Development of Seed Production Technology of CIMMYT Tropical Single Cross Maize Hybrids

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genetic Materials and Experimental Locations
2.2. Details of Experiments
2.3. Data Collection and Analysis
3. Results and Discussion
3.1. Parental Seed Management
3.1.1. Male Inbred Line T10
3.1.2. Female Inbred Lines
3.2. Hybrid Seed Production
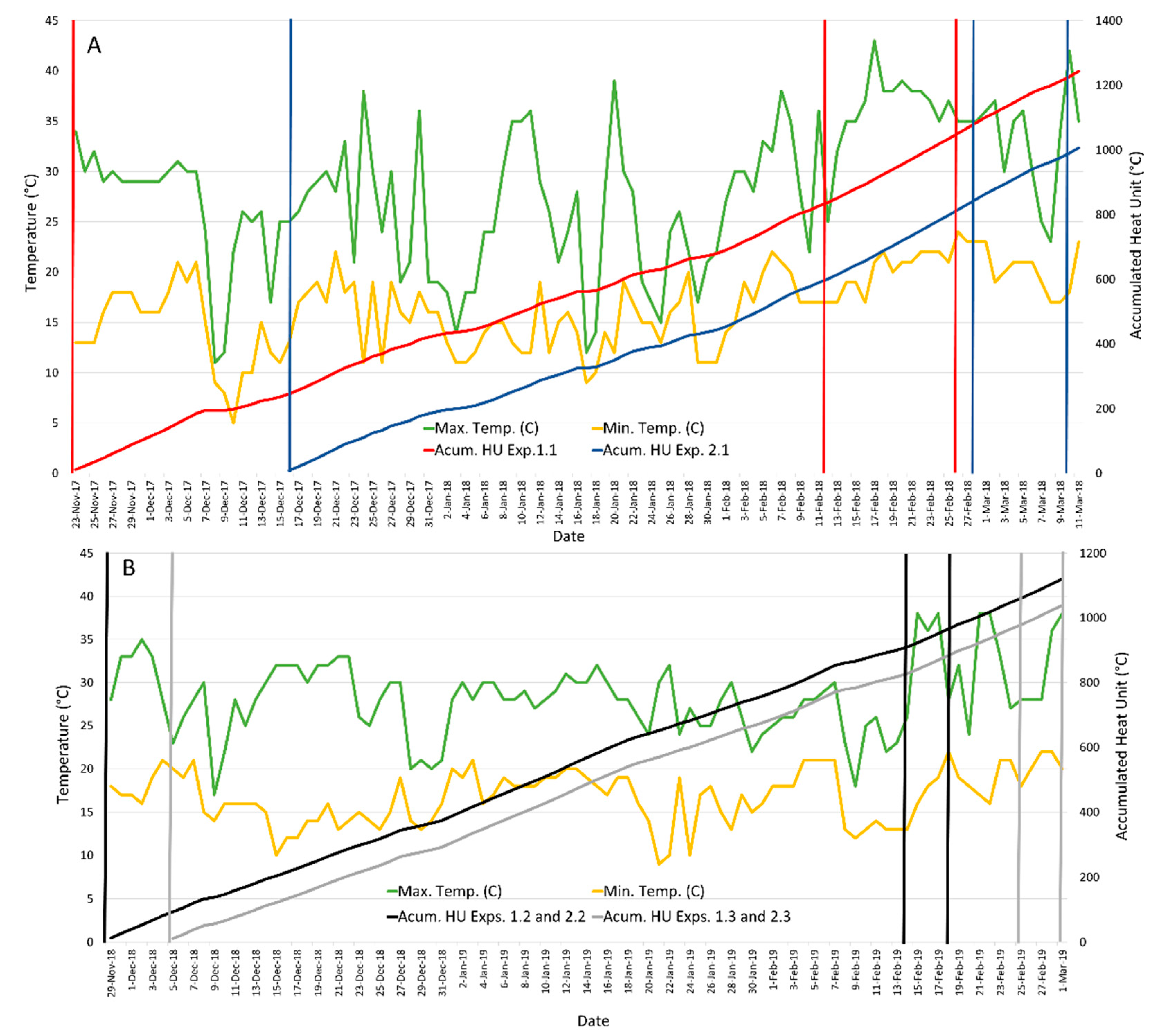
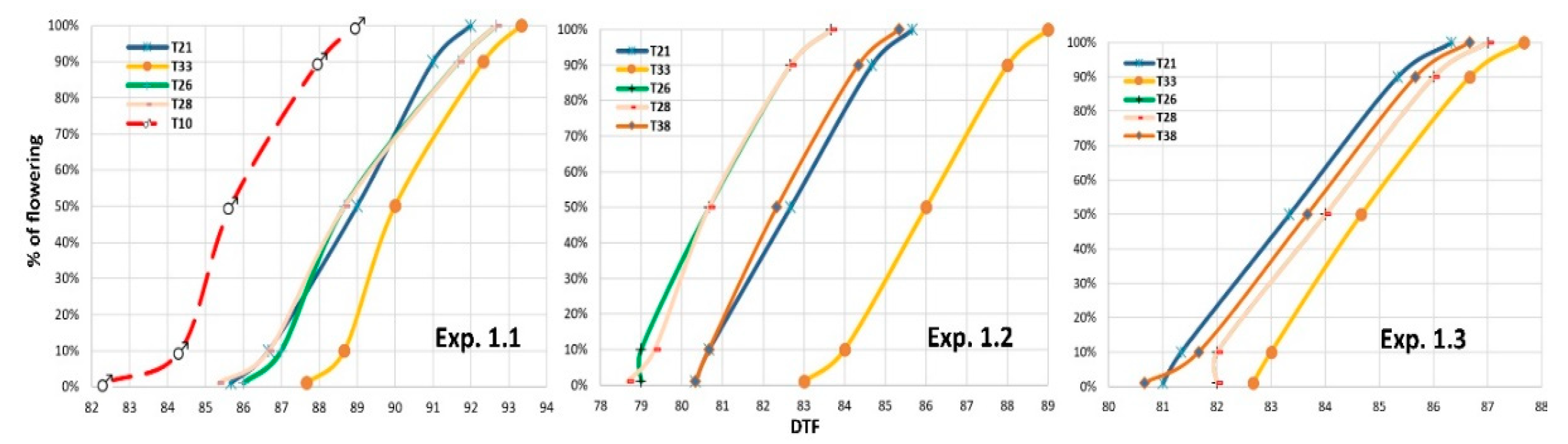
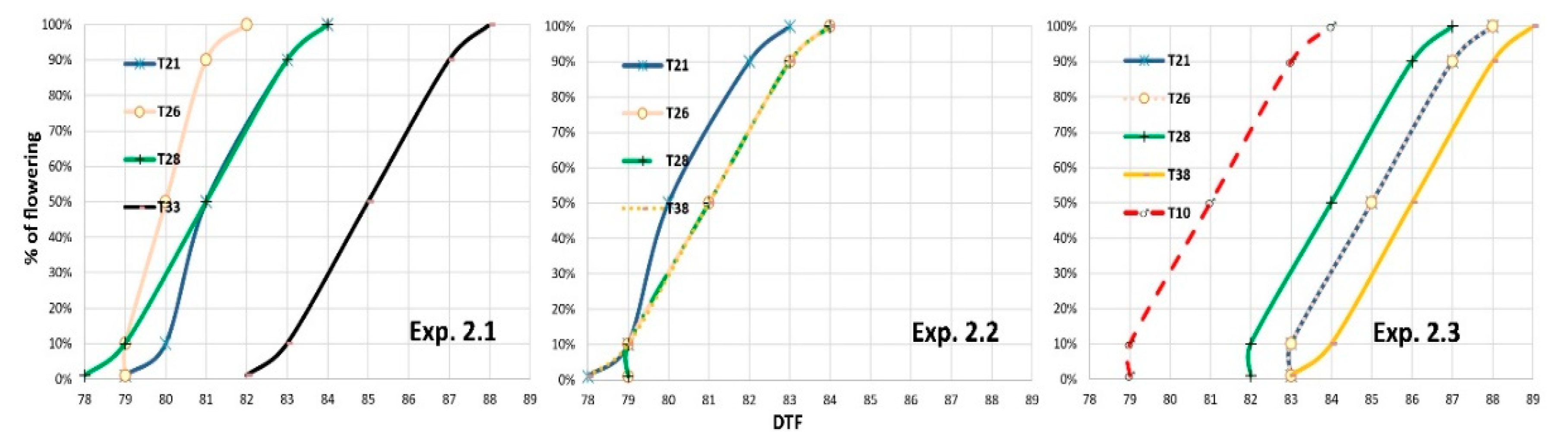
3.2.1. Floral Synchronization for Hybrid Seed Production
3.2.2. Female Height of Stigmas and Male Height Tassels
3.2.3. Effective Seed Yield
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| DAP | 23 Nov. 2017 | 16 Dec. 2017 | 29 Nov. 2018 | 05 Dec. 2018 | ||||
|---|---|---|---|---|---|---|---|---|
| HU day | HU acum. | HU day | HU acum. | HU day | HU acum. | HU day | HU acum. | |
| 75 | 15.0 | 744.5 | 16.5 | 848.0 | 10.0 | 884.5 | 14.5 | 869.0 |
| 76 | 16.0 | 760.5 | 16.5 | 864.5 | 7.5 | 892.0 | 15.0 | 884.0 |
| 77 | 15.5 | 776.0 | 14.5 | 879.0 | 8.0 | 900.0 | 14.5 | 898.5 |
| 78 | 15.0 | 791.0 | 15.0 | 894.0 | 9.5 | 909.5 | 11.0 | 909.5 |
| 79 | 12.5 | 803.5 | 15.5 | 909.5 | 13.0 | 922.5 | 13.5 | 923.0 |
| 80 | 9.5 | 813.0 | 15.5 | 925.0 | 14.0 | 936.5 | 13.0 | 936.0 |
| 81 | 13.5 | 826.5 | 15.5 | 940.5 | 14.5 | 951.0 | 15.5 | 951.5 |
| 82 | 11.0 | 837.5 | 12.0 | 952.5 | 15.0 | 966.0 | 14.0 | 965.5 |
| 83 | 13.5 | 851.0 | 10.0 | 962.5 | 14.5 | 980.5 | 13.0 | 978.5 |
| 84 | 14.5 | 865.5 | 13.5 | 976.0 | 11.0 | 991.5 | 14.0 | 992.5 |
| 85 | 14.5 | 880.0 | 14.0 | 990.0 | 13.5 | 1005.0 | 15.0 | 1007.5 |
| 86 | 13.5 | 893.5 | 16.5 | 1006.5 | 13.0 | 1018.0 | 16.0 | 1023.5 |
| 87 | 15.5 | 909.0 | 11.5 | 1018.0 | 15.5 | 1033.5 | 15.0 | 1038.5 |
| 88 | 16.0 | 925.0 | 9.5 | 1027.5 | 14.0 | 1047.5 | 15.0 | 1053.5 |
| 89 | 15.0 | 940.0 | 14.0 | 1041.5 | 13.0 | 1060.5 | 15.5 | 1069.0 |
| 90 | 15.5 | 955.5 | 14.0 | 1055.5 | 14.0 | 1074.5 | 14.0 | 1083.0 |
References
- FAO Cereal Supply and Demand Brief. Available online: http://www.fao.org/worldfoodsituation/csdb/en/ (accessed on 2 May 2020).
- FAO. Crop Prospects and Food Situation—Quarterly Global Report No. 1; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Córdova-Téllez, L.; Caballero-García, M.A.; Hernández-Nicolás, N.Y.; Ríos-Santos, E. Boletín Informativo de Producción de Semilla Calificada por el SNICS; SADER: Mexico City, Mexico, 2019; 96p. [Google Scholar]
- Córdova-Téllez, L.; González-Santos, R.; Padilla-Vaca, E.; Chávez-Bravo, J. Acciones del Servicio Nacional de Inspección y Certificación de Semillas (SNICS-SAGARPA) para el desarrollo del campo mexicano. Agroproductividad 2018, 11, 3–8. [Google Scholar]
- Jugenheimer, R.W. Maíz Variedades Mejoradas, Métodos de Cultivo y Producción de Semillas; LIMUSA: Mexico City, Mexico, 1990; pp. 489–502. [Google Scholar]
- Espinosa-Calderón, A.; Sierra-Macías, M.; Gómez-Montiel, N. Seed technology and production of improved maize varieties from INIFAP without PRONASE. Agron. Mesoam. 2002, 14, 117–121. [Google Scholar] [CrossRef] [Green Version]
- Espinosa, A.; Gómez, N.; Sierra, M.; Betanzos, E.; Caballero, F.; Coutiño, B.; Palafox, A.; Rodríguez, F.; García, A.; Cano, O. Technology and seed production of maize hybrids and open pollinated varieties of quality protein maize (QPM) in Mexico. Agron. Mesoam. 2003, 14, 223–228. [Google Scholar] [CrossRef] [Green Version]
- McCoy, A.; Roth, M.; Shay, R.; Noel, Z.; Jayawardana, M.; Longley, R.; Bonito, G.; Chilvers, M. Identification of fungal communities within the tar spot complex of corn in Michigan via next-generation sequencing. Phytobiomes J. 2019, 3, 235–243. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, R.S.; Hernández, R.V. Producción de Semilla de maíz Caso CIR Pacífico Centro; Folleto Técnico Núm. 4; Campo Experimental Centro-Altos de Jalisco: Tepatitlán de Morelos, Mexico, 2017; 61p. [Google Scholar]
- Noriega-González, L.; Preciado-Ortiz, R.; Andrio-Enríquez, E.; Terrón-Ibarra1, A.; Covarrubias-Prieto, J. Phenology, plant growth and floral synchrony of the parental lines of H-374C QPM maize hybrid. Rev. Mex. Cienc. Agríc. 2011, 2, 489–500. [Google Scholar] [CrossRef] [Green Version]
- Parthasarathi, T.; Velu, G.; Prabhakaran, J. Impact of crop heat units on growth and developmental physiology of future crop production: A Review. R Res. Rev. J. Crop Sci. Technol. 2013, 2, 1–11. [Google Scholar]
- Dahmardeh, M. Effects of sowing date on the growth and yield of maize cultivars (Zea mays L.) and the growth temperature requirements. Afr. J. Biotechnol. 2012, 11, 12450–12453. [Google Scholar] [CrossRef]
- Ghamghami, M.; Ghahreman, N.; Irannejad, P.; Ghorbaniet, K. Comparison of data mining and GDD-based models in discrimination of maize phenology. Int. J. Plant Prod. 2019, 13, 11–22. [Google Scholar] [CrossRef]
- MacRobert, J.F.; Setimela, P.S.; Gethi, J.; Worku, M. Maize Hybrid Seed Production Manual; International Maize and Wheat Improvement Center (CIMMYT): Mexico City, Mexico, 2014; pp. 1–26. [Google Scholar]
- Romero, J.; Willcox, M.; Burgueno, J.; Romay, C.; Swarts, K.; Trachsel, S.; Preciado, E.; Terron, A.; Delgado, H.; Vidal, V.; et al. A study of allelic diversity underlying flowering-time adaptation in maize landraces. Nat. Genet. 2017, 49, 476–480. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Bravo, A.; Vidal-Martínez, V.; Bojórquez-Serrano, J.; García-Paredes, D. Respuesta del maíz al impacto ambiental ocurrido en las etapas de floración y ciclo vegetativo. Rev. Mex. Cienc. Agríc. 2014, 10, 2035–2045. [Google Scholar] [CrossRef] [Green Version]
- Vallejo, D.H.L.; Ramírez, J.L.; Chuela, M.; Ramírez, R. Manual de Producción de Semilla de Maíz. Estudio de Caso; Folleto Técnico Núm. 14; Campo Experimental Uruapan, INIFAP, CIRPAC: Guadalajara, Mexico, 2008; 96p. [Google Scholar]
- Virgen-Vargas, J.; Zepeda-Bautista, R.; Ávila-Perches, M.; Espinosa-Calderón, A.; Arellano-Vázquez, J.; Gámez-Vázquez, A. Producción de semilla de líneas progenitoras de maíz: Densidad de población e interacción. Agron. Mesoam. 2014, 25, 323–335. [Google Scholar] [CrossRef]
- El-Abady, M.I. Influence of maize seed size/shape, planted at different depths and temperatures on seed emergence and seedling vigor. Res. J. Seed Sci. 2015, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Torres, F.; Lewis, B.D.; Carballo, A.; Estrada, G. Técnicas para sincronizar floración en la producción de semilla híbrida de maíz. Agric. Téc. Méx. 2004, 30, 89–100. [Google Scholar]
- Espinosa, A.; Tadeo, M. Evaluación de desespigue mecánico en híbridos dobles de maíz, en los valles altos de México. Agron. Mesoam. 2016, 9, 90–92. [Google Scholar] [CrossRef]
- Cervantes-Ortiz, F.; Cadenas-Tepoxteco, J.; Raya-Pérez, J.; Andrio-Enríquez, E.; Rangel-Lucio, J.; Guevara-Acevedo, L.; Rodríguez-Herrera, S.; Mendoza-Elos, M. Silk Balling answer to soil moisture and population density in maize lines. Rev. Mex. Cienc. Agríc. 2015, 6, 231–241. [Google Scholar] [CrossRef] [Green Version]



| Female Inbred Lines | Male Inbred Line | Pedigree | Single-Cross Hybrids | Year of Release |
|---|---|---|---|---|
| T21 | T10 | T21/T10 | CLTHW14001 | 2015 |
| T33 | T33/T10 | CLTHW15002 | 2016 | |
| T26 | T26/T10 | CLTHW15005 | 2016 | |
| T28 | T28/T10 | CLTHW15007 | 2016 | |
| T38 | T38/T10 | CLTHW15109 | 2017 |
| Exp. 1 | Exp. 2 | ||||
|---|---|---|---|---|---|
| 2018A | 2019A | 2018A | 2019A | ||
| Exp. 1.1 23 Nov. 2017 | Exp. 1.2 29 Nov. 2018 | Exp. 1.3 05 Dec. 2018 | Exp. 2.1 16 Dec. 2017 | Exp. 2.2 29 Nov. 2018 | Exp. 2.3 05 Dec. 2018 |
| T10 | T10 | ||||
| T21 | T21 | T21 | T21 | T21 | T21 |
| T26 | T26 | T26 | T26 | T26 | T26 |
| T28 | T28 | T28 | T28 | T28 | T28 |
| T33 | T33 | T33 | T33 | ||
| T38 | T38 | T38 | T38 | ||
| Concept & (n) | DTA (days) | DT (cm) | SY (kg/ha) | |||
|---|---|---|---|---|---|---|
| 0% | 10% | 50% | 90% | |||
| Sowing dates | ||||||
| Mean Exp. 1.1 (3) | 83 | 84 | 86 | 88 | 204 | 8001 |
| Mean Exp. 2.3 (9) | 79 | 79 | 81 | 83 | 190 | 7206 |
| Difference Exp. 1.1 vs. 2.3 | 4 * | 5 * | 5 * | 5 * | 14 * | 795 * |
| Plant densities | ||||||
| D1 82,667 pt/ha (3) | 78 | 79 | 81 | 83 | 186 | 7011 |
| D2 93,333 pt/ha (3) | 78 | 79 | 81 | 83 | 194 | 7452 |
| D3 101,333 pt/ha (3) | 80 | 80 | 82 | 84 | 190 | 7158 |
| D1 vs. D2 | 0 | 0 | 0 | 0 | 8 | 441 |
| D1 vs. D3 | 2 * | 1 | 1 | 1 | 4 | 147 |
| D2 vs. D3 | 2 * | 1 | 1 | 1 | 4 | 294 |
| Male line | ||||||
| Averages T10 (12) | 80 | 81 | 83 | 85 | 193 | 7407 |
| Source | df | DTS (Days) | DS (cm) | ESY (kg/ha) | |||
|---|---|---|---|---|---|---|---|
| 0% | 10% | 50% | 90% | ||||
| Dates | 2 | 110.71 ** | 142.13 ** | 144.18 ** | 172.76 ** | 56.46 ns | 1125080.04 * |
| Lines | 4 | 8.91 ns | 10.54 * | 9.15 ns | 7.94 ns | 514.02 * | 2344102.13 ** |
| Dates × lines | 7 | 2.42 ns | 2.53 ns | 3.85 * | 4,42 ** | 65.99 ns | 122992.65 ns |
| CV (%) | 1.53 | 1.44 | 1.43 | 1.23 | 7.23 | 7.82 | |
| Source | df | DTS (days) | DS (cm) | ESY (kg/ha) | |||
|---|---|---|---|---|---|---|---|
| 0% | 10% | 50% | 90% | ||||
| Sowing dates (SD) | 2 | 241.37 ** | 206.22 ** | 205.00 ** | 194.00 ** | 4173.49 * | 667064.46 * |
| Lines (L) | 4 | 21.65 * | 32.14 ** | 27.80 * | 23.67 * | 1200.31 ns | 7507367.3 ** |
| Plant densities (PD) | 2 | 24.87 ** | 24.19 ** | 30.06 ** | 32.66 ** | 39.41 ns | 468768.26 * |
| PD × L | 8 | 1.31 ns | 1.04 ns | 1.32 ns | 2,80 * | 31.17 ns | 164074.32 ns |
| SD × L | 5 | 2.47 ns | 3.10 * | 3.63 ** | 3.30 * | 509.33 ** | 447710.28 ** |
| SD × PD | 4 | 25.15 ** | 25.87 ** | 26.19 ** | 25.84 ** | 191.89 ** | 199473.09 ns |
| SD × PD × L | 10 | 1.99 ns | 1.63 ns | 1.56 ns | 1.63 ns | 55.58 ns | 177789.01 ns |
| CV (%) | 1.36 | 1.31 | 1.30 | 1.34 | 7.25 | 8.29 | |
| Concept and (n) | DTS (days) | DS (cm) | ESY (kg/ha) | |||
|---|---|---|---|---|---|---|
| 0% | 10% | 50% | 90% | |||
| Sowing dates | ||||||
| Exp. 1.1; 23 Nov. 2017 (15) | 86 a | 87 a | 89 a | 92 a | 93 | 3783 a |
| Exp. 1.2; 29 Nov. 2018 (15) | 80 c | 81 b | 82 b | 84 b | 92 | 3614 a |
| Exp. 1.3; 05 Dec. 2018 (15) | 82 b | 82 b | 84 b | 86 b | 89 | 3158 b |
| Female lines | ||||||
| T21 (9) | 82 | 83 b | 85 | 87 | 105 a | 3994 a |
| T26 (9) | 82 | 83 b | 84 | 87 | 86 b | 4009 a |
| T28 (9) | 82 | 83 b | 84 | 87 | 87 b | 3499 b |
| T33 (9) | 84 | 85 a | 87 | 89 | 89 b | 2845 c |
| T38 (6) | 81 | 81 b | 83 | 85 | 89 b | 2976 c |
| Concept and (n) | DTS (days) | DS (cm) | ESY (kg/ha) | |||
|---|---|---|---|---|---|---|
| 0% | 10% | 50% | 90% | |||
| Sowing dates | ||||||
| Exp. 2.1; 16 Dec. 2017 (36) | 79 b | 80 b | 82 b | 84 b | 110 a | 3877 a |
| Exp. 2.2; 29 Nov. 2018 (36) | 78 b | 79 c | 81 c | 83 c | 90 b | 3936 a |
| Exp. 2.3; 05 Dec. 2018 (36) | 83 a | 83 a | 85 a | 87 a | 88 b | 3727 b |
| Plant densities | ||||||
| 82,667 pt/ha (39) | 80 a | 81 a | 83 b | 85 b | 96 | 3699 a |
| 93.333 pt/ha (39) | 79 b | 80 b | 82 c | 84 c | 95 | 3911 a |
| 101333 pt/ha (39) | 81 a | 82 a | 83 a | 86 a | 97 | 3929 a |
| Female lines | ||||||
| T21 (27) | 80 b | 81 b | 82 b | 84 b | 107 | 4345 a |
| T26 (27) | 80 ab | 81 b | 82 b | 85 b | 92 | 4198 a |
| T28 (27) | 80 b | 80 b | 82 b | 84 b | 91 | 3832 b |
| T33 (9) | 82 a | 83 a | 85 a | 87 a | 107 | 2786 d |
| T38 (18) | 80 ab | 81 b | 83 ab | 85 ab | 86 | 3125 c |
| Experiment/Plant Density (plants/ha) | T21 | T26 | T28 | T33 | T38 |
|---|---|---|---|---|---|
| Exp. 1.1: 71,111 | 97 | 121 | 118 | 109 | |
| Exp. 2.3: 82,667 | 93 | 100 | 101 | 100 | |
| Exp. 2.3: 93,333 | 100 | 109 | 103 | 104 | |
| Exp. 2.3: 101,333 | 101 | 110 | 98 | 107 | |
| Mean | 98 | 110 | 105 | 109 | 103 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chassaigne-Ricciulli, A.A.; Mendoza-Onofre, L.E.; Córdova-Téllez, L.; Carballo-Carballo, A.; San Vicente-García, F.M.; Dhliwayo, T. Development of Seed Production Technology of CIMMYT Tropical Single Cross Maize Hybrids. Agriculture 2020, 10, 259. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10070259
Chassaigne-Ricciulli AA, Mendoza-Onofre LE, Córdova-Téllez L, Carballo-Carballo A, San Vicente-García FM, Dhliwayo T. Development of Seed Production Technology of CIMMYT Tropical Single Cross Maize Hybrids. Agriculture. 2020; 10(7):259. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10070259
Chicago/Turabian StyleChassaigne-Ricciulli, Alberto A., Leopoldo E. Mendoza-Onofre, Leobigildo Córdova-Téllez, Aquiles Carballo-Carballo, Félix M. San Vicente-García, and Thanda Dhliwayo. 2020. "Development of Seed Production Technology of CIMMYT Tropical Single Cross Maize Hybrids" Agriculture 10, no. 7: 259. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10070259