Earthworm Inoculation Improves Upland Rice Crop Yield and Other Agrosystem Services in Madagascar


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
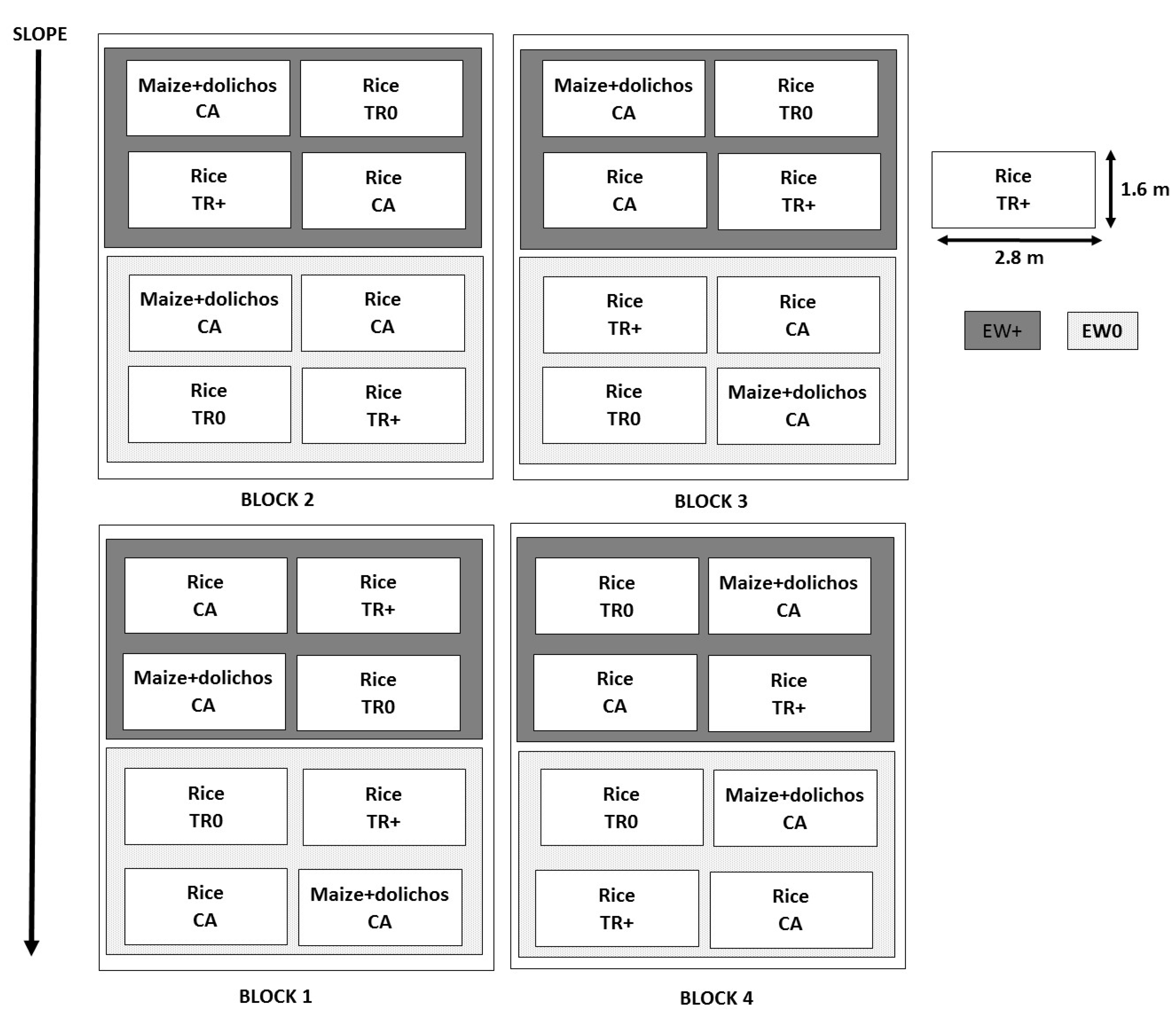
2.2. Experimental Design and Plot Preparation
2.3. Earthworm Inoculation
2.4. Soil and Plant Analyses
2.5. Statistical Analyses
3. Results
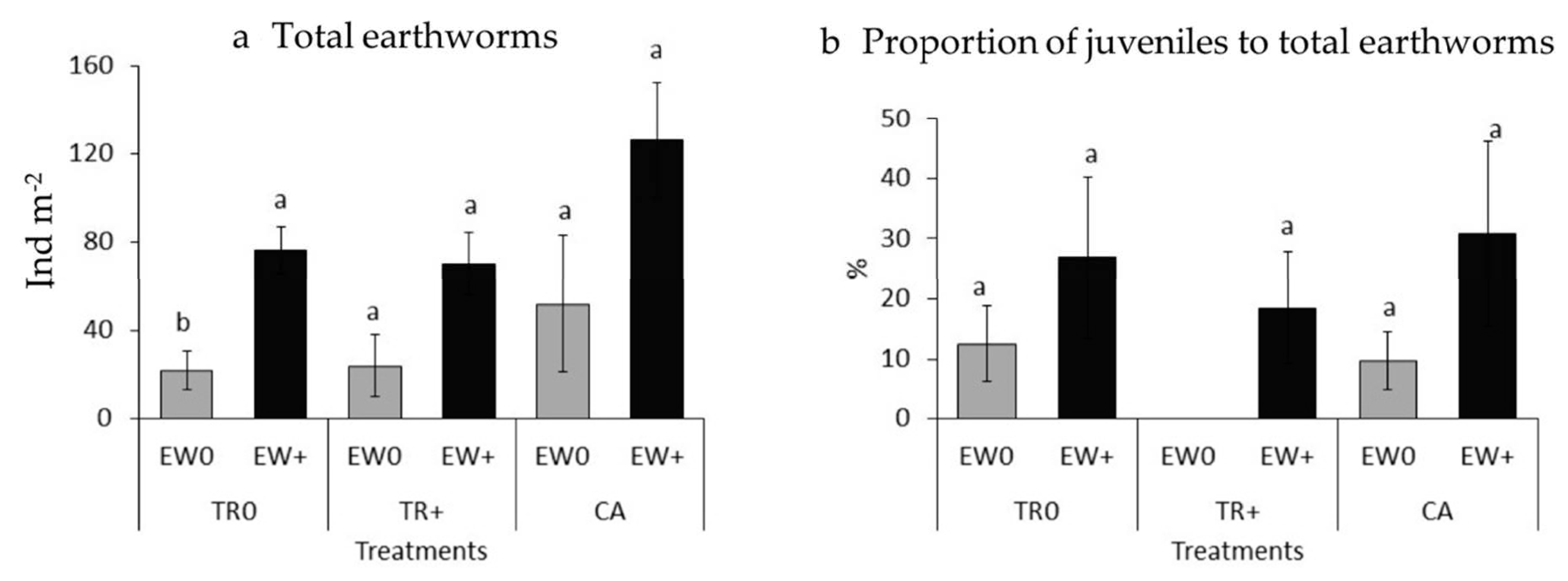
3.1. Soil Biological Properties
3.2. Soil Physico-Chemical Properties
3.2.1. Soil Properties at the Beginning of the Experiment
3.2.2. Soil Properties at the End of the 3rd Cropping Season
3.2.3. Changes in Soil Properties with Time
3.3. Plant Parameters
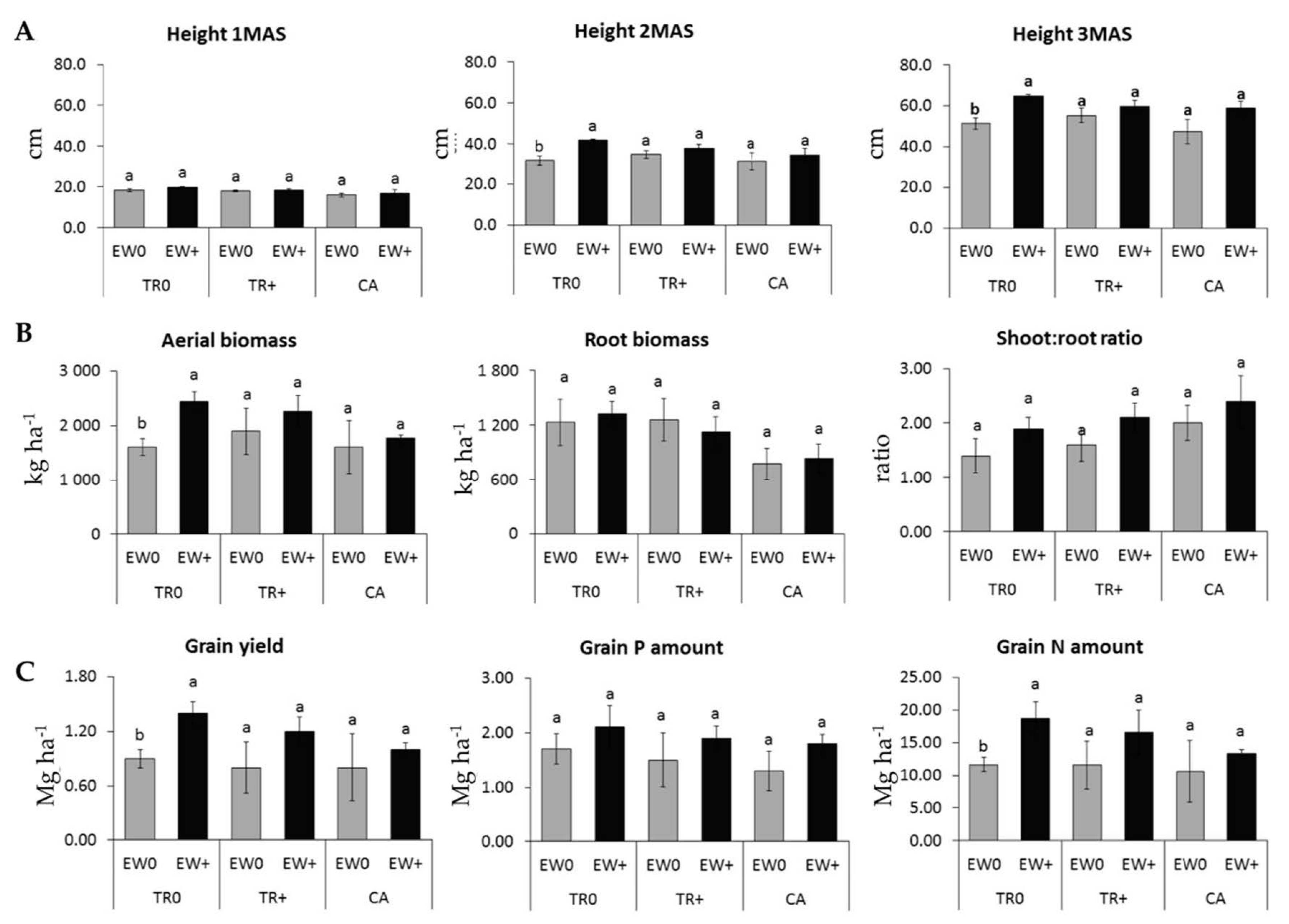
3.3.1. Rice Growth at the End of the 3rd Cropping Season
3.3.2. Rice Grain Yield and Nutrient Acquisition at the end of the 3rd Cropping Season
4. Discussion
4.1. Success of Earthworm Inoculation
4.2. Effects on Soil and Plant Properties
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, W.; Hendrix, P.F.; Dame, L.E.; Burke, R.A.; Wu, J.; Neher, D.A.; Li, J.; Shao, Y.; Fu, S. Earthworms Facilitate Carbon Sequestration through Unequal Amplification of Carbon Stabilization Compared with Mineralization. Nat. Commun. 2013, 4, 2576. [Google Scholar] [CrossRef] [PubMed]
- Van Groenigen, J.W.; Van Groenigen, K.J.; Koopmans, G.F.; Stokkermans, L.; Vos, H.M.; Lubbers, I.M. How Fertile Are Earthworm Casts? a Meta-Analysis. Geoderma 2019, 338, 525–535. [Google Scholar] [CrossRef]
- Bityutskii, N.P.; Kaidun, P.; Yakkonen, K. Earthworms Can Increase Mobility and Bioavailability of Silicon in Soil. Soil Biol. Biochem. 2016, 99, 47–53. [Google Scholar] [CrossRef]
- Blanchart, E.; Albrecht, A.; Brown, G.G.; Decaëns, T.; Duboisset, A.; Lavelle, P.; Mariani, L.; Roose, E. Effects of Tropical en-Dogeic Earthworms on Soil Erosion: A Review. Agric. Ecosyst. Environ. 2004, 104, 303–315. [Google Scholar] [CrossRef]
- Blanchart, E.; Marilleau, N.; Chotte, J.-L.; Drogoul, A.; Perrier, E.; Cambier, C. SWORM: An Agent-Based Model to Simulate the Effect of Earthworms on Soil Structure. Eur. J. Soil Sci. 2008, 60, 13–21. [Google Scholar] [CrossRef]
- Bertrand, M.; Blouin, M.; Barot, S.; Charlier, A.; Marchand, D.; Roger-Estrade, J. Biocontrol of Eyespot Disease on Two Winter Wheat Cultivars by an Anecic Earthworm (Lumbricus terrestris). Appl. Soil Ecol. 2015, 96, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Blanchart, E.; Ratsiatosika, O.; Raveloson, H.; Razafimbelo, T.; Razafindrakoto, M.; Sester, M.; Becquer, T.; Bernard, L.; Trap, J. Nitrogen Supply Reduces the Earthworm-Silicon Control on Rice Blast Disease in a Ferralsol. Appl. Soil Ecol. 2020, 145, 103341. [Google Scholar] [CrossRef]
- Bernard, L.; Chapuis-Lardy, L.; Razafimbelo, T.; Razafindrakoto, M.; Pablo, A.-L.; Legname, E.; Poulain, J.; Brüls, T.; O’Donohue, M.; Brauman, A.; et al. Endogeic Earthworms Shape Bacterial Functional Communities and Affect Organic Matter Mineralization in a Tropical Soil. ISME J. 2011, 6, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Butenschoen, O.; Poll, C.; Langel, R.; Kandeler, E.; Marhan, S.; Scheu, S. Endogeic Earthworms Alter Carbon Translocation by Fungi at the Soil–Litter Interface. Soil Biol. Biochem. 2007, 39, 2854–2864. [Google Scholar] [CrossRef]
- Villenave, C.; Rabary, B.; Kichenin, E.; Djigal, D.; Blanchart, E. Earthworms and Plant Residues Modify Nematodes in Tropical Cropping Soils (Madagascar): A Mesocosm Experiment. Appl. Environ. Soil Sci. 2010, 2010, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Eisenhauer, N. The Action of an Animal Ecosystem Engineer: Identification of the Main Mechanisms of Earthworm Impacts on Soil Microar-Thropods. Pedobiologia 2010, 53, 343–352. [Google Scholar] [CrossRef]
- Van Groenigen, J.W.; Lubbers, I.M.; Vos, H.M.J.; Brown, G.G.; De Deyn, G.B.; Van Groenigen, K.J. Earthworms Increase Plant Production: A Meta-Analysis. Sci. Rep. 2015, 4, 6365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavelle, P.; Barois, I.; Martin, A.; Zaidi, Z.; Schaefer, R. Management of Earthworm Populations in Agro-Ecosystems: A Possible Way to Maintain Soil Quality? In Ecology of Arable Land—Perspectives and Challenges; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 1989; pp. 109–122. [Google Scholar]
- Jouquet, P.; Blanchart, E.; Capowiez, Y. Utilization of Earthworms and Termites for the Restoration of Ecosystem Functioning. Appl. Soil Ecol. 2014, 73, 34–40. [Google Scholar] [CrossRef]
- Wubs, E.R.J.; Van Der Putten, W.H.; Bosch, M.; Bezemer, T.M. Soil Inoculation Steers Restoration of Terrestrial Ecosystems. Nat. Plants 2016, 2, 16107. [Google Scholar] [CrossRef] [Green Version]
- Brun, J.J.; Cluzeau, D.; Tréhen, P.; Bouché, M.B. Biostimulation: Perspectives et Limites de L’Amélioration Biologique Des Sols par Stimulation ou Introduction d’Espèces Lombriciennes. Rev. Ecol. Biol. Sols 1987, 24, 685–701. [Google Scholar]
- Butt, K.R. Earthworms in Soil Restoration: Lessons Learned from United Kingdom Case Studies of Land Reclamation. Restor. Ecol. 2008, 16, 637–641. [Google Scholar] [CrossRef]
- Butt, K.R.; Frederickson, J.; Morris, R.M. The Earthworm Inoculation Unit Technique: An Integrated System for Cultivation and Soil-Inoculation of Earthworms. Soil Biol. Biochem. 1997, 29, 251–257. [Google Scholar] [CrossRef]
- Lavelle, P.; Barros, E.; Blanchart, E.; Brown, G.G.; Desjardins, T.; Mariani, L.; Rossi, J.-P. SOM Management in the Tropics: Why Feeding the Soil Macrofauna? Nutr. Cycl. Agroecosyst. 2001, 61, 53–61. [Google Scholar] [CrossRef]
- Butt, K.R. Inoculation of Earthworms into Reclaimed Soils: The UK Experience. Land Degrad. Dev. 1999, 10, 565–575. [Google Scholar] [CrossRef]
- Pashanasi, B.; Lavelle, P.; Alegre, J.; Charpentier, F. Effect of the Endogeic Earthworm Pontoscolex Corethrurus on Soil Chemical Characteristics and Plant Growth in a Low-Input Tropical Agroecosystem. Soil Biol. Biochem. 1996, 28, 801–810. [Google Scholar]
- Senapati, B.; Lavelle, P.; Giri, S.; Pashanasi, B.; Alegre, J.; Decaëns, T.; Jimenez, J.J.; Albrecht, A.; Blanchart, E.; Mahieu, M.; et al. In-Soil Earthworm Technologies for Tropical Agroecosystems. In Earthworm Management in Tropical Agroecosystems; Lavelle, P., Brussaard, L., Hendrix, P., Eds.; CABI Publishing: Wallingford, UK, 1999; pp. 199–238. [Google Scholar]
- Blanchart, E.; Albrecht, A.; Chevallier, T.; Hartmann, C. The Respective Roles of Biota (Roots and Earthworms) in the Restoration of Physical Properties in Vertisol under a Digitaria De-cumbens Pasture (Martinique). Agric. Ecosyst. Environ. 2004, 103, 343–355. [Google Scholar] [CrossRef]
- Senapati, B.K.; Lavelle, P.; Panigrahi, P.K.; Giri, S.; Brown, G.G. Restoring Soil Fertility and Enhancing Productivity in Indian Tea Plantations with Earthworms and Organic Fertilizers. In Proceedings of the International Technical Workshop on Biological Management of Soil Ecosystems for Sustainable Agriculture, Londrina, Brazil, 24–27 June 2002; FAO (Food and Agriculture Organization): Rome, Italy, 2003; Volume 1, pp. 172–190. [Google Scholar]
- Spain, A.; Lavelle, P.; Mariotti, A. Stimulation of Plant Growth by Tropical Earthworms. Soil Biol. Biochem. 1992, 24, 1629–1633. [Google Scholar] [CrossRef]
- Raminoarison, M.; Razafimbelo, T.; Rakotoson, T.; Becquer, T.; Blanchart, E.; Trap, J. Multiple-Nutrient Limitation of Upland Rainfed Rice in Ferralsols: A Greenhouse Nutrient-Omission Trial. J. Plant Nutr. 2019, 43, 270–284. [Google Scholar] [CrossRef]
- Razafindrakoto, M.; Csuzdi, C.S.; Blanchart, E. New and Little-Known Giant Earthworms from Madagascar (Oligochaeta: Ky-notidae). Afr. Invertebr. 2011, 52, 285–294. [Google Scholar] [CrossRef]
- Csuzdi, C.; Razafindrakoto, M.; Blanchart, E. New and Little-Known Earthworm Species from Central Madagascar (Oligo-chaeta: Kynotidae). Zootaxa 2012, 3578, 36–42. [Google Scholar] [CrossRef] [Green Version]
- Razafindrakoto, M.; Csuzdi, C.; James, S.; Blanchart, E. New Earthworms from Madagascar with Key to the Kynotus Species (Oligochaeta: Kynotidae). Zool. Anz. A J. Comp. Zool. 2017, 268, 126–135. [Google Scholar] [CrossRef]
- Hong, Y.; Razafindrakoto, M.; Blanchart, E.; Szederjesi, T.; Csuzdi, C. New Acanthodrilid Species from Madagascar (Clitellata, Acanthodrilidae). Acta Zool. Acad. Sci. Hung. 2019, 65, 215–233. [Google Scholar] [CrossRef]
- Rakotomanga, D.; Blanchart, E.; Rabary, B.; Randriamanantsoa, R.; Razafindrakoto, M.; Autfray, P. Diversité de la Macrofaune Des Sols Cultivés Sur Les Hautes Terres de Madagascar. Biotechnol. Agron. Soc. Environ. 2016, 20, 495–507. [Google Scholar]
- Taheri, S.; Pelosi, C.; Dupont, L. Harmful or Useful? A Case Study of the Exotic Peregrine Earthworm Morphospecies Pontoscolex Corethrurus. Soil Biol. Biochem. 2018, 116, 277–289. [Google Scholar] [CrossRef]
- Coq, S.; Barthès, B.G.; Oliver, R.; Rabary, B.; Blanchart, E. Earthworm Activity Affects Soil Aggregation and Organic Matter Dynamics According to the Quality and Localization of Crop Residues—an Experimental Study (Madagascar). Soil Biol. Biochem. 2007, 39, 2119–2128. [Google Scholar] [CrossRef]
- Trap, J.; Blanchart, E.; Ratsiatosika, O.; Razafindrakoto, M.; Becquer, T.; Andriamananjara, A.; Morel, C. Effects of the Earthworm Pontoscolex Corethrurus on Rice P Nutrition and Plant-Available Soil P in a Tropical Ferralsol. Appl. Soil Ecol. 2021, 160, 103867. [Google Scholar] [CrossRef]
- Fonte, S.J.; Quintero, D.C.; Velásquez, E.; Lavelle, P. Interactive Effects of Plants and Earthworms on the Physical Stabilization of Soil Organic Matter in Aggregates. Plant Soil 2012, 359, 205–214. [Google Scholar] [CrossRef]
- André, J.; Villenave, C.; Charpentier, F.; Bernier, N.; Lavelle, P.; Ponge, J.; Derouard, L.; Pashanasi, B.; Rossi, J.-P. Effects of Earthworms on Soil Organic Matter and Nutrient Dynamics at a Landscape Scale over Decades. In Earthworm Ecology; Informa UK Limited: London, UK, 2004; pp. 145–160. [Google Scholar]
- Hooper, D.J. Extraction of Free-Living Stages from Soil. In Laboratory Methods for Work with Plant and Soil Nematodes; Southey, J.F.E., Ed.; Her Majesty’s Stationary Office: London, UK, 1986; pp. 5–30. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Rabary, B. Impact du Semis Direct Sous Couverture Végétale Sur la Macrofaune ET la Microflore des Sols Ferrallitiques D’Andranomanelatra, Antsirabe, Hautes Terres Malgaches. Ph.D. Thesis, University of Antananarivo, Antananarivo, Madagascar, 2011; p. 211. [Google Scholar]
- Brown, G.G.; Pashanasi, B.; Villenave, C.; Patron, J.C.; Senapati, B.K.; Giri, S.; Barois, I.; Lavelle, P.; Blanchart, E.; Blakemore, R.J.; et al. Effects of Earthworms on Plant Production in the Tropics. in Earthworm Management. In Tropical Agroecosystems; Lavelle, P., Brussaard, L., Hendrix, P., Eds.; CABI Publishing: Wallingford, UK, 1999; pp. 87–148. [Google Scholar]
- Blanchart, E.; Bernoux, M.; Sarda, X.; Siqueira Neto, M.; Cerri, C.C.; Piccolo, M.; Douzet, J.M.; Scopel, E.; Feller, C. Effect of Direct Seeding Mulch-Based Systems on Soil Carbon Storage and Macrofauna in Central Brazil. Agric. Conspec. Sci. 2007, 72, 81–87. [Google Scholar]
- Eriksen-Hamel, N.S.; Speratti, A.B.; Wang, W.; Légère, A.; Madramootoo, C.A. Earthworm Populations and Growth Rates Related to Long-Term Crop Residue and Tillage Management. Soil Tillage Res. 2009, 104, 311–316. [Google Scholar] [CrossRef]
- Henneron, L.; Bernard, L.; Hedde, M.; Pelosi, C.; Villenave, C.; Chenu, C.; Bertrand, M.; Girardin, C.; Blanchart, E. Fourteen Years of Evidence for Positive Effects of Conservation Agriculture and Organic Farming on Soil Life. Agron. Sustain. Dev. 2014, 35, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Lavelle, P.; Spain, A.V.; Fonte, S.; Bedano, J.C.; Blanchart, E.; Galindo, V.; Grimaldi, M.; Jimenez, J.J.; Velasquez, E.; Zangerlé, A. Soil Aggregation, Ecosystem Engineers and the C Cycle. Acta Oecol. 2020, in press. [Google Scholar] [CrossRef]
- Brown, G.G.; Edwards, C.A.; Brussaard, L. How Earthworms Affect Plant Growth: Burrowing into the Mechanisms. In Earth-Worm Ecology, 2nd ed.; Edwards, C.A., Ed.; CRC Press: Boca Raton, FL, USA, 2004; Chapter 2; p. 441. [Google Scholar]
- Blanchart, E.; Albrecht, A.; Alegre, J.; Duboisset, A.; Villenave, C.; Pashanasi, B.; Lavelle, P.; Brussaard, L. Effects of Earthworms on Soil Structure and Physical Properties. In Earthworm Management in Tropical Agroecosystems; Lavelle, P., Brussaard, L., Eds.; CABI Publishing: Wallingford, UK, 1999; pp. 149–172. [Google Scholar]
- Muscolo, A.; Bovalo, F.; Gionfriddo, F.; Nardi, S. Earthworms Humic Matter Produces Auxin-Like Effects on Daucus Carota Cell Growth and Nitrate Metabolism. Soil Biol. Biochem. 1999, 31, 1303–1311. [Google Scholar] [CrossRef]
- Blouin, M. Chemical Communication: An Evidence for Co-evolution between Plants and Soil Organisms. Appl. Soil Ecol. 2018, 123, 409–415. [Google Scholar] [CrossRef]
- Brouwer, R. Functional Equilibrium: Sense or Nonsense? Neth. J. Agric. Sci. 1983, 31, 335–348. [Google Scholar] [CrossRef]
- Kreuzer, K.; Bonkowski, M.; Langel, R.; Scheu, S. Decomposer Animals (Lumbricidae, Collembola) and Organic Matter Distri-Bution Affect the Performance of Lolium Perenne (Poaceae) and Trifolium Repens (Fabaceae). Soil Biol. Biochem. 2004, 36, 2005–2011. [Google Scholar] [CrossRef]
- Coulis, M.; Bernard, L.; Gerard, F.; Hinsinger, P.; Claude, P.; Villeneuve, M.; Blanchart, E. Endogeic Earthworms Modify Soil Phosphorus, Plant Growth and Interactions in a Legume–Cereal Intercrop. Plant Soil 2014, 379, 149–160. [Google Scholar] [CrossRef]
- Razafimbelo, T.M.; Albrecht, A.; Oliver, R.; Chevallier, T.; Chapuis-Lardy, L.; Feller, C. Aggregate Associated-C and Physical Protection in a Tropical Clayey Soil under Malagasy Conventional and No-Tillage Systems. Soil Tillage Res. 2008, 98, 140–149. [Google Scholar] [CrossRef]
- Parr, J.; Papendick, R.; Hornick, S.; Meyer, R. Soil Quality: Attributes and Relationship to Alternative and Sustainable Agriculture. Am. J. Altern. Agric. 1992, 7, 5–11. [Google Scholar] [CrossRef]
- Altieri, M.A. Agroecology: The Science of Sustainable Agriculture; Westview Press: Boulder, CO, USA, 1995; p. 182. [Google Scholar]
- Ranaivoson, L.; Naudin, K.; Ripoche, A.; Affholder, F.; Rabeharisoa, R.L.; Corbeels, M. Agro-Ecological Functions of Crop Residues under Conservation Agriculture. A Review. Agron. Sustain. Dev. 2017, 37, 26. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Units | Treatments | One-Way ANOVA | ||
|---|---|---|---|---|
| EW0 | EW+ | Plot Position | ||
| Total C content | g kg−1 | 16.8 ± 0.6 | 14.3 ± 0.6 | 0.005 |
| Total N content | g kg−1 | 1.84 ± 0.09 | 1.56 ± 0.05 | <0.001 |
| Pi content | mg kg−1 | 10.65 ± 0.83 | 4.33 ± 0.55 | <0.001 |
| Total C stock | Mg ha−1 | 19.3 ± 0.8 | 16.7 ± 0.7 | 0.028 |
| Total N stock | Mg ha−1 | 2.11 ± 0.11 | 1.83 ± 0.08 | 0.011 |
| Pi stock | kg ha−1 | 12.1 ± 0.9 | 5.1 ± 0.7 | <0.001 |
| Bulk density | g cm−3 | 1.14 ± 0.04 | 1.18 ± 0.04 | 0.058 |
| Soil Variables | Units | Treatments | |||||
|---|---|---|---|---|---|---|---|
| TR0 | TR+ | CA | |||||
| EW0 | EW+ | EW0 | EW+ | EW0 | EW+ | ||
| Chemical soil variables Total C content | g kg−1 | 23.7 ± 1.1 | 22.6 ± 0.6 | 24.3 ± 1.0 | 22.7 ± 1.4 | 23.6 ± 0.6 | 22.2 ± 1.0 |
| NH4+ content | g kg−1 | 4.18 ± 0.83 | 4.60 ± 0.51 | 4.41 ± 0.43 | 4.14 ± 0.67 | 4.38 ± 0.54 | 5.07 ± 0.28 |
| Pi content | mg kg−1 | 11.86 ± 1.90 | 5.70 ± 1.93 | 13.36 ± 1.56 | 8.99 ± 2.64 | 14.38 ± 0.90 | 9.73 ± 2.02 |
| Total C stock | Mg ha−1 | 29.7 ± 1.8 | 28.1 ± 1.0 | 30.0 ± 1.4 | 28.6 ± 1.9 | 29.3 ± 1.3 | 28.1 ± 1.6 |
| Total NH4+ stock | Mg ha−1 | 5.21 ± 0.99 | 5.70 ± 0.59 | 5.44 ± 0.51 | 5.20 ± 0.84 | 5.41 ± 0.64 | 6.42 ± 0.41 |
| Pi stock | kg ha−1 | 14.9 ± 2.5 | 7.1 ± 2.5 | 16.5 ± 2.0 | 11.3 ± 3.3 | 17.8 ± 0.9 | 12.4 ± 2.7 |
| Physical soil variables Bulk density | g cm−3 | 1.25 ± 0.02 | 1.24 ± 0.01 | 1.23 ± 0.01 | 1.26 ± 0.01 | 1.24 ± 0.03 | 1.27 ± 0.02 |
| Macroaggregates | % | 17.3 ± 0.7 | 30.0 ± 2.3 | 18.5 ± 0.8 | 24.2 ± 2.0 | 16.3 ± 1.5 | 30.2 ± 2.3 |
| Factors | Chemical Soil Variables | Physical Soil Variables | ||||||
|---|---|---|---|---|---|---|---|---|
| Total C Content | NH4+ | Pi | Total C | NH4+ | Pi | Soil | Bulk | |
| Content | Content | Stock | Stock | Stock | Macroaggregates | Density | ||
| Main effects | ||||||||
| Earthworm inoculation (E) | 0.397 | 0.216 | 0.276 | 0.708 | 0.281 | 0.225 | <0.001 | 0.214 |
| Cropping system (S) | 0.840 | 0.643 | 0.632 | 0.827 | 0.466 | 0.704 | 0.384 | 0.702 |
| Block | 0.029 | 0.066 | 0.255 | 0.031 | 0.128 | 0.166 | 0.490 | 0.077 |
| Interaction | ||||||||
| E*S | 0.878 | 0.620 | 0.966 | 0.963 | 0.433 | 0.954 | 0.063 | 0.423 |
| Covariates | ||||||||
| Initial total C content | 0.999 | 0.229 | 0.801 | |||||
| Initial total N content | 0.210 | 0.325 | 0.417 | |||||
| Initial Pi content | 0.289 | 0.729 | 0.276 | |||||
| Initial total C stock | 0.756 | 0.141 | 0.393 | |||||
| Initial total N stock | 0.336 | 0.378 | 0.278 | |||||
| Initial Pi stock | 0.240 | 0.602 | 0.148 | |||||
| Soil Variables | Units | Treatments | |||||
|---|---|---|---|---|---|---|---|
| TR0 | TR+ | CA | |||||
| EW0 | EW+ | EW0 | EW+ | EW0 | EW+ | ||
| ∆C content | g kg−1 | 6.02 ± 0.76 | 7.48 ± 0.65 | 4.92 ± 1.30 | 7.40 ± 1.71 | 8.55 ± 1.49 | 9.77 ± 1.70 |
| ∆Pi content | mg kg−1 | 0.04 ± 0.71 | 1.29 ± 1.88 | 2.94 ± 1.35 | 3.63 ± 2.63 | 4.01 ± 1.28 | 5.66 ± 2.40 |
| ∆C stock | Mg ha−1 | 8.95 ± 1.43 | 10.30 ± 0.74 | 7.78 ± 1.19 | 10.35 ± 2.30 | 12.54 ± 2.02 | 14.12 ± 2.23 |
| ∆Pi stock | kg ha−1 | 1.12 ± 1.29 | 1.91 ± 2.51 | 4.54 ± 1.66 | 4.92 ± 3.38 | 6.23 ± 1.31 | 3.20 ± 0.28 |
| Soil Variables | Units | Three-Way ANOVA | |||
|---|---|---|---|---|---|
| Earthworm Inoculation (E) | Cropping System (S) | Block | E*S | ||
| ∆C content | g kg−1 | 0.135 | 0.089 | 0.397 | 0.882 |
| ∆Pi content | mg kg−1 | 0.465 | 0.132 | 0.834 | 0.970 |
| ∆C stock | Mg ha−1 | 0.218 | 0.056 | 0.415 | 0.934 |
| ∆Pi stock | kg ha−1 | 0.659 | 0.120 | 0.709 | 0.971 |
| Plant Variables | Three-Way ANOVA | |||
|---|---|---|---|---|
| Main Effects | Interaction | |||
| Earthworm Inoculation (E) | Cropping System (S) | Block | E*S | |
| Plant growth | ||||
| Height 1MAS | 0.306 | 0.028 | 0.086 | 0.771 |
| Height 2MAS | 0.028 | 0.351 | 0.713 | 0.375 |
| Height 3MAS | 0.008 | 0.403 | 0.983 | 0.477 |
| Shoot biomass | 0.126 | 0.084 | 0.268 | 0.280 |
| Grain biomass | 0.047 | 0.359 | 0.631 | 0.813 |
| Shoot biomass | 0.060 | 0.300 | 0.535 | 0.584 |
| Root biomass | 0.984 | 0.094 | 0.761 | 0.828 |
| Total biomass | 0.179 | 0.120 | 0.847 | 0.673 |
| Shoot:root ratio | 0.091 | 0.352 | 0.262 | 0.997 |
| Plant nutrition | ||||
| Shoot N amount | 0.680 | 0.310 | 0.184 | 0.314 |
| Grain N amount | 0.037 | 0.421 | 0.345 | 0.843 |
| Aboveground N amount | 0.063 | 0.486 | 0.271 | 0.714 |
| Shoot P amount | 0.140 | 0.547 | 0.311 | 0.671 |
| Grain P amount | 0.056 | 0.392 | 0.065 | 0.848 |
| Aboveground P amount | 0.167 | 0.637 | 0.070 | 0.781 |
| Plant production | ||||
| Grain yield | 0.040 | 0.281 | 0.668 | 0.802 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ratsiatosika, O.; Razafindrakoto, M.; Razafimbelo, T.; Rabenarivo, M.; Becquer, T.; Bernard, L.; Trap, J.; Blanchart, E. Earthworm Inoculation Improves Upland Rice Crop Yield and Other Agrosystem Services in Madagascar. Agriculture 2021, 11, 60. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11010060
Ratsiatosika O, Razafindrakoto M, Razafimbelo T, Rabenarivo M, Becquer T, Bernard L, Trap J, Blanchart E. Earthworm Inoculation Improves Upland Rice Crop Yield and Other Agrosystem Services in Madagascar. Agriculture. 2021; 11(1):60. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11010060
Chicago/Turabian StyleRatsiatosika, Onja, Malalatiana Razafindrakoto, Tantely Razafimbelo, Michel Rabenarivo, Thierry Becquer, Laetitia Bernard, Jean Trap, and Eric Blanchart. 2021. "Earthworm Inoculation Improves Upland Rice Crop Yield and Other Agrosystem Services in Madagascar" Agriculture 11, no. 1: 60. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11010060