
Effects of Composts Made from Broiler Chicken Residues and Blended with Biochar on the Minerals and Phenolic Compounds in Parsley (Petroselinum crispum Mill.)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Production of Organic Substrates
2.2. Treatments and Parsley Growing Conditions
2.3. Determination of Fresh and Dry Matter Yield
2.4. Determination of P, K, and N Contents in the Organic Substrates and Parsley
2.5. Determination of Total Phenolics (TPC), Total Flavonoids (TFC), and the Antioxidant Activity of Parsley Samples
2.6. Separation and Identification of Individual Phenolic Compounds
2.7. Determination of Total Anthocyanins (TAC) in Parsley Samples
2.8. Statistical Analysis
3. Results and Discussion
3.1. Chemical Characteristics of the Composts and the Organic Substrates (Compost-Biochar Mixtures)
3.2. Parsley Yield as Affected by the Different Organic Substrates
3.3. Nutritional Quality of Parsley Based on the Different Organic Substrates
3.4. Antioxidant Activity of Parsley Based on the Different Organic Substrates
3.5. Individual Flavonoids in Parsley Based on the Different Organic Substrates
3.6. PCA of Data
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Escobar, A.C.N.; Nascimento, A.L.; Gomes, J.G.; Borba, R.V.; Alves, C.C.; Costa, C.A. Avaliação da produtividade de três cultivares de salsa em função de diferentes substratos. Hortic. Bras. 2010, 28, S2671–S2676. [Google Scholar]
- Tamele, R.A.; Ueno, H.; Toma, Y.; Morita, N. Nitrogen recoveries and nitrogen use efficiencies of organic fertilizers with different C/N ratios in maize cultivation with low-fertile soil by 15N method. Agriculture 2020, 10, 272. [Google Scholar] [CrossRef]
- Morales, M.M.; Comerford, N.B.; Behling, M.; de Abreu, D.C.; Guerrini, I.A. Biochar chemistry in a weathered tropical soil: Kinetics of phosphorus sorption. Agriculture 2021, 11, 295. [Google Scholar] [CrossRef]
- Šubová, E.; Sasáková, N.; Zigo, F.; Mindžáková, I.; Vargová, M.; Kachnič, J.; Laktičová, K.V. Amendment of livestock manure with natural zeolite-clinoptilolite and its effect on decomposition processes during composting. Agriculture 2021, 11, 980. [Google Scholar] [CrossRef]
- Santos, C.; Goufo, P.; Fonseca, J.; Pereira, J.; Ferreira, L.; Coutinho, J.; Trindade, H. Effect of lignocellulosic and phenolic compounds on ammonia, nitric oxide and greenhouse gas emissions during composting. J. Clean. Prod. 2018, 171, 548–556. [Google Scholar] [CrossRef]
- Frišták, V.; Pipíška, M.; Turčan, V.; Bell, S.M.; Laughinghouse, H.D., IV; Ďuriška, L.; Soja, G. Preparation and characterization of novel magnesium composite/walnut shells-derived biochar for as and P sorption from aqueous solutions. Agriculture 2021, 11, 714. [Google Scholar] [CrossRef]
- Santos, F.T.; Ludwig, F.; Costa, L.A.M.; Costa, M.S.S.M. Nutrition and growth of potted gerbera according to mineral and organic fertilizer. Rev. Bras. Hortic. Ornam. 2015, 21, 251–258. [Google Scholar] [CrossRef]
- Santos, F.T.; Ludwig, F.; Costa, L.A.M.; Costa, M.S.S.M.; Remor, M.B.; Silva, P.E.R. Growth analysis of potted gerbera conducted with mineral fertilization and organic fertigation. Cienc. E Investig. Agraria 2016, 43, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Fornes, F.; Carrión, C.; García de la Fuente, R.; Puchades, R.; Abad, M. Leaching composted lignocellulosic wastes to prepare container media: Feasibility and environmental concerns. J. Environ. Manag. 2010, 91, 1747–1755. [Google Scholar] [CrossRef]
- Karaal, G.; Ugur, A. Lepidium sativum cultivation in organic fertilizer added hazelnut husk compost. Ekoloji Derg. 2014, 23, 33–39. [Google Scholar] [CrossRef]
- Prayogo, C.; Jones, J.E.; Baeyens, J.; Gary, D. Impact of biochar on mineralisation of C and N from soil and willow litter and its relationship with microbial community biomass and structure bending. Biol. Fertil. Soils 2014, 50, 695–702. [Google Scholar] [CrossRef]
- Dumroese, R.K.; Heiskanen, J.; Englund, K.; Tervahauta, A. Pelleted biochar: Chemical and physical properties show potential use as a substrate in container nurseries. Biomass Bioenergy 2011, 35, 2018–2027. [Google Scholar] [CrossRef]
- Tian, Y.; Suna, X.; Li, S.; Wang, H.; Wang, L.; Cao, J.; Zhang, L. Biochar made from green waste as peat substitute in growth media for Calathea rotundifola cv. Fasciata. Sci. Hortic. 2012, 143, 15–18. [Google Scholar] [CrossRef]
- Goufo, P.; Singh, R.K.; Cortez, I. A Reference list of phenolic compounds (including stilbenes) in grapevine (Vitis vinifera L.) roots, woods, canes, stems, and leaves. Antioxidants 2020, 9, 398. [Google Scholar] [CrossRef]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: Chemistry, bioavailability and effects on health. Nat. Prod. Rep. 2009, 26, 1001–1043. [Google Scholar] [CrossRef]
- Goufo, P.; Henrique, H. Factors influencing antioxidant compounds in rice. Crit. Rev. Food Sci. Nutr. 2017, 57, 893–922. [Google Scholar] [CrossRef]
- Naczk, M.; Shahidi, F. Extraction and analysis of phenolics in food. J. Chromatogr. A 2004, 1054, 95–111. [Google Scholar] [CrossRef]
- Deng, J.; Cheng, W.; Yang, G. A novel antioxidant activity index (AAU) for natural products using the DPPH assay. Food Chem. 2011, 125, 1430–1435. [Google Scholar] [CrossRef]
- Oliveira, G.L.S. Determination in vitro of the antioxidant capacity of natural products by the DPPH•method: Review study. Rev. Bras. Plant. Med. 2015, 17, 36–44. [Google Scholar] [CrossRef]
- Liberal, Â.; Fernandes, Â.; Polyzos, N.; Petropoulos, S.A.; Dias, M.I.; Pinela, J.; Petrović, J.; Soković, M.; Ferreira, I.C.F.R.; Barros, L. Bioactive properties and phenolic compound profiles of turnip-rooted, plain-leafed and curly-leafed parsley cultivars. Molecules 2020, 25, 5606. [Google Scholar] [CrossRef]
- Luthria, D. Influence of experimental conditions on the extraction of phenolic compounds from parsley (Petroselinum crispum) flakes using a pressurized liquid extractor. Food Chem. 2008, 107, 745–752. [Google Scholar] [CrossRef]
- Epifanio, N.M.M.; CavalcantI, L.R.I.; Santos, K.F.; Duarte, P.S.C.; Kachlicki, P.; Ożarowski, M.; Riger, C.J.; Chaves, D.S.A. Chemical characterization and in vivo antioxidant activity of parsley (Petroselinum crispum). Food Funct. 2020, 11, 5346–5356. [Google Scholar] [CrossRef]
- Vora, S.R.; Patil, R.B.; Pillai, M.M. Oxidative stress associated alterations in lysosomal enzymes and modulatory effect of Petroselinum crispum (Mill) Nyman Ex. A.W. hill leaf extract on mouse brain. Int. J. Sci. Res. 2012, 7, 64–68. [Google Scholar]
- Nitsche, P.R.; Caramori, P.H.; Ricce, W.S.; Pinto, L.F.D. Atlas Climático do Estado do Paraná, 1st ed.; Instituto Agronomico Do Paraná (IAPAR): Londrina, Brazil, 2019; pp. 1–216. [Google Scholar]
- Lana, M.C.; Fey, R.; Francoloso, J.F.; Richart, A.; Fontaniva, S. Análise Química de Solo e Tecido Vegetal: Práticas de Laboratório, 1st ed.; Edunioeste: Cascavel, Brazil, 2010; pp. 1–130. [Google Scholar]
- Silva, C.F. Manual de Análises Químicas de Solos, Plantas e Fertilizantes, 2nd ed.; Embrapa (Empresa Brasileira de Pesquisa Agropecuária) Agricultura Digital e Embrapa Solos: Brasília, Brazil, 2009; pp. 1–627. [Google Scholar]
- Malavolta, E. ABC da Adubação, 5th ed.; Agronômica Ceres: São Paulo, Brazil, 1989; pp. 1–292. [Google Scholar]
- Li, Z.; Zhao, X.; Sandhu, A.K.; Gu, L. Effects of exogenous abscisic acid on yield, antioxidant capacities, and phytochemical contents of greenhouse grown lettuces. J. Agric. Food Chem. 2010, 58, 6503–6509. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A., Jr. Colorimetry of total phenolics with phosphomolybdic phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Huber, L.S.; Rodriguez-Amaya, D.B.; Rodrigues, M.I. Optimization and validation of HPLC methodology for determining flavonols and flavones in vegetables. Rev. Inst. Adolfo Lutz 2007, 66, 142–151. [Google Scholar]
- Rodriguez-Saona, L.E.; Giusti, M.M.; Wrolstad, R.E. Anthocyanin pigment composition of red-fleshed potatoes. J. Food Sci. 1998, 63, 458–465. [Google Scholar] [CrossRef]
- Jackson, D.A. Stopping rules in principal components analysis: A comparison of heuristical and statistical approaches. Ecology 1993, 74, 2204–2214. [Google Scholar] [CrossRef]
- Norhafizah, M.Z.; Ismail, B.S.; Chuah, T.S. Herbicidal activity of Pennisetum purpureum (Napier grass). Afr. J. Biotechnol. 2012, 11, 6269–6273. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, R.; Nascimento, D.A.M.; Silva, D.A.; Alves, A.G. Alterações nos teores de clorofilas em plantas de feijão-caupi cultivadas sob diferentes fontes de nitrogênio. Rev. Educ. Agrícola Superior 2012, 27, 94–96. [Google Scholar] [CrossRef]
- Lechtenberg, M.; Zumdick, S.; Gerhards, C.; Schmidt, T.J.; Hensel, A. Evaluation of analytical markers characterising different drying methods of parsley leaves (Petroselinum crispum L.). Die Pharm. 2007, 62, 949–954. [Google Scholar] [CrossRef] [PubMed]
- van Raij, B.; Cantarella, H.; Quaggio, J.A.; Furlani, A.M.C. Boletim Técnico 100: Recomendações de Adubação e Calagem Para o Estado de São Paulo, 2nd ed.; Instituto Agronômico & Fundação IAC: Campinas, Brazil, 1997; pp. 1–285. [Google Scholar]
- Malavolta, E. O Fósforo na planta e interações com outros elementos. In Fósforo na Agricultura Brasileira, 1st ed.; Yamada, T., Abdalla, S.R.S., Eds.; Associacao Brasileira para Pesquisa da Potassa e do Fosfato (Piracicaba, SP): Piracicaba, Brazil, 2004; pp. 35–98. [Google Scholar]
- Wei, Y.; Zhao, Y.; Xi, B.; Wei, Z.; Li, X.; Cao, Z. Changes in phosphorus fractions during organic wastes composting from different sources. Bioresour. Technol. 2015, 189, 349–356. [Google Scholar] [CrossRef]
- Rheinheimer, D.R.S.; Gatibonil, L.C.; Kaminskil, J. Factors affecting the phosphorus availability and the fertilization management in no-tillage system. Cienc. Rural 2008, 38, 576–586. [Google Scholar]
- Cavar, S.; Vidic, D.; Maksimovic, M. Volatile constituents, phenolic compounds, and antioxidant activity of Calamintha glandulosa (Req.) Bentham. J. Sci. Food Agric. 2012, 93, 1758–1764. [Google Scholar] [CrossRef] [PubMed]
- Kumaran, A.; Karunakaran, R.J. In vitro antioxidant activities of methanol extracts of five Phyllanthus species from India. Food Sci. Technol. 2007, 40, 344–352. [Google Scholar] [CrossRef]
- Olsson, M.E.; Andersson, C.S.; Oredsson, S.; Berglund, R.H.; Gustavsson, K.E. Antioxidant levels and inhibition of cancer cell proliferation in vitro by extracts from organically and conventionally cultivated strawberries. J. Agric. Food Chem. 2006, 54, 1248–1255. [Google Scholar] [CrossRef]
- Naczk, M.; Shahidi, F. Phenolics in cereals, fruits and vegetables: Occurrence, extraction and analysis. J. Pharm. Biomed. Anal. 2006, 41, 1523–1542. [Google Scholar] [CrossRef]
- Meyer, H.; Bolarinwa, A.; Wolfram, G.; Linseisen, J. Bioavailability of apigenin from apiin-rich parsley in humans. Ann. Nutr. Metab. 2006, 50, 167–172. [Google Scholar] [CrossRef]
- Kaiser, A.; Carle, R.; Kammerer, D.R. Effects of blanching on polyphenol stability of innovative paste-like parsley (Petroselinum crispum (Mill.) Nym ex A. W. Hill) and marjoram (Origanum majorana L.) products. Food Chem. 2013, 138, 1648–1656. [Google Scholar] [CrossRef]
- El-Zaeddi, H.; Calín-Sánchez, H.; Nowicka, P.; Martínez-Tomé, J.; Noguera-Artiaga, L.; Burló, F.; Wojdyło, A.; Carbonell-Barrachina, A.A. Preharvest treatments with malic, oxalic, and acetylsalicylic acids affect the phenolic composition and antioxidant capacity of coriander, dill and parsley. Food Chem. 2017, 226, 179–186. [Google Scholar] [CrossRef]
- Farzaei, M.H.; Abbasabadi, Z.; Assdekani, M.R.S.; Rahimi, R.; Farzaei, F. Parsley: A review of ethnopharmacology, phytochemistry and biological activities. J. Tradit. Chin. Med. 2013, 6, 815–826. [Google Scholar] [CrossRef] [Green Version]
- Hostetler, G.L.; Riedl, K.M.; Schwatz, S.L. Endogenous enzymes, heat, and pH affect flavone profiles in parsley (Petroselinum crispum var. neapolitanum) and celery (Apium graveolens) during juice processing. J. Agric. Food Chem. 2012, 11, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boldizsár, I.; Fuzfaib, Z.; Molnár-Perlb, I. Characterization of the endogenous enzymatic hydrolyses of Petroselinum crispum glycosides: Determined by chromatography upon their sugar and flavonoid products. J. Chromatogr. A 2013, 1293, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Kocsis, T.; Kotroczó, Z.; Kardos, L.; Biró, B. Optimization of increasing biochar doses with soil–plant–microbial functioning and nutrient uptake of maize. Environ. Technol. Innov. 2020, 20, 101191. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Organic Substrate | Proportion of Compost + Proportion of Biochar | N (g kg−1) | P (g kg−1) | K (g kg−1) | EC (dS m−1) | pH |
|---|---|---|---|---|---|---|
| B60 | 40% of bagasse–chicken compost + 60% of biochar | 21.52 ± 0.87 g | 1.72 ± 0.25 ef | 7.35 ± 0.28 d | 5.07 ± 0.26 g | 8.16 ± 0.21 a |
| B45 | 55% of bagasse–chicken compost + 45% of biochar | 25.13 ± 1.06 f | 2.13 ± 0.25 cde | 6.76 ± 0.44 ef | 5.71 ± 0.74 f | 7.64 ± 0.46 a |
| B30 | 70% of bagasse–chicken compost + 30% of biochar | 28.69 ± 3.12 ej | 2.77 ± 0.23 b | 6.09 ± 0.51 def | 6.75 ± 1.32 f | 7.31 ± 0.63 a |
| B15 | 85% of bagasse–chicken compost + 15% of biochar | 29.98 ± 1.88 e | 2.75 ± 0.26 b | 5.67 ± 0.64 f | 8.45 ± 0.75 def | 6.87 ± 1.18 a |
| B00 | 100% of bagasse–chicken compost + 0% of biochar | 31.18 ± 0.88 e | 3.07 ± 0.18 a | 5.40 ± 1.04 f | 10.40 ± 0.93 d | 6.85 ± 0.14 a |
| S60 | 40% of sawdust–chicken compost + 60% of biochar | 14.56 ± 0.98 h | 1.62 ± 0.07 fg | 6.17 ± 1.03 ef | 2.67 ± 0.14 i | 8.69 ± 0.37 a |
| S45 | 55% of sawdust–chicken compost + 45% of biochar | 19.83 ± 1.50 g | 1.99 ± 0.24 de | 5.89 ± 0.46 ef | 3.18 ± 0.50 hi | 8.36 ± 0.44 a |
| S30 | 70% of sawdust–chicken compost + 30% of biochar | 20.63 ± 1.60 g | 2.25 ± 0.13 cd | 5.64 ± 0.49 f | 3.62 ± 0.36 h | 8.04 ± 0.12 a |
| S15 | 85% of sawdust–chicken compost + 15% of biochar | 23.63 ± 0.78 fg | 2.59 ± 0.19 bc | 4.57 ± 0.27 fg | 5.03 ± 0.51 g | 7.43 ± 0.48 a |
| S00 | 100% of sawdust–chicken compost + 0% of biochar | 26.23 ± 1.34 ef | 3.00 ± 0.15 a | 3.88 ± 0.21 g | 5.37 ± 0.42 g | 7.30 ± 0.75 a |
| T60 | 40% of tree–chicken compost + 60% of biochar | 21.50 ± 1.93 g | 1.29 ± 0.22 g | 7.43 ± 0.34 d | 3.44 ± 0.58 h | 8.94 ± 0.53 a |
| T45 | 55% of tree–chicken compost + 45% of biochar | 21.54 ± 1.26 g | 1.41 ± 0.24 g | 7.63 ± 0.15 d | 3.79 ± 0.47 h | 8.79 ± 1.17 a |
| T30 | 70% of tree–chicken compost + 30% of biochar | 29.58 ± 2.25 e | 1.72 ± 0.12 ef | 7.16 ± 1.10 de | 4.53 ± 0.73 gh | 8.25 ± 0.75 a |
| T15 | 85% of tree–chicken compost + 15% of biochar | 36.33 ± 1.10 cde | 1.88 ± 0.30 de | 7.25 ± 0.71 d | 6.40 ± 0.62 f | 7.64 ± 0.48 a |
| T00 | 100% of tree–chicken compost + 0% of biochar | 38.81 ± 2.11 cd | 2.15 ± 0.29 cde | 6.85 ± 0.94 de | 6.28 ± 0.73 f | 7.59 ± 0.41 a |
| N60 | 40% of napier–chicken compost + 60% of biochar | 21.18 ± 2.54 g | 1.16 ± 0.21 g | 9.61 ± 0.78 c | 5.56 ± 0.64 fg | 8.94 ± 1.62 a |
| N45 | 55% of napier–chicken compost + 45% of biochar | 25.43 ± 0.73 f | 1.75 ± 0.22 ef | 9.74 ± 0.67 c | 6.72 ± 0.35 f | 8.75 ± 1.04 a |
| N30 | 70% of napier–chicken compost + 30% of biochar | 27.74 ± 1.93 ef | 2.17 ± 0.66 cd | 9.72 ± 0.85 c | 8.35 ± 0.61 ef | 8.31 ± 0.17 a |
| N15 | 85% of napier–chicken compost + 15% of biochar | 34.42 ± 3.21 d | 2.34 ± 0.24 bc | 10.17 ± 0.82 bc | 9.82 ± 0.40 de | 8.39 ± 0.81 a |
| N00 | 100% of napier–chicken compost + 0% of biochar | 40.67 ± 2.92 c | 2.30 ± 0.41 cd | 10.95 ± 0.48 bc | 11.70 ± 0.52 d | 7.60 ± 0.69 a |
| C60 | 40% of cotton–chicken compost + 60% of biochar | 24.42 ± 2.59 f | 1.79 ± 0.30 ef | 10.54 ± 0.24 bc | 6.55 ± 0.24 f | 8.61 ± 0.50 a |
| C45 | 55% of cotton–chicken compost + 45% of biochar | 32.89 ± 1.57 de | 2.30 ± 0.30 bc | 10.74 ± 0.21 bc | 7.94 ± 0.58 ef | 8.25 ± 0.07 a |
| C30 | 70% of cotton–chicken compost + 30% of biochar | 39.76 ± 0.90 c | 2.64 ± 0.16 b | 12.15 ± 0.50 ab | 13.20 ± 0.78 c | 7.81 ± 0.27 a |
| C15 | 85% of cotton–chicken compost + 15% of biochar | 44.96 ± 0.91 b | 2.99 ± 0.23 ab | 12.13 ± 0.90 ab | 15.52 ± 0.94 b | 7.79 ± 0.25 a |
| C00 | 100% of cotton–chicken compost + 0% of biochar | 53.41 ± 2.54 a | 3.43 ± 0.21 a | 13.34 ± 0.44 a | 18.30 ± 0.82 a | 7.38 ± 0.13 a |
| Organic Substrate | Fresh Matter Yield (g pot−1 FW) | Dry Matter Yield (g pot−1 DW) | N Content (g kg−1) | P Content (g kg−1) | K Content (g kg−1) |
|---|---|---|---|---|---|
| B60 | 25.12 ± 1.40 bc | 4.33 ± 0.35 b | 22.89 ± 2.34 de | 2.11 ± 0.29 cd | 26.06 ± 1.21 ef |
| B45 | 27.91 ± 2.09 b | 3.88 ± 0.41 bc | 32.38 ± 1.65 bc | 2.60 ± 0.14 bc | 30.35 ± 3.78 cd |
| B30 | 25.26 ± 4.90 bc | 3.80 ± 0.82 bc | 34.40 ± 1.14 ab | 2.84 ± 0.23 ab | 27.03 ± 1.03 def |
| B15 | 19.01 ± 2.87 cd | 2.74 ± 0.40 de | 35.48 ± 0.64 a | 2.23 ± 0.21 bcd | 34.25 ± 4.31 bc |
| B00 | 19.00 ± 1.90 de | 2.34 ± 0.34 def | 33.82 ± 2.95 bc | 3.03 ± 0.24 a | 25.28 ± 1.21 f |
| S60 | 21.05 ± 1.55 cd | 3.11 ± 0.07 cd | 21.73 ± 1.36 de | 2.07 ± 0.16 cd | 28.31 ± 2.12 de |
| S45 | 27.10 ± 1.19 b | 4.38 ± 0.10 b | 23.31 ± 2.53 cde | 1.99 ± 0.16 cde | 29.24 ± 0.31 cd |
| S30 | 26.95 ± 2.10 bc | 4.33 ± 0.28 b | 29.81 ± 1.40 bcd | 2.33 ± 0.11 bc | 26.76 ± 2.30 ef |
| S15 | 36.83 ± 2.58 a | 5.65 ± 0.32 a | 28.57 ± 0.27 bcd | 2.38 ± 0.14 bc | 26.11 ± 2.70 ef |
| S00 | 26.13 ± 4.11 bc | 3.85 ± 0.68 bc | 35.37 ± 1.40 a | 3.28 ± 0.23 a | 29.89 ± 2.01 cd |
| T60 | 9.84 ± 0.20 f | 1.61 ± 0.04 fg | 15.21 ± 0.43 e | 2.29 ± 0.05 bcd | 28.98 ± 0.89 de |
| T45 | 12.70 ± 0.66 ef | 1.82 ± 0.16 ef | 25.43 ± 1.57 cd | 1.68 ± 0.25 de | 48.25 ± 2.15 a |
| T30 | 16.46 ± 0.80 de | 2.64 ± 0.11 de | 26.85 ± 0.65 cd | 2.25 ± 0.20 bcd | 45.97 ± 3.26 a |
| T15 | 17.05 ± 0.40 de | 2.49 ± 0.13 de | 30.76 ± 0.38 bc | 2.95 ± 0.04 ab | 26.35 ± 0.66 ef |
| T00 | 13.87 ± 0.27 def | 1.79 ± 0.07 fg | 30.03 ± 1.13 bc | 3.03 ± 0.45 a | 28.98 ± 2.02 de |
| N60 | 3.46 ± 1.07 g | 1.18 ± 0.44 g | 23.32 ± 2.47 cde | 1.12 ± 0.10 ef | 47.88 ± 1.63 a |
| N45 | 9.60 ± 2.90 f | 1.26 ± 0.50 fg | 30.41 ± 1.85 bc | 1.88 ± 0.05 cde | 45.10 ± 1.56 a |
| N30 | 2.94 ± 1.86 g | 0.41 ± 0.26 h | 9.17 ± 5.17 f | 1.00 ± 1.00 f | 6.80 ± 3.80 g |
| N15 | 0.00 ± 0.00 h | 0.00 ± 0.00 i | NA | NA | NA |
| N00 | 0.00 ± 0.00 h | 0.00 ± 0.00 i | NA | NA | NA |
| C60 | 16.31 ± 3.14 de | 2.81 ± 0.59 cde | 26.18 ± 0.48 cd | 1.64 ± 0.19 def | 36.40 ± 2.46 b |
| C45 | 15.50 ± 2.90 de | 2.81 ± 0.60 cde | 31.14 ± 3.06 bc | 1.64 ± 0.29 def | 36.18 ± 2.18 b |
| C30 | 19.20 ± 0.75 cd | 3.24 ± 0.06 cd | 31.18 ± 0.55 bc | 1.75 ± 0.08 de | 45.19 ± 1.86 a |
| C15 | 3.22 ± 1.22 g | 0.41 ± 0.17 h | 29.82 ± 0.60 bcd | 1.75 ± 0.07 de | 45.19 ± 1.85 a |
| C00 | 2.00 ± 0.49 g | 0.27 ± 0.08 h | 29.82 ± 0.60 bcd | 1.75 ± 0.07 de | 45.19 ± 1.40 a |
| Organic Substrate | DPPH Scavenging Activity (%) | DPPH Scavenging Activity (g TE g−1) | Total Flavonoids (mg CE g−1) | Total Phenolics (mg GAE g−1) | Total Anthocyanins (mg CGE g−1) |
|---|---|---|---|---|---|
| B60 | 48.49 ± 2.56 bc | 17.08 ± 1.00 b | 20.44 ± 2.05 e | 9.22 ± 0.42 a | 0.34 ± 0.02 cd |
| B45 | 38.61 ± 4.07 e | 13.60 ± 1.63 d | 15.14 ± 1.49 gh | 6.95 ± 0.64 cde | 0.26 ± 0.01 ef |
| B30 | 30.07 ± 2.34 gh | 16.32 ± 3.52 bc | 16.84 ± 0.79 e | 6.54 ± 0.35 cde | 0.25 ± 0.00 efg |
| B15 | 28.77 ± 2.79 gh | 9.70 ± 1.17 g | 13.70 ± 0.60 f | 6.24 ± 0.40 de | 0.32 ± 0.01 de |
| B00 | 27.41± 2.42 gh | 9.34 ± 0.91 g | 15.88 ± 1.50 e | 6.29 ± 0.69 cde | 0.35 ± 0.04 bc |
| S60 | 34.73 ± 2.94 f | 12.19 ± 1.25 ef | 27.18 ± 1.16 a | 7.39 ± 0.88 bc | 0.34 ± 0.05 cd |
| S45 | 35.49 ± 1.00 f | 12.49 ± 0.30 e | 20.97 ± 1.40 ef | 7.31 ± 0.96 bc | 0.28 ± 0.00 def |
| S30 | 26.14 ± 0.58 h | 9.47 ± 0.74 g | 19.01 ± 1.41 d | 5.30 ± 0.26 def | 0.40 ± 0.03 b |
| S15 | 29.52 ± 2.49 gh | 14.64 ± 2.19 c | 19.75 ± 0.87 cd | 6.69 ± 1.03 cd | 0.36 ± 0.02 bc |
| S00 | 34.81 ± 2.35 f | 12.32 ± 0.93 e | 9.71 ± 0.58 h | 6.69 ± 0.51 cd | 0.22 ± 0.01 g |
| T60 | 47.63 ± 0.94 c | 17.57 ± 0.28 b | 24.48 ± 0.79 b | 7.93 ± 0.49 b | 0.32 ± 0.05 de |
| T45 | 50.65 ± 5.72 b | 21.79 ± 4.04 a | 22.76 ± 1.16 bc | 6.96 ± 0.93 bcd | 0.20 ± 0.01 g |
| T30 | 47.61 ± 2.19 c | 17.17 ± 0.79 b | 23.50 ± 1.99 bc | 8.20 ± 0.40 b | 0.23 ± 0.01 fg |
| T15 | 41.48 ± 1.98 de | 14.96 ± 0.64 c | 20.03 ± 2.23 cd | 7.23 ± 0.46 bc | 0.45 ± 0.07 ab |
| T00 | 32.62 ± 0.65 g | 11.17 ± 0.26 f | 24.27 ± 1.52 b | 6.92 ± 0.52 bcd | 0.23 ± 0.01 fg |
| N60 | 32.90 ± 1.94 g | 11.44 ± 0.85 f | 15.70 ±0.30 fg | 3.74 ± 0.02 fg | 0.34 ± 0.00 cd |
| N45 | 62.26 ± 5.44 a | 20.52 ± 3.65 a | 13.64 ± 1.05 f | 6.34 ± 0.61 cde | 0.40 ± 0.04 b |
| N30 | 34.62 ± 7.82 f | 13.89 ± 2.21 cd | 11.55 ± 1.30 g | 2:12 ± 0.09 g | 0.48 ± 0.00 a |
| N15 | NA | NA | NA | NA | NA |
| N00 | NA | NA | NA | NA | NA |
| C60 | 47.08 ± 6.95 c | 13.27 ± 0.41 d | 8.43 ± 0.54 h | 8.58 ± 0.96 ab | 0.28 ± 0.02 def |
| C45 | 42.78 ± 2.30 d | 15.30 ± 0.99 c | 11.53 ± 1.80 g | 8.53 ± 1.51 ab | 0.37 ± 0.05 bc |
| C30 | 34.26 ± 3.24 f | 13.86 ± 0.73 d | 13.85 ± 0.84 f | 8.40 ± 0.77 ab | 0.32 ± 0.01 de |
| C15 | 37.21 ± 1.82 ef | 13.02 ± 0.79 de | 13.01 ± 0.75 f | 5.19 ± 0.28 ef | 0.26 ± 0.00 ef |
| C00 | 38.48 ± 1.17 e | 13.94 ± 0.65 cd | 13.68 ± 0.38 f | 6.57 ± 0.87 cd | 0.46 ± 0.03 a |
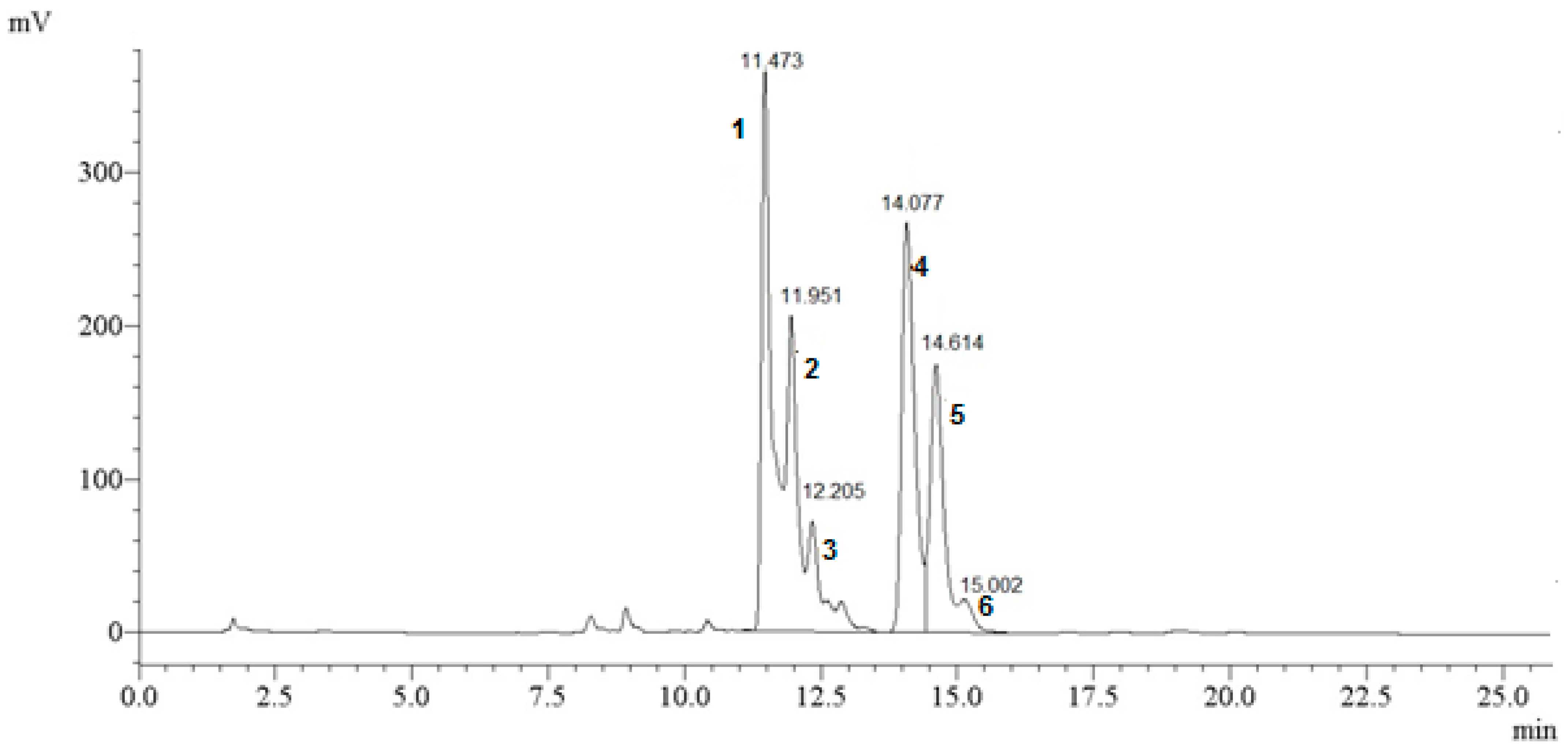
| Organic Substrate | Apigenin-7-apiosylglucoside (1) | Diosmetin-apiosylglucoside (2) | Diosmetin-apiosylglucoside Isomer (3) | Apigenin-malonyl-apiosylglucoside (4) | Diosmetin-malonyl-apiosylglucoside (5) | Apigenin-malonylglucoside (6) |
|---|---|---|---|---|---|---|
| B60 | 18.69 ± 1.20 cd | 6.96 ± 0.56 c | 1.75 ± 0.14 d | 3.51 ± 0.15 e | 2.15 ± 0.25 f | 0.15 ± 0.05 f |
| B45 | 13.96 ± 1.78 ef | 6.96 ± 0.60 c | 1.49 ± 0.20 ef | 1.21 ± 0.48 i | 0.79 ± 0.07 j | ND |
| B30 | 13.82 ± 2.10 ef | 5.19 ± 0.16 efg | 1.47 ± 0.07 ef | 4.01 ± 0.58 d | 2.63 ± 0.18 d | 0.15 ± 0.00 f |
| B15 | 12.96 ± 0.24 fg | 3.55 ± 0.85 i | 1.16 ± 0.09 g | 4.62 ± 0.16 c | 2.44 ±0.32 de | 0.14 ± 0.01 f |
| B00 | 18.32 ± 1.97 cd | 4.83 ± 0.55 fg | 1.31 ± 0.16 efg | 1.35 ± 0.80 ijk | 0.69 ± 0.06 jk | ND |
| S60 | 20.08 ± 1.43 b | 6.16 ± 0.68 cde | 1.50 ± 0.16 ef | 2.96 ± 0.63 f | 1.37 ± 0.15 g | 0.44 ± 0.17 c |
| S45 | 25.21 ± 1.75 a | 6.96 ± 0.38 c | 2.35 ± 0.38 b | 3.53 ± 0.30 e | 1.69 ± 0.16 g | 0.26 ± 0.10 de |
| S30 | 16.30 ± 1.19 de | 5.39 ± 0.95 def | 1.24 ± 0.08 fg | 2.02 ± 0.42 hi | 1.20 ± 0.10 gh | ND |
| S15 | 19.74 ± 1.36 bc | 6.61 ± 0.35 cd | 1.66 ± 0.28 def | 2.99 ± 0.00 f | 1.42 ± 0.02 fg | ND |
| S00 | 17.67 ± 0.86 cde | 6.46 ± 0.62 cd | 1.65 ± 0.27 d | 4.42 ± 0.83 c | 2.29 ± 0.03 ef | ND |
| T60 | 12.95 ± 0.74 fg | 5.20 ± 0.46 defg | 7.35 ± 0.20 a | 5.15 ± 0.11 b | 5.25 ± 0.43 a | 1.45 ± 0.12 a |
| T45 | 21.50 ± 0.99 b | 7.52 ± 1.41 bc | 2.34 ± 0.32 b | 5.39 ± 0.37 a | 3.44 ± 0.09 b | 0.47 ± 0.05 c |
| T30 | 24.35 ± 2.88 a | 9.98 ± 0.52 a | 2.46 ± 0.25 b | 3.96 ± 0.46 d | 2.34 ± 0.42 e | 0.43 ± 0.07 c |
| T15 | 21.22 ± 2.94 b | 6.95 ± 0.71 c | 1.98 ± 0.12 c | 5.22 ± 0.47 ab | 3.09 ± 0.25 c | 0.36 ± 0.06 d |
| T00 | 18.38 ± 1.51 cd | 6.18 ± 0.55 cde | 1.66 ± 0.15 def | 3.76 ± 0.02 de | 2.35 ± 0.18 e | 0.12 ± 0.01 f |
| N60 | 12.30 ± 0.03 fg | 4.87 ± 0.51 fg | 1.28 ± 0.27 fg | 0.82 ± 0.09 j | 0.40 ± 0.00 k | 0.01 ± 0.00 e |
| N45 | 15.57 ± 1.34 def | 5.87 ± 0.60 de | 1.27 ± 0.05 fg | 0.63 ± 0.00 j | 0.79 ± 0.07 jk | 0.01 ± 0.00 c |
| N30 | 10.72 ± 1.18 g | 4.43 ± 0.54 gh | 0.42 ± 0.06 h | 0.07 ± 0.00 k | 0.00 ± 0.00 l | 0.01 ± 0.00 c |
| N15 | NA | NA | NA | NA | NA | NA |
| N00 | NA | NA | NA | NA | NA | NA |
| C60 | 23.34 ± 1.66 ab | 8.10 ± 0.79 b | 2.15 ± 0.29 bc | 2.14 ± 0.40 gh | 1.58 ± 0.19 fg | 0.64 ± 0.02 b |
| C45 | 15.30 ± 1.05 def | 5.61 ± 0.56 de | 1.62 ± 0.09 def | 4.56 ± 0.34 c | 3.06 ± 0.14 c | 0.20 ± 0.05 e |
| C30 | 15.04 ± 0.78 def | 6.29 ± 1.18 cde | 1.41 ± 0.22 ef | 3.83 ± 0.37 de | 2.17 ± 0.25 f | 0.22 ± 0.03 e |
| C15 | 13.29 ± 0.79 efg | 4.08 ± 0.22 h | 1.09 ± 0.08 g | 2.39 ± 0.59 fg | 1.02 ± 0.15 i | ND |
| C00 | 17.96 ± 0.24 cde | 5.38 ± 0.53 def | 1.38 ± 0.17 efg | 1.68 ± 0.18 i | 1.18 ± 0.12 hi | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, F.T.; Trindade, H.; Costa, M.S.S.M.; Costa, L.A.M.; Goufo, P. Effects of Composts Made from Broiler Chicken Residues and Blended with Biochar on the Minerals and Phenolic Compounds in Parsley (Petroselinum crispum Mill.). Agriculture 2021, 11, 1168. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11111168
Santos FT, Trindade H, Costa MSSM, Costa LAM, Goufo P. Effects of Composts Made from Broiler Chicken Residues and Blended with Biochar on the Minerals and Phenolic Compounds in Parsley (Petroselinum crispum Mill.). Agriculture. 2021; 11(11):1168. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11111168
Chicago/Turabian StyleSantos, Francielly T., Henrique Trindade, Mônica S. S. M. Costa, Luiz A. M. Costa, and Piebiep Goufo. 2021. "Effects of Composts Made from Broiler Chicken Residues and Blended with Biochar on the Minerals and Phenolic Compounds in Parsley (Petroselinum crispum Mill.)" Agriculture 11, no. 11: 1168. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11111168