1. Introduction
The olive is one of the most ancient fruit tree species, and is traditionally cultivated in the Mediterranean basin with great socioeconomic impact for the countries in the region. Since 2000, there has been a remarkable rapid expansion of olive cultivation in countries such as Australia, China, Argentina, Chile and New Zealand. Therefore, great numbers of new olive trees are required for new olive orchards and for the replacement of the existing traditional ones.
Asexual propagation by leafy cuttings is the most efficient and economical method of olive propagation [
1]. However, leafy cuttings of different olive cultivars exhibit differences in their rooting ability [
2]. Therefore, they have been categorized into three groups, easy (100−66% rooting), moderate (66−33% rooting) and hard-to-root (33−0% rooting) [
3]. The differences in genotypes, the kind of hormone applied and its concentration, the rooting medium, the nutritional status of mother plants, and the phenological stage are some of the possible factors that affect the rooting ability in olive cuttings [
4,
5,
6,
7]. Furthermore, endogenous factors such as polyamines and phenolic compounds, as well as the interactions between them, may influence the primary triggers for root initiation [
8]. Cristofori et al. [
9] proposed that auxin in conjunction with phenolics, flavonoids and/or polyamines may increase the root promoting stimulus.
“Kalamata” is one of the most important Greek cultivars (cvs) worldwide, highly appreciated for the production of black table olives. The excellent fruit quality and the special morphological characteristics of the fruit and tree distinguish this cv from other table olive cvs and make it very desirable to customers. Unfortunately, leafy cuttings of Kalamata present a very low rooting ability (0−5%) and thus it is characterized as a hard-to-root cv [
3]. Earlier studies have shown that the exogenous applied auxins failed to promote rooting in Kalamata cuttings, or they had only a slight rooting effect [
3,
10].
Girdling of the shoot phloem is a horticultural practice that blocks the downward translocation of carbohydrates, hormones, and other possible root-promoting factors [
4,
11]. The resulting series of physiological changes, such as the accumulation of various organic substances that occur above the ring, affect the hormonal balance of the shoot, thus promoting root initiation [
12,
13]. The positive effect of girdling on root initiation has usually been attributed to the increase in carbohydrate content in the part of the shoot above the girdle which is used as the cutting source [
4,
12,
14]. However, the effect of girdling on carbohydrate concentration varies, depending on the size of branches, time of application, size of the girdle, as well as the kind of tree [
15].
Basak et al. [
11] found that girdling increased soluble sugar concentration and the rooting ability in mangrove trees. Additionally, girdling was found to be the most effective method for the rooting of cv. “Domat” olive cuttings [
16]. However, Usta [
16] only reported the effect of girdling on the rooting percentage of olive cuttings, without determining the concentrations and changes of any endogenous rooting co-factor during the early phases of rhizogenesis. On the other hand, Wilson [
17] did not confirm any positive effect of girdling on rooting of
Eucalyptus grandis Hill ex Maid. stem cuttings. That was also the case for the
Rhododendron arboreum girdled cuttings [
18]. Furthermore, the effect of girdling on endogenous polyamines and phenolic compounds and its relationship with root initiation has not been investigated in olive trees so far.
The objectives of this study were: (a) to assess the effect of shoot girdling of olive mother plants on the rooting potential of the leafy stem cuttings of the recalcitrant cv Kalamata; (b) to examine the changes in the concentration of biochemical factors in the cuttings’ bases induced by shoot girdling; and (c) to review the relationship between girdling, changes in biochemical factors, and the rooting potential of the cuttings.
4. Discussion
Based on the results of the PCA analysis, it was clear that girdling had a distinguished effect on both the rooting performance of the cuttings as well as on the endogenous rooting co-factors. Girdling significantly increased the rooting performance of Kalamata cuttings independently of the season it was performed. Similar results on the effect of girdling on rooting ability of olive cuttings were also found by Usta [
16]. Girdling has been found to promote rooting of semi-hardwood cuttings of
Artocarpus heterophyllus Lam. [
25], as well as of apple [
26] and of mangrove species [
11,
27]. A number of biochemical parameters, such as carbohydrates, phenolic compounds and PAs, affected the root induction of Kalamata cuttings. Girdling stimulated the substantial accumulation of these rooting cofactors above the girdle zone, as has been reported by other researchers [
11,
12], and thus significant effects were exhibited on the root initiation of Kalamata cuttings. Girdling also increased callus formation and resulted in a reduction in rotten cuttings, effects that also might favor root formation.
According to Biswas [
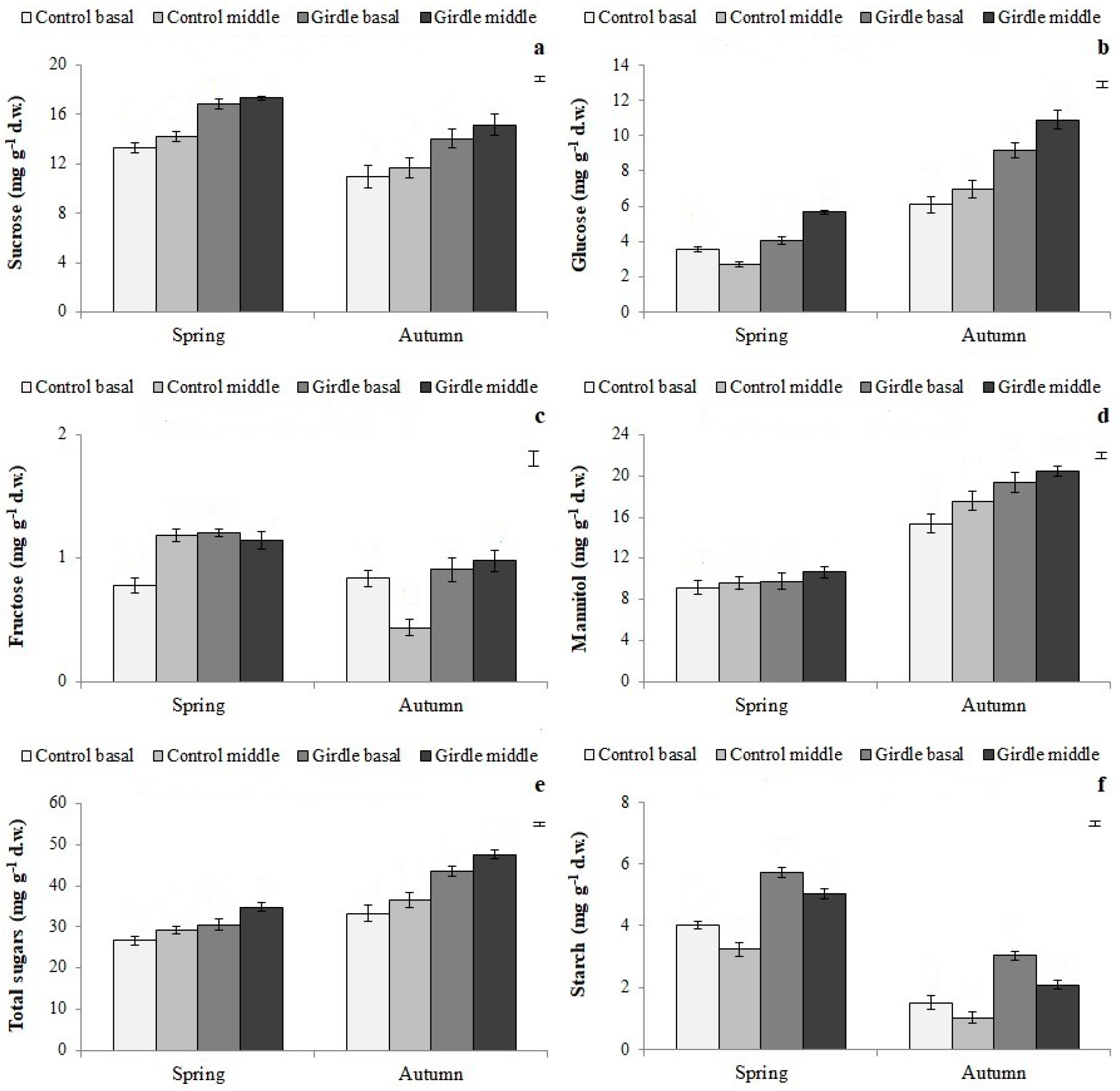
25], girdling blocks the basipetal movement of carbohydrates and other root-promoting factors and thus may positively affect root initiation. In the present study, high concentrations of soluble sugars, total sugars, and starch (
Figure 3) were found in the girdled Kalamata cuttings, in accordance with the literature [
11,
12,
13,
14,
28]. Furthermore, it is known that root formation relies on the utilization of soluble sugars [
11], because carbohydrates are used as energy sources during rhizogenesis [
1]. Carbohydrates can also improve the stimulatory effect of IBA by affecting auxin metabolism [
29], and are structural constituents of root primordia, acting as carbon skeleton donors [
30]. Thus, the increased carbohydrate concentration detected in the cuttings deriving from the girdled shoots might be partly responsible for the observed improvement of their rooting ability.
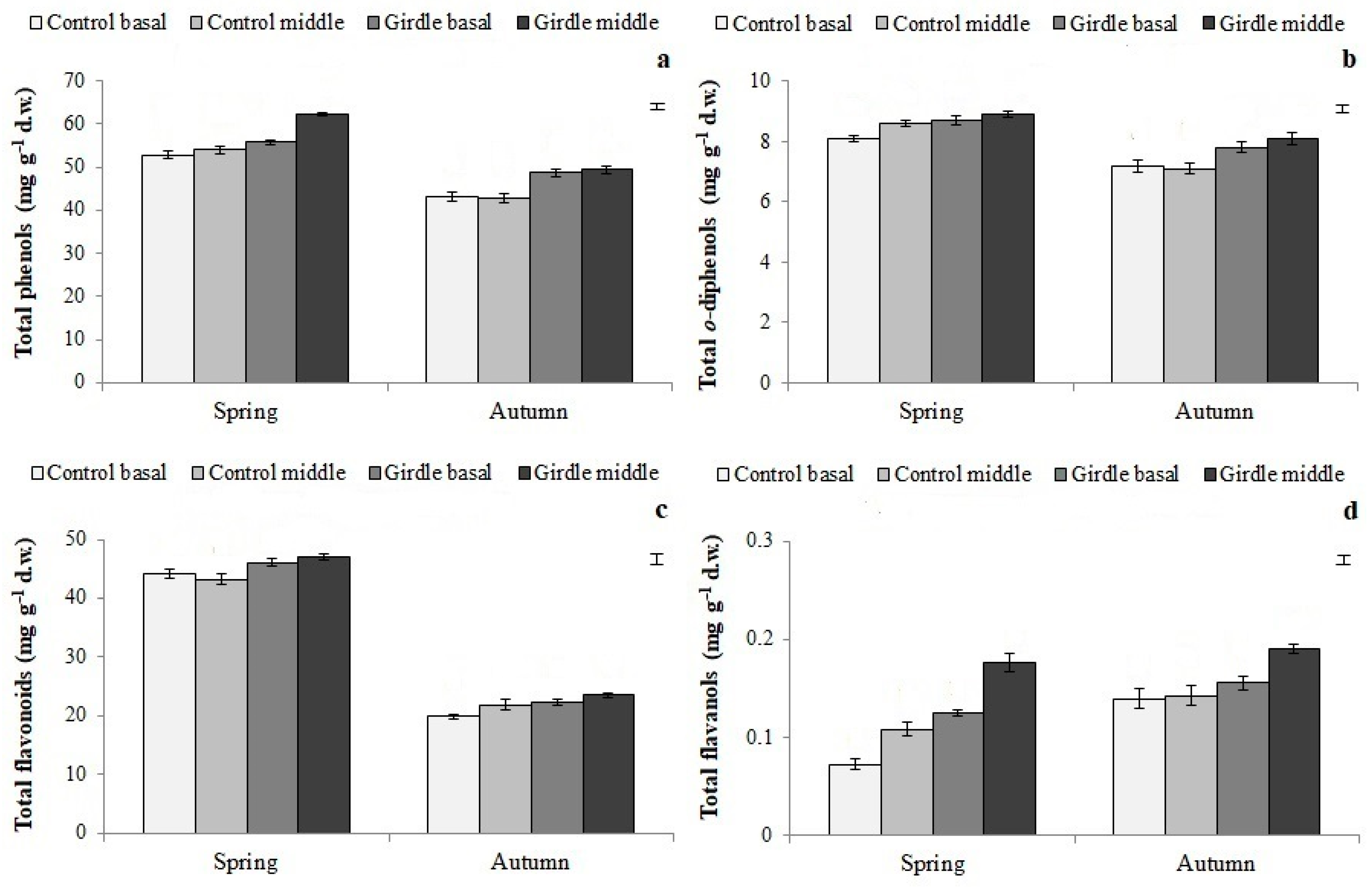
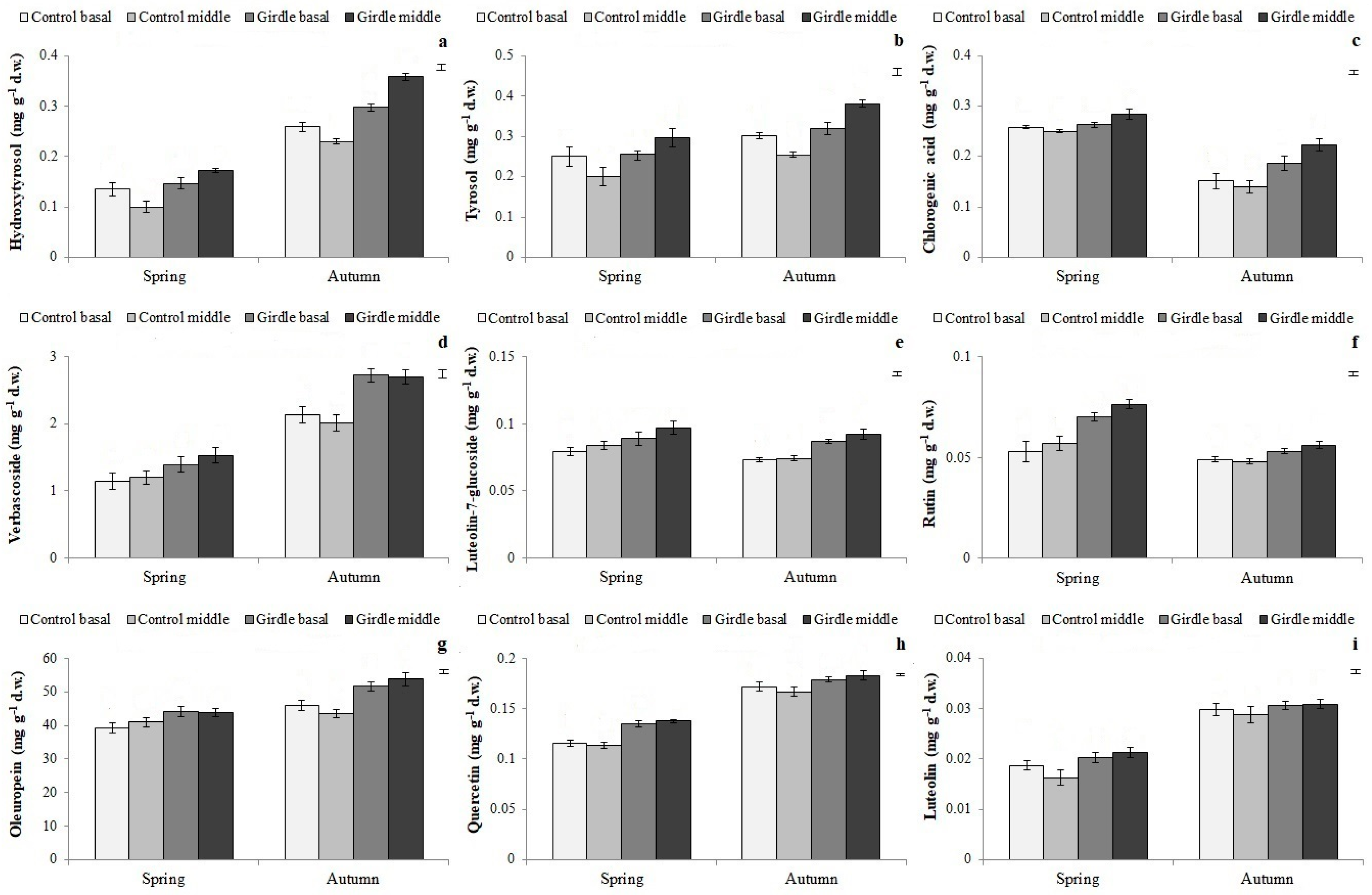
Apart from carbohydrates, girdling generally increased the concentration of total and individual phenolic compounds in both seasons (
Figure 6 and
Figure 7,
Table 4 and
Table 5). Kuokkanen et al. [
31], as well as Roussos et al. [
32], justified the accumulation of phenolic compounds in fruits from girdled scaffolds based on the hypothesis of growth differentiation balance. According to this theory, when carbohydrate biosynthesis exceeds the demand for growth, the excess carbon can be used for carbon-based secondary metabolites such as phenolic compounds [
31]. In the present study, girdling enhanced carbohydrate accumulation above the girdle zone; therefore, it can be assumed that part of the excess carbohydrates might have served as substrates for the biosynthesis of phenolic compounds. Furthermore, Kubota et al. [
33] and Khandaker et al. [
34] attributed the increase in the phenolic concentration of the girdled shoots to the enhancement of the activity of phenylalanine ammonia lyase, the key enzyme in phenolic compound biosynthesis, which might also be the case here.
Tyrosol, luteolin-7-glucoside and rutin, as well as total
o-diphenols, which are considered to act protectively against indole-3-acetic acid catabolism, protecting it from oxidation, during the induction and initiation phases [
24,
35,
36], were found, based on the PCA results (
Table 6), to be related with the increased rooting performance of the girdled cuttings. These results indicate the possible positive effect of specific phenolic compounds on the adventitious rooting process. There are not much data in the literature regarding the effect of girdling on phenolic compounds concentration; therefore, the present data offer new insights into the effects of girdling on secondary metabolism and the accumulation of rooting co-factors.
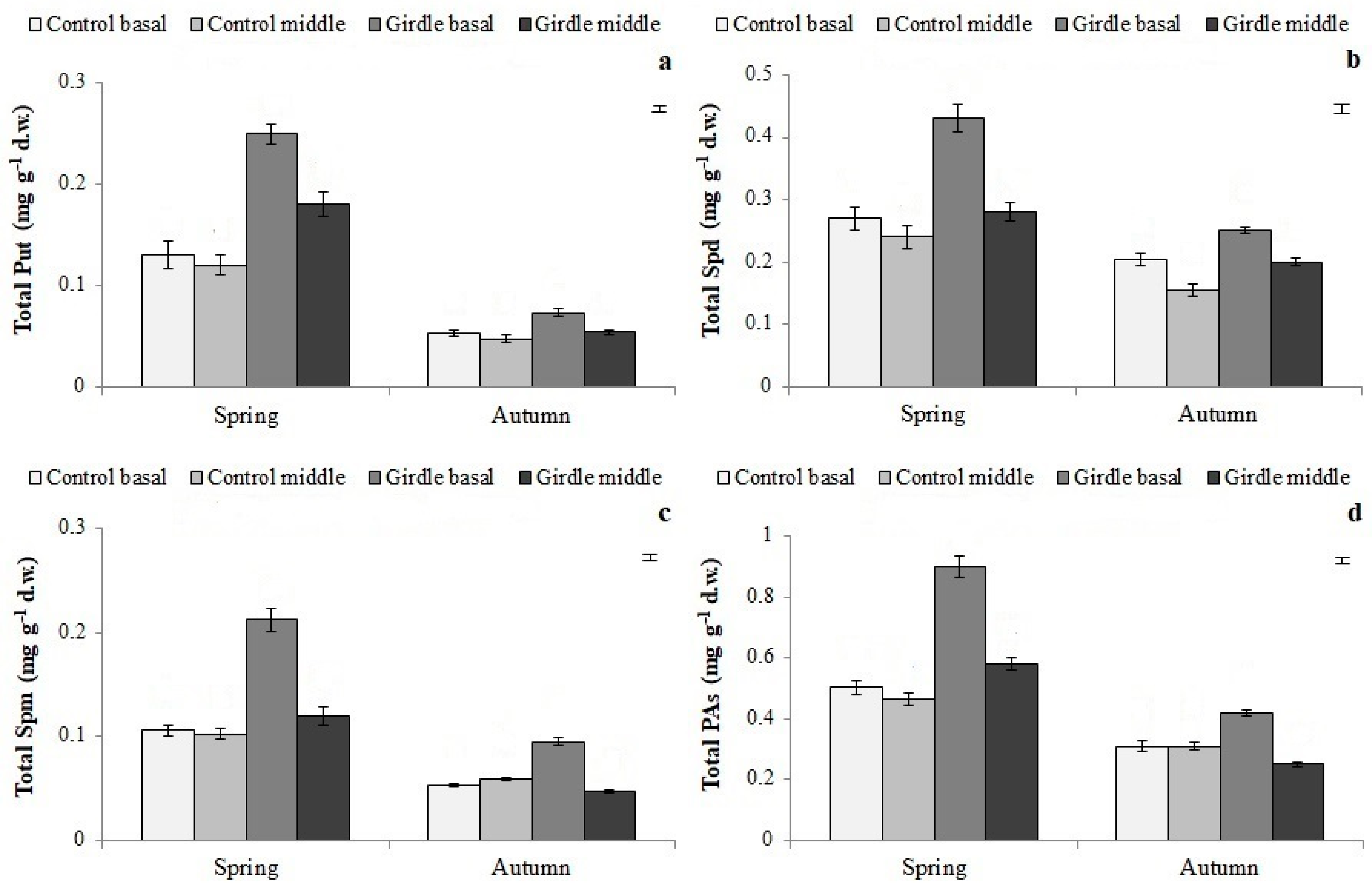
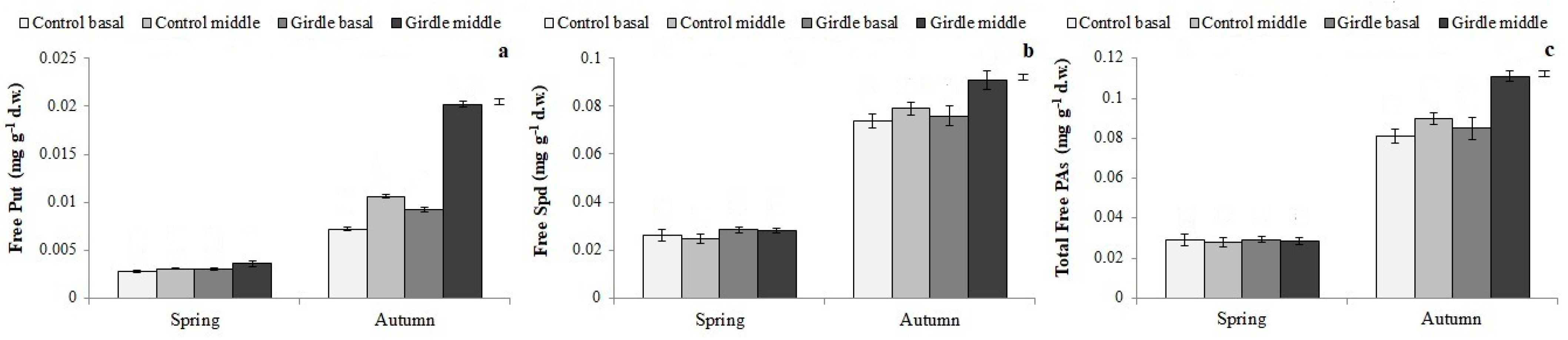
The concentration of PAs was also affected by girdling (
Figure 4 and
Figure 5,
Table 3). The increased level of free PAs, and especially of free Put, has been correlated with root induction [
37,
38], whereas bound PAs have been reported to have possible roles in rhizogenesis [
39], which is not yet fully clarified. Therefore, based on the rooting results of the present study, it can be assumed that the accumulation of PAs in the cuttings deriving from girdled shoots, especially in their free form, may have enhanced cell division and morphogenesis [
40,
41,
42], thus promoting cell differentiation into root primordia, under the effect of the exogenously applied auxin. Previous research in olive cuttings [
21,
43,
44] and other plant species [
9] also indicates that the increased concentration of endogenous PAs promotes rooting and improves root quality.
The type of the cutting (basal or middle part), as well as the time of cutting collection, may also influence the rooting potential of Kalamata cuttings (
Table 1 and
Table 2). Seasonal and even monthly or weekly changes in rooting performance have been reported in stone fruit rootstock cuttings [
45], in
Prunus cuttings [
18,
46], in
Lobostemon fruticosus [
47], and in many other species as well. These reports indicate that the rooting potential of the cuttings is significantly affected by the physiological state of the plant. According to Hartmann et al. [
4] and Tsafouros et al. [
45], the different ratio of endogenous root-promoting versus -inhibiting factors and/or the level of reserves changes with time, and therefore influences rooting performance.
Glucose, mannitol, and total sugars, independently of the cutting type, were higher in autumn than in spring, the period when the highest rooting percentage was recorded (
Figure 3). A seasonal variation in the rooting ability, which was associated with changes in carbohydrate concentrations in olive cuttings, was also found in the previous studies of Denaxa et al. [
19], Wiesman and Lavee [
29], and Del Rio et al. [
48], and in other plant species [
30]. These results strengthen the significant role of both the season and carbohydrate pool on the rooting potential of cuttings.
Polyamines were also affected by the season; the levels of free PAs, which have a key role in cell division [
40], were significantly higher in cuttings taken in autumn than in those in spring (
Figure 4 and
Figure 5). Furthermore, in autumn, the concentration of individual phenolic compounds, except for chlorogenic acid, rutin and luteolin-7-glucoside, was increased compared to the control (
Figure 7). Therefore, the increased rooting ability of autumn cuttings might be attributed to the increased endogenous concentrations of carbohydrates, free Pas, and specific phenolic compounds found at that time, compared to those of spring. The synergistic effect of the above rooting cofactors in the rooting potential of the stem cuttings has also been reported by Hartmann et al. [
4].
As far as the part of the mother shoot used as the cutting source is concerned, the present results indicated that the middle part of girdled cuttings exhibited the highest rooting percentage (
Table 1 and
Table 2). Similar results have been reported for a variety of plants, i.e.,
Lobostemon fruticosus [
47],
Santalum austrocaledonicum [
49], and the rootstocks GF 677, Rootpac-R and Cadaman [
45]. The concentration of the endogenous co-factors found in the present study may explain the different response of the two types of cuttings; the middle part was characterized by higher concentrations of sucrose, glucose, mannitol, free PAs, total phenolic compounds, luteolin-7-glucoside, and rutin (
Figure 3,
Figure 4,
Figure 5,
Figure 6 and
Figure 7). In agreement with the present results, Tombesi et al. [
50] reported that the part of the mother shoot used for cuttings affected their rooting ability and is closely related to carbohydrate accumulation, in particular to soluble sugar content, at the cutting’s base during the rooting process. This is also the case for PA, and phenolic compounds [
24,
36,
51], because their high endogenous content has been correlated with high rooting percentages in olive cuttings [
21] and in various other tree species [
52,
53].
Conclusively, the present study has shown that girdling significantly increased the rooting ability of olive cuttings by as much as 220% (from 5 to 16% in cuttings from the middle part, in autumn). Furthermore, girdling positively affected the concentration of soluble carbohydrates, free and total PAs, total phenolic compounds, as well as individual phenolic compounds. Nonetheless, further research is needed in order to draw conclusions on the overall efficiency of girdling on inducing olive cutting rooting, by using more hard-to-root olive cultivars as well as by performing girdling in different seasons. The accumulation of endogenous root-promoting substances above the girdled zone, such as carbohydrates, in combination with the increased concentrations of phenolic compounds and PAs, could be responsible for the recorded increase in the rooting potential of olive cuttings. However, according to Wiesman and Lavee [
3] the economic threshold for cutting propagation is estimated to be around 20% of successful rooting. Thus, more research is necessary in order to further improve the root induction of olive cv Kalamata cuttings.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}