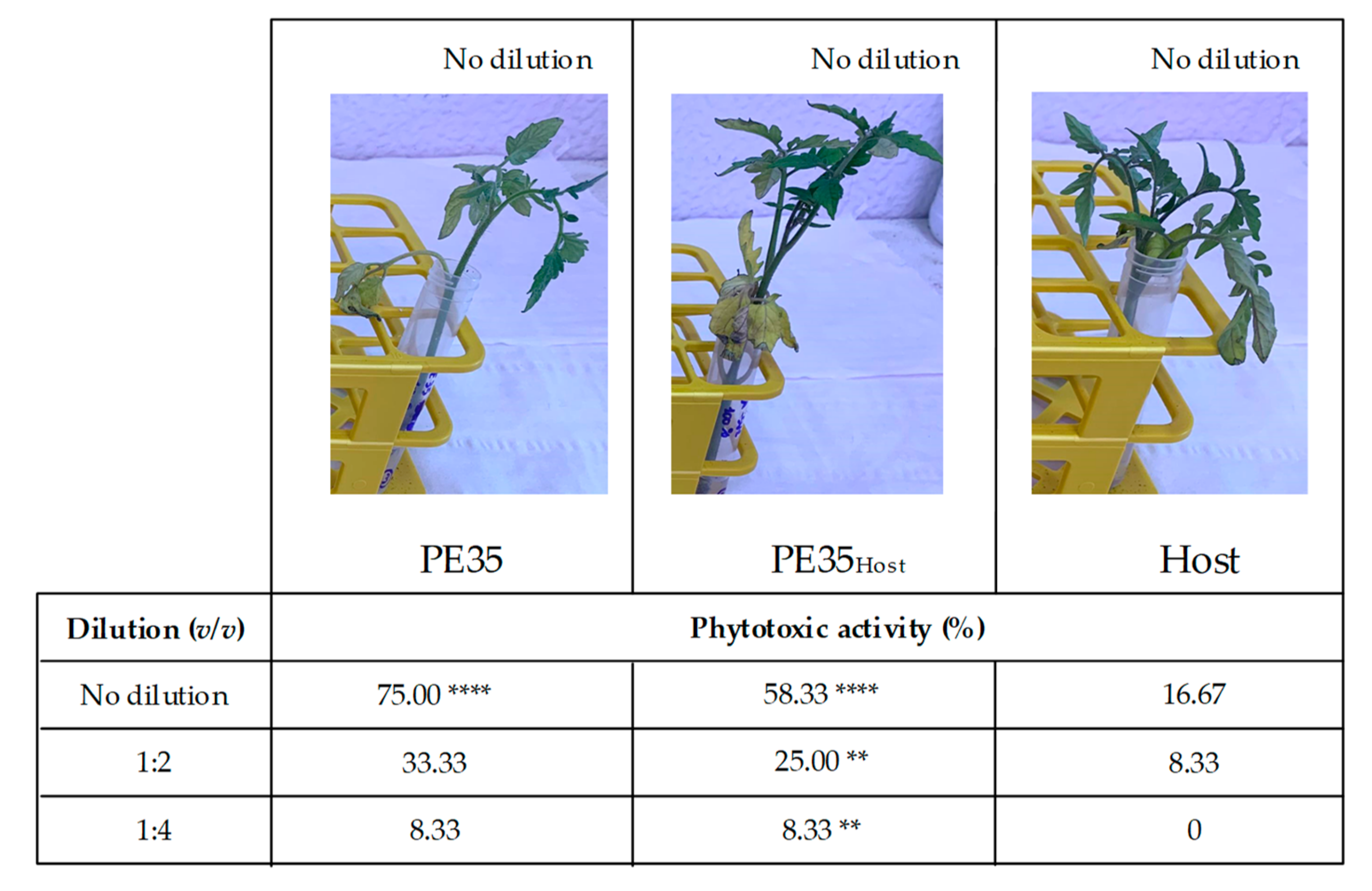
2.4. Phytotoxicity Assay
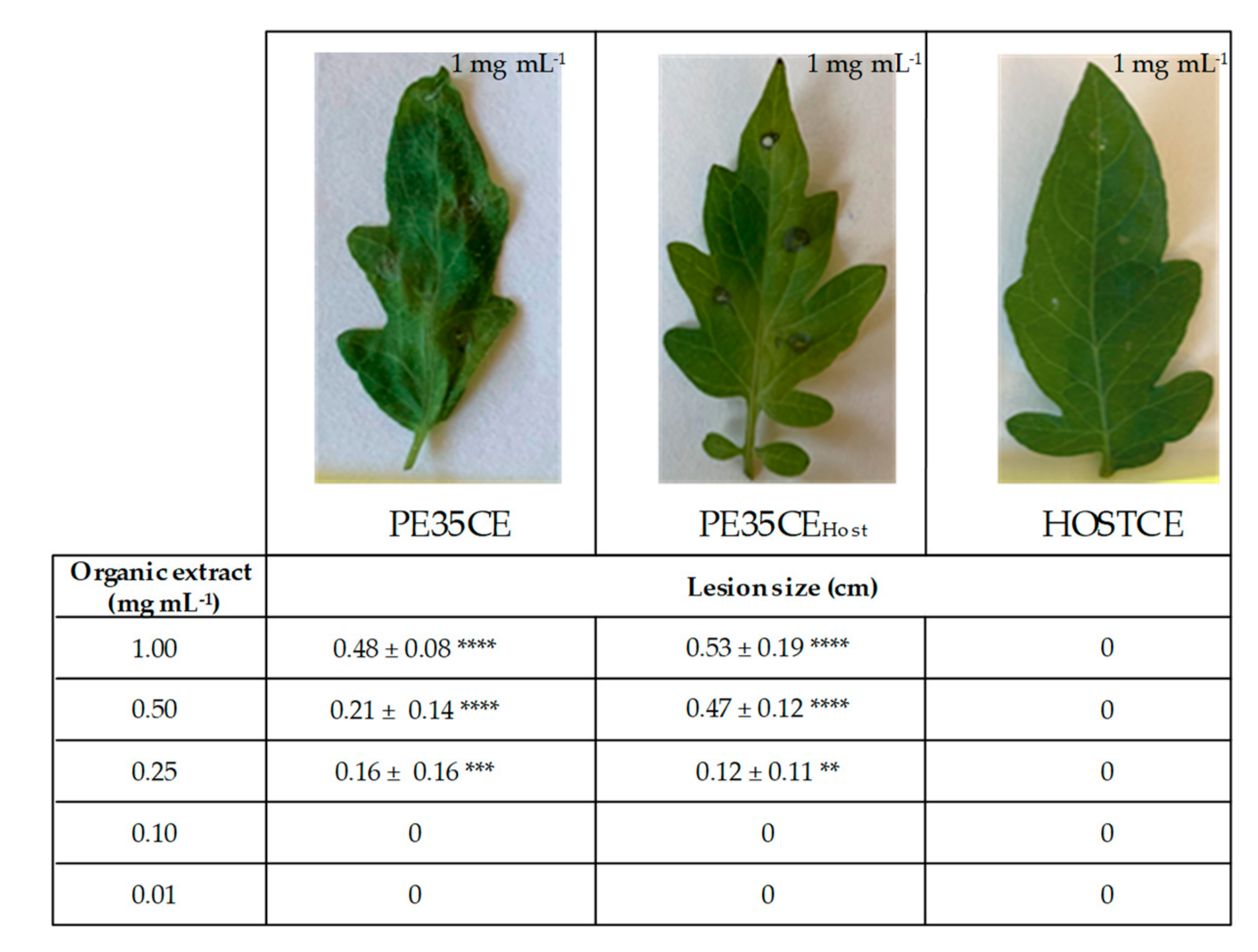
2.5. Cytotoxicity Assay
2.6. Statistical Analysis
3. Results
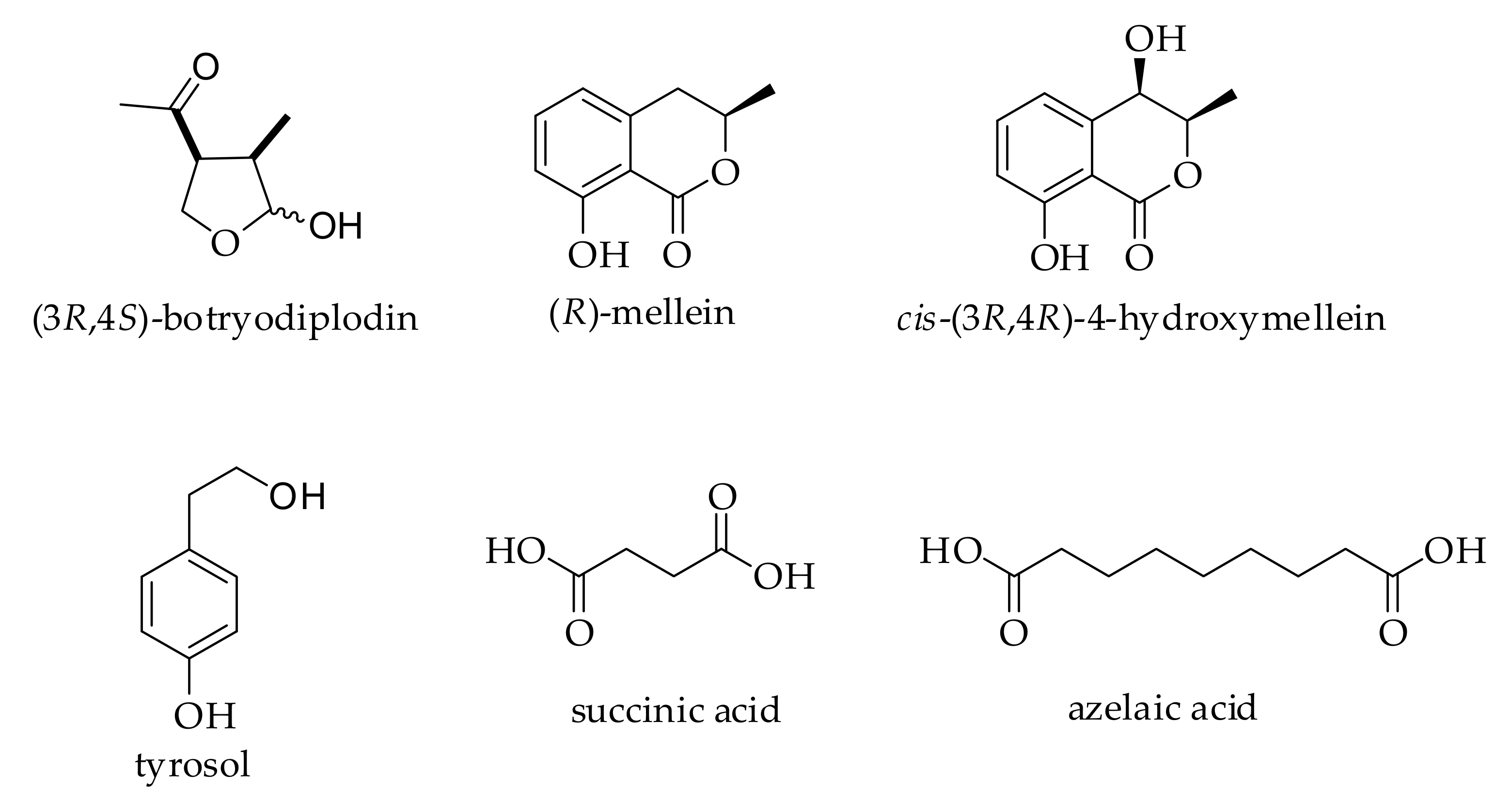
3.1. Identification and Distribution of Metabolites
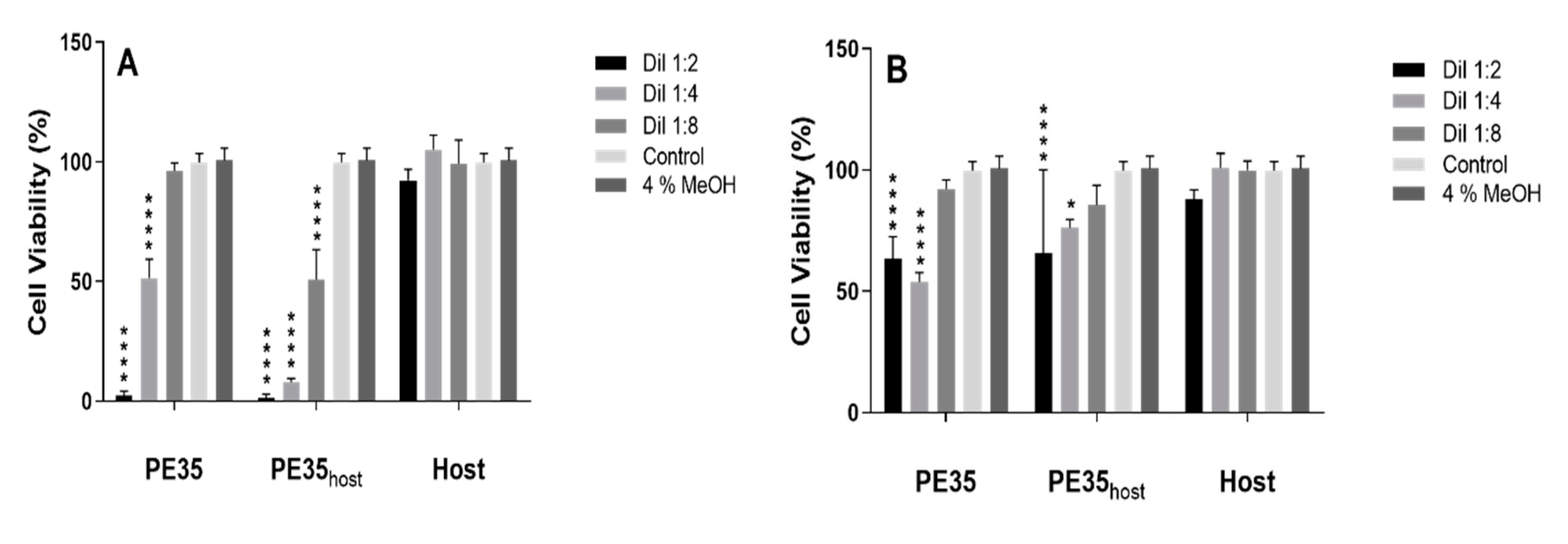
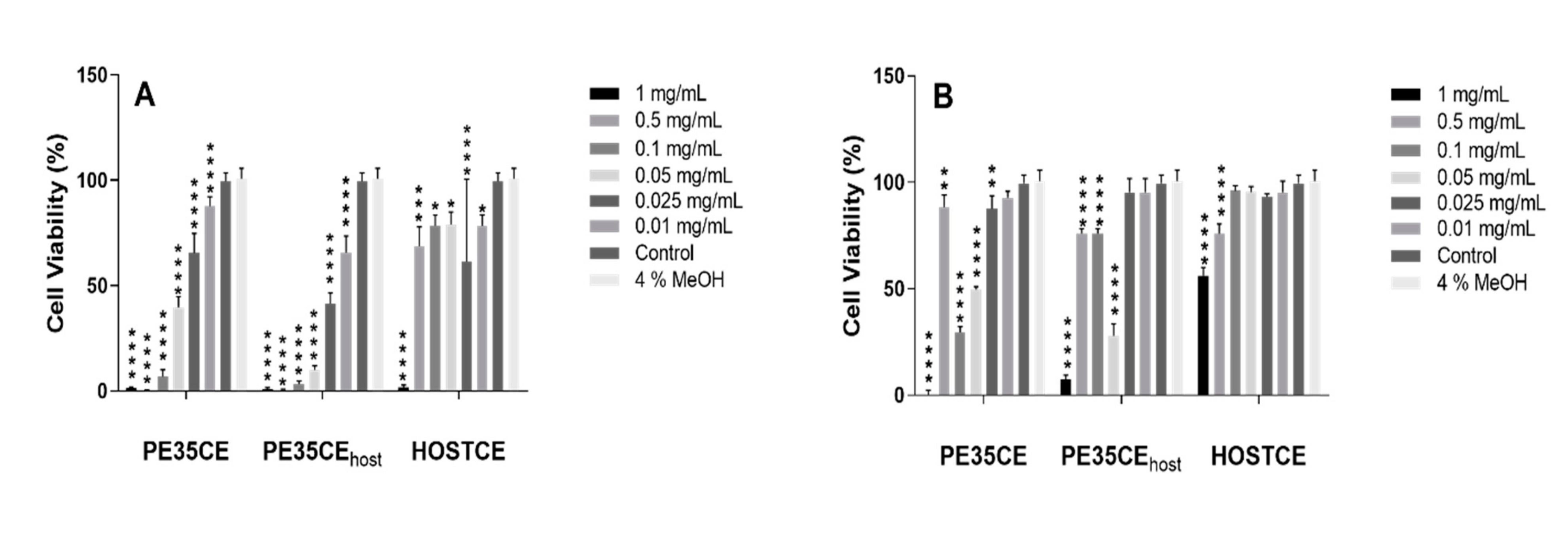
3.2. Phytotoxicity and Cytotoxicity Assays
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- House, S.M. Reproductive biology of eucalypts. In Eucalypt Ecology: Individuals to Ecosystems; Cambridge University Press: Cambridge, UK, 1997; pp. 30–55. ISBN 0-521-49740. [Google Scholar]
- Coppen, J.J. Eucalyptus: The Genus Eucalyptus; CRC Press: Boca Raton, FL, USA, 2002; ISBN 0-203-21943-0. [Google Scholar]
- Zhang, J.; An, M.; Wu, H.; Liu, D.L.; Stanton, R. Chemical composition of essential oils of four Eucalyptus species and their phytotoxicity on silverleaf nightshade (Solanum elaeagnifolium Cav.) in Australia. Plant Growth Regul. 2012, 68, 231–237. [Google Scholar] [CrossRef]
- Dhakad, A.K.; Pandey, V.V.; Beg, S.; Rawat, J.M.; Singh, A. Biological, medicinal and toxicological significance of Eucalyptus leaf essential oil: A review. J. Sci. Food Agric. 2018, 98, 833–848. [Google Scholar] [CrossRef] [PubMed]
- Myburg, A.A.; Potts, B.M.; Marques, C.M.; Kirst, M.; Gion, J.M.; Grattapaglia, D.; Grima-Pettenatti, J. Eucalypts. In Forest Trees; Springer: Berlin/Heidelberg, Germany, 2007; pp. 115–160. ISBN 978-3-540-34540-4. [Google Scholar]
- Wingfield, M.J.; Slippers, B.; Hurley, B.P.; Coutinho, T.A.; Wingfield, B.D.; Roux, J. Eucalypt pests and diseases: Growing threats to plantation productivity. South For. 2008, 70, 139–144. [Google Scholar] [CrossRef]
- Burgess, T.I.; Wingfield, M.J. Pathogens on the move: A 100-year global experiment with planted eucalypts. Bioscience 2017, 67, 14–25. [Google Scholar] [CrossRef]
- Barradas, C.; Phillips, A.J.; Correia, A.; Diogo, E.; Bragança, H.; Alves, A. Diversity and potential impact of Botryosphaeriaceae species associated with Eucalyptus globulus plantations in Portugal. Eur. J. Plant Pathol. 2016, 146, 245–257. [Google Scholar] [CrossRef]
- Slippers, B.; Fourie, G.; Crous, P.W.; Coutinho, T.A.; Wingfield, B.D.; Carnegie, A.J.; Wingfield, M.J. Speciation and distribution of Botryosphaeria spp. on native and introduced Eucalyptus trees in Australia and South Africa. Stud. Mycol. 2004, 50, 343–358. [Google Scholar]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [Green Version]
- Dissanayake, A.J.; Phillips, A.J.L.; Li, X.H.; Hyde, K.D. Botryosphaeriaceae: Current status of genera and species. Mycosphere 2016, 7, 1001–1073. [Google Scholar] [CrossRef]
- Gezahgne, A.; Roux, J.; Slippers, B.; Wingfield, M.J.; Hare, P.D. Identification of the causal agent of Botryosphaeria stem canker in Ethiopian Eucalyptus plantations. S. Afr. J. Bot. 2004, 70, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Slippers, B.; Burgess, T.; Pavlic, D.; Ahumada, R.; Maleme, H.; Mohali, S.; Rodas, C.; Wingfield, M.J. A diverse assemblage of Botryosphaeriaceae infect Eucalyptus in native and non-native environments. South For. 2009, 71, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Soni, K.K.; Dadwal, V.S. Charcoal root and stem rot of Eucalyptus. Eur. J. For. Pathol. 1985, 15, 397–401. [Google Scholar] [CrossRef]
- Agarwal, D.K. Macrophomina: Taxonomy, Ecology, Diseases and their Management. In Taxonomy and Ecology of Indian Fungi; IK International Publishing House: New Dehli, India, 2010; pp. 59–78. ISBN 978-93-80026-92-3. [Google Scholar]
- De, B.K.; Chattopadhya, S.B.; Arjunan, G. Effect of potash on stem rot diseases of jute caused by Macrophomina phaseolina. J. Mycopathol. Res. 1992, 30, 51–55. [Google Scholar]
- Su, G.; Suh, S.O.; Schneider, R.W.; Russin, J.S. Host specialization in the charcoal rot fungus, Macrophomina phaseolina. Phytopathology 2001, 91, 120–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.N. Macrophomina phaseolina as causal agent for charcoal rot of sunflower. Mycopathology 2007, 5, 111–118. [Google Scholar]
- Jacob, C.J.; Krarup, C.; Díaz, G.A.; Latorre, B.A. A severe outbreak of charcoal rot in cantaloupe melon caused by Macrophomina phaseolina in Chile. Plant Dis. 2013, 97, 141–142. [Google Scholar] [CrossRef]
- Sánchez, S.; Henríquez, J.L.; Urcola, L.A.; Scott, A.; Gambardella, M. Susceptibility of strawberry cultivars to root and crown rot caused by Macrophomina phaseolina. J. Berry Res. 2016, 6, 345–354. [Google Scholar] [CrossRef]
- Kaur, S.; Dhillon, G.S.; Brar, S.K.; Vallad, G.E.; Chand, R.; Chauhan, V.B. Emerging phytopathogen Macrophomina phaseolina: Biology, economic importance and current diagnostic trends. Crit. Rev. Microbiol. 2012, 38, 136–151. [Google Scholar] [CrossRef]
- Dhar, T.K.; Siddiqui, K.A.; Ali, E. Structure of phaseolinone, a novel phytotoxin from Macrophomina phaseolina. Tetrahedron Lett. 1982, 23, 5459–5462. [Google Scholar]
- Mahato, S.B.; Siddiqui, K.A.; Bhattacharya, G.; Ghosal, T.; Miyahara, K.; Sholichin, M.; Kawasaki, T. Structure and stereochemistry of phaseolinic acid: A new acid from Macrophomina phaseolina. J. Nat. Prod. 1987, 50, 245–247. [Google Scholar] [CrossRef]
- Guida, M.; Salvatore, M.M.; Salvatore, F. A strategy for GC/MS quantification of polar compounds via their silylated surrogates: Silylation and quantification of biological amino acids. J. Anal. Bioanal. Tech. 2015, 6, 263–279. [Google Scholar]
- Félix, C.; Duarte, A.S.; Vitorino, R.; Guerreiro, A.C.; Domingues, P.; Correia, A.C.; Alves, A.; Esteves, A.C. Temperature modulates the secretome of the phytopathogenic fungus Lasiodiplodia theobromae. Front. Plant Sci. 2016, 7, 1096. [Google Scholar]
- Duarte, A.S.; Cavaleiro, E.; Pereira, C.; Merino, S.; Esteves, A.C.; Duarte, E.P.; Tomás, J.M.; Correia, A.C. Aeromonas piscicola AH-3 expresses an extracellular collagenase with cytotoxic properties. Lett. Appl. Microbiol. 2015, 60, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Ammerman, N.C.; Beier-Sexton, M.; Azad, A.F. Growth and maintenance of vero cell lines. Curr. Protoc. Microbiol. 2008, 11, A.4E.1–A.4E.7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvatore, M.M.; Nicoletti, R.; Salvatore, F.; Naviglio, D.; Andolfi, A. GC–MS approaches for the screening of metabolites produced by marine-derived Aspergillus. Mar. Chem. 2018, 206, 19–33. [Google Scholar] [CrossRef]
- NIST Standard Reference Data. Available online: http://www.nist.gov/srd/nist1a.cfm (accessed on 3 February 2020).
- Félix, C.; Salvatore, M.M.; DellaGreca, M.; Meneses, R.; Duarte, A.S.; Salvatore, F.; Naviglio, D.; Gallo, M.; Jorrino-Novo, J.V.; Alves, A.; et al. Production of toxic metabolites by two strains of Lasiodiplodia theobromae, isolated from a coconut tree and a human patient. Mycologia 2018, 110, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Félix, C.; Salvatore, M.M.; DellaGreca, M.; Ferreira, V.; Duarte, A.S.; Salvatore, F.; Naviglio, D.; Gallo, M.; Alves, A.; Esteves, A.C.; et al. Secondary metabolites produced by grapevine strains of Lasiodiplodia theobromae grown at two different temperatures. Mycologia 2019, 111, 466–476. [Google Scholar] [CrossRef]
- Schwartz, R.A.; Kapila, R. Macrophomina phaseolina: An overlooked cutaneous infection, seed rot disease in humans. Int. J. Dermatol. 2019. [CrossRef]
- Tan, D.H.; Sigler, L.; Gibas, C.F.; Fong, I.W. Disseminated fungal infection in a renal transplant recipient involving Macrophomina phaseolina and Scytalidium dimidiatum: Case report and review of taxonomic changes among medically important members of the Botryosphaeriaceae. Med. Mycol. 2008, 46, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, A.; Wickes, B.L.; Romanelli, A.M.; Debelenko, L.; Rubnitz, J.E.; Sutton, D.A.; Thompson, E.H.; Fothergill, A.W.; Rinaldi, M.G.; Hayden, R.T.; et al. Cutaneous infection caused by Macrophomina phaseolina in a child with acute myeloid leukemia. J. Clin. Microbiol. 2009, 47, 1969–1972. [Google Scholar] [CrossRef] [Green Version]
- Arora, P.; Dilbaghi, N.; Chaudhury, A. Opportunistic invasive fungal pathogen Macrophomina phaseolina prognosis from immunocompromised humans to potential mitogenic RBL with an exceptional and novel antitumor and cytotoxic effect. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 101–107. [Google Scholar] [CrossRef]
- Premamalini, T.; Ambujavalli, B.T.; Vijayakumar, R.; Rajyogahandh, S.V.; Kalpana, S.; Kindo, A.J. Fungal keratitis caused by Macrophomina phaseolina a case report. Med. Mycol. Case Rep. 2012, 1, 123–126. [Google Scholar] [CrossRef]
- Romero Luna, M.P.; Mueller, D.; Mengistu, A.; Singh, A.K.; Hartman, G.L.; Wise, K.A. Advancing our understanding of charcoal rot in soybeans. J. Integr. Pest. Manag. 2017, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Ramezani, M.; Shier, W.T.; Abbas, H.K.; Tonos, J.L.; Baird, R.E.; Sciumbato, G.L. Soybean charcoal rot disease fungus Macrophomina phaseolina in Mississippi produces the phytotoxin (−)-botryodiplodin but no detectable phaseolinone. J. Nat. Prod. 2007, 70, 128–129. [Google Scholar] [CrossRef] [PubMed]
- Abbas, H.K.; Bellaloui, N.; Accinelli, C.; Smith, J.R.; Shier, W.T. Toxin production in soybean (Glycine max L.) plants with charcoal rot disease and by Macrophomina phaseolina, the fungus that causes the disease. Toxins 2009, 11, 645. [Google Scholar] [CrossRef] [Green Version]
- Salvatore, M.M.; Giambra, S.; Naviglio, D.; DellaGreca, M.; Salvatore, F.; Burruano, S.; Andolfi, A. Fatty acids produced by Neofusicoccum vitifusiforme and N. parvum, fungi associated with grapevine Botryosphaeria dieback. Agriculture 2018, 8, 189. [Google Scholar] [CrossRef] [Green Version]
- Dongdong, Z.; Jin, Y.; Yang, T.; Yang, Q.; Wu, B.; Chen, Y.; Luo, Z.; Liang, L.; Liu, Y.; Xu, A.; et al. Antiproliferative and immunoregulatory effects of azelaic acid against acute myeloid leukemia via activation of notch signaling pathway. Front. Pharmacol. 2019, 10, 1396. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Suero, M.; Benard-Gellon, M.; Chong, J.; Laloue, H.; Stempien, E.; Abou-Mansour, E.; Fontaine, F.; Larignon, P.; Mazet-Kieffer, F.; Farine, S.; et al. Extracellular compounds produced by fungi associated with Botryosphaeria dieback induce differential defence gene expression patterns and necrosis in Vitis vinifera cv. Chardonnay cells. Protoplasma 2014, 251, 1417–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andolfi, A.; Maddau, L.; Cimmino, A.; Linaldeddu, B.T.; Franceschini, A.; Serra, S.; Basso, S.; Melck, D.; Evidente, A. Cyclobotryoxide, a phytotoxic metabolite produced by the plurivorous pathogen Neofusicoccum australe. J. Nat. Prod. 2012, 75, 1785–1791. [Google Scholar] [CrossRef] [PubMed]
- Andolfi, A.; Maddau, L.; Cimmino, A.; Linaldeddu, B.T.; Basso, S.; Deidda, A.; Serra, S.; Evidente, A. Lasiojasmonates A–C, three jasmonic acid esters produced by Lasiodiplodia sp., a grapevine pathogen. Phytochemistry 2014, 103, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Andolfi, A.; Mugnai, L.; Luque, J.; Surico, G.; Cimmino, A.; Evidente, A. Phytotoxins produced by fungi associated with grapevine trunk diseases. Toxins 2011, 3, 1569–1605. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salvatore, M.M.; Félix, C.; Lima, F.; Ferreira, V.; Naviglio, D.; Salvatore, F.; Duarte, A.S.; Alves, A.; Andolfi, A.; Esteves, A.C. Secondary Metabolites Produced by Macrophomina phaseolina Isolated from Eucalyptus globulus. Agriculture 2020, 10, 72. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10030072
Salvatore MM, Félix C, Lima F, Ferreira V, Naviglio D, Salvatore F, Duarte AS, Alves A, Andolfi A, Esteves AC. Secondary Metabolites Produced by Macrophomina phaseolina Isolated from Eucalyptus globulus. Agriculture. 2020; 10(3):72. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10030072
Chicago/Turabian StyleSalvatore, Maria Michela, Carina Félix, Fernanda Lima, Vanessa Ferreira, Daniele Naviglio, Francesco Salvatore, Ana Sofia Duarte, Artur Alves, Anna Andolfi, and Ana Cristina Esteves. 2020. "Secondary Metabolites Produced by Macrophomina phaseolina Isolated from Eucalyptus globulus" Agriculture 10, no. 3: 72. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10030072