Systemic Acquired Resistance Induced by Compatible and Incompatible Tomato Mosaic Viruses Effectively Controls Bacterial Spot and Speck Diseases in Tomato



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
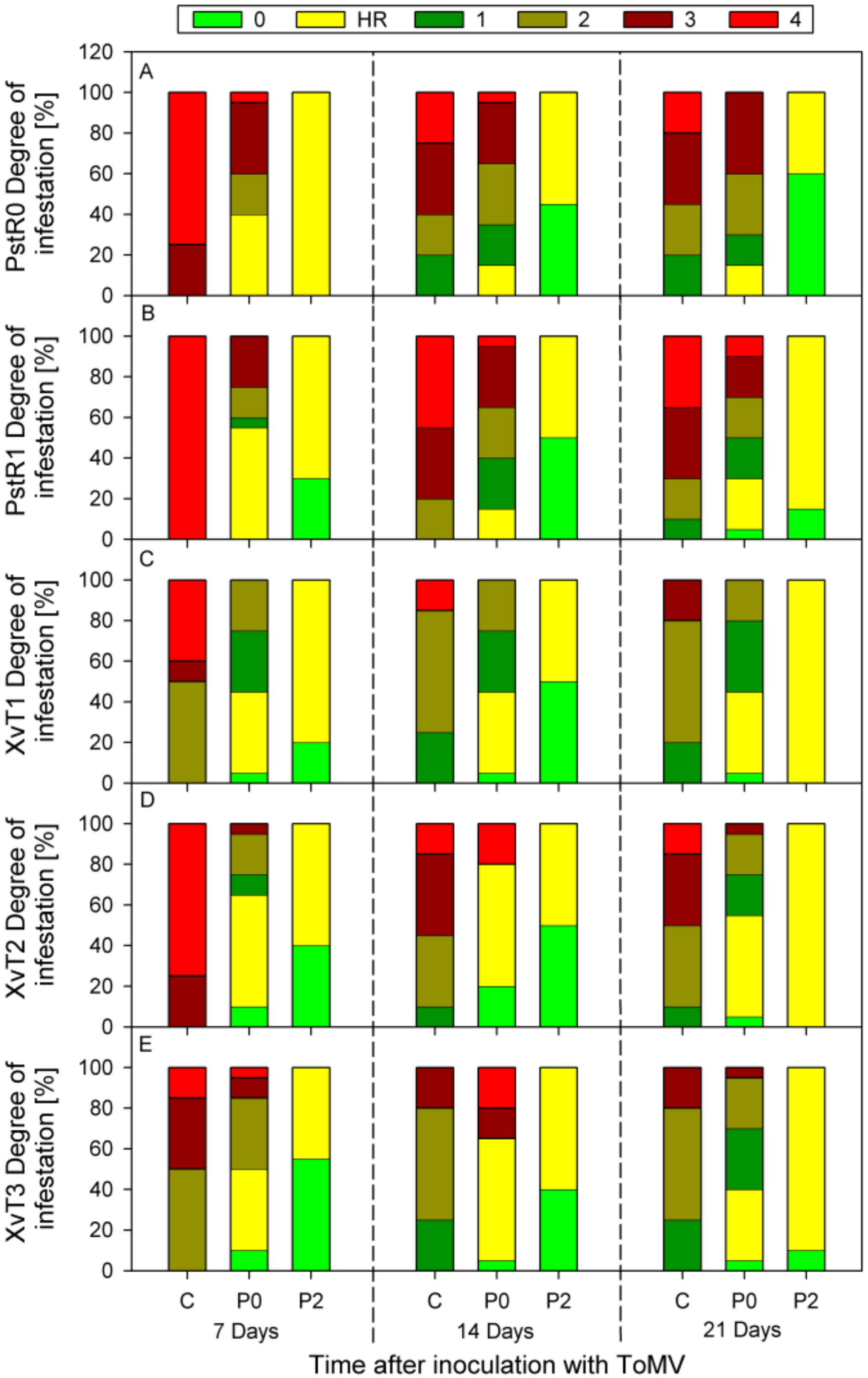
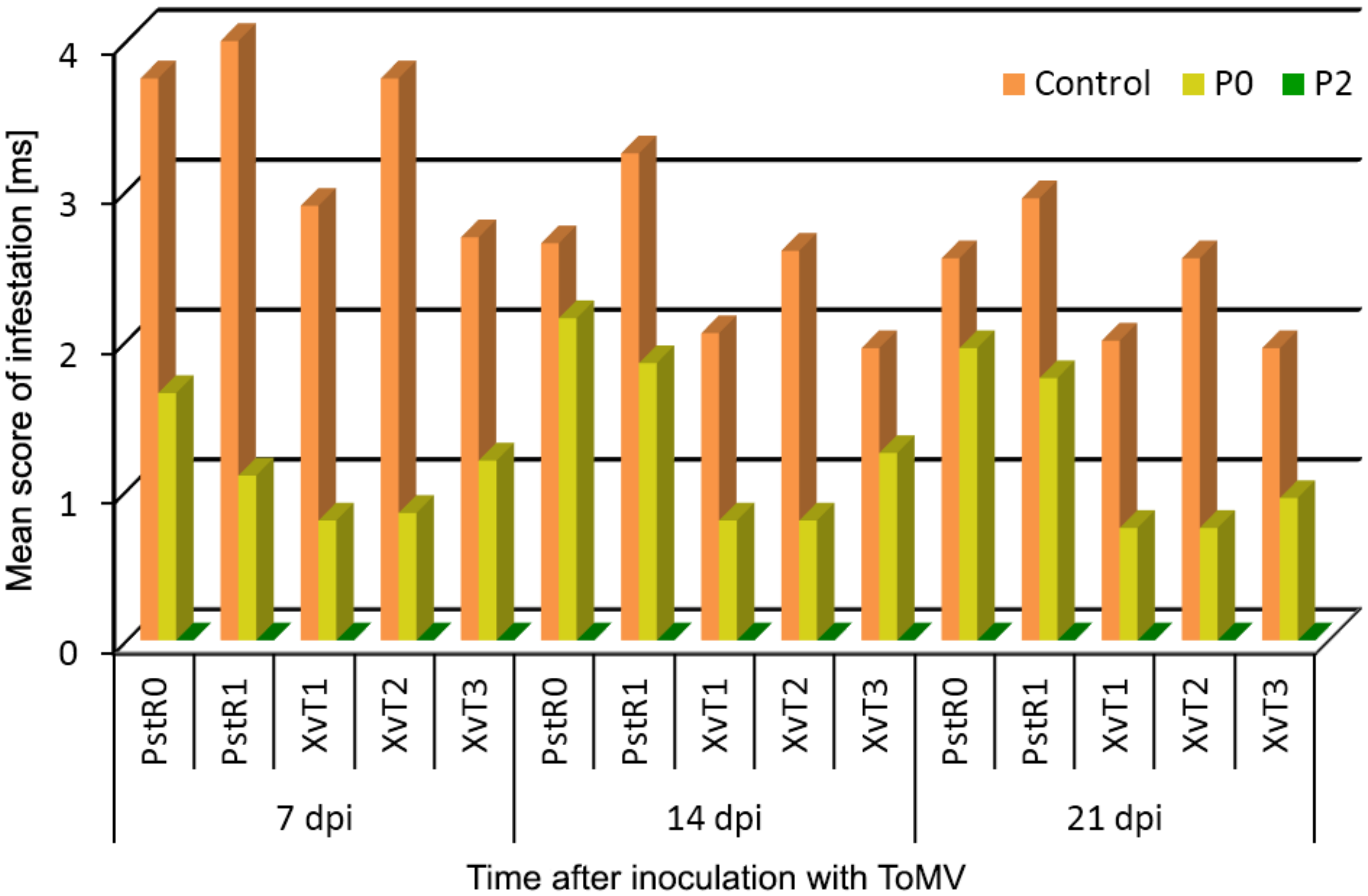
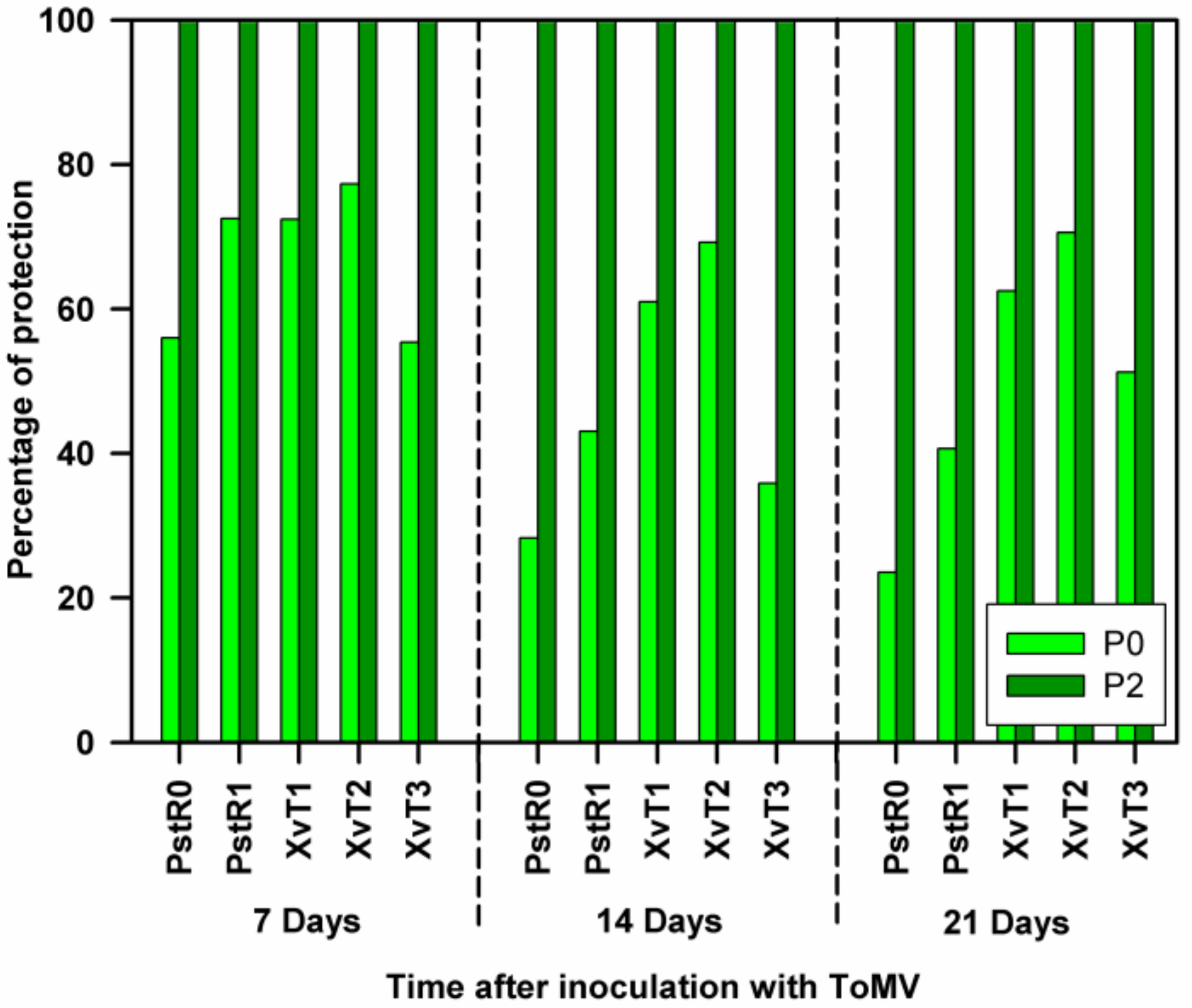
3.1. Assessment of Disease Severity and SAR Effectiveness in Tomato Plants Pre-Inoculated with Compatible and Incompatible ToMV Strains and Challenge Inoculated with Bacterial Strains
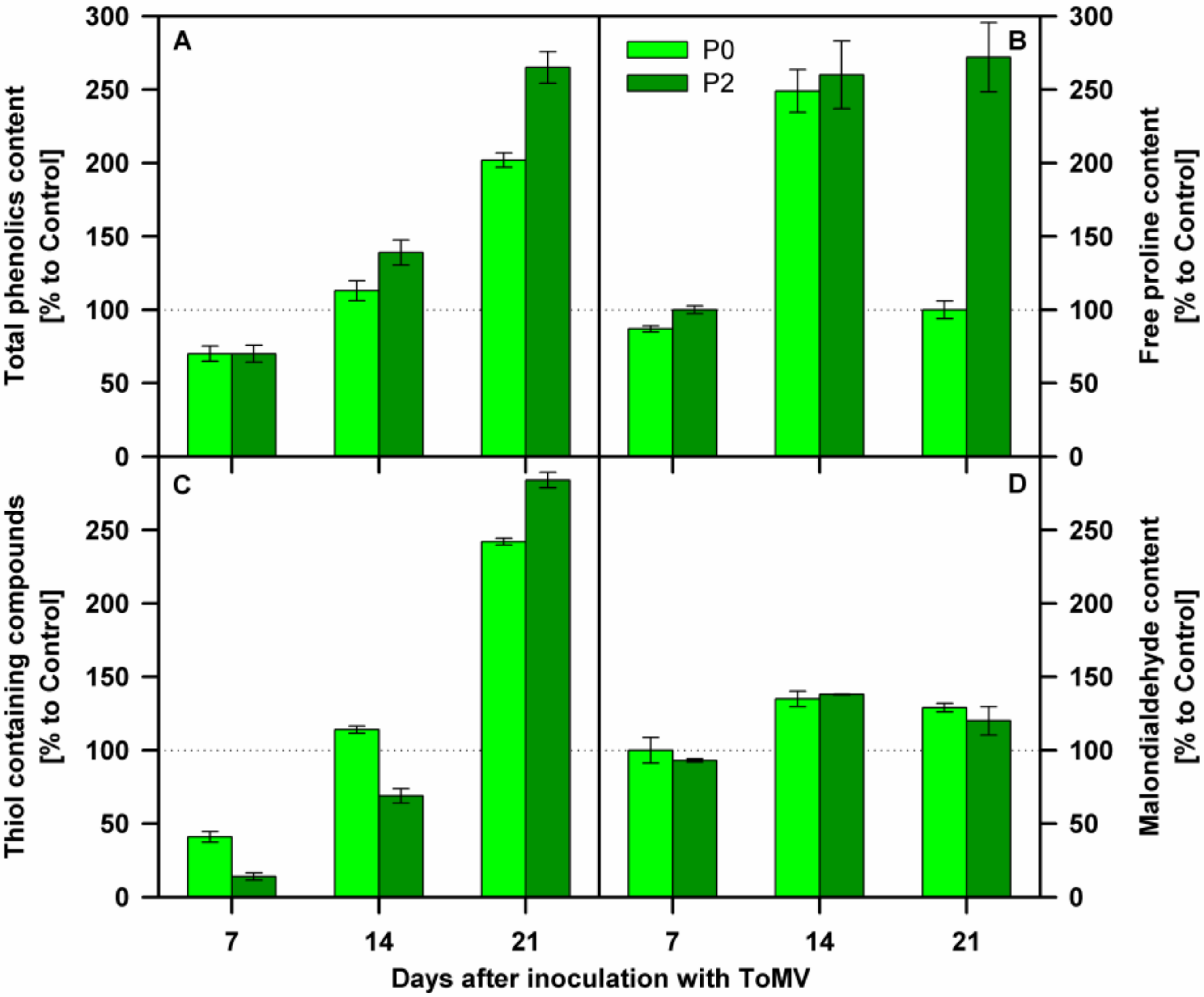
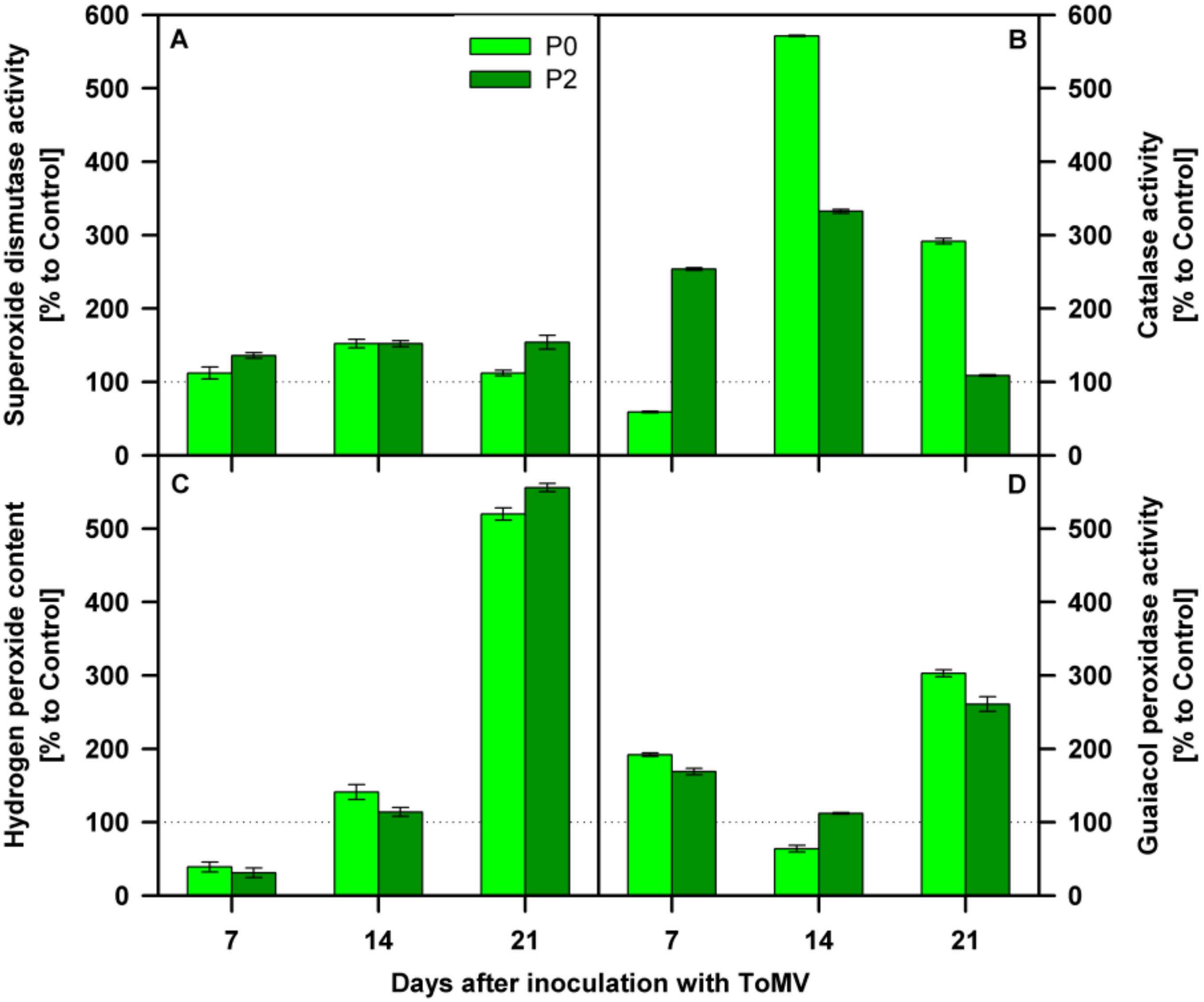
3.2. Biochemical Analyses of Tomato Plants Inoculated with Compatible and Incompatible ToMV Strains
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- OECD. Chapter 2. Tomato (Solanum lycopersicum). In Safety Assessment of Transgenic Organisms in the Environment; OECD Consensus Documents, Harmonisation of Regulatory Oversight in Biotechnology; OECD Publishing: Paris, France, 2017; Volume 7, pp. 69–104. [Google Scholar] [CrossRef]
- Ullah, N.; Ali, A.; Ahmad, M.; Fahim, M.; Din, N.; Ahmad, F. Evaluation of tomato genotypes against tomato mosaic virus (ToMV) and its effect on yield contributing parameters. Pak. J. Bot. 2017, 49, 1585–1592. [Google Scholar]
- Irfan, M.; Hayat, S.; Ahmad, A. Oxidative stress management in plants. Regulation of antioxidant system under biotic stress in plants. Chapter 14. In Antioxidants; Parvaz, A., Umar, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 283–303. [Google Scholar]
- Shetty, N.P.; Jorgensen, H.J.L.; Jensen, J.; Collinge, D.; Shetty, H.S. Roles of reactive oxygen species in interactions between plants and pathogens. Eur. J. Plant Pathol. 2008, 121, 267–280. [Google Scholar] [CrossRef]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as key players in plant stress signalling. J. Exp. Bot. 2014, 65, 1229–1240. [Google Scholar] [CrossRef] [PubMed]
- Hernández, J.A.; Gullner, G.; Clemente-Moreno, M.J.; Künstler, A.; Juhász, C.; Díaz-Vivancos, P.; Király, L. Oxidative stress and antioxidative responses in plant-virus interactions. Physiol. Mol. Plant Pathol. 2016, 94, 134–148. [Google Scholar] [CrossRef] [Green Version]
- Luna, E.; Bruce, T.J.A.; Roberts, M.R.; Flors, V.; Ton, J. Next-generation systemic acquired resistance. Plant Physiol. 2012, 158, 844–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryals, J.A.; Neuenschwander, U.H.; Willits, M.G.; Molina, A.; Steiner, H.-Y.; Hunt, M.D. Systemic acquired resistance. Plant Cell 1996, 8, 1809–1819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Király, L.; Barna, B.; Király, Z. Plant resistance to pathogen infection: Forms and mechanisms of innate and acquired resistance. J. Phytopathol. 2007, 155, 385–396. [Google Scholar] [CrossRef]
- Pfitzner, A.J.P. Resistance to tobacco mosaic virus and tomatomosaic virus in tomato. Chapter B4. In Natural Resistance Mechanisms of Plants to Viruses; Loebenstein, G., Carr, J.P., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 399–413. [Google Scholar]
- Stoimenova, E.; Bogatzevska, N. Tomato mosaic virus induced systemic resistance against bacterial spot and speck diseases in tomato. Plant Prot. 2006, 17, 109–115. [Google Scholar]
- Stoimenova, E.; Bogatzevska, N. Tomato mosaic virus induced systemic resistance against single and mix bacterial infection of Pseudomonas syringae pv. tomato and Xanthomonas vesicatoria on tomato. Plant Prot. 2008, 19, 70–73. [Google Scholar]
- Stoimenova, E.; Bogatzevska, N. Systemic acquired resistance induced by salicylic acid and tomato mosaic virus against bacterial spot and speck diseases in tomato. Plant Prot. 2009, 20, 100–104. [Google Scholar]
- Anfoka, G.; Buchenauer, H. Systemic acquired resistance in tomato against Phytophthora infestans by pre-inoculation with tobacco necrosis virus. Physiol. Mol. Plant. Pathol. 1997, 50, 85–101. [Google Scholar] [CrossRef]
- Kang, B.C.; Yeam, I.; Jahn, M.M. Genetics of plant virus resistance. Ann. Rev. Phytopathol. 2005, 43, 581–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conrath, U. Systemic acquired resistance. Plant Signal. Behav. 2006, 1, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Chambers, S.; Merriman, P. Perennation and control of Pseudomonas tomato in Victoria. Austr. J. Agric. Res. 1975, 26, 657–663. [Google Scholar] [CrossRef]
- Sotirova, V.; Beleva, L. Resistance of wild species, varieties and cultivars to Xanthomonas vesicatoria (Doidge) Dowson. C. R. Acad. Agric. 1975, 8, 43–47. [Google Scholar]
- Bates, L. Rapid determination of free proline content for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Swain, T.; Goldstein, L. Methods in Polyphenol Chemistry; Pridham, J.B., Ed.; Pergamon Press: Oxford, UK, 1964; pp. 131–146. [Google Scholar]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–75. [Google Scholar] [CrossRef]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Kramer, G.; Norman, H.; Krizek, D.; Mirecki, R. Influence of UV-B radiation on polyamines, lipid peroxidation and membrane lipids in cucumber. Phytochemistry 1991, 30, 2101–2108. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase. Improved assay and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Dias, I.; Costa, M. Effect of low salt concentration on nitrate reductase and peroxidase of sugar beet leaves. J. Exp. Bot. 1983, 34, 197–202. [Google Scholar] [CrossRef]
- Aebi, M. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Meshi, T.; Motoyoshi, F.; Maeda, T.; Yoshiwoka, S.; Watanabe, H.; Okada, Y. Mutations in the tobacco mosaic virus 30-kD protein gene overcome Tm-2 resistance in tomato. Plant Cell 1989, 1, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Strasser, M.; Pfitzner, A.J. The double-resistance-breaking Tomato mosaic virus strain ToMV1-2 contains two independent single resistance-breaking domains. Arch. Virol. 2007, 152, 903–914. [Google Scholar] [CrossRef]
- Zitter, T. Tobacco Mosaic Virus (TMV) Control; Cornell University: Ithaca, NY, USA, 2012; Available online: https://cuaes.cals.cornell.edu/sites/cuaes.cals.cornell.edu/files/shared/documents/TMV_ToMV_CALS_TomZitter.pdf (accessed on 13 July 2020).
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Gholi-Tolouie, S.; Sokhandan-Bashir, N.; Davari, M.; Sedghi, M. The effect of salicylic and jasmonic acids on tomato physiology and tolerance to Cucumber mosaic virus (CMV). Eur. J. Plant Pathol. 2018, 151, 101–116. [Google Scholar] [CrossRef]
- Ramos, O.F.; Smith, C.M.; Fritz, A.K.; Madl, R.L. Salicylic acid-mediated synthetic elicitors of systemic acquired resistance administered to wheat plants at jointing stage induced phenolics in mature grains. Crop Sci. 2017, 57, 3122–3128. [Google Scholar] [CrossRef]
- Anuradha, C.; Selvarajan, R.; Vasantha, S.; Suresha, G.S. Biochemical characterization of compatible plant virus interaction: A case study with bunchy top virus-banana host-pathosystem. Plant Pathol. J. 2015, 14, 212–222. [Google Scholar] [CrossRef] [Green Version]
- Balogun, O.S.; Teraoka, T. Time-course analysis of the accumulation of phenols in tomato seedlings infected with Potato Virus X and Tobacco mosaic virus. Biokemistri 2004, 16, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Mahjabeen; Akhtar, K.P.; Sarwar, N.; Saleem, M.Y.; Asghar, M.; Iqbal, Q.; Jamil, F.F. Effect of cucumber mosaic virus infection on morphology, yield and phenolic contents of tomato. Arch. Phytopathol. Plant Prot. 2012, 45, 766–782. [Google Scholar] [CrossRef]
- Hassan, O.; Chang, T.; Hossain, A. Changes in the secondary compounds of persimmon leaves as a defense against circular leaf spot caused by Plurivorosphaerella nawae. PloS ONE 2020, 15, e0230286. [Google Scholar] [CrossRef] [PubMed]
- Kuzniak, E. The ascorbate-glutathione cycle and related redox signals in plant-pathogen interactions. In Ascorbate-Glutahione Pathway and Stress Tolerance in Plants; Anjum, N.A., Umar, S., Chan, M.-T., Eds.; Springer: Dordrecht, Germany, 2010; pp. 115–136. [Google Scholar]
- Gullner, G.; Zechmann, B.; Künstler, A.; Király, L. The signaling roles of glutathione in plant disease resistance. Chapter 15. In Glutathione in Plant Growth, Development, and Stress Tolerance; Hossain, M.A., Mostofa, M.G., Diaz-Vivancos, P., Burritt, D.J., Fujita, M., Tran, L.-S.P., Eds.; Springer: Cham, Switzerland, 2017; pp. 331–337. ISBN 978-3-319-66682-2. [Google Scholar]
- Künstler, A.; Király, L.; Kátay, G.; Enyedi, A.J.; Gullner, G. Glutathione can compensate for salicylic acid defciency in tobacco to maintain resistance to Tobacco Mosaic Virus. Front. Plant Sci. 2019, 10, 1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagai, A.; Torres, P.B.; Duarte, L.M.L.; Chaves, A.L.R.; Macedo, A.F.; Floh, E.I.S.; de Oliveira, L.F.; Zuccarelli, R.; dos Santos, D.Y.A.C. Signaling pathway played by salicylic acid, gentisic acid, nitric oxide, polyamines and non-enzymatic antioxidants in compatible and incompatible Solanum-tomato mottle mosaic virus interactions. Plant Sci. 2020, 290, 110274. [Google Scholar] [CrossRef] [PubMed]
- Fabro, G.; Kovács, I.; Pavet, V.; Szabados, L.; Alvarez, M.E. Proline accumulation and AtP5CS2 gene activation are induced by plant-pathogen incompatible interactions in Arabidopsis. Mol. Plant Microbe Interact. 2004, 17, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Gholi-Tolouie, S.; Davari, M.; Sokhandan-Bashir, N.; Sedghi, M. Influence of salicylic and jasmonic acids on the antioxidant systems of tomato (Solanum lycopersicum cv. Superchief) plants under biotic stresses’. Iran. J. Plant Physiol. 2018, 8, 2345–2351. [Google Scholar]
- Jiang, J.-F.; Li, J.-G.; Dong, Y.-H. Effect of calcium nutrition on resistance of tomato against bacterial wilt induced by Ralstonia solanacearum. Eur. J. Plant Pathol. 2013, 136, 547–555. [Google Scholar] [CrossRef]
- Almagro, L.; Gomez Ros, L.V.; Belchi-Navarro, S.; Bru, R.; Ros Barcelo, A.; Pedreno, M.A. Class III peroxidases in plant defence reactions. J. Exp. Bot. 2009, 60, 377–390. [Google Scholar] [CrossRef] [Green Version]
- Chassot, C.; Buchala, A.; Schoonbeek, H.-J.; Metraux, J.-P.; Lamotte, O. Wounding of Arabidopsis leaves causes a powerful but transient protection against Botrytis infection. Plant J. 2008, 55, 555–567. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shopova, E.; Mihailova, B.; Todorova, D.; Sergiev, I.; Stoimenova, E. Systemic Acquired Resistance Induced by Compatible and Incompatible Tomato Mosaic Viruses Effectively Controls Bacterial Spot and Speck Diseases in Tomato. Agriculture 2020, 10, 302. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10070302
Shopova E, Mihailova B, Todorova D, Sergiev I, Stoimenova E. Systemic Acquired Resistance Induced by Compatible and Incompatible Tomato Mosaic Viruses Effectively Controls Bacterial Spot and Speck Diseases in Tomato. Agriculture. 2020; 10(7):302. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10070302
Chicago/Turabian StyleShopova, Elena, Bistra Mihailova, Dessislava Todorova, Iskren Sergiev, and Elisaveta Stoimenova. 2020. "Systemic Acquired Resistance Induced by Compatible and Incompatible Tomato Mosaic Viruses Effectively Controls Bacterial Spot and Speck Diseases in Tomato" Agriculture 10, no. 7: 302. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture10070302