Mortierella Species as the Plant Growth-Promoting Fungi Present in the Agricultural Soils


1
Department of Industrial and Environmental Microbiology, Institute of Biological Sciences, Faculty of Biology and Biotechnology, Maria Curie-Sklodowska University, Akademicka 19, 20-033 Lublin, Poland
2
Department of Plant Physiology and Biophysics, Institute of Biological Sciences, Faculty of Biology and Biotechnology, Maria Curie-Sklodowska University, Akademicka 19, 20-033 Lublin, Poland
*
Author to whom correspondence should be addressed.
Agriculture 2021, 11(1), 7; https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11010007
Submission received: 4 December 2020
/
Revised: 19 December 2020
/
Accepted: 21 December 2020
/
Published: 24 December 2020
(This article belongs to the Special Issue Plant-Microbe-Soil Interactions in Agroecosystems)
Abstract
:In the most recent scientific reports based on the DNA or RNA-analyses a widespread presence of the filamentous fungi, Mortierella in various environments has been shown. Some strains of this genus belong to the plant growth-promoting fungi (PGPF) and are found in the bulk soil, rhizosphere and plants tissues. These microorganisms are also often found in the extremely hostile environments, responsible for improving access to the bioavailable forms of P and Fe in the soils, the synthesis of phytohormones and 1-aminocyclopropane-1-carboxylate (ACC) deaminase, and last but not least the protection of agricultural plants from pathogens. Furthermore, earlier reports classified Mortierella spp. as the saprotrophic microorganisms isolated from the forest litter, and nowadays their status as a very valuable decomposers in the agricultural soils was confirmed. The key features like the ability to survive under very unfavorable environmental conditions and the utilization of carbon sources contained in polymers like cellulose, hemicellulose, chitin make these fungi efficient as the agricultural inoculants. The growing interest in the application of Mortierella spp. is mainly due to the potential use of this genus in the increase of the nutrient uptake efficiency, positive effect in crop protection against adverse conditions, and reduction of chemical fertilizers and pesticides applied. Moreover, activities of Mortierella species selected from the wild or cultivated plants influence the soil microbiota and support the performance of the beneficial microorganisms enhancing significantly crop yield.
1. Introduction
Soils are very diverse systems consisting of various types of organic and inorganic components. One of the most essential constituents of the soil are microorganisms with the ability to degrade a number of compounds and to create a very dynamic soil environment of the transformed substances acting as decomposers. Their activities result in a higher bioavailability of the key elements such as N, organic C, P, Fe, and other substances not exclusively for themselves, but also for other organisms which is crucial in agricultural soils [1,2]. The reduced application of inorganic synthetic fertilizers in favor of biofertilizers containing microorganisms has an influence in the increasing uptake and availability of mineral nutrients for plants. A specific group of these environmentally friendly microbial fertilizers can be effectively applied on the seeds, plant surfaces, or to the soil in order to promote plant growth by supplying them with the nutrients indispensable for their proper functioning [3]. The development and availability of the new methods of identification of microorganisms within the last 20 years led to the detection of plant growth-promoting Mortierella species in the soils all over the world. The present paper is focused on the plant growth-promoting abilities and the other properties of Mortierella species which seem to be helpful in enhancing plant development in agricultural applications.
2. From the Pioneering Microorganisms to the Important Agricultural Inoculants
2.1. Isolation and Identification of Mortierella spp. from the Soils of the World
Over the last two decades numerous studies indicated the occurrence of Mortierella species in diversified environments, e.g., rocks, caves, mines, asbestos fibers, rivers, lakes, plant tissues soils, and rhizospheres (including agricultural areas) at each latitude [4,5,6,7,8,9]. Recently, researches employing molecular methods revealed that not only the Aspergillus and Penicillium genera, but also Mortierella strains are the most abundant filamentous fungi in the soils around the world [10,11,12,13,14,15,16]. Moreover, the Mortierella omnipresence may be confirmed by in the environment the effects of 35-year application of an organic fertilizer to the agricultural soil in China (Anhui Province) in order to promote plant growth. Mortierella elongata was found to be the dominant fungal species in this soil [17].
New species of Mortierella, being constantly discovered, are the most common soil-inhabiting fungi. Moreover, they are one of the largest genus of Mucorales within Zygomycetes. In turn, filamentous fungi with asseptate hyphae (coenocytic hyphae) belong to the Mortierellales order. Mortierella species form colonies often described as pale white, white or whitish (sometimes little dark whitish at the center) zonate, forming a typical rosette pattern [7,8,15,18,19]. In the course of time, the reverse side of the colonies may become yellowish. Cottony aerial hyphae of Mortierella species form one of the simplest sporangia (without or with rudimentary columella) which are pigmented or colorless [11]. The Mortierella colony growth may be accompanied by the garlic-like odor [15,20]. After incubation at 5–10 °C, these fungi are known as forming sporangia on the nutrient-poor media [21], such as water agar [11]. Contrarily to these results, Mortierella antarctica investigated by Ozimek et al. [8] failed sporulation on this medium (water agar) at low temperatures and the genetic analysis was required to confirm the species affiliation. However, Kuhlman [22] observed that neither the low temperature nor the different substrates affected the type of spore formation of Mortierella species, i.e., M. ericetorum, M. parvispora, M. bainieri, M. candelabrum, M. elongata, M. minutissima, M. pulchella, M. zychae and M. marburgensis. On the other hand, the studied Mortierella species often formed spores on the hemp seed agar or/and diluted Pablum agar. It is probable that this medium was prepared from Pablum—a product for malnutrition of infants consisting of the ground and precooked oatmeal, yellow corn meal, bone meal, dried brewer’s yeast, and powdered Alfalfa (Medicago sativa) leaf [23]. In addition to the mineral compounds, Pablum is rich in vitamins A, B1, B2, D, E and reduced iron. Due to its composition, “Pablum Cereal Agar” is recommended as a medium stimulating sporulation of fungi [24,25]. Furthermore, Gao et al. [26] obtained very good sporulation of Mortierella isabelina on medium with xylose. The lack of morphological characteristics of Mortierella in the past has resulted in difficulties in identification of the species and can lead to the problems with the storage of fungal strains in the collections. Moreover, the comparison of culture-based and culture-independent methods mostly supports the higher taxonomic precision of the methods based on DNA or RNA analyses which allowed to confirm the abundance of Mortierella species in different environments [10].
The positive contribution of these fungi in the agricultural soils (Table 1) as well as in the plant tissues should lead to the discovery of the optimal culture conditions and spore-forming media enabling analysis of the complex interactions between plants and microorganisms. It is worth highlighting that the pathogenic species are very rare in this genus, that is why Mortierella species are very promising sources of plant growth-promoting inoculants for agriculture [27].
2.2. Mortierella Features Useful for Agricultural Inoculation
According to literature, plant growth-promoting microorganisms (PGPM) are divided into plant growth-promoting bacteria (PGPB) or rhizobacteria (PGPR) as well as plant growth-promoting fungi (PGPF) [37]. In the case of PGPM or precisely PGPF, filamentous fungi have a significant advantage over bacteria resulting from their growth. Filamentous mycelium stabilizes the soil structure, penetrates the pores of soil and the solid substrates like rocks and minerals into their cavities, slots and cracks followed by their physical changes related to the structure and size. In order to occupy large areas by hyphae, they excrete acidic compounds useful in drilling microtunnels in the solid surroundings [38,39]. The recent researches focused on the interactions between minerals (abiotic compounds) and filamentous fungi (biotic compounds) interoperable in bioweathering [39,40,41].
Microorganisms isolated from the extremely nutrient-poor (oligotrophic) environments, e.g., bare rocks, sandstones, newly created dunes, and areas destroyed by fires may be used as very efficient components of biopreparations used in agriculture [38,39]. Their radially-spreading mycelium, covers small fragments of the fragmented substrate. As it was discovered by scientists, the microfungal activity was observed near the weathered stalactite collected from the Penico cave located in South Portugal [42]. The identification of fungi by rRNA sequencing revealed that some spherical cells were Mortierella sporangiospores. The photographs taken in scanning electron microscopy demonstrated not only the presence of the subeuhedral, but also rounded shaped calcite crystals where the Mortierella hyphae was detected. The fungal hyphal tips and networks around crystals suggested microbial calcite dissolution [42]. These documented properties of the Mortierella species fungi may explain the prevalence of their occurrence in the cultivated soils.
Fungi belonging to the Mortierella genus are regularly recorded in Arctic, Antarctic, and Alpine soils where with the low temperatures of ground and water the activities of a number of processes in the oligotrophic conditions decreases [43,44,45]. Some of Mortierella isolates defined as psychrotrophs or psychrophiles are referred to as “snow mold” due to the ability to adapt to severe conditions (low temperatures) [46,47]. Psychrotrophs and psychrophiles grow at low temperatures, i.e., about 5 °C or below, with the optimum temperature at about 15 °C or lower for psychrophiles or above 15 °C with maximal growth at higher than 20 °C for psychrotrophs [8,48].
The studies on the molecular mechanisms underlying cold adaptation of M. isabellina M6-22 in the extreme environmental conditions [47] confirmed the presence of a higher content of polyunsaturated fatty acids (PUFAs), which maintained the better membrane fluidity for microbial adaptation to low temperatures. Moreover, the agricultural soils are periodically influenced by low temperatures, especially within the range of the temperate climate. Among all fungi identified in the soils of samples from the Wyoming’s Agricultural Experimental Station in the USA about 8% of isolates were ice-nucleationactive (INA) and all of them were M. alpina strains [49].
Among 20 genera of psychrotrophic fungi originating from Arctic soils of Franz Joseph Land and growing at 0 °C on malt extract agar (MEA), Mortierella isolates belonging to the following species: M. alpina, M. exigua, M. hyalina, M. minutissima, and M. parvispora) were present [50]. A number of Mortierella fungi (oligotrophs) were able to grow slowly on the carbon- and/or nitrogen-free silica gel media. The growth on the carbon-deficient media suggests that the low level of available carbon in the soil should not be the limiting factor of growth of some Mortierella species. Mortierella hyaliana was able to grow at 0 °C as a psychrooligotrophic fungus on a silica gel containing N-source and C-deprived source [50].
The multiple agricultural biopreparation containing potentially useful microorganisms do not improve crop yields. One of the reasons for that may be the screening of strains which covers their ability to improve bioavailable forms of P, K, Fe, and others in the soil, while the occupation of niches by inoculants depends on the availability of the C substrate [51]. Free-living saprophytic fungi perform the major role in the carbon delivery from the decomposition of plant residues where many of the nutrients contained in the soil are deposited [40]. The periodic lack of bioavailable macro- and micro-element forms, the availability of which depends practically on the intensity of microbial transformations, is noticed under adverse environmental conditions.
Saprotrophic fungi, which degrade the polymers such as cellulose, hemicellulose, chitin, render plant residues more amenable to decomposition for other microorganisms forming the organic soil which is necessary for good quality of agricultural soil [52]. Saprotrophic Mortierella species are often isolated among forests soils and dead wood fungal communities [53,54]. For instance, Mortierella minutissima, M. alpina, and M. verticillata were identified in the forest soil in Romania [6] and one of Mortierella sp. selected by Gawas-Sakhalkar and Singh [55] revealed a high cellulase activity. In the studies on cellulose degradation in rye straw the ability to decompose plant remnants was also identified for Mortierella verticillata and Mortierella humilis [56].
Furthermore, Jackson [57] investigated fungi from the species of M. spinosa, M. elongata, M. alpina, M. humicola, M. minutissima, M. horticola, and M. exigua in the pasture soil and described a strong chitinolytic activity in the strains from root surface. Additionally, M. alpina was also found to decomposed hemicellulose. Again, the unargued potential of Mortierella was indicated in the experiments where peat substrate enriched with chitin was used in the cultivation of lettuce seedlings (Lactuca sativa var. capitata ‘Alexandria’) [58].
Two of the M. alpina rhizospheric isolates were characterized by a protease activity and both protease and urease activities were analysed for Mortierella simplex [55]. Additionally, there are reports of other activities of these fungi, e.g., in xylose (M. isabellina) [26] and starch (Mortierella alliacea and M. alpina) [59,60] degradation as well as a high amylolytic activity observed for Mortierella sp. and M. minutissima.
Access to carbon contained in different polymers like cellulose, hemicellulose, and chitin combined with staying active at lower temperatures provides fungi belonging to the Mortierella genus advantageous in the unfavorable soil conditions.
3. Plant Growth-Promoting Abilities of Mortierella Species
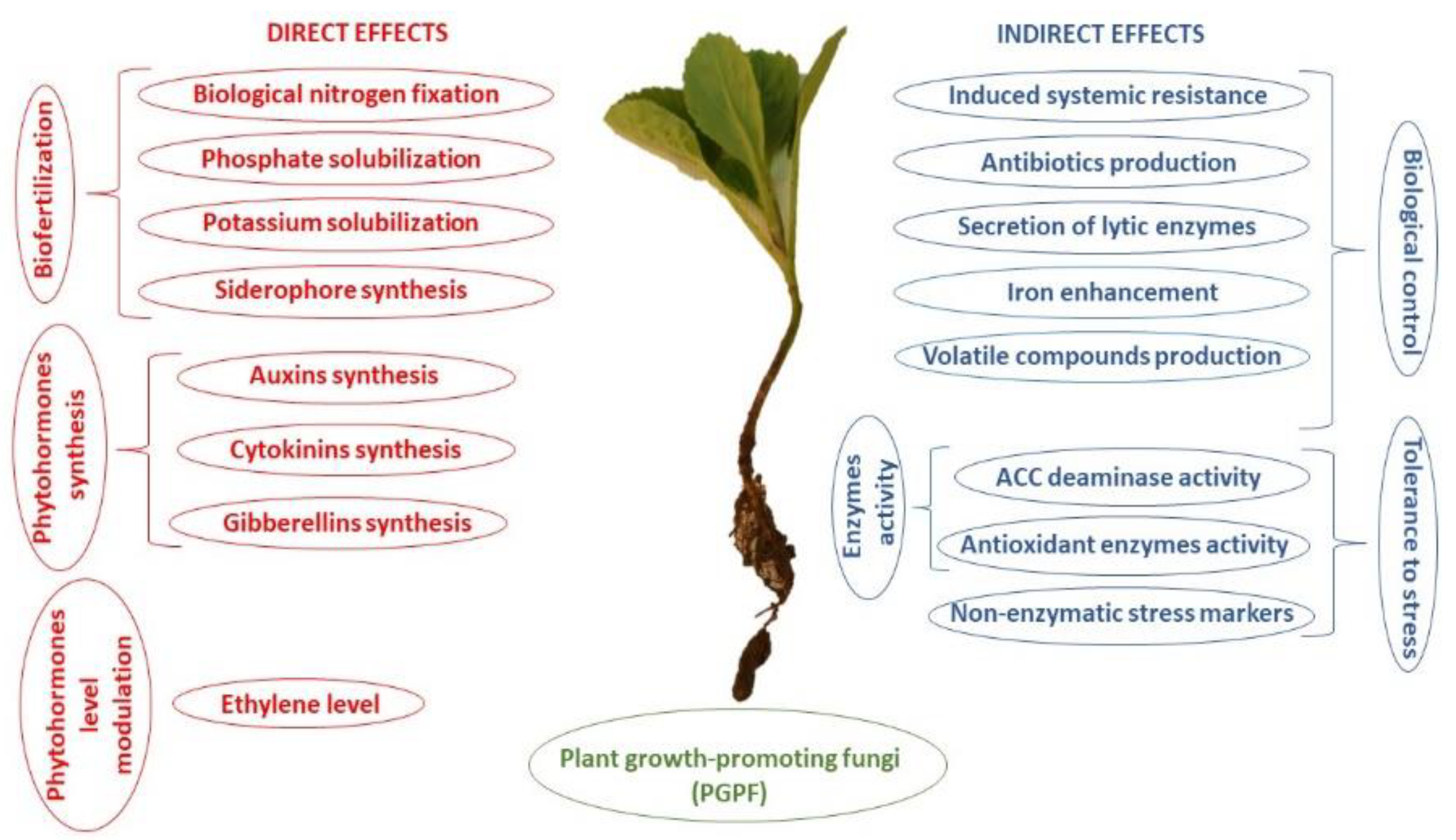
Mortierella species, representing plant growth-promoting abilities (Figure 1) are isolated from the agricultural soils (bulk soil, rhizosphere soil) (Table 1).
Recent studies of rhizosphere soils or endophytes of various plant species proved that the presence of Mortierella provides a higher resistance at soil-borne pathogens or/and improve plant growth (Table 2) [9,33,64,65].
3.1. Mortierella Fungi—Effective in Increasing Bioavailable Forms of P in Agricultural Soils
The fungal agricultural inoculants are free-living fungi or arbuscular mycorrhizal fungi (AMF) screened for plant growth-promoting abilities (Table 2) [68]. Innovations in the laboratory technology expanded our understanding of multiple interactions between plant and microorganisms resulting in the growth of crops. One of the most important traits of plant growth-promoting strains is enhancing the bioavailability of certain essential elements like P, K, and Fe. The sorption of bioavailable P (Piav) from the soil solution on the surface of the soil minerals is one of the limiting factors of the crop productivity. Mortierella species are described as the phosphate solubilizing fungi (PSF) [71,72,73,74], e.g., Mortierella sp. isolated from Andisol of Hawai increased the concentration of Piav [75].
Another mechanism reducing the P loss from the soil and improving the Piav level is desorption of P (“run off”) from the surface of the soil particles. Osorio et al. [76] investigated the effectiveness of Mortierella sp. to desorb Pi from the soil minerals: goethite, allophane kaolinite, and montmorillonite distributed in a number of agricultural soils [77]. Sorbed Piav can be held very strongly on the surfaces of some minerals, especially the one which is bounded by iron and aluminum hydroxides [78]. Mortierella sp. was effective in desorbing Piav from the soil minerals (except from allophane) differing in their Piav sorption capacity. The highest concentration of desorbed Piav was found for montmorillonite and kaolinite. The desorption also depended on the production of oxalic acid by Mortierella sp. [76].
Mortierella sp. isolate was detected in the soil dominated by montmorillonite [79]. The values of pH in the soil solution inoculated with microorganism at the end of the incubation period were significantly lower as compared to the non-inoculated samples. Osorio et al. [19] observed that Mortierella sp. effectively dissolved the rock phosphate by decreasing pH of the medium from 7.7 to 3.0. The low molecular weight of organic acids, like oxalic, malic, acetic, formic, gluconic, citric, lactic, 2-ketogluconic, and tartaric and citric acids secreted by filamentous fungi are considered to be one of the main mechanisms of the inorganic phosphate solubilization by Mortierella sp. [40,66,72]. Two Mortierella species cultivated on three different media synthetized various acids. In the presence of NH4Cl in the culture, mainly acetate, succinate and formate were excreted, while M. ramanniana synthetized lactate, succinate, acetate, and formate [80]. Moreover, silicates and aluminosilicates levels might be dissolved by the product of microfungal activity, the organic acids, i.e., oxalic acid [40]. This acid has also the capacity to desorb P from mineral surfaces contributing to the increase of bioavailable P in the environment [75,81,82,83].
In the agricultural soils (Table 1), Piav is supplemented with chemical fertilizers obtained from phosphate rocks, which are non-renewable resource. The problem lies in the fact that Piav applicated in many soils is fixed by cations resulting in low P-fertilization-use efficiency. In the acidic soils, most phosphate anions are fixed mostly by Fe and Al cations and in alkaline soils Ca cations are responsible for phosphate anions precipitation. The microbial solubilization of P-minerals naturally located in the soils or refixation of applied phosphates provide stable Piav source which is very important for sustainable agriculture [84].
Ceci et al. [85] showed the ability of M. globalpina to mobilize P from insoluble forms. This strain isolated from the soil on the solid medium with 5% insoluble tricalcium phosphate (TCP) as the only P source revealed halo zone of about 11.7 mm with growth diameter of 30.0 mm after seven days of incubation. The microscopic observations of mycelia obtained on a liquid medium with TCP have shown mineral precipitates loosely attached on the surface of fungal biomass and embedded in the fungal biomass.
Zhang et al. [67] (Table 2) investigated the interactions between phosphate solubilizing Mortierella sp. (PSM) with Glomus aggregatum and G. mossae (arbuscular mycorrhizal fungus—AMF). The inoculation of PSM isolated from the salty coastal soil samples in China (and one of the AMF strains) caused promotion of growth of halophyte, Kostelelzkya virginica, at free different salinity levels. K. virginica is a salt-tolerant plant effective in amelioration of the very saline soil conditions and in saline agriculture [86]. After combined inoculation of Mortierella and Glomus species, the higher concentration of Piav in all the inoculated bulk soils at different salinity levels were observed (as compared to the non-inoculated bulk soil and to the soil inoculated only with Glomus species). This promoting effect was very strong at the 100 mM NaCl level of salinity and about 40% higher shoot DW and above 100% higher root DW were achieved (as compared to the non-inoculated control at the same salinity conditions). These results may suggest that, there are non-stress conditions for Mortierella sp. at particular salt concentration in the soil [67].
Various proportions of two inoculants: Mortierella sp. (PSF) and Glomus mosseae (AMF) were prepared to study their efficiency in pot experiment [68]. Fungi were introduced to the rhizosphere soil of castor bean (Ricinus communis) seedlings, an important plant cultivated in China. With the increased ratio of Mortierella sp. to G. mosseae, the extension of AMF root colonization, elevation of Piav concentration in the soil solution and higher shoot and root weight were observed. This might also result in positive interactions between Mortierella sp. and G. mosseae and with the native microorganisms and beneficial changes in the activity of the soil enzymes (Table 2) [68].
The efficiency of phosphate solubilizing microorganisms as inoculants in the agricultural soils should not be studied only in vitro. The higher Piav level in the soil solution should not be the only criterium for screening PSM activity. On the other hand, the lack of the higher level of Piav in the soil after inoculation with PSM does not indicate inactivity of the inoculant. The content of Piav is changing very dynamically and the best effect of PSM efficiency can be the higher level of P in the inoculated plant and the higher yield [67]. Osorio and Habte [66] inoculated Leucaena leucocephala with Mortierella sp. isolated from the rhizosphere of mature Leucaena leucocephala and indicated this fungus as a very efficient phosphate rock (PR) solubilizer. In the experiment with Mortierella sp. (PSM) and Glomus sp. as coinoculant of the plant, the amount of Piav increased (over 73%) in the treatment with PR application, caused the higher content of P in the shoots and higher shoot DW of L. leucocephala (about 29%) (Table 2).
Two Mortierella alpina and one M. simplex strains isolated from rhizosphere revealed phosphatase activity [55]. In the preliminary screening of fifty six isolates from Arctic soils, M. alpina PG40 demonstrated the strongest phosphatase activity among Mortierella strains. M. simplex PG26 and Mortierella schmuckeri PG45 strains had low activity of this enzyme [55]. In most soils, the mineral compounds have a smaller share of the total pool of this element than the organic phosphorus compounds where the phosphate group is attached by proteins, sugars, lipids, and nucleotides. Phosphatases are enzymes synthetized by plant roots and microorganisms which are often induced by low concentration of Piav in the soil solution [24]. On the other hand, at the low level of anions bioavailable for plants (H2PO4− and HPO42−), the decrease in crop productivity is proved.
3.2. Siderophore Producing Mortierella Species—Efficient in the Increase of Fe Bioavailability
Iron is an essential micronutrient required for the plant and microbial growth. When present at lower content in the agricultural soils, Fe is responsible for reduction of plant productivity. In the upper soil layers under aerobic conditions, Fe solubility is low and it is mostly found in the form of Fe3+ which is not available to plants [87]. One of the specific mechanisms developed by microorganism is synthesis of Fe3+-complexing compounds followed by the transportation and accumulation of Fe. Siderophores are known to be synthetized and excreted by soil fungi as a response to Fe starvation in the environment and as a consequence, microbial inoculants with this ability may strongly enhance the plant growth and resistance through the sequestration of Fe from the soil [88,89].
To the best of our knowledge, only few studies have explored the capability of Mortierella species to Fe-complexation. In 1972, Bozarth and Goenaga extracted mycoferritine, the iron-binding protein, from mycelium of Mortierella alpina isolated from the soil [90]. After separation from the lyophilized mycelium, this protein contained 17% of Fe. On the basis of the results obtained from HPLC-analysis, it was indicated that rhizoferrin (polycarboxylate-type siderophore) was synthetized by Mortierella vinaceae cultivated under Fe-limiting conditions [91].
M. alpina CS10E4 strain investigated by Wani et al. [9] also formed orange halo zone on (CAS)-blue agar. Although siderophores are mainly specific for Fe3+, these chelating compounds also bind other metals, such as Cd2+, Cu2+, and Zn2+ [92,93,94]. The presence of optimal dose of bioavailable heavy metals in the soil might promote plant growth.
Mortierella turficola CQ1 strain isolated from the rhizosphere of Panax ginseng (Korean ginseng) showed positive reaction on (CAS)-blue agar. Moreover, on the liquid media amended with different concentrations of Zn2+ (50 to 250 µg/mL), the percentage of siderophore units in the culture of this strain was the highest at 200 µg Zn2+/mL [95].
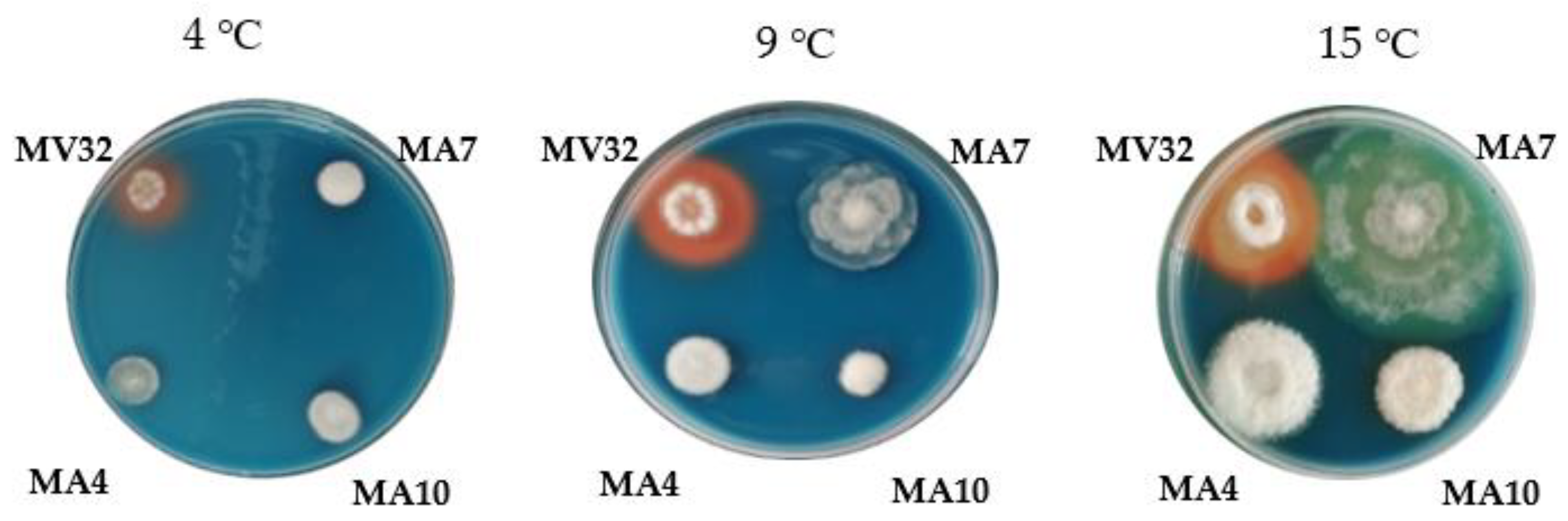
Until now, there is limited data concerning siderophore production by Mortierella species, especially these isolated from the area of harsh climatic conditions. Among four tested Mortierella strains isolated from Spitzbergen soils only M. verticillata DEM32 was capable of releasing these low-molecular mass compounds with a high affinity to Fe and change the color of solid medium after incubation at three different temperatures (Figure 2) (data not published). Moreover, except for some plant growth-promoting activities detected at lower temperatures [8], this fungus also revealed the production of siderophores at conditions found in the temperate climate soils making it an attractive inoculant for the agricultural soils.
3.3. Production of Phytoregulators by PGP-Mortierella Fungi
Among phytoregulators, phytohormones are chemicals involved in a variety of physiological and biochemical processes of plants at very low concentrations. The most important and well-characterized phytoregulator is 1-aminocyclopropane-1-carboxylate (ACC) deaminase and among phytohormones of well-established importance are auxins, gibberellins, cytokinins, ethylene, abscisic acid, but also brassinosteroids, jasmonates, and salicylic acid [96,97,98,99,100]. It is known that environmental fungi are also able to produce some phytoregulators, e.g., phytohormones (Table 2) [4,97,101].
Among auxins, the key hormone produced by fungi is indole-3-acetic acid (IAA). IAA biosynthesis can be tryptophane (Trp)-dependent or independent. IAA was found both in the culture medium and inside the fungal mycelium [97]. The higher level of IAA can be the result of the lower sensitivity of microorganisms to IAA or the differences in the regulation of IAA biosynthesis. In fungi, two pathways were identified, the indole-3-pyruvic acid (IPY) and the indole-3-acetamide (IAM) ones [101,102]. The knowledge of the complex cross-regulatory relations between hormone pathways which are useful in the regulation of plant development and stress responses [98,103,104,105] still requires deeper research and understanding. Soil microorganisms and plant endophytes producing phytohormones have a great physiological importance for microbe-plant interactions. Among other free living fungi, Mortierella species are known to produce a variety of phytoregulators, like IAA, gibberellic acid (GA), and ACC deaminase [8,70]. Their activity in the agricultural soil may play significant role in growth improvement and stress tolerance of cultivated plants enhancement.
M. alpina synthetized over 70 mg of IAA/L with Trp in medium [9]. In the research of efficient agricultural inoculant to the application in lower temperatures accompanying in a sprouting in temperate climate Mortierella spp. isolated from Spitzbergen were investigated [8]. M. verticillata DEM32 synthetised the highest amount of IAA at 15 °C with initial dose of Trp (about 1.5 mM). Moreover, at the same conditions the root and shoot weight of Winter wheat seedlings (Triticum aestivum cv. Arcadia), inoculated with this strain, was significantly higher (about 40%). At 9 °C psychrotrophic M. antarctica also produced IAA, however, no Trp was added in the medium confirming promoting effect on wheat seedlings. Additionally, both strains synthesized GA for growth promotion and elongation of plant cells. Among various plant growth-promoting fungal mechanisms is the synthesis of ACC deaminase. This commonly known phytoregulator is produced in order to control the ethylene content. Higher ethylene level in the plant tissues is often the result of various stresses and can lead to strong growth inhibition of the crop. M. antarctica DEM7 demonstrated the ability to synthetize ACC deaminase (cleaving plant ethylene precursor) [106] at 9 °C, 15 °C, and 20 °C, but with the highest efficiency at the lowest temperature [8]. Genome characteristics of M. elongata selected from the field soil showed the presence of biosynthesis genes of ubiquitous plant hormones IAA and abscisic acid (ABA) (Table 2) [17]. ABA-mediated signalling is not only involved in the plant developmental process but its higher concentration may be increased by the plant resistance to the biotic and abiotic stresses [107]. Li et al. [17] detected 40% increase of ABA and IAA in maize roots inoculated with saprotrophic M. elongata. Moreover, the plant biomass, soil enzyme parameters and concentration of bioavailable nutrients also increased (Table 2).
The fungal activity causes also higher concentration of regulators in its host plant tissue or/and they may affect plants auxin-responsive genes [108]. It is also known that Mortierella produces and accumulates IAA in its mycelia. After inoculation, the IAA level in roots colonized by fungus was significantly higher. Furthermore, the jasmonic acid (JA) level in plants colonized by M. hyalina tissue was over five-times higher compared to the concentration of its phytohormone in roots inoculated with other tested fungi [107,108]. Endophytic fungi (described below) produce and accumulate phytoregulators in their mycelia.
4. Endophytic Mortierella Strains
Endophytes are microbiota, i.e., bacteria and fungi able to colonize internal tissues of plants without symptoms [106]. They are isolated from different parts of plants, e.g., roots, stems, leaves, fruits, bulbs, and seeds [109,110]. Moreover, they can extend their niches, e.g., from roots to the aerial parts of the plant [111]. The intracellular growth of the endophytic fungi such as Mortierella chlamydospora and M. indohii results in the penetration of the cortex cells of the plant [112].
Understanding the interaction between endophytes and host-plants allows to notice the importance of composition of the plant’s microbiome for its optimal development and immunity [113]. The close associations of endophytes with agricultural plants could be beneficial in crop growth promotion based on the usefulness of products resulting from the trophic interactions [106].
Investigations involving Mortierella revealed that Morterella candelabrum was found in the seeds of Crataegus azarolus [109]; M. hyalina, M. isabellina, and M. ramanniana were isolated from the cones of Pinus densiflora growing in Japan and these fungi also were found among early stage decomposers of cones on the ground [110].
These endophytes were also isolated from the roots of Holcus lanatus—a grass occurring in humid soils in temperate zones around the world [114,115]; Huperzia serrate—a very important medicinal plant native to eastern Asia used for over 1000 years in China [116] and Pteridium aquilinum—a species common in moorlands and pastures of Great Britain (Mortierella sp. 1, M. sp. 2 and M. ramanniana var. ramanniana L.) [117]. Root-derived isolates of the endophytic fungi commonly existing in agricultural plant tissues could be beneficial for crops biopreparates. A few Mortierella strains: M. gamsii; M. verticillata and Mortierella zonata were found in Fragaria vesca roots growing in Essex in United Kingdom [118]. Moreover, remarkable growth parameters of F. vesca (number and length of leaves and roots) were associated with the occurrence of Mortierella endophytes in its tissues. M. hyalina and M. indohii were the most abundant species among this genus identified in the endophytic microbiome of tomato roots [111].
The Populus trichocarpa endophyte M. elongata PMI93 can alter the genes expression of host-plant involving GA, JA and ethylene signalling which results in an improvement of plant growth and plant DW (over 30% of DW in one year) [113].
The investigation of the microbiome composition in the healthy wild plants seems to be crucial for their optimal growth and development and is a significant direction in the development of sustainable agriculture. The morphological and physiological traits of the host plant, Crocus sativus, an important spice and medical plant, were improved in the presence of the endophytic M. alpina CS19E4 strain [9]. From 14 different sites in India, the fungal endophytes of the same plant (C. sativus) were analyzed and the efficient endophytes were isolated [119].
These endophytic fungi are also effective in the synthesis of phytohormones, e.g., IAA was detected in the mycelia of M. hyalina [52]. As a result, higher auxin and jasmonate levels in its host plant (Arabidopsis thaliana) tissues were demonstrated and the growth of Arabidopsis was promoted.
5. Contribution of the Mortierella Species in the Healthy Condition of Plants and Soils
Among plant growth-promoting microorganisms, the potential bio-inoculants should be able to increase agricultural production and immunity of the arable crops linking different activities [120] and lead to the limitations of toxic products application [121]. The diverse activities of Mortierella fungi is evidenced to effectively support plant immunity. These microorganisms are present in the healthy plant population on plantations and wild gatherings.
The positive impact of Mortierella sp. and M. elongata on the growth and development of the cultivated plants was demonstrated along with the presence of these microorganisms in the fungal community of the healthy Pisum sativum rhizosphere and bulk soil [121]. M. elongata strains were rarely detected in the samples from the infected plants. Over four hundred Mortierella isolates were indicated in the acidic soils of apple orchards collected in Chihuahua in Mexico, e.g., M. capitata, M. sp., M. gamsii aurium, M. alpina, but none of them were pathogenic to the apple rootstocks [33].
The presence of some microorganisms can enhance plant growth and additionally protects plants from disease and abiotic stresses through various mechanisms. A good example of this activity is the synthesis of iron-chelating compounds that are secreted by microorganisms providing them one of the most abundant elements in the soil. On the other hand, limitations in the biologically available forms of Fe (Fe2+) in the soil for pathogens could contribute in the reduction of their number present in the soil [88].
Similarly, Mortierella species possess ability to secrete the lytic enzymes which provide compounds like cellulose, hemicellulose, pectin or chitin available as nutrients. Simultaneously, these compounds are the main structural polysaccharide components of the plant and fungal cell wall and the ability to decompose them gives the possibility to eliminate pathogenic organisms from the soil and even to dominate the environment [122]. Mortierella elongata, the endophyte of Populus trichocarpa, may regulate the genes associated with hypersensitive response and cell wall degradation which are responsible for the plant immunity [113].
M. hyaliana promoting aerial growth of Arabidopsis thaliana simultaneously showed over 30% growth inhibition of Alternaria brassicae on the agar plate [52]. The free of pathogenic Fusarium oxysporum f. sp. cubense (caused Fusarium wilt of banana) soil was dominated by Mortierella species constituting over 36% of the total soil abundance [35]. Very similar results were indicated by Xiong et al. [123] while investigating vanilla soil—suppressive against Fusarium wilt disease. About 37% of the total fungal sequences belonged to the Mortierella genus showing the great potential of these fungi to promote agricultural plants.
Studies indicated that more vulnerable to soil-borne pathogens were the rhizosphere soils poor in Mortierella species [124]. To understand the multidirectional impact of inoculant on the soil microbial populations, the genetic analyses on the reshaping microbiomes are carried out. Li et al. [125] demonstrated that the introduction of M. capitata to maize rhizosphere promoted its growth “by altering the root gene expression levels”, which was also the result of change in the quality and quantity of the rhizosphere bacteria.
It is worth noting that some of these filamentous fungi might produce and accumulate PUFAs like arachidonic, γ-linolenic and eicosapentaenoic acids in their mycelium [126,127]. The synthesis of the fat compounds is one of the microbial cold adaptive mechanisms [128]. For that reason, in the 1990s, mostly psychrotrophic species of Mortierella were isolated from Arctic, Antarctic or alpine soils [50,129]. Polyunsaturated fatty acids which are essential for the human health are also very important nutrients and are widely used in medicine, pharmaceutics and cosmetics [127,130]. Some of PUFAs, e.g., arachidonic acid, are incorporated into the plant oxylipin pathway regulating defense processes in plants and as a result inducing plant resistance [120,131].
The effect of mixture containing lipids from Mortierella hygrophila on plant resistance was observed in the field experiments. The preparation composed of 30% of arachidonic acid stimulated the resistance of potato and sugar beet against Phytophthora infestans and Rhizoctonia solani and increased the yield from 11 to 14%. The mixture also limited the development of powdery mildew of grapes in about 40% [120]. Similarly, among eleven endophytes of Crocus sativus, M. alpina CS10E4 extract demonstrated antimicrobial activity and inhibited more than three pathogens (at least of about 50%) [119]. Moreover, the mechanism of enhancing the plants immunity may be based on the better control of different soil-borne pathogens by synthesis of antibiotics. In the genome of M. elongata also streptomycin, butirosin, and neomycin biosynthesis genes were described [17]. The strong influence of Mortierella on the growth of pathogenic species described Tagawa et al. [132]. M. chlamydospora was proposed as the one of potential agents in controlling of the potato scab pathogens, i.e., Streptomyces. This fungal strain isolated from the potato soil, was found to have a high antagonistic activity against Streptomyces turgidiscabiei. All these examples are pointing to the usefulness of some Mortierella spp. not only as the inoculants in order to improve crop nutrient acquisition, but also to inhibit plant pathogens.
6. Conclusions
In the face of the rapid climate change and the decrease of natural resources necessary to maintain the crop productivity at the current level, there is a clear need for new efficient solutions preserving and enhancing agricultural ecosystems. Mortierella species are common soil and endophytic fungi with various characteristics promoting plant growth and supporting the defense mechanisms in plants. The results achieved among a number of studies highlight positive influence of the applied Mortierella strains on soils activities and plants growth parameters (including crop species).
Recent studies focusing on the impact of the introduced inoculant to the soil microbiome have also proved that the activity of Mortierella fungi may have a beneficial effect on modifying soil microbiological composition and gene activity of some microorganisms.
Author Contributions
E.O. and A.H. prepared the manuscript. E.O. conducted the screening of Mortierella strains for siderophore activity. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by project of National Science Centre, Poland—MINIATURA 1 no. 2017/01/X/NZ9/00837.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support Figure 2 are available upon request from the authors.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ABA | abscisic acid |
| ACC | 1-aminocyclopropane-1-carboxylate |
| AMF | arbuscular mycorrhizal fungi |
| CAS | chrome azurol S |
| DW | dry weight |
| GA | gibberellic acid |
| IAA | indole-3-acetic acid |
| JA | jasmonate acid |
| PGP | plant growth-promoting |
| PGPB | plant growth-promoting bacteria |
| PGPR | plant growth-promoting rhizobacteria |
| PGPF | plant growth-promoting fungi |
| PGPM | plant growth-promoting microorganisms |
| PSF | phosphate solubilizing fungi |
| PSM | phosphate solubilizing microorganisms |
| PUFAs | polyunsaturated fatty acids |
| TCP | tricalcium phosphate |
| Trp | L-tryptophane |
References
- Berg, G.; Smalla, K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol. Ecol. 2009, 68, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.L.; Mortimer, P.E.; Ferry Slik, J.W.; Zou, X.-M.; Xu, J.; Feng, W.-T.; Qiao, L. Variation in forest soil fungal diversity along a latitudinal gradient. Fungal Divers 2014, 64, 305–315. [Google Scholar] [CrossRef]
- Malusá, E.; Sas-Paszt, L.; Ciesielska, J. Technologies for beneficial microorganisms inocula used as biofertilizers. Sci. World J. 2012, 2012, 491206. [Google Scholar] [CrossRef] [PubMed]
- Held, B.W.; Salomon, C.E.; Blanchette, R.A. Diverse subterranean fungi of an underground iron ore mine. PLoS ONE 2020, 15, e0234208. [Google Scholar] [CrossRef] [PubMed]
- Martino, E.; Prandi, L.; Fenoglio, I.; Bonfante, P.; Perotto, S.; Fubini, B. Soil fungal hyphae bind and attack asbestos fibers. Angew. Chem. Int. Ed. 2003, 42, 219–222. [Google Scholar] [CrossRef]
- Matei, G.; Matei, S.; Mocanu, V. Assessing the role of soil microbial communities of natural forest ecosystem. EuroBiotech J. 2020, 4, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.T.T.; Park, S.W.; Pangging, M.; Lee, H.B. Molecular and morphological confirmation of three undescribed species of Mortierella from Korea. Mycobiology 2019, 47, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Ozimek, E.; Jaroszuk-Ściseł, J.; Bohacz, J.; Korniłłowicz-Kowalska, T.; Tyśkiewicz, R.; Słomka, A.; Nowak, A.; Hanaka, A. Synthesis of indoleacetic acid, gibberellic acid and ACC-deaminase by Mortierella strains promote winter wheat seedlings growth under different conditions. Int. J. Mol. Sci. 2018, 19, 3218. [Google Scholar] [CrossRef] [Green Version]
- Wani, Z.A.; Kumar, A.; Sultan, P.; Bindu, K.; Riyaz-Ul Hassan, S.; Ashraf, N. Mortierella alpina CS10E4, an oleaginous fungal endophyte of Crocus sativus L. enhances apocarotenoid biosynthesis and stress tolerance in the host plant. Sci. Rep. 2017, 7, 8598. [Google Scholar] [CrossRef]
- Daghino, S.; Murat, C.; Sizzano, E.; Girlanda, M.; Perotto, S. Fungal diversity is not determined by mineral and chemical differences in serpentine substrates. PLoS ONE 2012, 7, e44233. [Google Scholar] [CrossRef] [Green Version]
- Domsch, K.H.; Gams, W.; Anderson, T.-H. Compendium of Soil Fungi, 2nd ed.; IHW-Verlag: Eching, Germany, 2007; pp. 1–672. [Google Scholar]
- Grządziel, J.; Gałązka, A. Fungal biodiversity of the most common types of Polish soil in a long-term microplot experiment. Front. Microbiol. 2019, 10, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, Q.; Zhang, J.; Ma, C.; Wang, F.; Chen, Y.; Zhang, C.; Zhang, H.; Zhang, J. Characterization and variation of the rhizosphere fungal community structure of cultivated tetraploid cotton. PLoS ONE 2019, 14, e0207903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smit, E.; Leeflang, P.; Glandorf, B.; van Elsas, J.D.; Wernars, K. Analysis of fungal diversity in the wheat rhizosphere by sequencing of cloned PCR-amplified genes encoding 18S rRNA and temperature gradient gel electrophoresis. Appl. Environ. Microbiol. 1999, 65, 2614–2621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, D.R.; Kim, S.W.; Adhikari, M.; Um, Y.H.; Kim, H.S.; Kim, C.; Lee, H.B.; Lee, Y.S. Three new records of Mortierella species isolated from crop field soil in Korea. Mycobiology 2015, 43, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Tian, J.; Bai, C.; Xiang, M.; Sun, J.; Liu, X. The biogeography of fungal communities in wetland sediments along the Changjiang River and other sites in China. ISME J. 2013, 7, 1299–1309. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Chen, L.; Redmile-Gordon, M.; Zhang, J.; Zhang, C.; Ning, Q.; Li, W. Mortierella elongata’s roles in organic agriculture and crop growth promotion in a mineral soil. Land Degrad. Dev. 2018, 29, 1642–1651. [Google Scholar] [CrossRef]
- Jiang, X.; Yu, H.; Xiang, M.; Liu, X.; Liu, X. Echinochlamydosporium variabile, a new genus and species of Zygomycota from soil nematodes. Fungal Divers 2011, 46, 43–51. [Google Scholar] [CrossRef]
- Osorio, N.W.; Habte, M.V.; Peláez, J.D.L. Effectiveness of a rock phosphate solubilizing fungus to increase soil solution phosphate impaired by the soil phosphate sorption capacity. Rev. Fac. Nal. Agr. 2015, 68, 7627–7636. [Google Scholar] [CrossRef]
- Gomes, E.C.Q.; Godinho, V.M.; Silva, D.A.S.; de Paula, M.T.R.; Vitoreli, G.A.; Zani, C.L.; Alves, T.M.A.; Junior, P.A.S.; Murta, S.M.F.; Barbosa, E.C.; et al. Cultivable fungi present in Antarctic soils: Taxonomy, phylogeny, diversity, and bioprospecting of antiparasitic and herbicidal metabolites. Extremophiles 2018, 22, 381–393. [Google Scholar] [CrossRef]
- Shimizu, S.; Jareonkitmongkol, S. Mortierella species (fungi): Production of C20 polyunsaturated fatty acids. In Medicinal and Aromatic Plants VIII. Biotechnology in Agriculture and Forestry; Bajaj, Y.P.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1995; Volume 33, pp. 308–325. [Google Scholar] [CrossRef]
- Kulhman, E.G. Variation in zygospore formation among species of Mortierella. Mycologia 1972, 64, 325–341. [Google Scholar] [CrossRef]
- Ham, M.P.; Smith, M.D. Fluorine balance studies on four infants. J. Nutr. 1954, 53, 15–223. [Google Scholar] [CrossRef] [PubMed]
- Atlas, R.M. Handbook of Media for Environmental Microbiology; CRC Press: Boca Raton, FL, USA, 1995; pp. 32–33. [Google Scholar]
- Lechevalier, M.P.; Lechevalier, H.A. A new genus of Actinomycetales: Waksmania gen. nov. J. Gen. Microbiol. 1957, 17, 104–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, D.; Zeng, J.; Zheng, Y.; Yu, X.; Chen, S. Microbial lipid production from xylose by Mortierella isabellina. Bioresour. Technol. 2013, 133, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Mehrotra, B.S.; Baijal, U. Species of Mortierella from India-III. Mycopathol. Mycol. Appl. 1963, 20, 49–54. [Google Scholar] [CrossRef]
- Tsao, P.H.; Guy, S.O. Inhibition of Mortierella and Pythium in a Phytophtora-isolation medium containing hymexazol. Phytopathology 1977, 67, 796–801. [Google Scholar] [CrossRef]
- Hýsek, J.; Brožová, J. The changes of soil microfungal spectrum simultaneous agricultural and forestry cultivation. Silva Gabreta 2001, 7, 185–190. [Google Scholar]
- Swer, H.; Dkhar, M.S.; Kayang, H. Fungal population and diversity in organically amended agricultural soils of Meghalaya, India. J. Org. Sys. 2011, 6, 3–12. [Google Scholar]
- Wagner, L.; Stielow, B.; Hoffmann, K.; Petkovits, T.; Papp, T.; Vágvölgyi, C.; de Hoog, G.S.; Verkley, G.; Voigt, K. A comprehensive molecular phylogeny of the Mortierellales (Mortierellomycotina) based on nuclear ribosomal DNA. Persoonia 2013, 30, 77–93. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.W.; Nguyen, T.T.T.; Lee, H.Y.M.H.; Kim, C.; Lee, H.B. Confirmation of two undescribed fungal species from Dokdo of Korea based on current classification system using multi loci. Mycobiology 2015, 43, 392–401. [Google Scholar] [CrossRef] [Green Version]
- Mares-Ponce de León, Y.; Muñoz-Castellanos, L.N.; Ruiz-Cisneros, M.F.; Pérez-Corral, D.A.; Ornelas-Paz, J.J.; Acosta-Muñiz, C.H.; Berlanga-Reyes, D.I.; Rios-Velasco, C. Morphological and molecular identification of Mortierella species associated to rhizosphere of apple trees with symptoms of root diseases. Mexican J. Phytopathol. 2017, 36, 184–195. [Google Scholar] [CrossRef] [Green Version]
- Silva, U.C.; Medeiros, J.D.; Leite, L.R.; Morais, D.K.; Cuadros-Orellana, S.; Oliveira, C.A.; de Paula Lana, U.G.; Gomes, E.A.; Dos Santos, V.L. Long-term rock phosphate fertilization impacts the microbial communities of maize rhizosphere. Front. Microbiol. 2017, 8, 1266. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Jing, T.; Chen, Y.; Wang, F.; Qi, D.; Feng, R.; Xie, J.; Li, H. Deciphering microbial diversity associated with Fusarium wilt-diseased and disease-free banana rhizosphere soil. BMC Microbiol. 2019, 19, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, X.; Xiao, S.; Zhao, Y.; Xu, Y.; Yang, H.; Zhang, L. Comparative analysis of rhizosphere soil physiochemical characteristics and microbial communities between rusty and healthy ginseng root. Sci. Rep. 2020, 10, 15756. [Google Scholar] [CrossRef]
- Hanaka, A.; Ozimek, E.; Majewska, M.; Rysiak, A.; Jaroszuk-Ściseł, J. Physiological diversity of Spitsbergen soil microbial communities suggests their potential as plant growth-promoting bacteria. Int. J. Mol. Sci. 2019, 20, 1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Scholl, L.; Kuyper, T.W.; Smits, M.M.; Landeweert, R.; Hoffland, E.; van Breemen, N. Rock-eating mycorrhizas: Their role in plant nutrition and biogeochemical cycles. Plant Soil 2008, 303, 35–47. [Google Scholar] [CrossRef]
- Daghino, S.; Martino, E.; Vurro, E.; Tomatis, M.; Girlanda, M.; Fubini, B.; Perotto, S. Bioweathering of chrysotile by fungi isolated in ophiolitic sites. FEMS Microbiol. Lett. 2008, 285, 242–249. [Google Scholar] [CrossRef] [Green Version]
- Gadd, G.M. Geomycology: Biogeochemical transformations of rocks, minerals, metals and radionuclides by fungi, bioweathering and bioremediation. Mycol. Res. 2007, 111, 3–49. [Google Scholar] [CrossRef]
- Gadd, G.M. Metal transformations. In Fungi in Bioremediation; Gadd, G.M., Ed.; Cambridge University Press: Cambridge, UK, 2001; pp. 359–383. [Google Scholar]
- Font, E.; Veiga-Pires, C.M.; Pozo, C.; Carvallo, A.C.; de Siqueira Neto, P.; Camps, S.; Fabre, S.; Mirão, J. Magnetic fingerprint of southern Portuguese speleothems and implications for paleomagnetism and environmental magnetism. J. Geophys. Res. Solid Earth 2014, 119, 7993–8020. [Google Scholar] [CrossRef]
- Widden, P.; Parkinson, D. Populations of fungi in a high arctic ecosystem. Can. J. Bot. 1979, 57, 2408–2417. [Google Scholar] [CrossRef]
- Kurek, E.; Korniłłowicz-Kowalska, T.; Słomak, A.; Melke, J. Characteristics of soil filamentous fungi communities isolated from various micro−relief forms in the high Arctic tundra (Bellsund region, Spitsbergen). Polish Polar Res. 2007, 28, 57–73. [Google Scholar]
- Pugh, G.J.F.; Allsopp, D. Microfungi on Signy Island, South Orkney Islands. Br. Antarct. Surv. Bull. 1982, 57, 55–67. [Google Scholar]
- Schmidt, S.K.; Wilson, K.L.; Meyer, A.F.; Gebauer, M.M.; King, A.J. Phylogeny of ecophysiology of opportunistic “snow molds” from a subalpine forest ecosystem. Microb. Ecol. 2008, 56, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, S.; Hu, B.; Lin, L.; Zhang, Q. RNA-sequence analysis reveals the cold adaption mechanism of Mortierella isabellina M6-22. Int. J. Agric. Biol. 2018, 20, 415–421. [Google Scholar] [CrossRef]
- Werner, S.; Peršoh, D.; Rambold, G. New aspects of the biology of Mortierella alliacea. Mycol. Progress 2016, 15, 1293–1301. [Google Scholar] [CrossRef]
- Fröhlich-Nowoisky, J.; Hill, T.C.J.; Pummer, B.G.; Yordanova, P.; Franc, G.D.; Pöschl, U. Ice nucleation activity in the widespread soil fungus Mortierella alpina. Biogeosciences 2015, 12, 1057–1071. [Google Scholar] [CrossRef] [Green Version]
- Bergero, R.; Girlanda, M.; Varese, G.; Intili, D.; Luppi, A.M. Psychrooligotrophic fungi from Arctic soils of Franz Joseph Land. Polar Biol. 1999, 21, 361–368. [Google Scholar] [CrossRef]
- Lee, J.-S.; Nam, B.; Lee, H.B.; Choi, Y.-J. Molecular phylogeny and morphology reveal the underestimated diversity of Mortierella (Mortierellales) in Korea. Kor. J. Mycol. 2018, 46, 375–382. [Google Scholar] [CrossRef]
- Johnson, J.M.; Ludwig, A.; Furch, A.C.U.; Mithöfer, A.; Scholz, S.; Reichelt, M.; Oelmüller, R. The beneficial root-colonizing fungus Mortierella hyalina promotes the aerial growth of Arabidopsis and activates calcium-dependent responses that restrict Alternaria brassicae-induced disease development in roots. Mol. Plant Microbe Interact. 2019, 32, 351–363. [Google Scholar] [CrossRef] [Green Version]
- Lindahl, B.D.; Ihrmark, K.; Boberg, J.; Trumbore, S.E.; Högberg, P.; Stenlid, J.; Finlay, F.D. Spatial separation of litter decomposition and mycorrhizal nitrogen uptake in a boreal forest. N. Phytol. 2007, 173, 611–620. [Google Scholar] [CrossRef] [Green Version]
- Mäkipää, R.; Rajala, T.; Schigel, D.; Rinne, K.T.; Pennanen, T.; Abrego, N.; Ovaskainen, O. Interactions between soil- and dead wood-inhabiting fungal communities during the decay of Norway spruce logs. ISME J. 2017, 11, 1964–1974. [Google Scholar] [CrossRef] [Green Version]
- Gawas-Sakhalkar, P.; Singh, S.M. Fungal community associated with Arctic moss, Tetraplodon mimoides and its rhizosphere: Bioprospecting for production of industrially useful enzymes. Curr. Sci. 2011, 100, 1701–1705. [Google Scholar]
- Varnaitė, R.; Paškevičius, A.; Raudonienė, V. Cellulose degradation in rye straw by micromycetes and their complexes. Ekologija 2008, 54, 29–31. [Google Scholar] [CrossRef]
- Jackson, R.M. Studies of fungi in pasture soils. III. Physiological studies on some fungal isolates from the root surface and from organic debris. N. Z. J. Agric. Res. 1965, 8, 878–888. [Google Scholar] [CrossRef] [Green Version]
- De Tender, C.; Mesuere, B.; Van der Jeugt, F.; Haegeman, A.; Ruttink, T.; Vandecasteele, B.; Dawyndt, P.; Debode, J.; Kuramae, E.E. Peat substrate amended with chitin modulates the N-cycle, siderophore and chitinase responses in the lettuce rhizobiome. Sci. Rep. 2019, 9, 9890. [Google Scholar] [CrossRef]
- Tanaka, Y.; Aki, T.; Hidaka, Y.; Furuya, Y.; Kawamoto, S.; Shigeta, S.; Ono, K.; Suzuki, O. Purification and characterization of a novel fungal α-glucosidase from Mortierella alliacea with high starch-hydrolytic activity. Biosci. Biotech. Bioch. 2002, 66, 2415–2423. [Google Scholar] [CrossRef]
- Cao, G.; Guan, Z.; Liu, F.G.; Liao, X.; Cai, Y. Arachidonic acid production by Mortierella alpina using raw crop materials. Acta Sci. Pol. Technol. Aliment. 2015, 14, 133–143. [Google Scholar] [CrossRef]
- Naziya, B.; Murali, M.; Amruthesh, K.N. Plant growth-promoting fungi (PGPF) instigate plant growth and induce disease resistance in Capsicum annuum L. upon infection with Colletotrichum capsici (Syd.) Butler & Bisby. Biomolecules 2020, 10, 41. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.M.; Sultana, F. Application and mechanisms of plant growth promoting fungi (PGPF) for phytostimulation. In Organic Agriculture; Das, S.K., Ed.; IntechOpen Limited: London, UK, 2020; pp. 1–30. [Google Scholar] [CrossRef]
- Jahagirdar, S.; Kambrekar, D.N.; Navi, S.S.; Kunta, M. Plant growth-promoting fungi: Diversity and classification. In Bioactive Molecules in Plant Defense Signaling in Growth and Stress; Jogaiah, S., Abdelrahman, M., Eds.; Springer: Cham, Switzerland, 2019; pp. 25–34. [Google Scholar]
- Frąc, M.; Lipiec, J.; Usowicz, B.; Oszust, K.; Brzezińska, M. Structural and functional microbial diversity of sandy soil under cropland and grassland. Peer J. 2020, 8, e9501. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, M.; Zhang, J.; Hu, Y.; Dunjiang, C.; Guo, J.; Wu, D.; Sun, G. Soil physicochemical properties and the rhizosphere soil fungal community in a mulberry (Morus alba L.)/alfalfa (Medicago sativa L.) intercropping system. Forests 2019, 10, 167. [Google Scholar] [CrossRef] [Green Version]
- Osorio, N.W.; Habte, M. Synergistic influence of an arbuscular mycorrhizal fungus and a P solubilizing fungus on growth and P uptake of Leucaena leucephala in an oxisol. Arid Land Res. Manag. 2001, 15, 263–274. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, X.; Li, G.; Qi, P. Interactions between arbuscular mycorrhizal fungi and phosphate-solubilizing fungus (Mortierella sp.) and their effects on Kostelelzkya virginica growth and enzyme activities of rhizosphere and bulk soils at different salinities. Biol. Fertil. Soils 2011, 47, 543–554. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.S.; Li, G.; Qin, F.F.; Zhou, M.X.; Qin, P.; Pan, S.M. Castor bean growth and rhizosphere soil property response to different proportions of arbuscular mycorrhizal and phosphate-solubilizing fungi. Ecol. Res. 2014, 29, 181–190. [Google Scholar] [CrossRef]
- Tamayo-Velez, A.; Osorio, N.W. Co-inoculation with an arbuscular mycorrhizal fungus and a phosphate-solubilizing fungus promotes the plant growth and phosphate uptake of avocado plantlets in a nursery. Botany 2017, 95, 539–545. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Bonito, G.; Hsu, C.; Hameed, K.; Vilgalys, R.; Liao, H.-L. Mortierella elongata increases plant biomass among non-leguminous crop species. Agronomy 2020, 10, 754. [Google Scholar] [CrossRef]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial phosphorus solubilization and its potential for use in sustainable agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. Springer Plus 2013, 2, 587–600. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, R.; Yandigeri, M.S.; Kashyap, S.; Alagawadi, A.R. Effect of salt on survival and P-solubilization potential of phosphate solubilizing microorganisms from salt affected soils. Saudi J. Biol. Sci. 2012, 19, 427–434. [Google Scholar] [CrossRef] [Green Version]
- Whitelaw, M.A. Growth promotion of plants inoculated with phosphate solubilizing fungi. Adv. Agron. 2000, 69, 99–151. [Google Scholar] [CrossRef]
- Osorno, L.; Osorio, N.W. Effect of carbon and nitrogen source and concentration on rock phosphate dissolution induced by fungi. J. Appl. Biotechnol. 2014, 2, 32–42. [Google Scholar] [CrossRef]
- Osorio, N.W.; Habte, M. Phosphate desorption from the surface of soil mineral particles by a phosphate-solubilizing fungus. Biol. Fertil. Soils 2013, 49, 481–486. [Google Scholar] [CrossRef]
- Onweremadu, E.U. Predicting phosphorus sorption characteristics in highly weathered soils of South-Eastern Nigeria. Res. J. Environ. Sci. 2007, 1, 47–55. [Google Scholar]
- do Carmo Horta, M.; Torrent, J. Phosphorus desorption kinetics in relation to phosphorus forms and sorption properties of Portuguese acid soils. Soil Sci. 2007, 172, 631–638. [Google Scholar] [CrossRef] [Green Version]
- Osorno, L.; Osorio, N.W.; Habte, M. Phosphate desorption by a soil fungus in selected Hawaiian soils differing in their mineralogy. Trop. Agric. 2018, 95, 154–166. [Google Scholar]
- Devêvre, O.; Garbaye, J.; Botton, B. Release of complexing organic acids by rhizosphere fungi as a factor in Norway spruce yellowing in acidic soils. Mycol. Res. 1996, 100, 1367–1374. [Google Scholar] [CrossRef]
- Certini, G.; Corti, G.; Ugolini, F.C. Vertical trends of oxalate concentration in two soils under Abies alba from Tuscany (Italy). J. Plant Nutr. Soil Sci. 2000, 163, 173–177. [Google Scholar] [CrossRef]
- Ström, L.; Owen, A.G.; Godbold, D.L.; Jones, D.L. Organic acid mediated P mobilization in the rhizosphere and uptake by maize roots. Soil Biol. Biochem. 2002, 34, 703–710. [Google Scholar] [CrossRef]
- Anstoetz, M.; Rose, T.J.; Clark, M.W.; Yee, L.H.; Raymond, C.A.; Vancov, T. Novel applications for oxalate phosphate-amine metal-organic frameworks (OPA-MOFs): Can an iron based OPA-MOF be used as slow release fertilizer? PLoS ONE 2015, 10, e0144169. [Google Scholar] [CrossRef]
- Malusà, E.; Pinzari, F.; Canfora, L. Efficacy of biofertilizers: Challenges to improve crop production. In Microbial Inoculants in Sustainable Agricultural Productivity; Singh, D., Singh, H., Prabha, R., Eds.; Springer: New Delhi, India, 2016; pp. 17–40. [Google Scholar] [CrossRef]
- Ceci, A.; Pinzari, F.; Russo, F.; Maggi, O.; Persiani, A.M. Saprotrophic soil fungi to improve phosphorus solubilisation and release: In vitro abilities of several species. Ambio 2018, 47, 30–40. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Ruan, C.H.; Qin, P.; Seliskar, D.M.; Gallagher, J.L. Kosteletzkya virginica, a halophytic species with potential for agroecotechnology in Jiangsu Province. China Ecol. Engin. 2003, 21, 271–276. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Giovannin, L.; Palla, M.; Agnolucci, M.; Avio, L.; Sbrana, C.; Turrini, A.; Giovannetti, M. Arbuscular mycorrhizal fungi and associated microbiota as plant biostimulants: Research strategies for the selection of the best performing inocula. Agronomy 2020, 10, 106. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, E.; Holmstrom, S.J.M. Siderophores in environmental research: Roles and applications: Minireview. Microb. Biotechnol. 2014, 7, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Bozarth, R.F.; Goenaga, A. Purification and properties of mycoferritin from Mortierella alpina. Can. J. Microbiol. 1972, 18, 619–622. [Google Scholar] [CrossRef] [PubMed]
- Thicken, A.; Winkclmann, G. Rhizoferrin: A complexone type siderophore of the Mucorales and Entomophthorales (Zygomycetes). FEMS Microbiol. Lett. 1992, 94, 37–42. [Google Scholar] [CrossRef]
- Johnstone, T.C.; Nolan, E.M. Beyond iron: Non-classical biological functions of bacterial siderophores. Dalton Trans. 2015, 14, 6320–6339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, M.; Sarkar, S.; Sarkar, B.; Sharma, B.K.; Bhattacharjee, S.; Tribedi, P. Microbial siderophores and their potential applications: A review. Environ. Sci. Pollut. Res. Int. 2016, 23, 3984–3999. [Google Scholar] [CrossRef] [PubMed]
- Watteau, F.; Berthelin, J. Microbial dissolution of iron and aluminium from soil minerals: Efficiency and specificity of hydroxamate siderophores compared to aliphatic acids. Eur. J. Soil Biol. 1994, 30, 1–9. [Google Scholar]
- Hussein, K.A.; Joo, J.H. Zinc ions affect siderophore production by fungi isolated from the Panax ginseng rhizosphere. J. Microbiol. Biotechnol. 2019, 29, 105–113. [Google Scholar] [CrossRef]
- Davies, P.J. The plant hormones: Their nature, occurrence, and functions. In Plant Hormones; Davies, P.J., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 1–15. [Google Scholar] [CrossRef]
- Tudzynski, B.; Sharon, A. Biosynthesis, biological role and application of fungal phytohormones. In Industrial Applications. The Mycota (A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research); Osiewacz, H.D., Ed.; Springer: Berlin/Heidelberg, Germany, 2002; Volume 10, pp. 183–211. [Google Scholar] [CrossRef]
- Depuydt, S.; Hardtke, C.S. Hormone signalling crosstalk in plant growth regulation. Curr. Biol. 2011, 21, R365–R373. [Google Scholar] [CrossRef]
- Hanaka, A.; Nowak, A.; Plak, A.; Dresler, S.; Ozimek, E.; Jaroszuk-Ściseł, J.; Wójciak-Kosior, M.; Sowa, I. Bacterial isolate inhabiting Spitsbergen soil modifies the physiological response of Phaseolus coccineus in control conditions and under exogenous application of methyl jasmonate and copper excess. Int. J. Mol. Sci. 2019, 20, 1909. [Google Scholar] [CrossRef] [Green Version]
- Souza, R.; Ambrosini, A.; Passaglia, L.M. Plant growth-promoting bacteria as inoculants in agricultural soils. Genet. Mol. Biol. 2015, 38, 401–419. [Google Scholar] [CrossRef] [PubMed]
- Basse, C.W.; Lottspeich, F.; Steglich, W.; Kahmann, R. Two potential indole-3-acetalde-hyde dehydrogenases in the pythopathogenic fungus Ustilago maydis. Eur. J. Biochem. 1996, 242, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.; Riov, J.; Sharon, M. Indole-3-acetic acid biosynthesis in Colletotrichum gloeosporioides f. sp. aeschynomene. App. Environ. Microbiol. 1998, 64, 5030–5032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munné-Bosch, S.; Müller, M. Hormonal cross-talk in plant development and stress responses. Front. Plant Sci. 2013, 4, 529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Ruyter-Spira, C.; Bouwmeester, H. The interaction between strigolactones and other plant hormones in the regulation of plant development. Front. Plant Sci. 2013, 4, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, J.A.; Benková, E. Cytokinin cross-talking during biotic and abiotic stress responses. Front. Plant Sci. 2013, 4, 451. [Google Scholar] [CrossRef] [Green Version]
- Kuźniar, A.; Włodarczyk, K.; Wolińska, A. Agricultural and other biotechnological applications resulting from trophic plant-endophyte interactions. Agronomy 2019, 9, 779. [Google Scholar] [CrossRef] [Green Version]
- Marusig, D.; Tombesi, S. Abscisic acid mediates drought and salt stress responses in Vitis vinifera—A review. Int. J. Mol. Sci. 2020, 21, 8648. [Google Scholar] [CrossRef]
- Woźniak, M.; Gałązka, A.; Tyśkiewicz, R.; Jaroszuk-Ściseł, J. Endophytic bacteria potentially promote plant growth by synthesizing different metabolites and their phenotypic/physiological profiles in the Biolog GEN III MicroPlateTM Test. Int. J. Mol. Sci. 2019, 20, 5283. [Google Scholar] [CrossRef] [Green Version]
- Czeczuga, B.; Muszyńska, E.; Godlewska, A.; Mazalska, B. Aquatic fungi and fungus-like organisms growing on seeds of 131 plant taxa. Nova Hedwigia 2009, 89, 451–467. [Google Scholar] [CrossRef]
- Kasai, K.; Morinaga, T.; Horikoshi, T. Fungal succession in the early decomposition process of pine cones on the floor of Pinus densiflora forests. Mycoscience 1995, 36, 325–334. [Google Scholar] [CrossRef]
- Manzotti, A.; Bergna, A.; Burow, M.; Jørgensen, H.J.L.; Cernava, T.; Berg, G.; Collinge, D.B.; Jensen, B. Insights into the community structure and lifestyle of the fungal root endophytes of tomato by combining amplicon sequencing and isolation approaches with phytohormone profiling. FEMS Microbiol. Ecol. 2020, 96, fiaa052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansel, P.J.; Young, T.W.K. Association of Mortierella chlamydospora and Mortierella indohii with plants in culture. Microbios. Lett. 1982, 19, 193–244. [Google Scholar]
- Liao, H.L.; Bonito, G.; Rojas, J.A.; Hameed, K.; Wu, S.; Schadt, C.W.; Labbé, J.; Tuskan, G.A.; Martin, F.; Grigoriev, I.V.; et al. Fungal endophytes of Populus trichocarpa alter host phenotype, gene expression, and rhizobiome composition. Mol. Plant Microbe Interact. 2019, 32, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Perera, R.H.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Jones, E.B.G.; McKenzie, E.H.C.; Stadler, M.; Lee, H.B.; Samarakoon, M.C.; Ekanayaka, A.H.; Camporesi, E.; et al. Fungi on wild seeds and fruits. Mycosphere 2020, 11, 2108–2480. [Google Scholar] [CrossRef]
- Márquez, S.; Bills, G.F.; Domínguez Acuña, L.; Zabalgogeazcoa, I. Endophytic mycobiota of leaves and roots of the grass Holcus lanatus. Fungal Divers 2010, 41, 115–123. [Google Scholar] [CrossRef]
- Ferreira, A.; Rodrigues, M.; Fortuna, A.; Falcão, A.; Alves, G. Huperzine A from Huperzia serrata: A review of its sources, chemistry, pharmacology and toxicology. Phytochem. Rev. 2016, 15, 51–85. [Google Scholar] [CrossRef]
- Petrini, O.; Fisher, P.J.; Petrini, L.E. Fungal endophytes of bracken (Pteridium aquilinum), with some reflections on their use in biological control. Sydowia 1992, 44, 282–293. [Google Scholar]
- Yokoya, K.; Postel, S.; Fang, R.; Sarasan, V. Endophytic fungal diversity of Fragaria vesca, a crop wild relative of strawberry, along environmental gradients within a small geographical area. Peer J. 2017, 5, e2860. [Google Scholar] [CrossRef]
- Wani, Z.A.; Mirza, D.N.; Arora, P.; Riyaz-ul-Hassan, S. Molecular phylogeny, diversity, community structure, and plant growth promoting properties of fungal endophytes associated with corms of saffron plant: An insight into the microbiome of Crocus sativus Linn. Fungal Biol. 2016, 120, 1509–1524. [Google Scholar] [CrossRef]
- Eroshin, V.K.; Dedyukhina, E.G. Effect of lipids from Mortierella hygrophila on plant resistance to phytopathogens. World J. Microbiol. Biotechnol. 2002, 18, 165–167. [Google Scholar] [CrossRef]
- Xu, L.; Ravnskov, S.; Larsen, J.; Nicolaisen, M. Linking fungal communities in roots, rhizosphere, and soil to the health status of Pisum sativum FEMS Microbiol. Ecol. 2012, 82, 736–745. [Google Scholar] [CrossRef] [Green Version]
- Jaroszuk-Ściseł, J.; Kurek, E.; Słomka, A.; Janczarek, M.; Rodzik, B. Activities of cell wall degrading enzymes in autolyzing cultures of three Fusarium culmorum isolates: Growth-promoting, deleterious and pathogenic to rye (Secale cereale). Mycologia 2011, 103, 929–945. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Li, R.; Ren, Y.; Liu, C.Q.; Wu, H.; Jousset, A.; Shen, Q. Distinct roles for soil fungal and bacterial communities associated with the suppression of vanilla Fusarium wilt disease. Soil Biol. Bioch. 2017, 107, 198–207. [Google Scholar] [CrossRef]
- Yu, C.; Hu, X.; Deng, W.; Li, Y.; Han, G.; Ye, C. Soil fungal community comparison of different mulberry genotypes and the relationship with mulberry fruit sclerotiniosis. Sci. Rep. 2016, 6, 28365. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhang, S.; Wang, Y.; Li, Y.; Li, P.; Chen, L.; Jie, X.; Hu, D.; Feng, B.; Yue, K.; et al. Rare fungus, Mortierella capitata, promotes crop growth by stimulating primary metabolisms related genes and reshaping rhizosphere bacterial community. Soil Biol. Biochem. 2020, 151, 108017. [Google Scholar] [CrossRef]
- Sakuradani, E.; Ando, A.; Ogawa, J.; Shimizu, S. Improved production of various polyunsaturated fatty acids through filamentous fungus Mortierella alpina breeding. Appl. Microbiol. Biotechnol. 2009, 84, 1–10. [Google Scholar] [CrossRef]
- Trytek, M.; Fiedurek, J. A novel psychrotrophic fungus, Mortierella minutissima, for D-limonene biotransformation. Biotechnol. Lett. 2005, 27, 149–153. [Google Scholar] [CrossRef]
- Robinson, C.H. Cold adaptation in Arctic and Antarctic fungi. New Phytol. 2001, 151, 341–353. [Google Scholar] [CrossRef]
- Figueredo, H.M.; Gonçalves, V.N.; Godinho, V.M.; Lopes, D.V.; Oliveira, F.S.; Rosa, L.H. Diversity and ecology of cultivable fungi isolated from the thermal soil gradients in Deception Island, Antarctica. Extremophiles 2020, 24, 219–225. [Google Scholar] [CrossRef]
- Dyal, S.D.; Narine, S.S. Implications for the use of Mortierella fungi in the industrial production of essential fatty acids. Food Res. Int. 2005, 38, 445467. [Google Scholar] [CrossRef]
- Zlotek, U.; Wójcik, W. Effect of arachidonic acid elicitation on lettuce resistance towards Botrytis cinerea. Sci. Hortic. 2014, 179, 16–20. [Google Scholar] [CrossRef]
- Tagawa, M.; Tamaki, H.; Manome, A.; Koyama, O.; Kamagata, Y. Isolation and characterization of antagonistic fungi against potato scab pathogens from potato field soils. FEMS Microbiol. Lett. 2010, 305, 136–142. [Google Scholar] [CrossRef] [PubMed]

Figure 2.
Production of siderophores by Mortierella strains on the Chrome azurol S agar medium at three temperatures of incubation, 4, 9, and 15 °C (data not published). Arrangement of the strains on the plate: clockwise from top left—M. verticillata DEM32 (MV32); M. antarctica DEM7 (MA7); bottom from left—M. antarctica DEM4 (MA4); M. antarctica DEM10 (MA10) (the psychrotrophic strains isolated from the Spitzbergen soils).
Figure 2.
Production of siderophores by Mortierella strains on the Chrome azurol S agar medium at three temperatures of incubation, 4, 9, and 15 °C (data not published). Arrangement of the strains on the plate: clockwise from top left—M. verticillata DEM32 (MV32); M. antarctica DEM7 (MA7); bottom from left—M. antarctica DEM4 (MA4); M. antarctica DEM10 (MA10) (the psychrotrophic strains isolated from the Spitzbergen soils).


{kind=link}
{kind=link}
Table 1.
The Mortierella species identified in the agricultural soils.
| Mortierella Species | Crop/Cultivar | Country (Area) | Ref. |
|---|---|---|---|
| Mortierella sp. (1–5) strains; M. alpina; M. hyalina; M. isabellina; M. spinosa; M. ramanniana var. angulispora; M. ramanniana var. ramanniana | soil samples from avocado and citrus field | Nd | [28] |
| M. polycephala | wheat rhizosphere soil (Triticum aestivum cv. Baldus) | Netherlands (Utrecht) | [14] |
| Mortierella sp. | soil samples from potato | Czech Republic (Lipovà near Kašperské Hory) | [29] |
| M. elongata; M. gamsii; M. nanna; M. parvispora | soil samples from maize (Zea mays) | India (Meghalaya) | [30] |
| M. gamsii; M. nanna; M. parvispora | soil samples from French bean (Phaseolus vulgaris) | India (Meghalaya) | [30] |
| M. chienii | soil samples from bamboo grove | Taiwan (near Tainan) | [31] |
| M. polygoonia | soil samples from Solanum tuberosum | Netherlands (Wegeningen) | [32] |
| M. ambigua; M. indohii; M. zychae | soil samples from agricultural fields | Korea | [15] |
| M. elongata | soil samples from maize | China | [17] |
| M. alpina; M. gamsii; M. capitata; Mortierella sp. | soil samples from apple orchards (Malus domestica) | Mexico (Chihuahua) | [33] |
| Mortierella sp. | rhizosphere soil samples from maize and sorghum | Brazil (Minas Gerais) | [34] |
| Mortierella sp. | rhizosphere soil samples from Gossypium species— G. hirsutum cv. TM-1 G. barbadense cv. Hai 7124 | China (Shandong) | [13] |
| Mortierella sp. | soil samples from banana (Musa acuminata Cavendish cv. Brazil) farms | China (Hainan) | [35] |
| Mortierella sp. | soil samples from ginseng (Panax ginseng) farm | China (Jilin) | [36] |
nd—lack of data.
Table 2.
The effect of inoculation of agricultural plants with the plant growth-promoting Mortierella species.
Table 2.
The effect of inoculation of agricultural plants with the plant growth-promoting Mortierella species.
| Mortierella sp. (Co-Inoculants) | Origin of Mortierella sp. | Host Crops | Effect on Plant and Soil | Ref. |
|---|---|---|---|---|
| Mortierella sp. (G. aggregatum) | rhizospheric soil from mature L. leucocephala grown in Hawaii | leucaena (L. leucocephala) |
| [66] |
| Mortierella sp. (G. mosseae and/or G. aggregatum) | salt-affected coastal soil in China | Virginia saltmarsh mallow (Kosteletzkya virginica) as a potential seed crop for saline soils |
| [67] |
| Mortierella sp. (G. mosseae) | salt-affected coastal soil in China | castor bean (Ricinus communis cv. “Zi Bi”) |
| [68] |
| M. elongata | soil from 35-year experiment | corn (Zea mays) |
| [17] |
| Mortierella sp. strain HI-27 (Rhizoglomus fasciculatum) | rhizospheric soil from mature L. leucocephala grown in Hawaii | avocado (Persea americana cv. ‘Hass’) |
| [69] |
| M. alpina CS10E4 | C. sativus grown in India | saffron crocus (Crocus sativus) |
| [9] |
| M. elongata PMI 77 M. elongata PMI 94 M. elongata PMI 624 | Populus deltoides P. trichocarpa grown in the USA | watermelon (Citrullus lanatus) corn (Zea mays) tomato (Solanum lycopersicum) squash (Cucurbita) bahiagrass (Paspalum notatum) |
| [70] |
DW—dry weight; PR—phosphate rock; Piav—bioavailable P; IAA—indole-3-acetic acid; ABA—abscisic acid; JA—jasmonic acid.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ozimek, E.; Hanaka, A. Mortierella Species as the Plant Growth-Promoting Fungi Present in the Agricultural Soils. Agriculture 2021, 11, 7. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11010007
AMA Style
Ozimek E, Hanaka A. Mortierella Species as the Plant Growth-Promoting Fungi Present in the Agricultural Soils. Agriculture. 2021; 11(1):7. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11010007
Chicago/Turabian StyleOzimek, Ewa, and Agnieszka Hanaka. 2021. "Mortierella Species as the Plant Growth-Promoting Fungi Present in the Agricultural Soils" Agriculture 11, no. 1: 7. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11010007
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.