
Physiological Screening for Drought Tolerance Traits in Vegetable Amaranth (Amaranthus tricolor) Germplasm

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
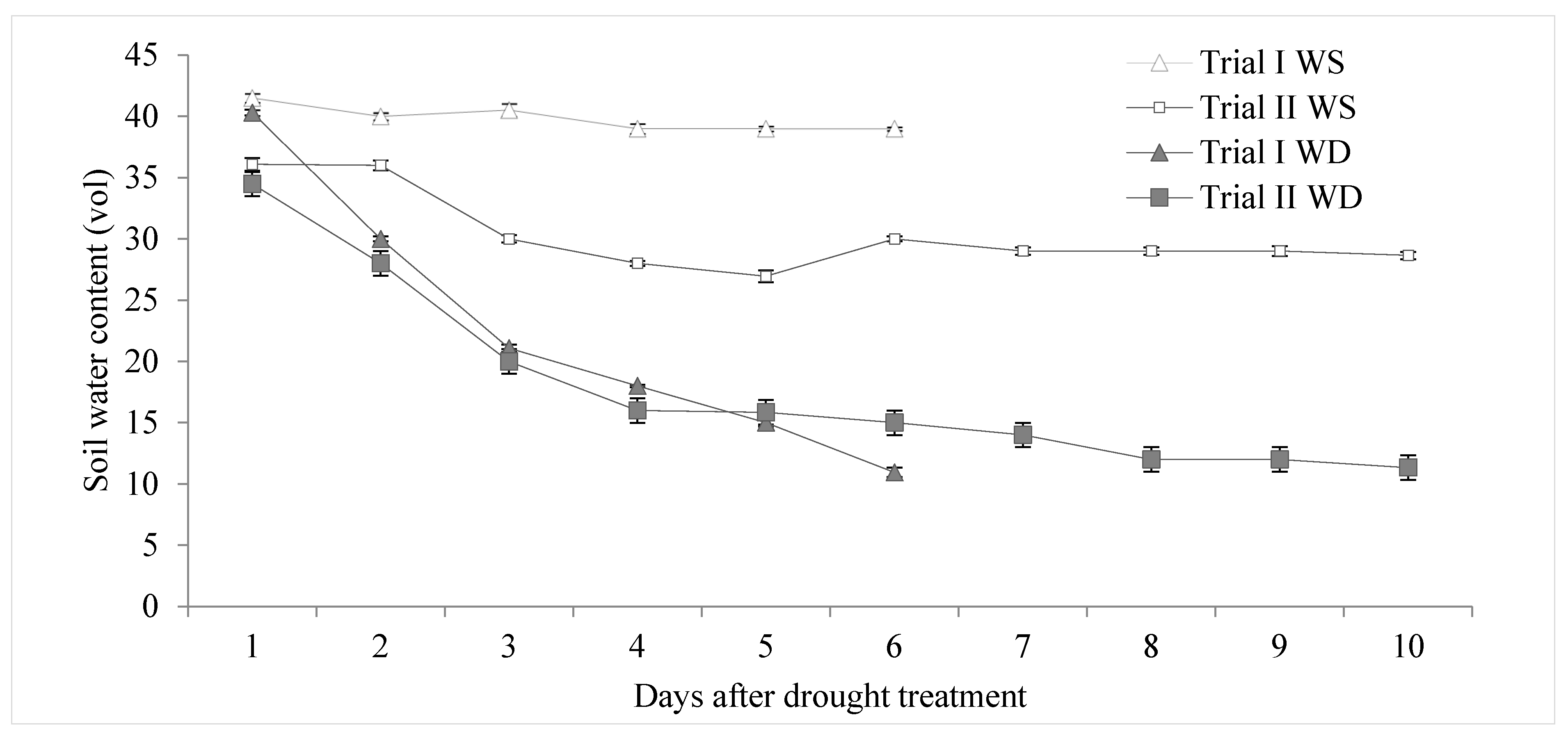
2.2. Water Treatments, Experimental Design and Growing Conditions
2.3. Plant Physiological Parameters
2.4. Total Yield and Leaf and Stem Biomass Partitioning
2.5. Rewatering Assessments
2.6. Drought Tolerance Indices
2.7. Statistical Analysis
3. Results
3.1. Combined Analysis of Water Treatment, Accession and Trial Environments on Growth and Plant Physiology
3.2. Genotypic Variations in Growth and Plant Physiology in Response to Drought Stress
3.2.1. Yield and Biomass Partitioning
3.2.2. Relative Water Content
3.2.3. Chlorophyll Fluorescence
3.2.4. Leaf Gas Exchange
3.3. Genotypic Variation in Drought Adaptability and Recovery
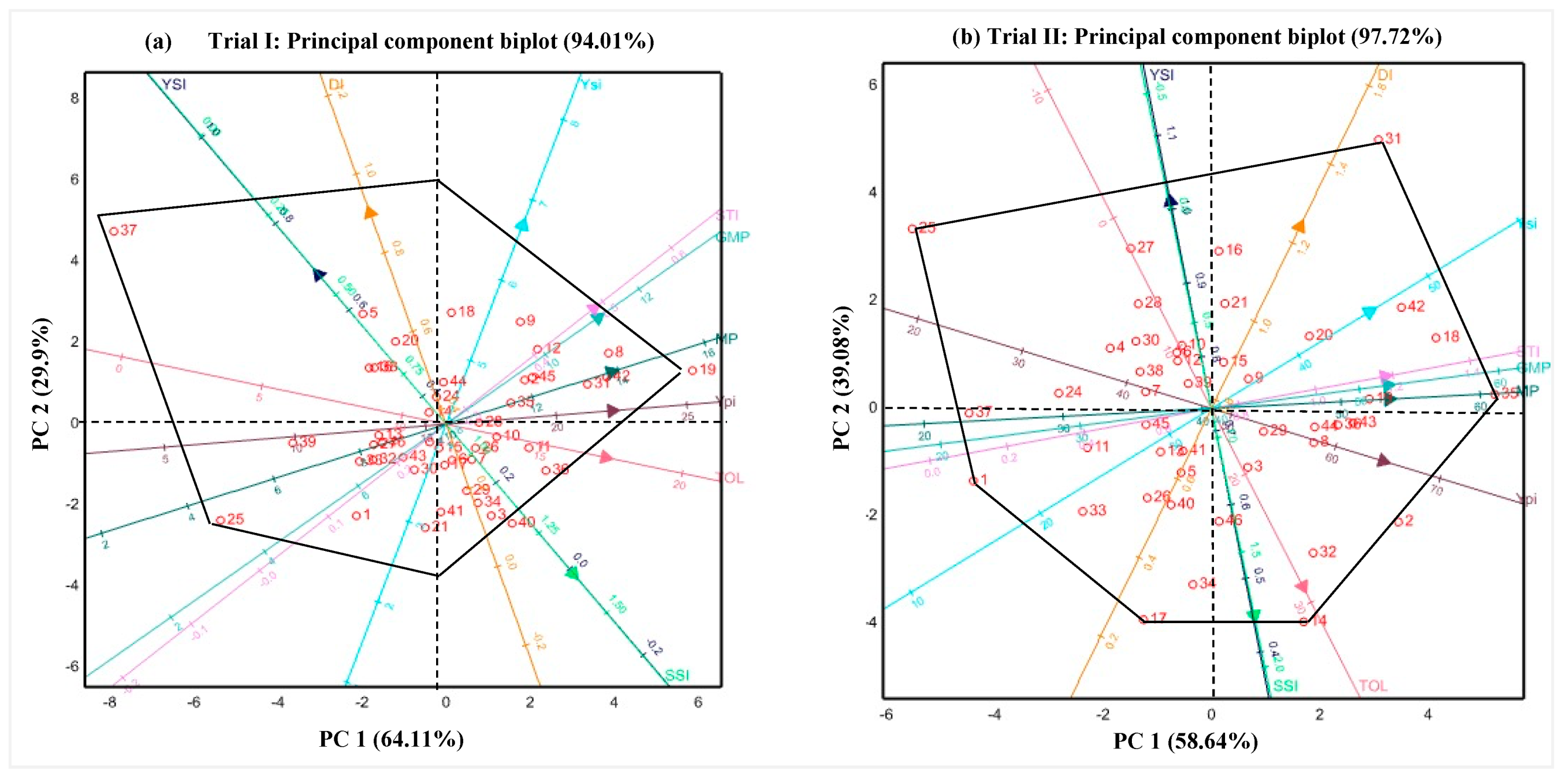
3.4. Interrelationship between Growth, Physiological Traits and Drought Adaptability
3.5. Selection of Drought Tolerance Indices
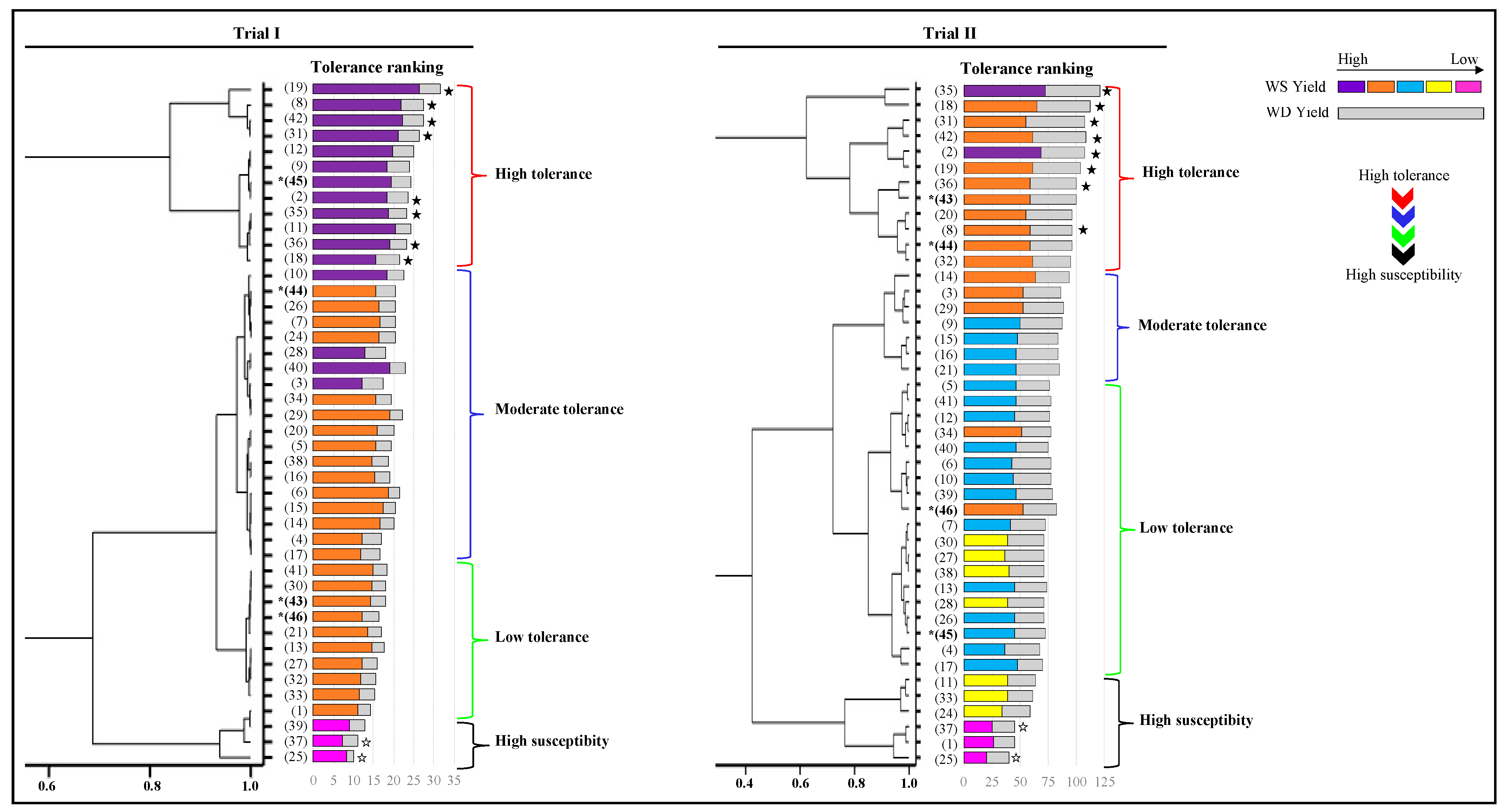
3.6. Drought Tolerance Ranking of 44 Amaranth Accessions
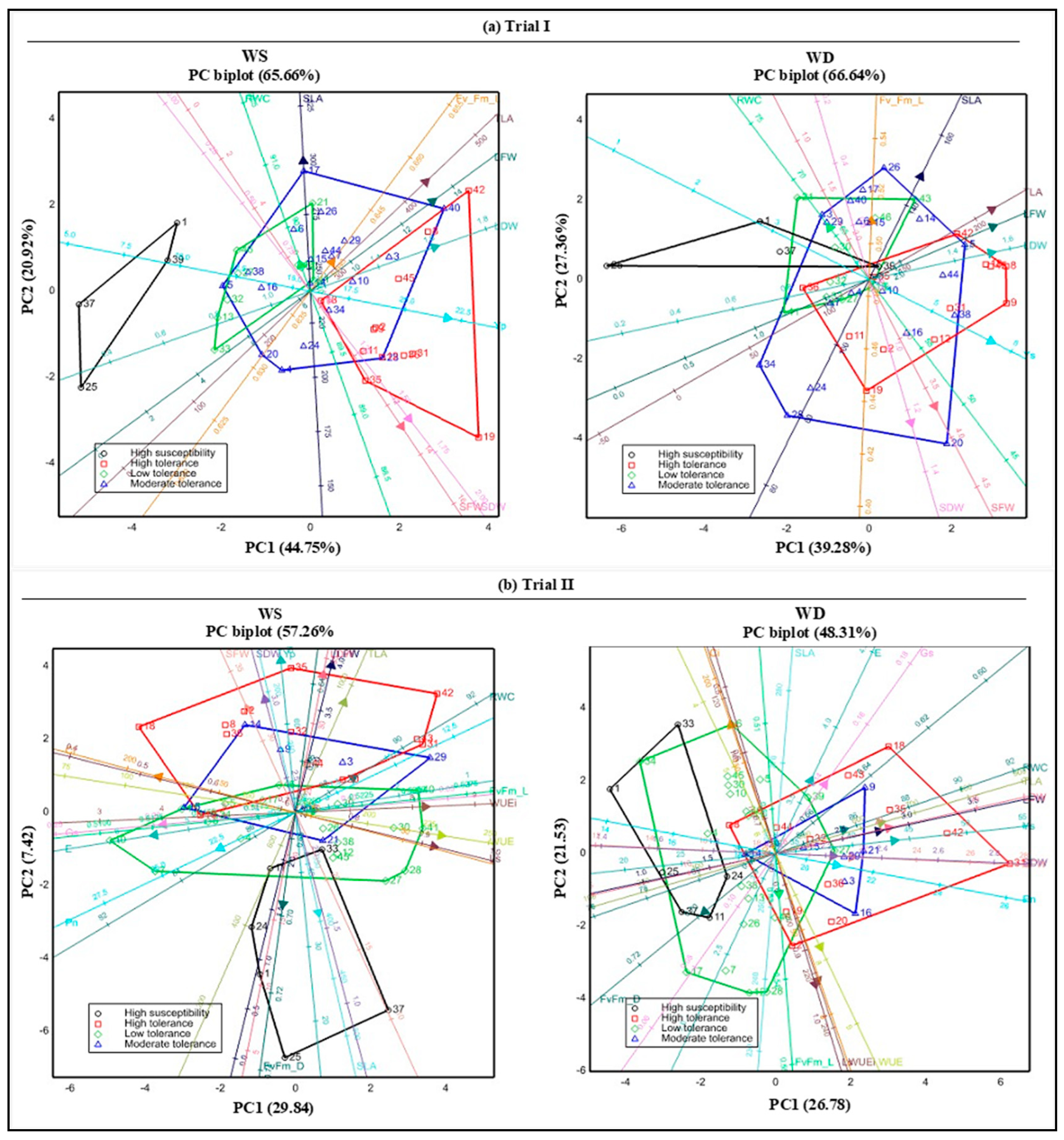
3.7. Association of Tolerance Grouping with Plant Growth and Physiology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Huang, J. Crop production under drought and heat stress: Plan responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [Green Version]
- Kogan, F.; Guo, W.; Yang, W. Drought and food security prediction from NOAA new generation of operational satellites. Geomat. Nat. Hazards Risk 2019, 10, 651–666. [Google Scholar] [CrossRef] [Green Version]
- Vermeulen, S.J.; Campbell, B.M.; Ingram, J.S.I. Climate change and food system. Annu. Rev. Environ. Resour. 2012, 37, 195–222. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.; Liu, Y.; Ishaq, M.; Shah, T.; Ilyas, A.; Din, I.U. Climate change and its impact on the yield of major food crops: Evidence from Pakistan. Foods 2017, 6, 39. [Google Scholar] [CrossRef] [PubMed]
- Twongyirwe, R.; Mfitumukiza, D.; Barasa, B.; Naggayi, B.R.; Odongo, H.; Nyakato, V.; Mutoni, G. Perceived effects of drought on household food security in South-western Uganda: Coping responses and determinants. Weather. Clim. Extrem. 2019, 24, 100201. [Google Scholar] [CrossRef]
- Chivenge, P.; Mabhaudhi, T.; Modi, A.T.; Mafongoya, P. The potential role of neglected and underutilised crop species as future crops under water scarce conditions in Sub-Saharan Africa. Int. J. Environ. Res. Public Health 2015, 12, 5685–5711. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, M.A.; Mayes, S.; Massawe, F. Crop diversification through a wider use of underutilised crops: A strategy to ensure food and nutrition security in the face of climate change. In Sustainable, Solutions for Food Security: Combating Climate Change by Adaptation; Sarkar, A., Sensarma, S.R., van Loon, G.W., Eds.; Springer Nature: Cham, Swizerland, 2019; pp. 125–149. [Google Scholar]
- Das, S. Systematics and taxonomic delimitation of vegetable, grain and weed amaranths: A morphological and biochemical approach. Genet. Resour. Crop Evol. 2012, 59, 289–303. [Google Scholar] [CrossRef]
- Jiménez-Aguilar, D.M.; Grusak, M.A. Minerals, vitamin C, phenolics, flavonoids and antioxidant activity of Amaranthus leafy vegetables. J. Food Compos. Anal. 2017, 58, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, A.; Shukla, S. Amaranth: A new millennium crop of nutraceutical values. Crit. Rev. Food Sci. Nutr. 2013, 53, 109–125. [Google Scholar] [CrossRef]
- Wu, X.; Blair, M.W. Diversity in grain amaranths and relatives distinguished by genotyping by sequencing (GBS). Front. Plant Sci. 2017, 8, 1960. [Google Scholar] [CrossRef] [Green Version]
- Jamalluddin, N.; Massawe, F.; Symonds, R.C. Transpiration efficiency of amaranth (Amaranthus sp.) in response to drought stress. J. Hortic. Sci. Biotechnol. 2018, 94, 448–459. [Google Scholar] [CrossRef] [Green Version]
- Stetter, M.G.; Zeitler, L.; Steinhaus, A.; Kroener, K.; Biljecki, M.; Schmid, K.J. Crossing methods and cultivation conditions for rapid production of segregating populations in three grain amaranth species. Front. Plant Sci. 2016, 7, 816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Stützel, H. Leaf water relations of vegetable amaranth (Amaranthus spp) in response to soil drying. Eur. J. Agron. 2002, 16, 137–150. [Google Scholar] [CrossRef]
- Liu, F.; Stützel, H. Leaf expansion, stomatal conductance and transpiration of vegetable amaranth (Amaranthus spp.) in response to soil drying. J. Am. Soc. Hortic. Sci. 2002, 127, 878–883. [Google Scholar] [CrossRef] [Green Version]
- Slabbert, R.; Krüger, G.H.J. Assessment of changes in photosystem II structure and function as affected by water deficit in Amaranthus hypochondriacus L. and Amaranthus hybridus L. Plant Physiol. Biochem. 2011, 49, 978–998. [Google Scholar] [CrossRef]
- Khanam, U.K.S.; Oba, S. Phenotypic plasticity of vegetable amaranth, Amaranthus tricolor L. under a natural climate. Plant Prod. Sci. 2014, 17, 166–172. [Google Scholar] [CrossRef] [Green Version]
- Kadereit, G.; Borsch, T.; Weising, K.; Freitag, H. Phylogeny of Amaranthaceae and Chenopodiaceae and the evolution of C4 photosynthesis. Int. J. Plant Sci. 2003, 164, 959–986. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Drought stress effects on growth, ROS markers, compatible solutes, phenolics, flavonoids, and antioxidant activity in Amaranthus tricolor. Appl. Biochem. Biotech. 2018, 186, 999–1016. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Drougt stress enhances nutritional bioactive compounds, phenolic acids and antioxidant capacity of Amaranthus leafy vegetable. BMC Plant Biol. 2018, 18, 258. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Stützel, H. Biomass partitioning, specific leaf area and water use efficiency of vegetable amaranth (Amaranthus spp.) in response to drought stress. Sci. Hortic. 2004, 102, 15–27. [Google Scholar] [CrossRef]
- Hura, T.; Grzesiak, S.; Hura, K.; Thiemt, E.; Tokarz, K.; Wedzony, M. Physiological and biochemical tools useful in drought tolerance detection in genotypes of winter triticale: Accumulation of ferulic acid correlates with drought tolerance. Ann. Bot. 2007, 100, 767–775. [Google Scholar] [CrossRef] [Green Version]
- Slabbert, R.; Krüger, G.H.J. Antioxidant enzyme activity, proline accumulation, leaf area and cell membrane stability in water stressed Amaranthus leaves. S. Afr. J. Bot. 2014, 95, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Luoh, J.W.; Begg, C.B.; Symonds, R.C.; Ledesma, D.; Yang, R. Nutritional yield of African indigenous vegetables in water-deficient and water-sufficient conditions. Food Nutr. Sci. 2014, 5, 812–822. [Google Scholar] [CrossRef] [Green Version]
- Babayev, H.; Mehvaliyeva, U.; Aliyeva, M.; Feyziyev, Y.; Guliyev, N. The study of NAD-malic enzyme in Amaranthus cruentus L. under drought. Plant Physiol. Biochem. 2014, 81, 84–89. [Google Scholar] [CrossRef]
- Jomo, O.M.; Netondo, G.W.; Musyimi, D.M. Drought inhibition of chlorophyll content among seven Amaranthus species. IARJSET 2016, 3, 1362–1371. [Google Scholar]
- Tsutsumi, N.; Tohya, M.; Nakashima, T.; Ueno, U. Variations in structural, biochemical, and physiological traits of photosynthesis and resource use efficiency in Amaranthus species (NAD-ME-type C4). Plant Prod. Sci. 2017, 20, 300–312. [Google Scholar] [CrossRef] [Green Version]
- Hura, T.; Hura, K.; Grzesiak, M.; Rzepka, A. Effect of long-term drought stress on leaf gas exchange and fluorescence parameters in C3 and C4 plants. Acta Physiol. Plant. 2007, 29, 103–113. [Google Scholar] [CrossRef]
- Johnson, D.A.; Asay, K.H. Viewpoint: Selection for improved response in cool-season grasses. J. Range Manag. 1993, 46, 194–202. [Google Scholar] [CrossRef]
- Rao, I.M. Role of physiology in improving crop adaptation to abiotic stresses in the tropics: The case of common bean and tropical forages. In Handbook of Plant and Crop Physiology, 2nd ed.; Pessarakli, M., Ed.; CRC Press: Boca Raton, FL, USA, 2001; p. 32. [Google Scholar]
- Hoyos-Villegas, V.; Song, Q.; Kelly, J.D. Genome-wide association analysis for drought tolerance and associated traits in common bean. Plant Genome 2017, 10, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, G.C.J. Effective selection criteria for assessing stress tolerance. In Proceedings of the International Symposium on Adaptation of Vegetables and Other, Food Crops in Temperature and Water Stress, Tainan, Taiwan, 13–18 August 1992; Kuo, C.G., Ed.; AVRDC: Shanhua, Taiwan, 1992; pp. 257–270. [Google Scholar]
- Mitra, J. Genetics and genetic improvement of drought resistance in crop plants. Curr. Sci. 2001, 80, 758–763. [Google Scholar]
- Farshadfar, E.; Sutka, J. Multivariate analysis of drought tolerance in wheat substitution lines. Cereal Res. Commun. 2002, 31, 33–39. [Google Scholar] [CrossRef]
- Andini, R.; Yoshida, S.; Yoshida, Y.; Ohsawa, R. Amaranthus genetic resources in Indonesia: Morphological and protein content assessment in comparison with worldwide amaranths. Genet. Resour. Crop Evol. 2013, 60, 2115–2128. [Google Scholar] [CrossRef] [Green Version]
- Sogbohossou, E.O.D.; Achigan-Dako, E.G. Phenetic differentiation and use type delimitation in Amaranthus spp. from worldwide origins. Sci. Hortic. 2014, 178, 31–42. [Google Scholar] [CrossRef]
- Sarker, U.; Islam, M.T.; Rabbani, M.G.; Oba, S. Genotypic variability for nutrient, antioxidant, yield and yield contributing traits in vegetable amaranth. J. Food Agric. Environ. 2014, 12, 168–174. [Google Scholar]
- Sarker, U.; Islam, M.T.; Rabbani, M.G.; Oba, S. Variability, heritability and genetic association in vegetable amaranth (Amaranthus tricolor L.). Span. J. Agric. Res. 2015, 13, 17. [Google Scholar] [CrossRef] [Green Version]
- Gerrano, D.A.; van Rensburg, W.S.J.; Mavengahama, S.; Bairu, M.; Venter, S.; Adebola, P. Qualitative morphological diversity of Amaranthus species. J. Trop. Agric. 2017, 55, 12–20. [Google Scholar]
- Thapa, R.; Blair, M. Morphological assessment of cultivated and wild amaranth species diversity. Agronomy 2018, 8, 272. [Google Scholar] [CrossRef] [Green Version]
- Gonzáles, L.; Gonzáles-Vilar, M. Determination of relative water content. In Handbook of Plant Ecophysiology Techniques; Reigosa, M.J., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001; pp. 207–212. [Google Scholar]
- Baker, N.R.; Rosenqvist, E. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities. J. Exp. Bot. 2004, 55, 1607–1621. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, L.; Shangguan, Z. Leaf gas exchange and fluorescence of two winter wheat varieties in response to drought stress and nitrogen supply. PLoS ONE 2016, 11, e0165733. [Google Scholar] [CrossRef] [Green Version]
- Yin, C.; Berninger, F.; Li, C. Photosynthetic responses of Populus przewalski subjected to drought stress. Photosynthetica 2006, 44, 62–68. [Google Scholar] [CrossRef]
- Fischer, R.A.; Maurer, R. Drought resistance in spring wheat cultivars. I. Grain yield response. Aust. J. Agric. Res. 1978, 29, 897–907. [Google Scholar] [CrossRef]
- Lan, J. Comparison of evaluating methods for agronomic drought resistance in crops. Acta Agric. Boreali-Occident. Sin. 1998, 7, 85–87. [Google Scholar]
- Hossain, A.B.S.; Sears, A.G.; Cox, T.S.; Paulsen, G.M. Desiccation tolerance and its relationship to assimilate partitioning in winter wheat. Crop Sci. 1990, 30, 622–627. [Google Scholar] [CrossRef]
- Bouslama, M.; Schapaugh, W.T. Stress tolerance in soybean. Part 1: Evaluation of three screening techniques for heat and drought tolerance. Crop Sci. 1984, 24, 933–937. [Google Scholar] [CrossRef]
- Kamoshita, A.; Babu, R.C.; Boopathi, N.M.; Fukai, S. Phenotypic and genotypic analysis of drought-resistance traits for development of rice cultivars adapted to rainfed environments. Field Crops Res. 2008, 109, 1–23. [Google Scholar] [CrossRef]
- Bahadur, A.; Chattarjee, A.; Kumar, R.; Singh, M.; Naik, P.S. Physiological and biochemical basis of drought tolerance in vegetables. Water 2011, 38, 1–16. [Google Scholar]
- Kumar, R.; Solankey, S.; Singh, M. Breeding for drought tolerance in vegetables. Veg. Sci. 2012, 39, 1–15. [Google Scholar]
- Bertolli, S.C.; Mazzafera, P.; Souza, G.M. Why is it so difficult to identify a single indicator of water stress in plants? A proposal for multivariate analysis to assess emergent properties. Plant Biol. 2014, 16, 578–585. [Google Scholar] [CrossRef]
- Gilbert, M.E.; Medina, V. Drought adaptation mechanisms should guide experimental design. Trends Plant Sci. 2016, 21, 639–647. [Google Scholar] [CrossRef] [Green Version]
- Hao, Z.F.; Li, X.H.; Su, Z.J.; Xie, C.X.; Li, M.S.; Liang, X.L.; Weng, J.F.; Zhang, D.G.; Li, L.; Zhang, S.H. A proposed selection criterion for drought resistance across multiple environments in maize. Breed. Sci. 2011, 61, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.B.; El-Sadek, A.N. Evaluation of drought tolerance indices for wheat (Triticum aestivum L.) under irrigated and rainfed conditions. Commun. Biometry Crop Sci. 2016, 11, 77–89. [Google Scholar]
- Ma, Q.Q.; Wang, W.; Li, Y.H.; Li, D.Q.; Zou, Q. Alleviation of photoinhibition in drought-stressed wheat (Triticum aestivum) by foliar-applied glycinebetaine. J. Plant Physiol. 2006, 163, 165–175. [Google Scholar] [CrossRef]
- Naya, L.; Ladrera, R.; Ramos, J.; González, E.M.; Arrese-Igor, C.; Minchin, F.R.; Becana, M. The response of carbon metabolism and antioxidant defences of alfalfa nodules to drought stress and to the subsequent recovery of plants. Plant Physiol. 2007, 144, 1104–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef]
- Iseki, K.; Homma, K.; Irie, T.; Endo, T.; Shiraiwa, T.; Iseki, K.; Shiraiwa, T. The long-term changes in midday photoinhibition in rice (Oryza sativa L.) growing under fluctuating soil water conditions. Plant Prod. Sci. 2013, 16, 287–294. [Google Scholar] [CrossRef]
- Uzildaya, B.; Turkana, I.; Sekmena, A.H.; Ozgura, R.; Karakayab, H.C. Comparison of ROS formation and antioxidant enzymes in Cleome gynandra (C4) and Cleome spinosa (C3) under drought stress. Plant Sci. 2012, 182, 59–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, A. Principal Estimates of Plant Water Status for Selection Work; AFRA Regional Training Course on Selection Methods for Drought Tolerance in Cereals and Legumes South Africa; ARC-Roodeplaat: Pretoria, South Africa, 1998. [Google Scholar]
- Vargas-Ortiz, E.; Ramírez-Tobias, H.M.; González-Escobar, J.L.; Gutiérrez-Garcia, A.K.; Bojórquez-Velázquez, E.; Espitia-Rangel, E.; de la Rosa, A.P.B. Biomass, chlorophyll fluorescence, and osmoregulation traits let differentiation of wild and cultivated Amaranthus under water stress. J. Photochem. Photobiol. B Biol. 2021, 220, 112210. [Google Scholar] [CrossRef] [PubMed]
- Zegaoui, Z.; Planchais, S.; Cabassa, C.; Djebbar, R.; Abrous, O.; Carol, P. Variation in relative water content, proline accumulation and stress gene expression in two cowpea landraces under drought. J. Plant Physiol. 2018, 218, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Bac-Molenaar, J.A.; Granier, C.; Keurentjes, J.J.B.; Vreugdenhil, D. Genome-wide association mapping of time-dependent growth responses to moderate drought stress in Arabidopsis: GWAS mapping of growth responses to moderate drought. Plant Cell Environ. 2016, 39, 88–102. [Google Scholar] [CrossRef] [Green Version]
- Rosales, M.A.; Cuellar-Ortiz, S.M.; Arrieta-Montiel, M.P.; Acosta-Gallegos, J.; Covarrubias, A.A. Physiological traits related to terminal drought resistance in common bean (Phaseolus vulgaris L.): Drought resistance-related traits in common bean. J. Sci. Food Agric. 2011, 93, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Soltys-Kalina, D.; Marczewski, W. The effect of drought stress on the leaf relative water content and tuber yield of a half-sib family of ‘Katahdin’-derived potato cultivars. Breed. Sci. 2016, 66, 328–331. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Wang, S.; Cao, B.; Cao, D.; Leng, G.; Li, H.; Yin, L.; Shan, L.; Deng, X. Genotypic variation in growth and physiological response to drought stress and re-watering reveals the critical role of recovery in drought adaptation in maize seedlings. Front. Plant Sci. 2016, 6, 1241. [Google Scholar] [CrossRef] [Green Version]
- Blum, A. Stress, strain, signalling, and adaptation—Not just a matter of definition. J. Exp. Bot. 2016, 67, 562–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, G.J.; Arndt, S.K. Osmotic Adjustment under drought conditions. In Plant Responses to Drought Stress; Aroca, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 199–229. [Google Scholar]
- Liu, C.C.; Liu, Y.G.; Guo, K.; Fan, D.Y. Effect of drought on pigments, osmotic adjustment and antioxidant enzymes in six woody plant species in karst habitats of southwestern China. Environ. Exp. Bot. 2011, 71, 174–183. [Google Scholar] [CrossRef]
- Zivcak, M.; Brestic, M.; Sytar, O. Osmotic adjustment and plant adaptation to drought stress. In Drought Stress Tolerance in Plants; Hossain, M., Wani, S., Bhattacharjee, S., Burritt, D., Tran, L.S., Eds.; Springer: Cham, Switzerland; Basel, Switzerland; Auckland, New Zealand, 2016; Volume 1, pp. 105–143. [Google Scholar]
- González-Rodríguez, T.; Cisneros-Hernández, I.; Acosta Bayona, J.; Ramírez-Chavez, E.; Martínez-Gallardo, N.; Mellado-Mojica, E.; López-Pérez, M.G.; Molina-Torres, J.; Délano-Frier, J. Identification of factors linked to higher water-deficit stress tolerance in Amaranthus hypochondriacus compared to other grain amaranths and A. hybridus, their ancestor. Plants 2019, 8, 239. [Google Scholar] [CrossRef] [Green Version]
- Malnoë, A.; Schultink, A.; Shahrasbi, S.; Rumeau, D.; Havaux, M.; Niyogi, K.K. The plastid lipocalin LCNP is required for sustained photoprotective energy dissipation in Arabidopsis. Plant Cell 2018, 30, 196–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, W.W., III; Zarter, C.R.; Ebbert, V.; Demmig-Adams, B. Photoprotective strategies of overwintering evergreens. Bioscience 2004, 4, 41–49. [Google Scholar] [CrossRef]
- Nemeskéri, E.; Helyes, J. Physiological responses of selected vegetable crop species to water stress. Agronomy 2019, 9, 447. [Google Scholar] [CrossRef] [Green Version]
- Li, R.H.; Guo, P.G.; Michael, B.; Stefania, G.; Salvatore, C. Evaluation of chlorophyll content and fluorescence parameters as indicators of drought tolerance in barley. Agric. Sci. China 2006, 5, 751–757. [Google Scholar] [CrossRef]
- Bahadur, A.; Kumar, R.; Mishra, U.; Rai, A.; Singh, M. Physiological approaches for screening of tomato genotypes for moisture stress tolerance. In Proceedings of the National Conference of Plant Physiology (NCPP-2010), Varanasi, India, 25–27 November 2010; p. 142. [Google Scholar]
- Miyake, C.; Amako, K.; Shiraishi, N.; Sugimoto, T. Acclimation of tobacco leaves to high light intensity drives the plastoquinone oxidation system-relationship among the fraction of open PSII centers, non-photochemical quenching of Chl fluorescence and the maximum quantum yield of PSII in the dark. Plant Cell Physiol. 2009, 50, 730–743. [Google Scholar] [CrossRef]
- Kohzuma, K.; Cruz, J.A.; Akashi, K.; Hoshiyasu, S.; Munekage, N.Y.; Yokota, A.; Kramer, D.M. The long-term responses of the photosynthetic proton circuit to drought. Plant Cell Environ. 2009, 32, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Bartoli, C.G.; Gomez, F.; Gergoff, G.; Guiamet, J.J.; Puntarulo, S. Up-regulation of the mitochondrial alternative oxidase pathway enhances photosynthetic electron transport under drought conditions. J. Exp. Bot. 2005, 56, 1269–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghate, T.; Deshpande, S.; Bhargava, S. Accumulation of stem sugar and its remobilisation in response to drought stress in a sweet sorghum genotype and its near-isogenic lines carrying different stay-green loci. Plant Biol. 2017, 19, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Perrier, L.; Rouan, L.; Jaffuel, S.; Clément-Vidal, A.; Roques, S.; Soutiras, A.; Baptiste, C.; Bastianelli, D.; Fabre, D.; Dubois, C.; et al. Plasticity of sorghum stem biomass accumulation in response to water deficit: A multiscale analysis from internode tissue to plant level. Front. Plant Sci. 2017, 8, 1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinheiro, C.; Chaves, M.M. Photosynthesis and drought: Can we make metabolic connections from available data? J. Exp. Bot. 2011, 62, 869–882. [Google Scholar] [CrossRef] [Green Version]
- Irving, L. Carbon assimilation, biomass partitioning and productivity in grasses. Agriculture 2015, 5, 1116–1134. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Ortiz, E.; Espitia-Rangel, E.; Tiessen, A.; Délano-Frier, J.P. Grain amaranths are defoliation tolerant crop species capable of utilizing stem and root carbohydrate reserves to sustain vegetative and reproductive growth after leaf loss. PLoS ONE 2013, 8, e67879. [Google Scholar] [CrossRef]
- Alfonso, S.U.; Brüggemann, W. Photosynthetic responses of a C3 and three C4 species of the genus Panicum (s.l.) with different metabolic subtypes to drought stress. Photosynth. Res. 2012, 112, 175–191. [Google Scholar] [CrossRef]
- Shabala, S.; White, R.G.; Djordjevic, M.A.; Ruan, Y.L.; Mathesius, U. Root-to-shoot signalling: Integration of diverse molecules, pathways and functions. Funct. Plant Biol. 2016, 43, 87–104. [Google Scholar] [CrossRef] [Green Version]
- Talebi, R.; Fayaz, F.; Naji, A.M. Effective selection criteria for assessing drought stress tolerance in durum wheat (Triticum durum Desf.). Gen. Appl. Plant Physiol. 2009, 35, 64–74. [Google Scholar]
- Panthuwan, G.S.; Fokai, M.; Cooper, S.; Rajatasereekul, S.O.; O’Toole, J.C. Yield response of rice genotypes to different types of drought under rainfed lowlands. Part 1: Grain yield and yield components. Field Crop Res. 2002, 41, 45–54. [Google Scholar]
- Gholipouri, A.; Sedghi, M.R.; Sharifi, S.; Nazari, N.M. Evaluation of drought tolerance indices and their relationship with grain yield in wheat cultivars. Recent Res. Sci. Technol. 2009, 1, 195–198. [Google Scholar]
- Anwar, J.; Subhani, G.M.; Hussain, M.; Ahmad, J.; Hussain, M.; Munir, M. Drought tolerance indices and their correlation with yield in exotic wheat genotypes. Pak. J. Bot. 2011, 43, 1527–1530. [Google Scholar]
- Tardieu, F. Plant response to environmental conditions: Assessing potential production, water demand, and negative effects of water deficit. Front. Physiol. 2013, 4, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entry | Accessions | Germplasm | ID | Origin Country |
|---|---|---|---|---|
| 1 | AV-TRI 2 | AVRDC | VI055356 | Bangladesh |
| 2 | AV-TRI 18 | AVRDC | VI044446 | India |
| 3 | AV-TRI 26 | AVRDC | VI049006 | Thailand |
| 4 | AV-TRI 33 | AVRDC | VI050610-A | Viet Nam |
| 5 | AV-TRI 34 | AVRDC | VI050609-A | Viet Nam |
| 6 | AV-TRI 39 | AVRDC | VI054572 | Philippines |
| 7 | AV-TRI 40 | AVRDC | VI054571 | Philippines |
| 8 | AV-TRI 44 | AVRDC | VI048286 | Bangladesh |
| 9 | AV-TRI 49 | AVRDC | VI047504 | Bangladesh |
| 10 | AV-TRI 51 | AVRDC | VI057270 | Cambodia |
| 11 | AV-TRI 53 | AVRDC | VI042979 | Indonesia |
| 12 | AV-TRI 54 | AVRDC | VI042978 | Indonesia |
| 13 | AV-TRI 56 | AVRDC | VI058498 | India |
| 14 | AV-TRI 57 | AVRDC | VI044426 | Malaysia |
| 15 | AV-TRI 58 | AVRDC | VI055139 | Malaysia |
| 16 | AV-TRI 68 | AVRDC | VI050111 | Taiwan |
| 17 | AV-TRI 69 | AVRDC | VI049431 | Taiwan |
| 18 | AV-TRI 3 | AVRDC | VI055353 | Bangladesh |
| 19 | AV-TRI 11 | AVRDC | VI047795 | Bangladesh |
| 20 | AV-TRI 24 | AVRDC | VI044396-A | Pakistan |
| 21 | AV-TRI 31 | AVRDC | VI050615-A | Viet Nam |
| 22 | AV-TRI 20 | AVRDC | VI043725 | Malaysia |
| 23 | AV-TRI 21 | AVRDC | VI043724 | Malaysia |
| 24 | US-TRI 3 | USDA | Ames 29505 | Brazil |
| 25 | US-TRI 6 | USDA | PI 478310 | China |
| 26 | US-TRI 13 | USDA | Ames 2039 | Indonesia |
| 27 | US-TRI 14 | USDA | Ames 5354 | Madagascar |
| 28 | US-TRI 15 | USDA | Ames 2029 | Malaysia |
| 29 | US-TRI 16 | USDA | Ames 29034 | Malaysia |
| 30 | US-TRI 19 | USDA | Ames 2199 | Taiwan |
| 31 | US-TRI 21 | USDA | PI 607446 | Thailand |
| 32 | US-TRI 24 | USDA | PI 632237 | USA |
| 33 | US-TRI 25 | USDA | Ames 5110 | West Africa |
| 34 | US-TRI 29 | USDA | Ames 26216 | China |
| 35 | US-TRI 39 | USDA | Ames 2132 | India |
| 36 | US-TRI 46 | USDA | Ames 5118 | Puerto Rico |
| 37 | US-TRI 47 | USDA | Ames 1993 | Taiwan |
| 38 | US-TRI 20 | USDA | Ames 2024 | Thailand |
| 39 | US-TRI 30 | USDA | Ames 5102 | Hong Kong |
| 40 | US-TRI 48 | USDA | Ames 1998 | Taiwan |
| 41 | US-TRI 49 | USDA | Ames 5134 | USA |
| 42 | US-TRI 51 | USDA | PI 633591 | Unknown |
| 43 | Local Red | LOCAL (Check) | var. BBS027 | Malaysia |
| 44 | Local PR | LOCAL (Check) | var. BBS014 | Malaysia |
| 45 | Thida | E-WEST (Check) | Thida | Tanzania |
| 46 | Zeya | E-WEST (Check) | Zeya | Tanzania |
| Yield (g) | LFW (g) | SFW (g) | LDW (g) | SDW (g) | TLA (cm2) | SLA (cm2g−1) | RWC | Fv’/Fm’ | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 100% WHC | 20% WHC | 100% WHC | 20% WHC | ||||||||
| Estimated variance | |||||||||||
| Trial (T) | 468.6 | 93.83 | 142.76 | 0.88 | 0.83 | 64634 | 4491 | 1.01 | 0 | 0.0049 | 0 |
| T × Water treatment (WT) | 2.4 | 1.02 | 0.23 | 0.06 | 0 | 2645 | 1213 | 2.08 | 7.1 | 0.0001 | 0.0003 |
| T × Accession(A) | 23.5 | 9.06 | 10.32 | 0.04 | 0.11 | 12758 | 2052 | 0.58 | 0 | 0.0001 | 0 |
| T × WT × A | −4.3 | −1.46 | −0.72 | 0.02 | −0.01 | −193 | 1121 | −0.05 | 0 | −0.0007 | 0 |
| Error Trial I | 19.82 | 6.316 | 4.75 | 0.14 | 0.1 | 4554 | 2042 | 32.01 | 227.1 | 0.003 | 0.0075 |
| Error Trial II | 123.3 | 35.52 | 41.82 | 0.54 | 0.41 | 39889 | 33123 | 39.91 | 153.5 | 0.0066 | 0.0077 |
| Wald tests for fixed effects | |||||||||||
| WT | 63.39 *** | 51.98 *** | 90.02 *** | 1.51 ns | 22.46 *** | 12.64 *** | 5.62 * | 0.77 ns | 76.65 *** | 0.01 ns | 5.44 * |
| A | 59.06 * | 65.23 * | 68.06 ** | 102.68 *** | 92.78 *** | 62.99 * | 30.44 ns | 67.57 * | 51.01 ns | 71.5 ** | 78.19 *** |
| WT × A | 132.96 *** | 204.03 *** | 116.47 *** | 48.86 ns | 55.82 ns | 91.3 *** | 26.51 ns | 37.86 ns | 61.2 * | 67.73 ** | 42.92 ns |
| Yield and Biomass Partitioning | |||||||
|---|---|---|---|---|---|---|---|
| Trial I | Yield (g) | LFW (g) | SFW (g) | TLA (cm2) | LDW (g) | SDW (g) | SLA (cm2g−1) |
| Mean | |||||||
| Water-sufficient | 15.78 | 8.42 | 7.37 | 265.49 | 1.15 | 0.94 | 239.62 |
| Water-deficient | 4.21 | 1.81 | 2.40 | 129.29 | 1.09 | 0.78 | 118.78 |
| SEM | 3.96 | 2.58 | 2.50 | 79.82 | 0.34 | 0.30 | 41.82 |
| p-values | |||||||
| Water treatment (WT) | 8838 * | 2882 * | 1626 * | 12,224,197 ** | 0.19 ns | 1.76 ns | 963,703 ** |
| Accession (A) | 28.88 *** | 12.52 *** | 13.07 *** | 17,104 ** | 0.47 *** | 0.32 *** | 4008 ** |
| WT × A | 20.54 *** | 8.28 ** | 6.90 *** | 7713 ** | 0.15 * | 0.08 ns | 2807 ns |
| Subplot error | 9.4 | 3.25 | 2.54 | 3677 | 0.1 | 0.06 | 1515 |
| Trial II | |||||||
| Mean | |||||||
| Water-sufficient | 48.11 | 23.2 | 24.90 | 682.32 | 2.48 | 2.27 | 305.06 |
| Water-deficient | 33.24 | 14.5 | 18.73 | 440.83 | 2.00 | 2.02 | 257.44 |
| SEM | 11.27 | 6.89 | 7.42 | 248.69 | 0.83 | 0.71 | 106.28 |
| p-values | |||||||
| Water treatment (WT) | 19,453 *** | 6690 ** | 3327 * | 5,132,472 ** | 20.34 ** | 5.48 ns | 199,527 * |
| Accession (A) | 6673 ** | 234.29 *** | 289.25 *** | 318,207 *** | 4.59 *** | 3.42 ** | 80,039 *** |
| WT × A | 115.11 ns | 42.42 * | 41.01 ns | 46,112 * | 0.41 ns | 0.36 ns | 41,261 ns |
| Subplot error | 94.9 | 26.7 | 36.19 | 31,761 | 0.49 | 0.39 | 33,114 |
| Relative Water Content (RWC) | Chlorophyll Fluorescence | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Light-Adapted Quantum Yield (Fv’/Fm’) | Dark-Adapted Quantum Yield (Fv/Fm) | ||||||||||||
| Trial I | Trial II | Trial I | Trial II | Trial II | |||||||||
| WHC | 100% | 20% | 100% | 50% | 20% | 100% | 20% | 100% | 50% | 20% | 100% | 50% | 20% |
| Mean | |||||||||||||
| Water-sufficient | 89.99 | 90.84 | 90.33 | 87.09 | 86.73 | 0.65 | 0.56 | 0.54 | 0.52 | 0.54 | 0.72 | 0.67 | 0.64 |
| Water-deficient | 87.13 | 61.12 | 90.65 | 82.92 | 75.98 | 0.64 | 0.49 | 0.54 | 0.53 | 0.53 | 0.72 | 0.67 | 0.63 |
| SEM | 3.25 | 3.63 | 2.08 | 4.63 | 3.27 | 0.02 | 0.03 | 0.04 | 0.03 | 0.04 | 0.03 | 0.04 | 0.04 |
| p-value | |||||||||||||
| Water treatment (WT) | 539.94 ns | 58,296 ** | 2.84 ns | 9124 ** | 40,687 * | 0.01 ns | 0.36 ns | 0.001 ns | 0.0002 ns | 0.03 ns | 0.02 ns | 0.02 ns | 0.03 ns |
| Accession (A) | 55.63 ** | 199.30 ns | 44.13 ns | 226.4 ** | 199.01 *** | 0.002 ns | 0.01 * | 0.003 ns | 0.008 *** | 0.01 *** | 0.006 ns | 0.01 * | 0.01 ** |
| WT × A | 28.12 ns | 229.20 ns | 39.60 ns | 121.50 ns | 181.64 ** | 0.001 ns | 0.06 ns | 0.003 ns | 0.002 ns | 0.005 ns | 0.003 ns | 0.04 ns | 0.01 ns |
| Subplot error | 30.08 | 209.3 | 39.98 | 129.6 | 94.61 | 0.002 | 0.06 | 0.002 | 0.003 | 0.006 | 0.005 | 0.004 | 0.01 |
| Leaf Gas Exchange | Photosynthesis, Pn (µmolCO2m−2s−1) | Stomatal Conductance, Gs (molH2Om−2s−1) | Transpiration, E (mmolH2Om−2s−1) | Intracellular [CO2], Ci (µmolCO2mol−1) | Instantaneous Water Use Efficiency, WUE (μmolmol−1) | Intrinsic WUE, WUEi (μmolmmol−1) | Stomatal Limitation, Ls | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WHC | 100% | 50% | 100% | 50% | 100% | 50% | 100% | 50% | 100% | 50% | 100% | 50% | 100% | 50% |
| Mean | ||||||||||||||
| Water-sufficient | 24.61 | 19.3 | 0.16 | 0.13 | 4.03 | 3.48 | 109.97 | 108.52 | 6.17 | 6.13 | 164.64 | 170.25 | 0.72 | 0.72 |
| Water-deficient | 33.04 | 19.07 | 0.22 | 0.12 | 4.99 | 3.17 | 111.67 | 95.78 | 7.05 | 6.35 | 160.01 | 176.45 | 0.71 | 0.75 |
| SEM | 7.99 | 6.27 | 0.06 | 0.05 | 1.11 | 1.34 | 42.27 | 50.56 | 1.35 | 1.67 | 29.55 | 37.53 | 0.11 | 0.13 |
| p-value | ||||||||||||||
| Water treatment (WT) | 591.57 ns | 2.62 ns | 0.305 ns | 0.010 ns | 739 ns | 8.49 ns | 77.53 ns | 16,542 ns | 65.33 ns | 5.20 ns | 1468 ns | 4227 ns | 0.01 ns | 0.09 ns |
| Accession (A) | 327.67 *** | 138.83 *** | 0.016 *** | 0.008 *** | 10,371 *** | 4.82 *** | 6.62 *** | 11,560 *** | 14.25 *** | 10.23 *** | 5234 *** | 4979 *** | 0.07 *** | 0.08 *** |
| WT × A | 155.85 ns | 119.82 *** | 0.007 ns | 0.007 *** | 4729 ns | 4.40 *** | 2.41 ns | 5523 ** | 6.27 ns | 8.44 *** | 2493 | 3346 * | 0.03 | 0.04 ** |
| Spit plot error | 47 | 55.25 | 0.003 | 0.003 | 1974 | 1.86 | 1.36 | 3248 | 1.73 | 2.93 | 880 | 2179 | 0.01 | 0.02 |
| Rewatering Assessment | |||||||
|---|---|---|---|---|---|---|---|
| Drought Stress | Recovery | ||||||
| DTF | DTW | LWS | DS | DTR | DS-1R | DS-5R | |
| Range | 0–4 | 1–8 | 0–5 | 0–7 | 1–6 | 0–9 | 0–9 |
| Mean | 3 | 3 | 3 | 2 | 2 | 4 | 5 |
| p-value | |||||||
| Accession | 2.67 ** | 2.87 ns | 2.209 ns | 2.22 ns | 5.73 *** | 16.24 *** | 29.61 *** |
| Error | 1.39 | 3.48 | 2.1 | 2.24 | 0.8 | 1.71 | 7.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jamalluddin, N.; Massawe, F.J.; Mayes, S.; Ho, W.K.; Singh, A.; Symonds, R.C. Physiological Screening for Drought Tolerance Traits in Vegetable Amaranth (Amaranthus tricolor) Germplasm. Agriculture 2021, 11, 994. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11100994
Jamalluddin N, Massawe FJ, Mayes S, Ho WK, Singh A, Symonds RC. Physiological Screening for Drought Tolerance Traits in Vegetable Amaranth (Amaranthus tricolor) Germplasm. Agriculture. 2021; 11(10):994. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11100994
Chicago/Turabian StyleJamalluddin, Norain, Festo J. Massawe, Sean Mayes, Wai Kuan Ho, Ajit Singh, and Rachael C. Symonds. 2021. "Physiological Screening for Drought Tolerance Traits in Vegetable Amaranth (Amaranthus tricolor) Germplasm" Agriculture 11, no. 10: 994. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11100994