Light Quality Affects Growth and Physiology of Carpesium triste Maxim. Cultured In Vitro



1
Department of Horticulture, Division of Applied Life Science (BK21 Plus Program), Graduate School of Gyeongsang National University, Jinju 52828, Korea
2
Institute of Agriculture and Life Science, Gyeongsang National University, Jinju 52828, Korea
3
Research Institute of Life Science, Gyeongsang National University, Jinju 52828, Korea
*
Author to whom correspondence should be addressed.
Agriculture 2020, 10(7), 258; https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10070258
Submission received: 1 June 2020
/
Revised: 26 June 2020
/
Accepted: 26 June 2020
/
Published: 2 July 2020
(This article belongs to the Section Agricultural Technology)
Abstract
:The objective of this study was to investigate the effect of light quality on the growth and contents of photosynthetic pigments and total flavonoids, the parameters of chlorophyll a fluorescence, antioxidant capacities, and enzyme activities. It was intended to preliminarily explore the internal mechanisms involved for the effect of light quality on the growth and physiology of Carpesium triste Maxim. Stem apex explants were cultured on the Murashige and Skoog (MS) medium under white (W), red (R), blue (B), or a 1:1 mixture of red and blue (RB) light-emitting diodes (LEDs) for four weeks. The RB induced sturdy plantlets and increased the contents of photosynthetic pigments and total flavonoids, photosynthetic electron transport and efficiency, 2-diphenyl-1-picrylhydrazyl (DPPH) free radical scavenging capacity, and activities of antioxidant enzymes in the plantlets. Taken together, combinations of monochromatic red and blue LEDs could be used for the best production of high-quality C. triste plantlets in vitro.
1. Introduction
Carpesium triste Maxim. is a rare perennial herbal species of the Carpesium genus that belongs to the Asteraceae family, which is widely distributed in South Korea, China, and Japan. It has a slender stem and is usually 40–100 cm in height. Its root, stem, leaf, and fruit have long been used as a traditional medicine in the treatment of fevers, mastitis, toothache, diarrhea, and even cancer [1] due to the anti-inflammatory, antifebrile, cytotoxic, and styptic properties of the biologically active compounds [2]. The usage and plant parts extracted for use as herbs in the Carpesium genus vary, according to the oral and literature records in different countries. For example, the shoots of Carpesium abrotanoides L. were used to treat fever and to inhibit inflammation in Korea [3]. The dried seeds of Carpesium abrotanoides L., also called Carpesii Fructus, were prescribed to treat diseases caused by parasites, such as ascariasis, cestodiasis, and enterobiasis in China [4,5]. Previous studies on the Carpesium genus have shown that the effective pharmacological ingredients mainly include flavonoids, sesquiterpene lactones (germacranolide and guaianolide type) [6,7,8], acyclic diterpenoids [9,10], thymol derivatives [11], and terpenoids [12,13]. In the Carpesium genus, previous studies were mainly focused on chemical compounds, officinal substances, and clinical applications. There has been no study that investigates plant regeneration and growth regulation of the Carpesium genus in in vitro conditions until now.
Light is a vital environmental factor that regulates the growth, development, morphogenesis, photosynthesis, and substance metabolism in plants [14,15]. Plants respond to the fluctuating light environment mainly via perceiving, capturing, and transducing functions of a series of photoreceptors. The artificial light environment became one of the core technologies for tissue culture after De Capite explored the effects of light intensity on the growth of Helianthus annuus, Parthenocissus tricuspidata, and Daucus carota [16]. Light is pivotal for the growth and development of plants as the energy source for photochemical reactions in photosynthesis and as the chief trigger signal that regulates diverse biological effects on plants including photomorphogenesis [17,18], photosynthesis [19,20], physiological–biochemical characteristics [19,21], and development of tissues or organs [22,23]. A broad range of the light spectrum from 260 nm to 780 nm modulates plant growth and development throughout their lifecycle. For example, the decreased red/far-red ratio elongated the stem and reorientation of leaves [24] and enhanced the tolerance of subzero temperature in Arabidopsis [25]. The supplemental UV-A and B LED or R LED to baby leaf lettuce increased the anthocyanins or phenolics concentrations, respectively [26]. The white and red lights were proven to be effective in promoting seed germination in Arabidopsis, and blue light promoted dormancy in barley and wheat [27]. Lately, light-emitting diodes (LEDs) are widely used in horticultural and photobiological applications in controlled environments for their unique performances. The LED has a high photoelectric transition efficiency, affordable price, portable size, long service life, low heat generation, and adjustable spectrum and intensity [28,29], making it an attractive light source. Previous studies have shown that red and blue LEDs have the greatest impact on plant growth due to the function of supplying energy in CO2 assimilation. The combination of red and blue LEDs, especially at a 1:1 ratio, could increase the accumulation of carbohydrates [30], could reduce levels of both leaf chlorosis and epinasty [31], and could promote growth and development of plants [32] in various species and, hence, often has been used in closed-type plant production systems.
Chlorophyll a fluorescence induction has been deeply studied since Kantsky and Hirsch discovered the negative correlation between fluorescence intensity and carbon dioxide fixation in 1931 [33]. Light energy absorbed by plants is utilized through three close correlated ways: absorption by photosynthetic pigments, heat loss caused by internal conversion, and fluorescence emission [34]. Measurement of chlorophyll a fluorescence could indirectly reflect the photosynthesis and heat dissipation. The parameters of chlorophyll a fluorescence have been used as indicators of the photochemical activity, photosynthetic electron transport, and photosynthetic apparatus status [35]. Previous studies have shown that parameters of chlorophyll a fluorescence in leaves of plants cultured in a controlled environment were affected by the light quality [36,37] and quantity [38,39], nutritional status [40,41], or environmental stresses [42,43].
Stress caused by the changeable light environment may cause the production of reactive oxygen species (ROS). The ROSs are crucial regulators of metabolism and plant stress responses, involving regulation of cell wall development and activation of antimicrobial defenses as signal molecules [44]. However, as toxic byproducts of aerobic metabolism, ROSs also tend to damage plants’ cells as well as photosynthetic apparatuses including DNA, RNA, protein, and membrane oxidation and damage [45]. The ROS could be mainly derived from the photosynthetic electron transport chain in photosystem I (PS I), photosystem II (PS II), and photorespiration [46], which can influence the stability of the chlorophyll and can decrease the photosynthetic activity. Plants have established mechanisms formed through long-term evolution to alleviate this damage. The heat dissipation [47] and the ROS scavengers by antioxidant enzymes, e.g., superoxide dismutase (SOD), catalase (CAT), peroxidase (POD), and ascorbate peroxidase (APX) [48], and by soluble antioxidants, e.g., flavonoids, anthocyanins, and various types of secondary metabolites, are main approaches of photoprotection [49].
This research focused on the effects of light quality provided by LEDs on growth and physiology of plantlets of C. triste and studied the responses of plantlets to light qualities. Growth parameters of shoot and root, contents of photosynthetic pigments and total flavonoids, parameters of chlorophyll a fluorescence, 2-diphenyl-1-picrylhydrazyl (DPPH) free radical scavenging capacity, and activities of antioxidant enzymes were investigated and analyzed. The ultimate aim of this research was to provide an effective approach and the basis of production of high-quality plantlets for a large scale and fast reproduction as a mean to conserve this wild germplasm resource.
2. Materials and Methods
2.1. Plant Materials and Sterilization Methods
Dried seeds of C. triste were obtained from the Korea National Arboretum (KNA) in Pocheon, Gyeonggi-do, South Korea. Healthy and uniform seeds were selected for surface sterilization. Dust and impurities were cleaned up with running tap water for 5 min, and then, the seeds were dipped in 70% ethanol for 5 min for preliminary disinfection. After being rinsed 5–6 times in distilled water, the seeds were immersed in a 3% (v/v) NaClO solution for 15 min. Afterward, the seeds were washed adequately with distilled water 5 times to remove the residual disinfectants and then dried on sterile tissue papers. After surface sterilization, seeds were cultured on the plant growth regulator (PGR)–free Murashige and Skoog (MS) medium supplemented with 3% (w/v) sucrose and 0.8% (w/v) agar.
2.2. Light Treatments and Growth Parameters
Stem apex explants from the 2-month-old plantlets derived from seed germination were cultured on the PGR–free MS medium and incubated in the plant growth chamber for four weeks with day/night temperatures of 24/18 °C and 16 hours of daily photoperiod under one of the following four light environments: white (W, 40 W tubes, Philips, The Netherlands), red (R, 621–710 nm), blue (B, 450–475 nm), or a 1:1 mixture of red and blue (RB) LEDs (LED, 400–700 nm, PSLED–1203–50A, Force Lighting Co.; Ltd., Hwaseong, Korea) at an intensity of 50 µmol·m−2·s−1 photosynthetic photon flux density (PPFD) measured with a photo/radiometer (HD 2102.2, Delta OHM, Padova, Italy). The spectral distributions were measured with a portable spectroradiometer (Spectra Light ILT 950, International Light Technologies, Inc., Peabody, MA, USA). The spectral quality of each light treatment is shown in Figure 1. Each treatment was repeated in three sets of 9 plantlets. Shoot and root lengths, number of leaves and roots, shoot diameter, and fresh and dry weights of shoots and roots were recorded after four weeks of exposure to the different lights. The dry weights of the shoots and roots were determined after drying for 48 hours at 60 °C.
2.3. Determination of Contents of Photosynthetic Pigments
A modified method described by Baglieri [50] was used to determine chlorophyll content: 500 mg of plant leaves was soaked in 10 mL 96% (v/v) ethanol for 10 hours at 4 °C for thorough decoloring, and after filtrating the leaf residues, extracts were applied to determine the contents of chlorophyll a and b and total carotenoids with a spectrophotometer at 665, 649, and 470 nm wavelengths, respectively. The calculations were based on formulae stated by Lichtenthaler and Wellburn [51]. The content of each photosynthetic pigment was measured three times, and the mean value was calculated.
2.4. Chlorophyll a Fluorescence Transient Measurement
Parameters of chlorophyll a fluorescence were measured with FluorPen (FP110/D, Photon Systems Instruments, Brno, Czech Republic). Intact and fully expanded leaves marked by the black arrows in Figure 2 were adapted to the dark for 20 min using detachable leaf-clips before measurement. The measuring light was set at 0.09 μmol·m−2·s−1, and the saturating light pulse was set at 3000 µmol·m−2·s−1 The FM/F0, FV/FM, and FV/F0 were calculated from the main parameters of chlorophyll a fluorescence. The calculated parameters from chlorophyll a fluorescence induction curve (OJIP), absorption (ABS), trapping (TR0), heat dissipation (DI0), and electron transport (ET0) per reaction center (RC) are related to specific energy fluxes. Parameters were measured in three replicates of 9 leaves. The formulas of OJIP protocol parameters were described by Strasser et al., as shown in Table 1 [52,53].
2.5. Determination of Activities of CAT, APX, POD, and SOD
One hundred milligrams of fresh leaf tissue were ground in liquid nitrogen and homogenized with a 1.5 mL 50 mM phosphate buffer (pH 7.0) containing 1 mM ethylenediaminetetraacetic acid (EDTA), 0.05% Triton X-100, and 2% polyvinylpyrrolidone (PVP). After being centrifuged at 13,000 rpm for 20 min at 4 °C, the supernatant of the homogenate was collected and used to measure the enzyme activities.
2.5.1. Catalase (CAT; EC 1.11.1.6)
The CAT activity was determined as described by Aebi [54]. The 3.0 mL assay system consisted of a 50 mM phosphate buffer (pH 7.0), 10 mM H2O2, and 0.1 mL of the extracted enzyme. The decrease in absorbance at 240 nm was monitored for 3 min. One unit of CAT activity represents 1 μmol H2O2 decomposed per minute. The specific CAT activity was calculated as the units of enzyme activity per mg protein of samples (U·mg−1·protein). The extinction coefficient of H2O2 at 240 nm was 40 mM−1·cm−1.
2.5.2. Ascorbate Peroxidase (APX; EC 1.11.1.11)
The APX activity was determined as described by Nakano and Asada [55]. The 3.0 mL assay system consisted of a 50 mM phosphate buffer (pH 7.0), 0.5 mM ascorbic acid, 0.1 mM H2O2, and 0.1 mL of the extracted enzyme. The decrease in the absorbance at 290 nm was monitored for 3 min. One unit of APX activity represents oxidation of 1 μmol of ascorbate per minute. The specific APX activity was calculated as the units of enzyme activity per mg protein of samples (U·mg−1·protein). The extinction coefficient was 28 mM−1·cm−1.
2.5.3. Peroxidase (POD; EC 1.11.1.7)
The POD activity was determined as described by Putter [56] with slight modifications. The 3.0 mL assay system consisted of a 50 mM phosphate buffer (pH 7.0), 10 mM H2O2, 20 mM guaiacol solution, and 0.1 mL extracted enzyme. The increase in the absorbance at 470 nm was recorded for 3 min. One unit of POD activity represents the formation of 1 μmol of tetraguaiacol per minute. The specific POD activity was calculated as the units of enzyme activity per mg protein of samples (U·mg−1·protein).
2.5.4. Superoxidase Dismutase (SOD; EC 1.15.1.1)
The SOD activity was determined as described by Beauchamp and Fridovich [57]. The 3.0 mL assay system consisted of a 50 mM phosphate buffer (pH 7.0), 0.1 mM EDTA, 2 mM riboflavin, and 0.1 mL of the extracted enzyme, which was incubated under light for 10 min. The reaction mixture contained a 50 mM phosphate buffer (pH 7.0) kept in dark and 2 mM riboflavin kept in light, which served as the reference and the blank, respectively. The absorbance at 560 nm was recorded, and one unit of SOD was defined as the amount of the enzyme that inhibited the nitroblue tetrazolium (NBT) reduction by 50%. The specific SOD activity was calculated as the units of enzyme activity per mg protein of samples (U·mg−1·protein).
2.6. Preparation of Plant Extracts
The extraction of 4-week-old plantlets as affected by light quality was performed according to the described protocol of Manivannan et al. [58]. Five hundred milligrams of finely powdered leaf samples were lyophilized with 5.0 mL 80% methanol (v/v) and extracted overnight. Then, the mixture was sonicated in an ultra-sonication bath (Bandelin Sonorex Super RK156BH, Berlin, Germany) for 30 min and centrifuged at 12,000 rpm for 10 min. The supernatant was employed for the assessment or stored at 4 °C.
2.7. Total Flavonoids Estimation
The total flavonoid quantification was determined based on the aluminum chloride colorimetric method as described by Chang et al. [59]. The 2 mL reaction mixture included 0.1 mL plant extracts, 1 mL 2% aluminum chloride solution, and 0.9 mL 80% methanol. The absorbance of the reaction mixture was incubated for 30 min and measured at 415 nm by using UV/Vis-spectrophotometer (LibraS22, Biochrom Ltd., Cambridge, UK). The total flavonoids were calculated according to a calibration curve of standard quercetin and expressed on the fresh weight basis as mg quercetin equivalent (QE)·mg−1.
2.8. 2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Free Radical Scavenging Capacity
The DPPH free radical scavenging capacity was determined as follows. A 20 μL extract of each sample or ascorbic acid standard solutions were added to 1.980 mL of a newly prepared DPPH solution and incubated under dark for 20 min. The absorbance of the reaction mixture was measured at 517 nm using a UV/V is spectrophotometer [60]. A standard curve was obtained by measuring the DPPH scavenging capacities of 50, 100, 150, and 200 mg·L−1 ascorbic acid. The free radical scavenging % was recorded by the equation (1 − As/Ac) × 100%, where As is the OD value of the sample extracts or ascorbic acid solution and Ac is the OD value of the DPPH solution. The DPPH scavenging capacity was expressed on a fresh weight basis as mg ascorbic acid equivalent (AAE) g−1.
2.9. Data Collection and Statistical Analysis
The assays were performed in triplicate to verify the reproducibility of the results. The data were collected and reported as the mean ± the standard error. A one-way analysis of variance (one-way ANOVA) was performed to evaluate the significant differences among the treatments, followed by a Tukey’s multiple range test (p < 0.05). Counting data were assessed via Kruskal–Wallis analysis of variance with the Chi-square test (p < 0.05). Statistical analysis was conducted using the SAS statistical software package (SAS Institute Inc., Cary, NC, USA). All figures were made in OriginPro software version 9.0.
3. Results
3.1. Effect of Light Quality on Growth and Development of Plantlets
As shown in Table 2, all growth indexes of C. triste plantlets showed extremely significant or significant differences under W, R, B, and RB LEDs. Under R, plantlets had the greatest shoot length (p < 0.05), which suggested that R played a positive role in promoting stem elongation. According to the Kruskal–Wallis Rank Sum Test, there was significant difference among the light qualities on the number of leaves (p = 0.023) and roots (p = 0.019). The R was the least effective in inducing new leaves, whereas W and RB were the best in increasing the leaf number (p < 0.05). Plantlets grown under R and RB had greater root lengths and number of roots than those grown under W and B. The shoot diameter of plantlets grown under RB was the greatest, followed in order by that of plantlets grown under W, R, and B (p < 0.05). The RB also increased the fresh and dry weights of shoots (p < 0.05). The above results indicated that plantlets displayed distinctly varying responses to the different light qualities. Plantlets grown under RB had a greater shoot diameter, dry matter accumulation, and root system development, which suggested that RB promotes the production of stout plantlets (Figure 2).
3.2. Effect of Light Quality on Contents of Photosynthetic Pigments
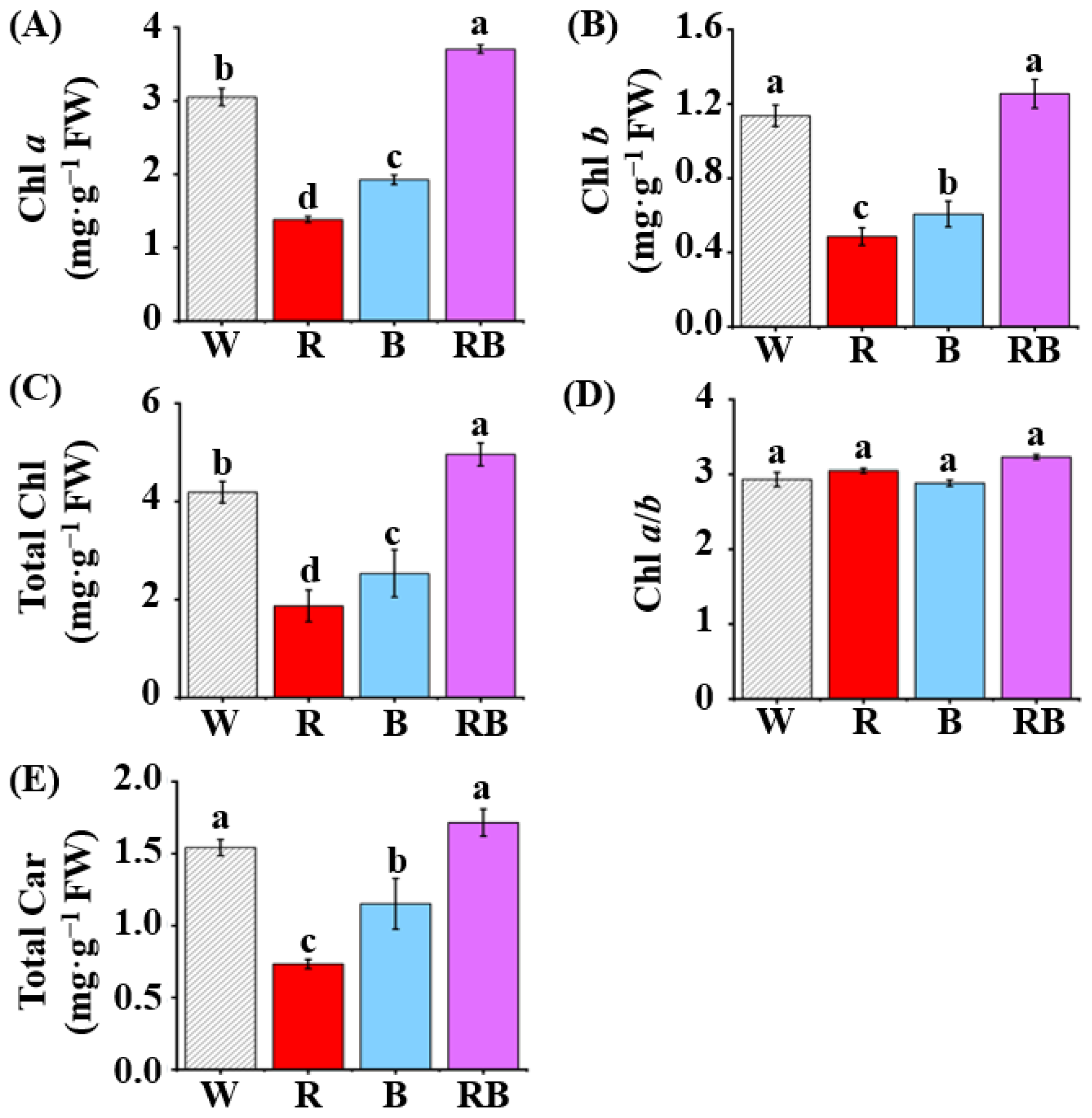
Chlorophyll a content in leaves of C. triste plantlets grown under RB was higher than that of plantlets grown under W, B, and R (p < 0.05, Figure 3A). The W or RB were the most effective promoters of chlorophyll b accumulation (p < 0.05, Figure 3B). Plantlets grown under RB had the highest total chlorophyll levels, followed by those grown under W, B, and R (p < 0.05, Figure 3C). The chlorophyll a/b ratio did not show much difference between treatments (p < 0.05, Figure 3D). In addition, W and RB were effective in increasing the content of total carotenoids in leaves of C. triste plantlets, whereas R and B were less effective in doing so (p < 0.05, Figure 3E). Above all, RB led to increased total chlorophyll and carotenoid accumulation, which indicated that light quality may influence the photosynthetic performance through the photosynthetic pigments.
3.3. Effect of Light Quality on Parameters of Chlorophyll a Fluorescence in Leaves
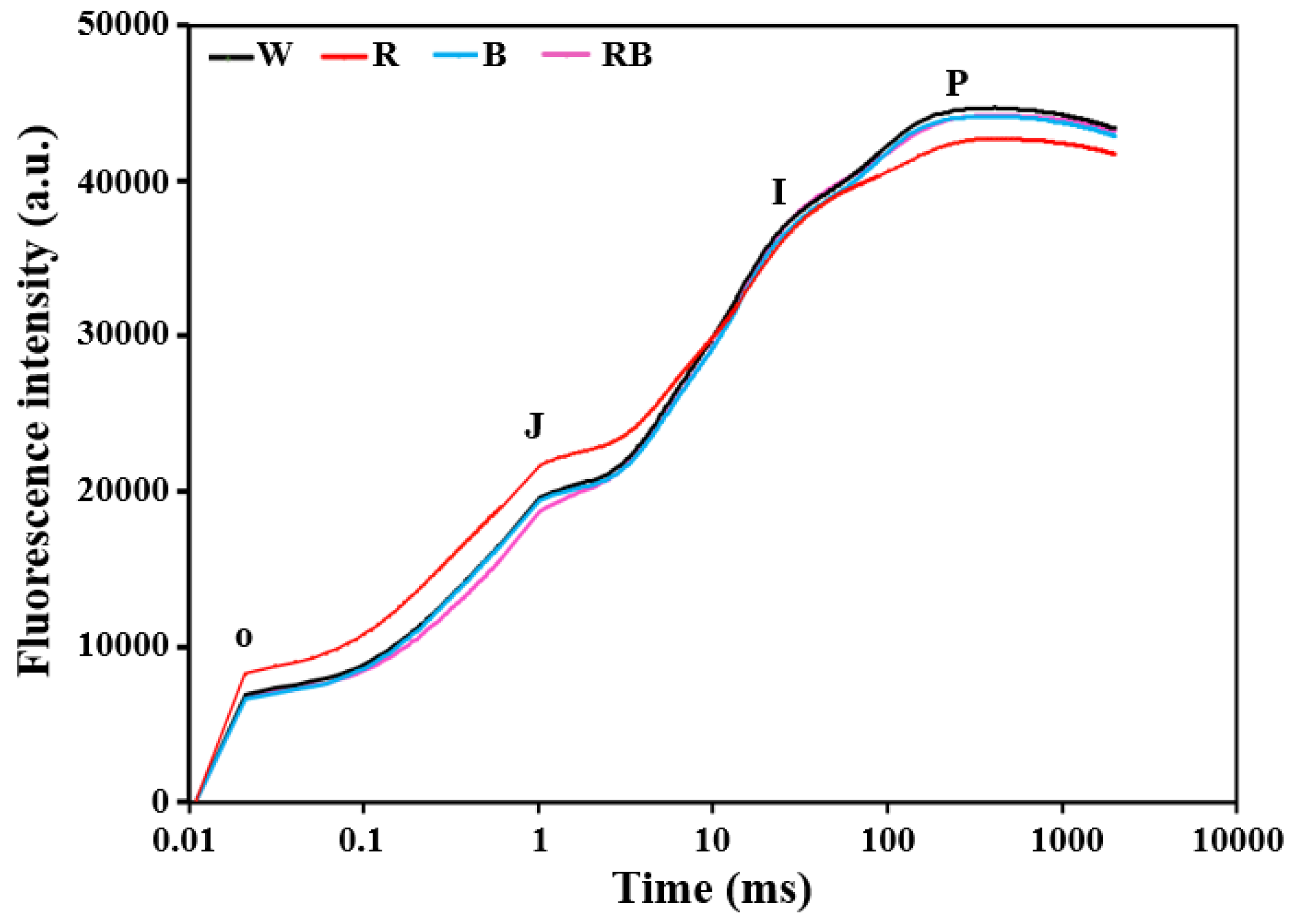
The OJIP method is considered an indicator of the state of the photosynthetic electron transport processes [61]. The OJ, JI, and IP phases represent the reduction of the acceptor side of PS II, of the plastoquinone (PQ)–pool, and of the acceptor side of PS I, respectively [40,41,42]. In this study, each step in OJIP curves exhibited a different response to light qualities (Figure 4). The fluorescence intensities were higher in the OJ phase under R. However, it showed the weakest signal in IP phase under R. RB induced lower fluorescence intensity in the OJ phase. For W and B, there was no significant difference in OJIP curves (Figure 4).
For parameters of chlorophyll a fluorescence, light quality influenced the minimum fluorescence (F0) but not the maximum fluorescence (FM) in C. triste leaves (p < 0.05, Table 3). It was observed that the F0 was appreciably higher in leaves of plantlets grown under R than that in plantlets grown under RB (p < 0.05, Table 3). The lowest electron transfer activity (FM/F0) was also observed in plantlets grown under R (p < 0.05, Table 3). The maximum quantum efficiency of PS II photochemistry (FV/FM) and the actual quantum yield of PS II (FV/F0) were significantly reduced with R (p < 0.05), which were decreases of 3.5% and 16.5% respectively, compared to the maximum value (Table 3).
The M0 reflects the approximate initial slope of the fluorescence transient, and SM is the normalized total complementary area. There was a great increase in the M0 value of plantlets grown under R (p < 0.05). The values of SM decreased in plantlets grown under W and R. In leaves of plantlets grown under RB, the lowest M0 and highest SM values were observed (p < 0.05, Table 3).
In C. triste leaves, the values of the quantum yield of electron transport (φE0) and the possibility of a trapped exciton moving an electron into the electron transport chain beyond QA (ψ0) were 10.9% and 7.5% lower, respectively, when grown under R than their corresponding highest values when grown under RB (p < 0.05, Table 3). In contrast, the quantum yield of heat dissipation (φD0) under R was approximately 18.8% higher than the lowest value (Table 3). Under R, specific energy fluxes per reaction center related to the light absorption flux (ABS/RC), trapped energy flux (TR0/RC), electron transport flux (ET0/RC), and dissipated energy flux (DI0/RC) were 24.5%, 27.1%, 10.8%, and 50.0% higher, respectively, than their lowest values under RB (Table 3). The performance index on an absorption basis (PIABS) in C. triste leaves was highest under RB (p < 0.05), followed in order by that under W and B, and R, where the minimum value was 51.7% of the maximum (Table 3).
3.4. Effect of Light Quality on Content of Total Flavonoids
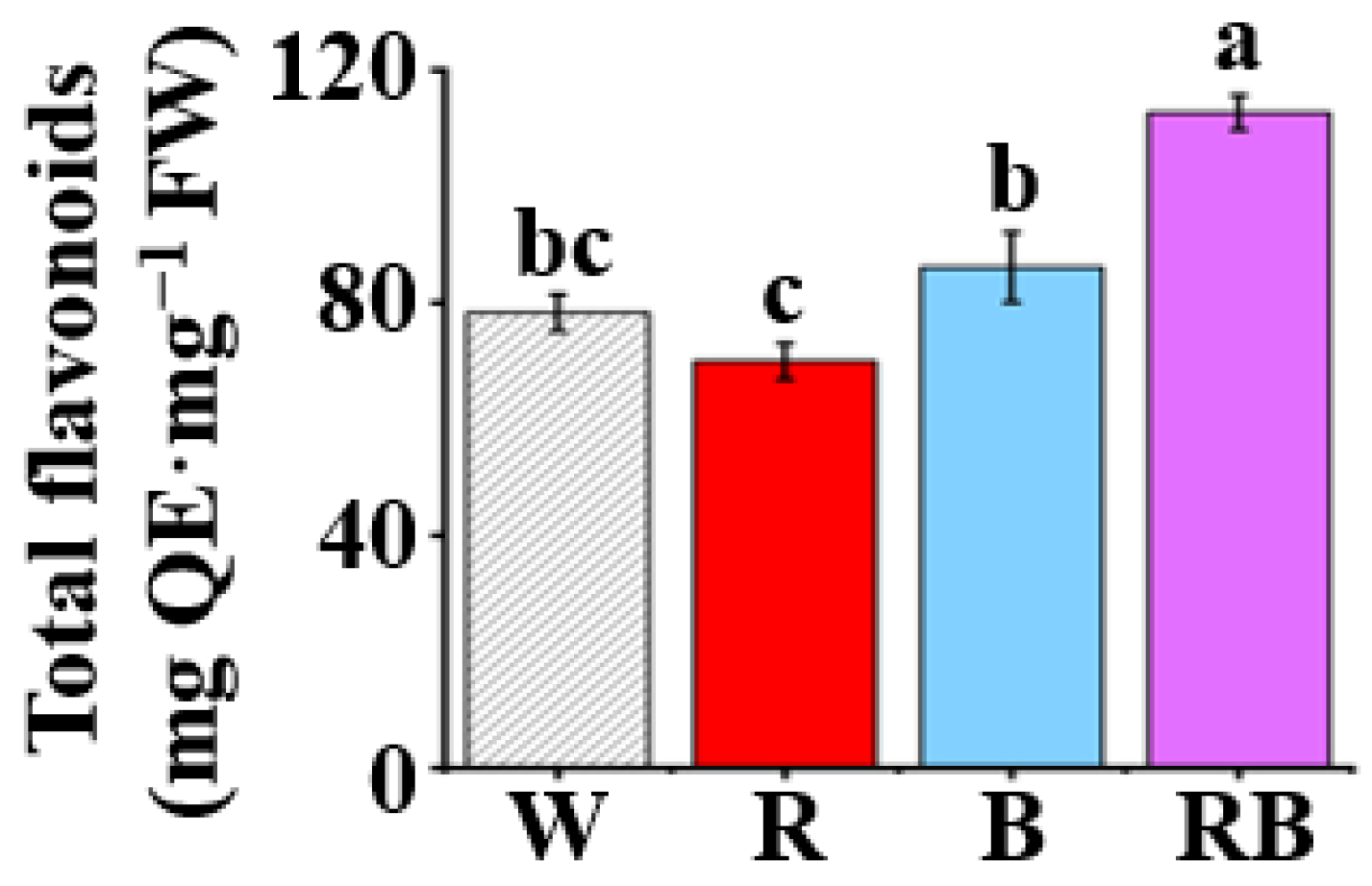
Content of total flavonoids represented as quercetin equivalent (QE) in the leaves of C. triste plantlets differed significantly (p < 0.05) among treatments of four light qualities after four weeks of treatment (Figure 5). Content of total flavonoids in leaves of plantlets under RB displayed the highest value as compared to other treatments, followed in order by that of plantlets grown under B, W, and R (p < 0.05). Compared to R treatment, content of total flavonoids in the leaves of RB, B, and W treatments increased by 61.0%, 23.2%, and 11.9%, respectively.
3.5. Effect of Light Quality on DPPH Radical Scavenging Capacity
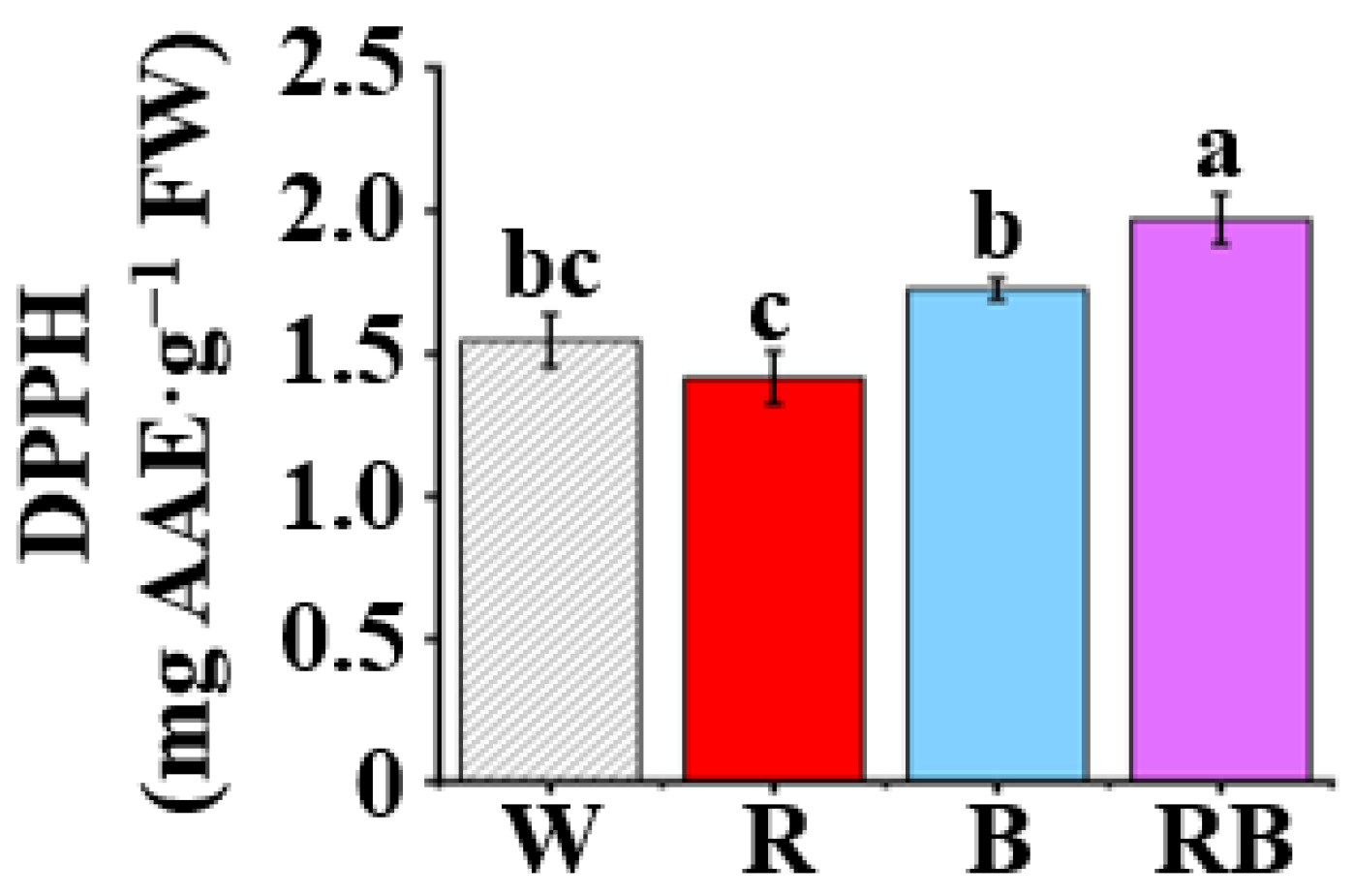
The DPPH is an organic radical with a deep purple color which will become yellow when it accepts an electron from an antioxidant. Therefore, the antioxidant capacity could be evaluated by the spectrophotometric measurements of the discoloration degree. The antioxidant capacities in C. triste plantlets grown under different light qualities, as measured by DPPH radical scavenging assays represented by ascorbic acid equivalent (AAE), were determined (Figure 6). The result showed that the highest antioxidant capacities were noted in the extracts of RB-treated plantlets (1.97 mg AAE·g−1 FW, p < 0.05) and that the extracts of B-treated plantlets exhibited moderate radical inhibition, followed by that of plantlets treated by W and R.
3.6. Effect of Light Quality on Activities of Antioxidant Enzymes
The activities of antioxidant enzymes in leaves of plantlets grown under different light qualities were determined in this study. The RB greatly increased the activities of CAT and APX compared to the other light qualities (p < 0.05), the respective values of which were 2.1- and 3.7-fold larger than the lowest values observed in leaves of plantlets grown under R (Figure 7A,B). The activities of POD and SOD were the lowest in leaves of plantlets grown under R, and the highest values were observed in those grown under W and RB (p < 0.05). For POD activity, the ratio of the highest to lowest was approximate 2.8-fold, and for SOD activity, the ratio of the highest to lowest was approximate 1.7-fold, respectively (p < 0.05, Figure 7C,D).
4. Discussion
The effects of light quality on the growth and development of C. triste plantlets were explored as light quality plays a vital role in regulating photosynthetic capacity, which may benefit rapid and high-quality propagation in vitro. This study showed that R led to longer shoots and roots in C. triste plantlets than other light qualities (p < 0.05, Table 2). These data were consistent with the results in various species such as Euphorbia millii [62], Tripterospermum japonicum [63], and Oncidium [64]. Plantlets grown under B had weak growth vigor, exhibiting the shortest shoots and roots, lowest fresh and dry weights, and slenderest stems (p < 0.05, Table 2). It was also reported that the increased internode length was due to the deficiency of blue light in Phaseolus vulgaris [65] and that blue light inhibited stem elongation in Pelargonium [66]. However, the highest specific growth rate and extracellular polysaccharide production of Porphyridium cruentum were observed under blue light [67]. Red light increased shoot height and rooting frequency in Convolvulus arvensis [68]. Many shreds of evidence are indicating that the combination of red and blue LEDs benefits the growth and development of plantlets cultured in vitro [69,70,71]. The combination of red and blue light enhanced plant above- and underground growth and development of Cymbidium by increasing the net leaf photosynthesis rate as well as the stomatal conductance in in vitro conditions [72,73]. These observations indicated that inductions and developments of shoot and root are probably dependent on the spectral quality of light. The reason underlying this phenomenon might be that the spectral energy distribution of red and blue lights was accordant with that of chlorophyll absorption [74]. The increased indole-3-acetic acid (IAA) oxidase activity caused by blue light [75,76] or promoted accumulation of gibberellin (GA) caused by red light [77] may be involved in the mechanisms with which light regulates shoot elongation. Reducing the red/far-red ratio promoted stem elongation and gave significant increases in contents of GA and IAA in both internodes and leaves in Helianthus annuus [78]. The decreased abscisic acid (ABA) content was observed with R light treatment [79]. In this study, the combination of red and blue LEDs in a 1:1 ratio increased the number of leaves and roots, fresh and dry weights of shoots, as well as shoot diameter (p < 0.05, Table 2), which led to a more compact and sturdy morphology of plantlets. The underlying mechanisms of signal interactions of light quality and plant hormones are well worth exploring in the future.
Photosynthetic pigments are another important factor for photosynthesis as well as plant growth. Light quality influences photosynthesis through the light absorption ability of the photosynthetic pigments [80]. Chlorophyll a mainly functions in photosystems, and chlorophyll b is concentrated in light-harvesting complexes (LHC) [81]. In this study, the highest chlorophyll a and total chlorophyll were observed in the leaves of C. triste plantlets grown under RB (p < 0.05, Figure 3A,C). W and RB promoted chlorophyll b and total carotenoid accumulation in leaves of plantlets (p < 0.05, Figure 3B,E). The chlorophyll a/b ratio indicates the amount of chlorophyll associated with the PS I and PS II complexes [82] and has a positive correlation with the net photosynthetic rate [83]. Blue light plays an important role in chlorophyll formation and chloroplast development [84]. Contents of chlorophyll and total carotenoids were higher under B than under R in this study, and it may because blue light was more effective than red light in the induction of photosynthetic pigments synthesis in leaves [85]. The change in the Chl a/b ratio often indicates a change in the ratio between the reaction center complexes of the photosystem and the LHC. In this study, there was no significant difference in the Chl a/b ratio between treatments (Figure 3D), and it indicated that the LHC of the thylakoid membranes was not altered much by light quality within the treatment time. Carotenoids are important components of the reaction center complex and act as accessory pigments in the antenna systems to perform functions of harvesting the light energy and improving the photosynthetic efficiency [86]. W and RB led to the highest content of total carotenoids in leaves of plantlets (p < 0.05), which indicated an enhanced photosynthetic performance of plants.
Chlorophyll a fluorescence acts as a subtle indicator of the photochemical activity and electron transfer of photosynthesis and is related to the physiological status of plants [87]. Based on the electron transport in the receptor side and donor side in PS II and the energy transfer in thylakoid membranes, Strasser and others built up the OJIP method on analysis and processing of rapid chlorophyll fluorescence [36,38]. The increase in leaf fluorescence transients of C. triste plantlets under different light qualities showed a typical OJIP shape (Figure 4). Under R treatment, it showed repressed fluorescence transients in the IP phase. This result was consistent with the previous study in Cyclocarya paliurus [88]. The increased value of F0 (p < 0.05, Table 3) suggested a decline in the light energy trapping rate in the PS II reaction center [89]. FM/F0, FV/FM, and FV/F0 are considered as indicators of the electron transfer activity and photosynthetic efficiency of plants [90] and could be used to evaluate the vigor and health of the photosynthetic apparatus [91]. The decreased values of FM/F0, FV/FM, and FV/F0 in leaves of C. triste plantlets grown under R (p < 0.05, Table 3) suggested that plants slightly underwent photoinhibition and had a reduced photochemical efficiency and PS II activity. M0 is related to QA reduction, and SM reflects the total energy required for QA reduction. The PS II fluorescence yield is positively correlated with the level of reduced QA, and oxidized QA quenched the fluorescence and leads to closure of the active reaction center [92]. The lowest M0 and highest SM values were observed in plantlets grown under RB (p < 0.05, Table 3). These suggested that RB induced more electrons transferring from QA– into the electron transport chain. In contrast, R had the opposite effect in the electron transport process from QA to QB. The similar decrease in the quantum yield of electron transport (φE0) (p < 0.05, Table 3) and the possibility of a trapped excitons moving an electron into the electron transport chain beyond QA (ψ0) (p < 0.05, Table 3) under R indicated that the acceptor side of PS II in the electron transport of PS II was inhibited. The ABS/RC represents the absorption flux of antenna chlorophyll per reaction center and reflects the ratio of photons absorbed by the chlorophyll pigments of reaction centers to the total numbers of active reaction centers [93]. The increased value of ABS/RC (p < 0.05, Table 3) indicated the decrease in antenna size of the reaction centers under R. Moreover, TR0/RC and ET0/RC represent the trapped energy flux and electron transport flux per reaction center, respectively, and the increased values under the R (p < 0.05, Table 3) were possibly due to the transportation of photons absorbed by inactive reaction center to active reaction centers. This was also verified with the increased value of heat dissipation per reaction center (DI0/RC) and the quantum yield of energy dissipation (φD0) (Table 3) [94]. Photons were unable to be captured effectively by the active centers, resulting in higher values of non-photochemical quenching. The above results indicated that C. triste plantlets grown under R underwent photoinhibition and had increased inactivated reaction centers and improved energy consumption in the activated reaction centers. PIABS is an indicator of plant vitality and reflects the functionality of both PS I and PS II. It is closely related to the activity of the photosynthetic apparatus. The significantly decreased PIABS (p < 0.05, Table 3) indicated that the energy conservation ability from photons absorbed by the PS II antenna to the reduction of QB and the activity of the photosynthetic apparatus were inhibited by R [95]. The reduced values of FV/FM and PIABS as well as increased values of parameters related to specific energy fluxes (ABS/RC, TR0/RC, ET0/RC, and DI0/RC), were observed in Cyclocarya paliurus and Brassica campestris under red or green LED as compared to blue or white LED [88,96]. Results were also found that monochromatic red light led to less efficiency in the transfer of excitons and higher energy dissipation in Calendula officinalis, indicating the occurrence of photoinhibition under the red light [97]. Plants grown under red light exhibit characteristics of shade plants, and a study in roses revealed that plants grown under monochromatic red or blue LED showed a more sensitive electron transport system under high light-intensity treatment as compared to those grown under combined red and blue or white LED [98]. The systematic analysis of parameters of chlorophyll a fluorescence revealed light energy trapping and transformation, electron transport, and activity of the photosynthetic apparatus under different light qualities. RB led to the highest values of M0 and PIABS, which indicated positive effects in the electron transport process from QA to QB and a higher activity of the photosynthetic apparatus.
Moreover, the sugar in the medium is thought to be the limitation of photosynthesis and development of photosynthetic apparatus. In several plant species cultured in in vitro conditions, the sugar reduces the need for carbohydrate production and therefore leads to a lower photosynthetic rate [99,100]. However, there are still studies showing the advantages of sugar on photosynthesis [101,102]. Plantlets treated without sugar showed deformed chloroplasts in the myrtle [103]. In this study, plantlets grown under 50 µmol·m−2·s−1 with 3% (w/v) sucrose did not show significant inhibitions of growth and photosynthesis. The 3% (w/v) sucrose revealed that it overcame the negative effect of strong light during culture on the functioning of photosynthesis and the xanthophyll cycle [104]. The interactions between sugar and light-induced signaling pathways in vitro are worth studying in the future.
ROSs could be derived from photochemical events, e.g., electron transport chains, as well as photorespiration under different light conditions, which can result in photodamage to PS II directly. Studies related to the effects of light qualities on nonenzymatic ROS scavengers showed that B stimulated the accumulation of total flavonoids and enhanced DPPH free radical scavenging capacity in callus cultures of Rhodiola imbricata [105] as well as the leaf and root extracts in Rehmannia glutinosa [106]. The combined red and blue LEDs in a 1:1 ratio also benefit the content of total flavonoids in Cnidium officinale callus and the activities of APX and SOD [107]. The present study also demonstrated similar findings, as shown in Figure 5, Figure 6 and Figure 7. CAT, APX, POD, and SOD are antioxidant enzymes reflecting the antioxidant capacity of plants. SOD, as the first activated antioxidant enzyme in the reactive oxygen scavenger process, catalyzes the superoxide anion free radicals (O2−) into hydrogen peroxide (H2O2) in plants. In the subsequent reactions, H2O2 is broken down by functions of CAT, APX, and the ascorbate–glutathione cycle [108,109,110]. A majority of classic PODs utilizes H2O2 to oxidize hydrogen donor substrates [111]. In addition to being involved in the defense responses of plants to biotic or abiotic stresses, antioxidant enzymes also positively regulate plant growth, differentiation, morphogenesis, and yield [112,113,114]. A previous study has shown that light quality could influence the metabolism of antioxidant enzymes [115]. RB significantly improved the activities of CAT and APX (p < 0.05, Figure 7A,B). The highest activities of POD and SOD were observed in the leaves of plantlets of C. triste grown under W and RB (p < 0.05, Figure 7C,D). Increased levels of ROS were observed in various plant species during senescence, and these changes may have resulted from the decline in the activities of certain antioxidant enzymes [116]. The decreased CAT activity was found in pea leaves which underwent senescence induced by a natural aging process or a dark condition [117]. Similarly, blue light caused higher specific activity of CAT in wheat as compared to other light treatments and delayed the onset of senescence of leaves, indicating the vital role of CAT in the prevention of senescence [118]. The increased activities of SOD, CAT, and POD as well as increased DPPH scavenging capacity in procyanidins-treated banana fruits suggested that these effectively scavenged excessive ROS and, therefore, alleviated lipid peroxidation and delayed senescence [119]. These findings show that the ability to scavenge superoxide anion radicals was improved under RB, which may delay senescence in plants. However, the antioxidant activity during senescence varied among plant species [116]. Elucidation of the mechanisms underlying plant senescence and ROS homeostasis will benefit with increasing yield and stress resistance.
5. Conclusions
This study investigated the growth, physiological responses, light energy trapping, transformation, and electron transport in the photosynthesis process of plants as affected by light quality. In conclusion, RB induced sturdy plantlets with well-developed shoots and roots; increased the content of photosynthetic pigments, photosynthetic electron transport, and efficiency and content of total flavonoids; and enhanced DPPH free radical scavenging capacity and activities of antioxidant enzymes in the plantlets. Hence, it could be used for the best production of high-quality C. triste plantlets in vitro. Future studies should focus on investigating the underlying molecular mechanisms of how combinations of monochromatic LEDs affect plants and the interactions between physiological regulatory and light-induced signaling pathways.
Author Contributions
Conceptualization, B.R.J.; methodology, B.R.J. and J.Z.; formal analysis, J.Z. and L.T.T.; resources, B.R.J. and Y.G.P.; data curation, J.Z. and L.T.T.; writing—original draft preparation, J.Z.; writing—review and editing, B.R.J.; project administration, B.R.J.; funding acquisition, B.R.J., Y.G.P., J.Z., and L.T.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Korea National Arboretum, project no. KNA1–2–25,16–3. J.Z. and L.T.T. were supported by a scholarship from the BK21 Plus Program, Ministry of Education, Republic of Korea.
Acknowledgments
This research was supported by the “useful plant collection and development of mass propagation protocol for the establishment of foundation of convergence platform using forest plants (KNA1–2–25,16–3)” project funded by the Korea National Arboretum. J.Z. and L.T.T. were supported by a scholarship from the BK21 Plus Program, Ministry of Education, Republic of Korea.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Moon, H.I. Compounds with antiproliferative activity on five human cancer cell lines from South Korean Carpesium triste. Nat. Prod. Commun. 2012, 7, 1934578X1200700704. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.P.; Wang, G.W.; Tian, X.H.; Yang, Y.X.; Liu, Q.X.; Chen, L.P.; Li, H.L.; Zhang, W.D. The genus Carpesium: A review of its ethnopharmacology, phytochemistry and pharmacology. J. Ethnopharmacol. 2015, 163, 173–191. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Hwang, K.H.; Kim, G.H. In vitro evaluation of anti–inflammatory activity for salad–food material Carpesium Abrotanoides. J. Food Biochem. 2013, 37, 18–25. [Google Scholar] [CrossRef]
- Xia, Q.; Luo, J.; Mei, X.; Wang, Y.; Huang, W.; Wang, J.; Yang, R.; Ma, Z.; Lin, R. A developmental toxicity assay of Carpesii Fructus on zebrafish embryos/larvae. Toxicol. Res. 2017, 6, 460–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Jin, G.; Tian, L.; Ebrahim, W.; Höfert, S.P.; Janiak, C.; Chen, J.F.; Guo, Z.Y.; Schäberle, T.F.; Liu, Z.; et al. New eremophilane–type sesquiterpenes and maleimide–bearing compounds from Carpesium abrotanoides L. Fitoterapia 2019, 138, 104294. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.R.; Suh, B.R.; Kim, J.G.; Ho Kim, Y.; Kim, D.K.; Moon, D.C. Sesquiterpene lactones from Carpesium triste var. manshuricum. Phytochemistry 1999, 52, 113–115. [Google Scholar] [CrossRef]
- Kim, M.R.; Lee, S.K.; Kim, C.S.; Kim, K.S.; Moon, D.C. Phytochemical constituents of Carpesium macrocephalum FR– et SAV. Arch. Pharm. Res. 2004, 27, 1029–1033. [Google Scholar] [CrossRef]
- Kim, M.R.; Hwang, B.Y.; Jeong, E.S.; Lee, Y.M.; Yoo, H.S.; Chung, Y.B.; Hong, J.T.; Moon, D.C. Cytotoxic germacranolide sesquiterpene lactones from Carpesium triste var. manshuricum. Arch. Pharm. Res. 2007, 30, 556. [Google Scholar] [CrossRef]
- Zee, O.P.; Kim, D.K.; Choi, S.U.; Lee, C.O.; Lee, K.R. A new cytotoxic acyclic diterpene from Carpesium divaricatum. Arch. Pharm. Res. 1999, 22, 225–227. [Google Scholar] [CrossRef]
- Gao, X.; Lin, C.J.; Jia, Z.J. Cytotoxic germacranolides and acyclic diterpenoides from the seeds of Carpesium triste. J. Nat. Prod. 2007, 70, 830–834. [Google Scholar] [CrossRef]
- Zee, O.P.; Kim, D.K.; Lee, K.R. Thymol derivatives from Carpesium divaricatum. Arch. Pharm. Res. 1998, 21, 618–620. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Chen, G. Study on terpenoid constituents from aerial part of Carpesium minum Hemsl. Chem. Res. Appl. 2012, 24, 1762–1765. [Google Scholar]
- Wu, H.B.; Wu, H.B.; Wang, W.S.; Liu, T.T.; Qi, M.G.; Feng, J.C.; Li, X.Y.; Liu, Y. Insecticidal activity of sesquiterpene lactones and monoterpenoid from the fruits of Carpesium abrotanoides. Ind. Crop. Prod. 2016, 92, 77–83. [Google Scholar] [CrossRef]
- Kami, C.; Lorrain, S.; Hornitschek, P.; Fankhauser, C. Light–regulated plant growth and development. Curr. Top. Dev. Biol. 2010, 91, 29–66. [Google Scholar]
- De Wit, M.; Galvão, V.C.; Fankhauser, C. Light–mediated hormonal regulation of plant growth and development. Annu. Rev. Plant Biol. 2016, 67, 513–537. [Google Scholar] [CrossRef]
- De Capite, L. Action of light and temperature on growth of plant tissue cultures in vitro. Am. J. Bot. 1955, 42, 869–873. [Google Scholar] [CrossRef]
- Eckardt, N.A. From darkness into light: Factors controlling photomorphogenesis. Plant Cell 2001, 13, 219–221. [Google Scholar] [CrossRef]
- Hu, W.; Lagarias, J.C. A tightly regulated genetic selection system with signaling–active alleles of phytochrome B. Plant Physiol. 2017, 173, 366–375. [Google Scholar] [CrossRef] [Green Version]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; Ieperen, W.V.; Harbinson, J. Blue light dose–responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef]
- Sun, W.; Ubierna, N.; Ma, J.Y.; Walker, B.J.; Kramer, D.M.; Cousins, A.B. The coordination of C4 photosynthesis and the CO2–concentrating mechanism in maize and Miscanthus × giganteus in response to transient changes in light quality. Plant Physiol. 2014, 164, 1283–1292. [Google Scholar] [CrossRef] [Green Version]
- Bantis, F.; Ouzounis, T.; Radoglou, K. Artificial LED lighting enhances growth characteristics and total phenolic content of Ocimum basilicum, but variably affects transplant success. Sci. Hortic. 2016, 198, 277–283. [Google Scholar] [CrossRef] [Green Version]
- Baque, M.A.; Hahn, E.J.; Paek, K.Y. Induction mechanism of adventitious root from leaf explants of Morinda citrifolia as affected by auxin and light quality. In Vitro Cell. Dev. Biol. Plant 2010, 46, 71–80. [Google Scholar] [CrossRef]
- Lin, Y.; Li, J.; Li, B.; He, T.; Chun, Z. Effects of light quality on growth and development of protocorm–like bodies of Dendrobium officinale in vitro. Plant Cell Tiss. Organ Cult. 2011, 105, 329–335. [Google Scholar] [CrossRef]
- Fankhauser, C.; Batschauer, A. Shadow on the plant: A strategy to exit. Cell 2016, 164, 15–17. [Google Scholar] [CrossRef] [Green Version]
- Franklin, K.A.; Whitelam, G.C. Light-quality regulation of freezing tolerance in Arabidopsis thaliana. Nat. Genet. 2007, 39, 1410–1413. [Google Scholar] [CrossRef]
- Li, Q.; Kubota, C. Effects of supplemental light quality on growth and phytochemicals of baby leaf lettuce. Environ. Exp. Bot. 2009, 67, 59–64. [Google Scholar] [CrossRef]
- Gubler, F.; Hughes, T.; Waterhouse, P.; Jacobsen, J. Regulation of dormancy in barley by blue light and after-ripening: Effects on abscisic acid and gibberellin metabolism. Plant Physiol. 2008, 147, 886–896. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.H.; Wheeler, R.M.; Sager, J.C.; Yorio, N.C.; Goins, G.D. Light–emitting diodes as an illumination source for plants: A review of research at Kennedy Space Center. Habitation 2005, 10, 71–78. [Google Scholar] [CrossRef]
- Bourget, C.M. An introduction to light–emitting diodes. HortScience 2008, 43, 1944–1946. [Google Scholar] [CrossRef] [Green Version]
- Heo, J.W.; Lee, C.W.; Paek, K.Y. Influence of mixed LED radiation on the growth of annual plants. J. Plant Biol. 2006, 49, 286–290. [Google Scholar] [CrossRef]
- Pham, M.D.; Hwang, H.; Park, S.W.; Cui, M.; Lee, H.; Chun, C. Leaf chlorosis, epinasty, carbohydrate contents and growth of tomato show different responses to the red/blue wavelength ratio under continuous light. Plant Physiol. Bioch. 2019, 141, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.W.; Lee, Y.B.; Kim, D.E.; Chang, Y.S.; Chun, C. Effects of supplementary LED lighting on growth and biochemical parameters in Dieffenbachia amoena ‘Camella’and Ficus elastica ‘Melany’. Korea J. Hortic. Sci. Technol. 2010, 28, 51–58. [Google Scholar]
- Jee, G. Sixty–three years since Kautsky: Chlorophyll a fluorescence. Aust. J. Plant Physiol. 1995, 22, 131–160. [Google Scholar]
- Stirbet, A.; Lazár, D.; Kromdijk, J. Chlorophyll a fluorescence induction: Can just a one–second measurement be used to quantify abiotic stress responses? Photosynthetica 2018, 56, 86–104. [Google Scholar] [CrossRef]
- Strasser, R.J. The grouping model of plant photosynthesis: Heterogeneity of photosynthetic units in thylakoids. In Photosynthesis: Structure and Molecular Organization of the Photosynthetic Apparatus; Akoyunoglou, G., Ed.; Balaban International Science Services: Philadelphia, PA, USA, 1981; Volume III, pp. 727–737. [Google Scholar]
- Strasser, R.J. The F0 and the O–J–I–P fluorescence rise in higher plants and algae. In Regulation of Chloroplast Biogenesis; Argyroudi–Akoyunoglou, J.H., Ed.; Springer: Boston, MA, USA, 1992; pp. 423–426. [Google Scholar]
- Strasser, R. On the OJIP fluorescence transient in leaves and D1 mutants of Chlamydomonas reinhardtii. In Research in Photosynthesis: Proceedings of the IXth International Congress on Photosynthesis, Nagoya, Japan, 30 August–4 September 1992; Murata, N., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1992; Volume II, pp. 29–32. [Google Scholar]
- Strasser, R.J.; Srivastava, A. Polyphasic chlorophyll a fluorescent transient in plants and cyanobacteria. Photochem. Photobiol. 1995, 61, 32–42. [Google Scholar] [CrossRef]
- Stirbet, A. On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and photosystem II: Basics and applications of the OJIP fluorescence transient. J. Photochem. Photobiol. B 2011, 104, 236–257. [Google Scholar] [CrossRef]
- Schreiber, U.; Neubauer, C.; Klughammer, C. Devices and methods for room–temperature fluorescence analysis. Philos. Trans. R. Soc. B 1989, 323, 241–251. [Google Scholar]
- Schansker, G.; Strasser, R.J. Quantification of non–QB–reducing centers in leaves using a far–red pre–illumination. Photosynth. Res. 2005, 84, 145–151. [Google Scholar] [CrossRef]
- Schansker, G.; Tóth, S.Z.; Holzwarth, A.R.; Garab, G. Chlorophyll a fluorescence: Beyond the limits of the QA model. Photosynth. Res. 2014, 120, 43–58. [Google Scholar] [CrossRef]
- Roháček, K. Chlorophyll fluorescence parameters: The definitions, photosynthetic meaning, and mutual relationships. Photosynthetica 2002, 40, 13–29. [Google Scholar] [CrossRef]
- Waszczak, C.; Carmody, M.; Kangasjärvi, J. Reactive oxygen species in plant signaling. Annu. Rev. Plant Biol. 2018, 69, 209–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oukarroum, A.; Bussotti, F.; Goltsev, V.; Kalaji, H.M. Correlation between reactive oxygen species production and photochemistry of photosystems I and II in Lemna gibba L. plants under salt stress. Environ. Exp. Bot. 2015, 109, 80–88. [Google Scholar] [CrossRef]
- Triantaphylidès, C.; Havaux, M. Singlet oxygen in plants: Production, detoxification and signaling. Trends Plant Sci. 2009, 14, 219–228. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Eghbaliferiz, S.; Iranshahi, M. Prooxidant activity of polyphenols, flavonoids, anthocyanins and carotenoids: Updated review of mechanisms and catalyzing metals. Phytother. Res. 2016, 30, 1379–1391. [Google Scholar] [CrossRef]
- Baglieri, A.; Cadili, V.; Monterumici, C.M.; Gennari, M.; Tabasso, S.; Montoneri, E.; Nardi, S.; Negre, M. Fertilization of bean plants with tomato plants hydrolysates. Effect on biomass production, chlorophyll content and N assimilation. Sci. Hortic. 2014, 176, 194–199. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Strasser, R.J.; Srivastava, A.; Tsimilli–Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. In Probing Photosynthesis: Mechanism, Regulation and Adaptation; Yunus, M., Pathre, U., Mohanty, P., Eds.; Taylor and Francis: Abingdon, UK, 2000; Chapter 25; pp. 445–483. [Google Scholar]
- Strasser, R.J.; Tsimilli–Michael, M.; Srivastava, A. Analysis of the chlorophyll fluorescence transient. In Chlorophyll Fluorescence: A Signature of Photosynthesis, Advances in Photosynthesis and Respiration; Papageorgiou, G.C., Ed.; Springer: Dordrecht, The Netherlands, 2004; Volume 19, pp. 321–362. [Google Scholar]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Packer, L., Ed.; Academic Press: New York, NY, USA, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate–specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Pütter, J. Peroxidase. In Method of Enzymatic Method Analysis; Bergmeyer, H.U., Ed.; Academic Press: New York, NY, USA, 1974; Volume 2, pp. 685–690. [Google Scholar]
- Beauchamp, C.; Fridovich, I. Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Manivannan, A.; Ren, X.; Kang, D.I.; Jeong, B.R. Effect of explant type and plant growth regulators on callus induction, growth and secondary metabolites production in Cnidium officinale Makino. Mol. Biol. Rep. 2018, 45, 1919–1927. [Google Scholar]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar]
- Khan, T.; Abbasi, B.H.; Khan, M.A.; Shinwari, Z.K. Differential effects of thidiazuron on production of anticancer phenolic compounds in callus cultures of Fagonia indica. Appl. Biochem. Biotechnol. 2016, 179, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Tóth, S.Z.; Schansker, G.; Garab, G.; Strasser, R.J. Photosynthetic electron transport activity in heat–treated barley leaves: The role of internal alternative electron donors to photosystem II. Biochim. Biophys. Acta. 2007, 1767, 295–305. [Google Scholar] [CrossRef] [Green Version]
- Dewir, Y.H.; Chakrabarty, D.; Kim, S.J.; Hahn, E.J.; Paek, K.Y. Effect of light–emitting diode on growth and shoot proliferation of Euphorbia millii and Spathiphyllum cannifolium. J. Korea Soc. Hortic. Sci. 2005, 46, 375. [Google Scholar]
- Moon, H.K.; Park, S.K.; Kim, Y.W.; Kim, C.S. Growth of Tsuru–rindo (Tripterospermum japonicum) cultured in vitro under various sources of light–emitting diode (LED) irradiation. J. Plant Biol. 2006, 49, 174–179. [Google Scholar] [CrossRef]
- Chung, J.P.; Huang, C.Y.; Dai, T.E. Spectral effects on embryogenesis and plantlet growth of Oncidium ‘Gower Ramsey’. Sci. Hortic. 2010, 124, 511–516. [Google Scholar] [CrossRef]
- Maas, F.M.; Bakx, E.J.; Morris, D.A. Photocontrol of stem elongation and dry weight partitioning in Phaseolus vulgaris L. by the blue-light content of photosynthetic photon flux. J. Plant Physiol. 1995, 146, 665–671. [Google Scholar] [CrossRef]
- Appelgren, M. Effects of light quality on stem elongation of Pelargonium in vitro. Sci. Hortic. 1991, 45, 345–351. [Google Scholar] [CrossRef]
- You, T.; Barnett, S.M. Effect of light quality on production of extracellular polysaccharides and growth rate of Porphyridium cruentum. Biochem. Eng. J. 2004, 19, 251–258. [Google Scholar] [CrossRef]
- Bonnett, H.T. Phytochrome regulation of endogenous bud development on root cultures of Convolvulus arvensis. Planta 1972, 106, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Hahn, E.J.; Heo, J.W.; Paek, K.Y. Effects of LEDs on net photosynthetic rate, growth and leaf stomata of chrysanthemum plantlets in vitro. Sci. Hortic. 2004, 101, 143–151. [Google Scholar] [CrossRef]
- Heo, J.W.; Shin, K.S.; Kim, S.K.; Paek, K.Y. Light quality affects in vitro growth of grape ‘Teleki 5BB’. J. Plant Biol. 2006, 49, 276–280. [Google Scholar] [CrossRef]
- Nhut, D.T.; Nam, N.B. Light emitting diodes (LEDs): An artificial lighting source for biological studies. In Proceedings of the 3rd International Conference of the Development of BME in Vietnam, Ho Chi Minh, Vietnam, 11–14 January 2010. [Google Scholar]
- Goins, G.D.; Yorio, N.C.; Sanwo, M.M.; Brown, C.S. Photomorphogenesis, photosynthesis, and seed yield of wheat plants grown under red light-emitting diodes (LEDs) with and without supplemental blue lighting. J. Exp. Bot. 1997, 48, 1407–1413. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Takamura, T.; Watanabe, H.; Endo, M.; Yanagi, T.; Okamoto, K. In vitro growth of Cymbidium plantlets cultured under superbright red and blue light-emitting diodes (LEDs). J. Hortic. Sci. Biotech. 1998, 73, 39–44. [Google Scholar] [CrossRef]
- Lee, S.H.; Tewari, R.K.; Hahn, E.J.; Paek, K.Y. Photon flux density and light quality induce changes in growth, stomatal development, photosynthesis and transpiration of Withania somnifera (L.) Dunal. plantlets. Plant Cell Tiss. Organ Cult. 2007, 90, 141–151. [Google Scholar] [CrossRef]
- Bellamine, J.; Penel, C.; Greppin, H. Proton pump and IAA sensitivity changes in spinach leaves during the flowering induction. Plant Physiol. Biochem. 1993, 31, 197–203. [Google Scholar]
- Moe, R.; Morgan, L.; Grindal, G. Growth and plant morphology of Cucumis sativus and Fuchsia x hybrid are influenced by light quality during the photoperiod and by diurnal temperature alternations. Acta Hortic. 2002, 580, 229–234. [Google Scholar] [CrossRef]
- Poudel, P.R.; Kataoka, I.; Mochioka, R. Effect of red–and blue–light–emitting diodes on growth and morphogenesis of grapes. Plant Cell Tiss. Org. 2008, 92, 147–153. [Google Scholar] [CrossRef]
- Kurepin, L.V.; Emery, R.N.; Pharis, R.P.; Reid, D.M. Uncoupling light quality from light irradiance effects in Helianthus annuus shoots: Putative roles for plant hormones in leaf and internode growth. J. Exp. Bot. 2007, 58, 2145–2157. [Google Scholar] [CrossRef] [Green Version]
- Weatherwax, S.C.; Ong, M.S.; Degenhardt, J.; Bray, E.A.; Tobin, E.M. The interaction of light and abscisic acid in the regulation of plant gene expression. Plant Physiol. 1996, 111, 363–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathy, B.C.; Brown, C.S. Root–shoot interaction in the greening of wheat seedlings grown under red light. Plant Physiol. 1995, 107, 407–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venema, J.H.; Villerius, L.; van Hasselt, P.R. Effect of acclimation to suboptimal temperature on chilling-induced photodamage: Comparison between a domestic and a high-altitude wild Lycopersicon species. Plant Sci. 2000, 152, 153–163. [Google Scholar] [CrossRef]
- Leong, T.Y.; Anderson, J.M. Adaptation of the thylakoid membranes of pea chloroplasts to light intensities. I. Study on the distribution of chlorophyll–protein complexes. Photosynth. Res. 1984, 5, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Tholen, D.; Pons, T.L.; Voesenek, L.A.C.J.; Poorter, H. Ethylene insensitivity results in down–regulation of Rubisco expression and photosynthetic capacity in tobacco. Plant Physiol. 2007, 144, 1305–1315. [Google Scholar] [CrossRef] [Green Version]
- Kurilčik, A.; Miklušytė–Čanova, R.; Dapkūnienė, S.; Žilinskaitė, S.; Kurilčik, G.; Tamulaitis, G.; Duchovskis, P.; Žukauskas, A. In vitro culture of Chrysanthemum plantlets using light–emitting diodes. Cent. Eur. J. Biol. 2008, 3, 161–167. [Google Scholar] [CrossRef]
- Bukhov, N.G.; Drozdova, I.S.; Bondar, V.V.; Mokronosov, A.T. Blue, red and blue plus red light control of chlorophyll content and CO2 gas exchange in barley leaves: Quantitative description of the effects of light quality and fluence rate. Physiol. Plant. 1992, 85, 632–638. [Google Scholar] [CrossRef]
- Goodwin, T.W.; Britton, G. Distribution and analysis of carotenoids. In Plant Pigments; Goodwin, T.W., Ed.; Academic Press: London, UK, 1988; pp. 61–132. [Google Scholar]
- Haehnel, W. Photosynthetic electron transport in higher plants. Annu. Rev. Plant Physiol. 1984, 35, 659–693. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, T.; Fang, S.; Zhou, M.; Qin, J. Responses of morphology, gas exchange, photochemical activity of photosystem II, and antioxidant balance in Cyclocarya paliurus to light spectra. Front. Plant Sci. 2018, 9, 1704. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Guo, P.; Michael, B.; Stefania, G.; Salvatore, C. Evaluation of chlorophyll content and fluorescence parameters as indicators of drought tolerance in barley. Agric. Sci. China 2006, 5, 751–757. [Google Scholar] [CrossRef]
- Shangguan, Z.; Shao, M.; Dyckmans, J. Effects of nitrogen nutrition and water deficit on net photosynthetic rate and chlorophyll fluorescence in winter wheat. J. Plant Physiol. 2000, 156, 46–51. [Google Scholar] [CrossRef]
- Clark, A.J.; Landolt, W.; Bucher, J.B.; Strasser, R.J. Beech (Fagus sylvatica) response to ozone exposure assessed with a chlorophyll fluorescence performance index. Environ. Pollut. 2000, 109, 501–507. [Google Scholar] [CrossRef]
- Ke, B. Photosynthesis–photobiochemistry and photobiophysics. In Advances in Photosynthesis; Ke, B., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001; Volume 10, pp. 1–763. [Google Scholar]
- Strasser, R.J.; Tsimilli–Michael, M.; Qiang, S.; Goltsev, V. Simultaneous in vivo recording of prompt and delayed fluorescence and 820–nm reflection changes during drying and after rehydration of the resurrection plant Haberlea rhodopensis. Biochim. Biophys. Acta 2010, 1797, 1313–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estaji, A.; Kalaji, H.M.; Karimi, H.R.; Roosta, H.R.; Moosavi–Nezhad, S.M. How glycine betaine induces tolerance of cucumber plants to salinity stress? Photosynthetica 2019, 57, 753–761. [Google Scholar] [CrossRef] [Green Version]
- Yusuf, M.A.; Kumar, D.; Rajwanshi, R.; Strasser, R.J.; Tsimilli–Michael, M.; Sarin, N.B. Overexpression of γ–tocopherol methyl transferase gene in transgenic Brassica juncea plants alleviates abiotic stress: Physiological and chlorophyll a fluorescence measurement. Biochim. Biophys. Acta 2010, 1797, 1428–1438. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Zhou, X.; Xu, R.; Wang, J.; Lin, Y.; Pang, J.; Wu, S.; Zhong, F. Comprehensive analysis of photosynthetic characteristics and quality improvement of purple cabbage under different combinations of monochromatic light. Front. Plant Sci. 2016, 7, 1788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aliniaeifard, S.; Seif, M.; Arab, M.; Zare Mehrjerdi, M.; Li, T.; Lastochkina, O. Growth and photosynthetic performance of Calendula officinalis under monochromatic red light. Int. J. Hortic. Sci. Technol. 2018, 5, 123–132. [Google Scholar]
- Bayat, L.; Arab, M.; Aliniaeifard, S.; Seif, M.; Lastochkina, O.; Li, T. Effects of growth under different light spectra on the subsequent high light tolerance in rose plants. AoB Plants 2018, 10, 1–17. [Google Scholar] [CrossRef]
- Van Huylenbroeck, J.M.; Debergh, P.C. Impact of sugar concentration in vitro on photosynthesis and carbon metabolism during ex vitro acclimatization of Spathiphyllum plantlets. Physiol. Plant. 1996, 96, 298–304. [Google Scholar] [CrossRef]
- Rybczyński, J.J.; Borkowska, B.; Fiuk, A.; Gawrońska, H.; Śliwińska, E.; Mikuła, A. Effect of sucrose concentration on photosynthetic activity of in vitro cultures Gentiana kurroo (Royle) germlings. Acta Physiol. Plant. 2007, 29, 445–453. [Google Scholar] [CrossRef]
- Trojan, A.; Gabrys, H. Chloroplast distribution in Arabidopsis thaliana (L.) depends on light conditions during growth. Plant Physiol. 1996, 111, 419–425. [Google Scholar] [CrossRef] [Green Version]
- Kadleček, P.; Rank, B.; Tichá, I. Photosynthesis and photoprotection in Nicotiana tabacum L. in vitro-grown plantlets. J. Plant Physiol. 2003, 160, 1017–1024. [Google Scholar] [CrossRef] [PubMed]
- Lucchesini, M.; Monteforti, G.; Mensuali-Sodi, A.; Serra, G. Leaf ultrastructure, photosynthetic rate and growth of myrtle plantlets under different in vitro culture conditions. Biol. Plant. 2006, 50, 161–168. [Google Scholar] [CrossRef]
- Eckstein, A.; Zięba, P.; Gabryś, H. Sugar and light effects on the condition of the photosynthetic apparatus of Arabidopsis thaliana cultured in vitro. J. Plant Growth Regul. 2012, 31, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, S.; Raghuvanshi, R.; Bhardwaj, P.; Sood, H.; Saxena, S.; Chaurasia, O.P. Influence of light quality on growth, secondary metabolites production and antioxidant activity in callus culture of Rhodiola imbricata Edgew. J. Photoch. Photobio. B Biol. 2018, 183, 258–265. [Google Scholar] [CrossRef]
- Manivannan, A.; Soundararajan, P.; Halimah, N.; Ko, C.H.; Jeong, B.R. Blue LED light enhances growth, phytochemical contents, and antioxidant enzyme activities of Rehmannia glutinosa cultured in vitro. Hortic. Environ. Biotechnol. 2015, 56, 105–113. [Google Scholar] [CrossRef]
- Manivannan, A.; Ren, X.; Jeong, B.R. Light elicited growth, antioxidant enzymes activities and production of medicinal compounds in callus culture of Cnidium officinale Makino. J. Photoch. Photobio. B Biol. 2019, 196, 111509. [Google Scholar]
- Hoque, M.A.; Banu, M.N.A.; Okuma, E.; Amako, K.; Nakamura, Y.; Shimoishi, Y.; Murata, Y. Exogenous proline and glycinebetaine increase NaCl–induced ascorbate–glutathione cycle enzyme activities, and proline improves salt tolerance more than glycinebetaine in tobacco Bright Yellow–2 suspension–cultured cells. J. Plant Physiol. 2007, 164, 1457–1468. [Google Scholar] [CrossRef]
- Mittova, V.; Volokita, M.; Guy, M.; Tal, M. Activities of SOD and the ascorbate-glutathione cycle enzymes in subcellular compartments in leaves and roots of the cultivated tomato and its wild salt-tolerant relative Lycopersicon pennellii. Physiol. Plant. 2000, 110, 42–51. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defense antioxidants–superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defense grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Veitch, N.C. Structural determinants of plant peroxidase function. Phytochem. Rev. 2004, 3, 3–18. [Google Scholar] [CrossRef]
- Genkov, T.; Ivanova, I. Effect of cytokinin–active phenylurea derivatives on shoot multiplication, peroxidase and superoxide dismutase activities of in vitro cultured carnation. Bulg. J. Plant Physiol. 1995, 21, 73. [Google Scholar]
- Blazquez, S.; Olmos, E.; Hernández, J.A.; Fernández–García, N.; Fernández, J.A.; Piqueras, A. Somatic embryogenesis in saffron (Crocus sativus L.). Histological differentiation and implication of some components of the antioxidant enzymatic system. Plant Cell Tiss. Organ Cult. 2009, 97, 49–57. [Google Scholar] [CrossRef]
- Shafi, A.; Chauhan, R.; Gill, T.; Swarnkar, M.K.; Sreenivasulu, Y.; Kumar, S.; Kumar, N.; Shankar, R.; Ahuja, P.S.; Singh, A.K. Expression of SOD and APX genes positively regulates secondary cell wall biosynthesis and promotes plant growth and yield in Arabidopsis under salt stress. Plant Mol. Biol. 2015, 87, 615–631. [Google Scholar] [CrossRef]
- Shohael, A.M.; Ali, M.B.; Yu, K.W.; Hahn, E.J.; Islam, R.; Paek, K.Y. Effect of light on oxidative stress, secondary metabolites and induction of antioxidant enzymes in Eleutherococcus senticosus somatic embryos in bioreactor. Process. Biochem. 2006, 41, 1176–1185. [Google Scholar] [CrossRef]
- Prochazkova, D.; Wilhelmova, N. Leaf senescence and activities of the antioxidant enzymes. Biol. Plant. 2007, 51, 401–406. [Google Scholar] [CrossRef]
- Pastori, G.M.; del Río, L.A. Natural senescence of pea leaves (an activated oxygen-mediated function for peroxisomes). Plant Physiol. 1997, 113, 411–418. [Google Scholar] [CrossRef] [Green Version]
- Causin, H.F.; Jauregui, R.N.; Barneix, A.J. The effect of light spectral quality on leaf senescence and oxidative stress in wheat. Plant Sci. 2006, 171, 24–33. [Google Scholar] [CrossRef]
- Chen, J.; Li, F.; Li, Y.; Wang, Y.; Wang, C.; Yuan, D.; Jiang, Y. Exogenous procyanidin treatment delays senescence of harvested banana fruit by enhancing antioxidant responses and in vivo procyanidin content. Postharvest Biol. Tec. 2019, 158, 110999. [Google Scholar] [CrossRef]
Figure 1.
Spectral qualities of the different light sources used in the experiment: W, white LED; R, red LED; B, blue LED; and RB, combined red and blue LEDs in a 1:1 ratio.
Figure 1.
Spectral qualities of the different light sources used in the experiment: W, white LED; R, red LED; B, blue LED; and RB, combined red and blue LEDs in a 1:1 ratio.

Figure 2.
A representative picture of C. triste plantlets as affected by light quality after four weeks of culture: The light qualities used were W, white LED; R, red LED; B, blue LED; and RB, combined red and blue LEDs in a 1:1 ratio. The arrows show mature leaves of plantlets which were treated with different light qualities and used for measurement of parameters of chlorophyll a fluorescence.
Figure 2.
A representative picture of C. triste plantlets as affected by light quality after four weeks of culture: The light qualities used were W, white LED; R, red LED; B, blue LED; and RB, combined red and blue LEDs in a 1:1 ratio. The arrows show mature leaves of plantlets which were treated with different light qualities and used for measurement of parameters of chlorophyll a fluorescence.

Figure 3.
Effect of light quality on contents of chlorophyll a (A) and chlorophyll b (B), total chlorophyll (C), chlorophyll a/b ratio (D), and content of total carotenoids (E) in leaves of C. triste plantlets after four weeks of culture: The light qualities used were W, white LED; R, red LED; B, blue LED; and RB, combined red and blue LEDs in a 1:1 ratio. The results were expressed as means ± standard errors (n = 3). Different letters indicate significant differences (p < 0.05) by the Tukey’s multiple range test.
Figure 3.
Effect of light quality on contents of chlorophyll a (A) and chlorophyll b (B), total chlorophyll (C), chlorophyll a/b ratio (D), and content of total carotenoids (E) in leaves of C. triste plantlets after four weeks of culture: The light qualities used were W, white LED; R, red LED; B, blue LED; and RB, combined red and blue LEDs in a 1:1 ratio. The results were expressed as means ± standard errors (n = 3). Different letters indicate significant differences (p < 0.05) by the Tukey’s multiple range test.

Figure 4.
Effect of light quality on OJIP transient of dark-adapted C. triste leaves plotted on a logarithmic time scale (0.01 ms to 1 s): The light qualities used were W, white LED; R, red LED; B, blue LED; and RB, combined red and blue LEDs in a 1:1 ratio.
Figure 4.
Effect of light quality on OJIP transient of dark-adapted C. triste leaves plotted on a logarithmic time scale (0.01 ms to 1 s): The light qualities used were W, white LED; R, red LED; B, blue LED; and RB, combined red and blue LEDs in a 1:1 ratio.

Figure 5.
Effect of light quality on the content of total flavonoids in leaves of C. triste plantlets after four weeks of culture: The light qualities used were W, white LED; R, red LED; B, blue LED; and RB, combined red and blue LEDs in a 1:1 ratio. The results were expressed as means ± standard errors (n = 3). Different letters indicate significant differences (p < 0.05) by the Tukey’s multiple range test.
Figure 5.
Effect of light quality on the content of total flavonoids in leaves of C. triste plantlets after four weeks of culture: The light qualities used were W, white LED; R, red LED; B, blue LED; and RB, combined red and blue LEDs in a 1:1 ratio. The results were expressed as means ± standard errors (n = 3). Different letters indicate significant differences (p < 0.05) by the Tukey’s multiple range test.

Figure 6.
Effect of light quality on 2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging capacity in leaves of C. triste plantlets after four weeks of culture: The light qualities used were W, white LED; R, red LED; B, blue LED; and RB, combined red and blue LEDs in a 1:1 ratio. The results were expressed as means ± standard errors (n = 3). Different letters indicate significant differences (p < 0.05) by the Tukey’s multiple range test.
Figure 6.
Effect of light quality on 2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging capacity in leaves of C. triste plantlets after four weeks of culture: The light qualities used were W, white LED; R, red LED; B, blue LED; and RB, combined red and blue LEDs in a 1:1 ratio. The results were expressed as means ± standard errors (n = 3). Different letters indicate significant differences (p < 0.05) by the Tukey’s multiple range test.

Figure 7.
Effect of light quality on activities of catalase (CAT) (A), ascorbate peroxidase APX (B), peroxidase (POD) (C), and superoxide dismutase (SOD) (D) in leaves of C. triste plantlets after four weeks of culture: The light qualities used were W, white LED; R, red LED; B, blue LED; and RB, combined red and blue LEDs in a 1:1 ratio. The results were expressed as means ± standard errors (n = 3). Different letters indicate significant differences (p < 0.05) by the Tukey’s multiple range test.
Figure 7.
Effect of light quality on activities of catalase (CAT) (A), ascorbate peroxidase APX (B), peroxidase (POD) (C), and superoxide dismutase (SOD) (D) in leaves of C. triste plantlets after four weeks of culture: The light qualities used were W, white LED; R, red LED; B, blue LED; and RB, combined red and blue LEDs in a 1:1 ratio. The results were expressed as means ± standard errors (n = 3). Different letters indicate significant differences (p < 0.05) by the Tukey’s multiple range test.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Extracted and technical parameters deduced from chlorophyll a fluorescence induction curve OJIP.
Table 1.
Extracted and technical parameters deduced from chlorophyll a fluorescence induction curve OJIP.
| Formula Abbreviation | Explanation |
|---|---|
| F0 | Minimum fluorescence in the dark-adapted state |
| FM | Maximum fluorescence at a saturating radiation pulse in the dark-adapted state |
| FV | Maximum variable fluorescence |
| FV/FM | Maximum quantum efficiency of PS II photochemistry |
| FM/F0 | Electron transfer activity |
| FV/F0 | Actual quantum yield of PS II |
| M0 = TR0/RC − ET0/RC = 4 (F300μs − F0)/(FM − F0) | Approximated initial slope of the fluorescence transient normalized on the FV |
| VJ = (FJ − F0)/FV | Relative variable fluorescence for J state |
| SM = Area/(FM − F0) | Normalized total complementary area |
| φP0 = (1 − F0)/F | Quantum yield of the QA reduction |
| φE0 = (1 − (F0/FM))·ψ0 | Quantum yield for electron transport |
| φD0 = 1 − φP0 | The quantum yield of energy dissipation |
| Ψ0 = 1 − VJ | Possibility of a trapped exciton moves an electron into the electron transport chain beyond QA |
| ABS/RC = M0·(1/VJ)·(FM/FV) | Absorption flux of antenna chlorophyll per reaction center |
| ET0/RC = M0·(1/VJ)·φE0 | Electron transport flux per reaction center |
| TR0/RC = M0·(1/VJ) | Trapped energy flux per reaction center |
| DI0/RC = ABS/RC − TR0/RC | Heat dissipation per reaction center |
| PIABS = (RC/ABS)·(φP0/(1 − φP0))·(ψ0/(1 − ψ0)) | Performance index per absorption flux |
Table 2.
Effect of light quality on growth and development of C. triste plantlets after four weeks of culture.
Table 2.
Effect of light quality on growth and development of C. triste plantlets after four weeks of culture.
| Light Quality | Shoot | Root | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Number | Length (cm) | Fresh Weight (g) | Dry Weight (g) | Diameter (mm) | Number | Length (cm) | Fresh Weight (g) | Dry Weight (g) | |
| W z | 5 ± 0.00 y a x | 10.19 ± 0.43 b | 0.66 ± 0.06 c | 0.09 ± 0.01 c | 5.72 ± 0.20 b | 6 ± 1.00 b | 4.78 ± 0.52 b | 1.18 ± 0.12 a | 0.16 ± 0.02 a |
| R | 3 ± 0.58 b | 14.30 ± 0.56 a | 0.97 ± 0.08 b | 0.13 ± 0.01 b | 5.22 ± 0.09 c | 8 ± 0.58 ab | 5.77 ± 0.30 a | 0.74 ± 0.04 b | 0.10 ± 0.01 b |
| B | 4 ± 0.58 b | 5.52 ± 0.22 d | 0.43 ± 0.04 d | 0.06 ± 0.01 d | 4.38 ± 0.10 d | 3 ± 0.58 c | 2.62 ± 0.22 c | 0.25 ± 0.05 c | 0.03 ± 0.01 c |
| RB | 6 ± 0.58 a | 7.00 ± 0.49 c | 1.25 ± 0.12 a | 0.17 ± 0.02 a | 6.11 ± 0.11 a | 9 ± 0.58 a | 5.55 ± 0.23 ab | 1.24 ± 0.10 a | 0.17 ± 0.01 a |
z The light qualities used were W, white LED; R, red LED; B, blue LED; and RB, combined red and blue LEDs in a 1:1 ratio. y Values represent the means ± standard errors (n = 9). x Means separation within columns were performed by the Tukey’s multiple range test at the 5% level. Different letters indicate significant differences (p < 0.05) by the Tukey’s multiple range test.
Table 3.
Effect of light quality on parameters of chlorophyll a fluorescence in leaves of C. triste cultured for 4 weeks.
Table 3.
Effect of light quality on parameters of chlorophyll a fluorescence in leaves of C. triste cultured for 4 weeks.
| Chlorophyll a Fluorescence Parameter | Light Quality | |||
|---|---|---|---|---|
| W z | R | B | RB | |
| F0 | 7954 ± 696 y ab x | 8399 ± 568 a | 7087 ± 326 ab | 6947 ± 443 b |
| FM | 46,827 ± 2489 a | 44,345 ± 1651 a | 43,532 ± 1297 a | 42,145 ± 2878 a |
| FV/FM | 0.83 ± 0.01 ab | 0.81 ± 0.02 b | 0.84 ± 0.00 a | 0.84 ± 0.00 a |
| FM/F0 | 5.90 ± 0.23 ab | 5.30 ± 0.49 b | 6.15 ± 0.12 a | 6.07 ± 0.03 a |
| FV/F0 | 4.90 ± 0.23 ab | 4.30 ± 0.49 b | 5.15 ± 0.12 a | 5.07 ± 0.03 a |
| M0 | 0.59 ± 0.05 ab | 0.76 ± 0.02 a | 0.57 ± 0.05 b | 0.53 ± 0.02 b |
| SM | 382 ± 14 c | 408 ± 19 c | 478 ± 19 b | 533 ± 14 a |
| φE0 | 0.52 ± 0.01 ab | 0.49 ± 0.02 b | 0.54 ± 0.00 a | 0.55 ± 0.02 a |
| φD0 | 0.17 ± 0.01 ab | 0.19 ± 0.02 a | 0.16 ± 0.00 b | 0.17 ± 0.00 ab |
| ψ0 | 0.63 ± 0.01 ab | 0.61 ± 0.02 b | 0.65 ± 0.00 ab | 0.66 ± 0.02 a |
| ABS/RC | 1.93 ± 0.12 ab | 2.29 ± 0.21 a | 1.94 ± 0.16 ab | 1.84 ± 0.15 b |
| ET0/RC | 1.02 ± 0.04 b | 1.13 ± 0.00 a | 1.05 ± 0.03 ab | 1.02 ± 0.06 b |
| TR0/RC | 1.61 ± 0.09 a | 1.92 ± 0.04 b | 1.62 ± 0.04 a | 1.51 ± 0.07 a |
| DI0/RC | 0.33 ± 0.03 b | 0.45 ± 0.03 a | 0.32 ± 0.03 b | 0.30 ± 0.02 b |
| PIABS | 4.44 ± 0.12 b | 2.98 ± 0.67 c | 4.89 ± 0.21 ab | 5.76 ± 0.08 a |
z The light qualities used were W, white LED; R, red LED; B, blue LED; and RB, combined red and blue LEDs in a 1:1 ratio. y Values represent means ± standard errors (n = 9). x Means separation within rows were performed by the Tukey’s multiple range test at the 5% level. Different letters indicate significant differences (p < 0.05) by the Tukey’s multiple range test.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhao, J.; Thi, L.T.; Park, Y.G.; Jeong, B.R. Light Quality Affects Growth and Physiology of Carpesium triste Maxim. Cultured In Vitro. Agriculture 2020, 10, 258. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10070258
AMA Style
Zhao J, Thi LT, Park YG, Jeong BR. Light Quality Affects Growth and Physiology of Carpesium triste Maxim. Cultured In Vitro. Agriculture. 2020; 10(7):258. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10070258
Chicago/Turabian StyleZhao, Jin, Luc The Thi, Yoo Gyeong Park, and Byoung Ryong Jeong. 2020. "Light Quality Affects Growth and Physiology of Carpesium triste Maxim. Cultured In Vitro" Agriculture 10, no. 7: 258. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10070258
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.