
Florpyrauxifen-Benzyl Selectivity to Rice

 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Florpyrauxifen-Benzyl Selectivity to Rice, as Affected by Planting Time, Stage, and Rate of Application
2.1.1. Site and Plant Material Description
2.1.2. Experimental Procedures
2.1.3. Evaluations
2.1.4. Statistical Analysis
2.2. Florpyrauxifen-Benzyl Selectivity to Rice, as Affected by P450 Inhibitors, and the Tolerance of Two Cultivars
2.3. Florpyrauxifen-Benzyl Selectivity to Rice, as Affected by Temperature Regimen and the Expression of Target Candidate Genes
2.3.1. Temperature and Rice Injury
2.3.2. Gene Expression of CYP450, GST, and Esterase Candidate Genes
Plant Material, Growth Conditions, and Treatments
RNA Extraction and cDNA Synthesis
Gene Expression Quantification by RT-qPCR
3. Results
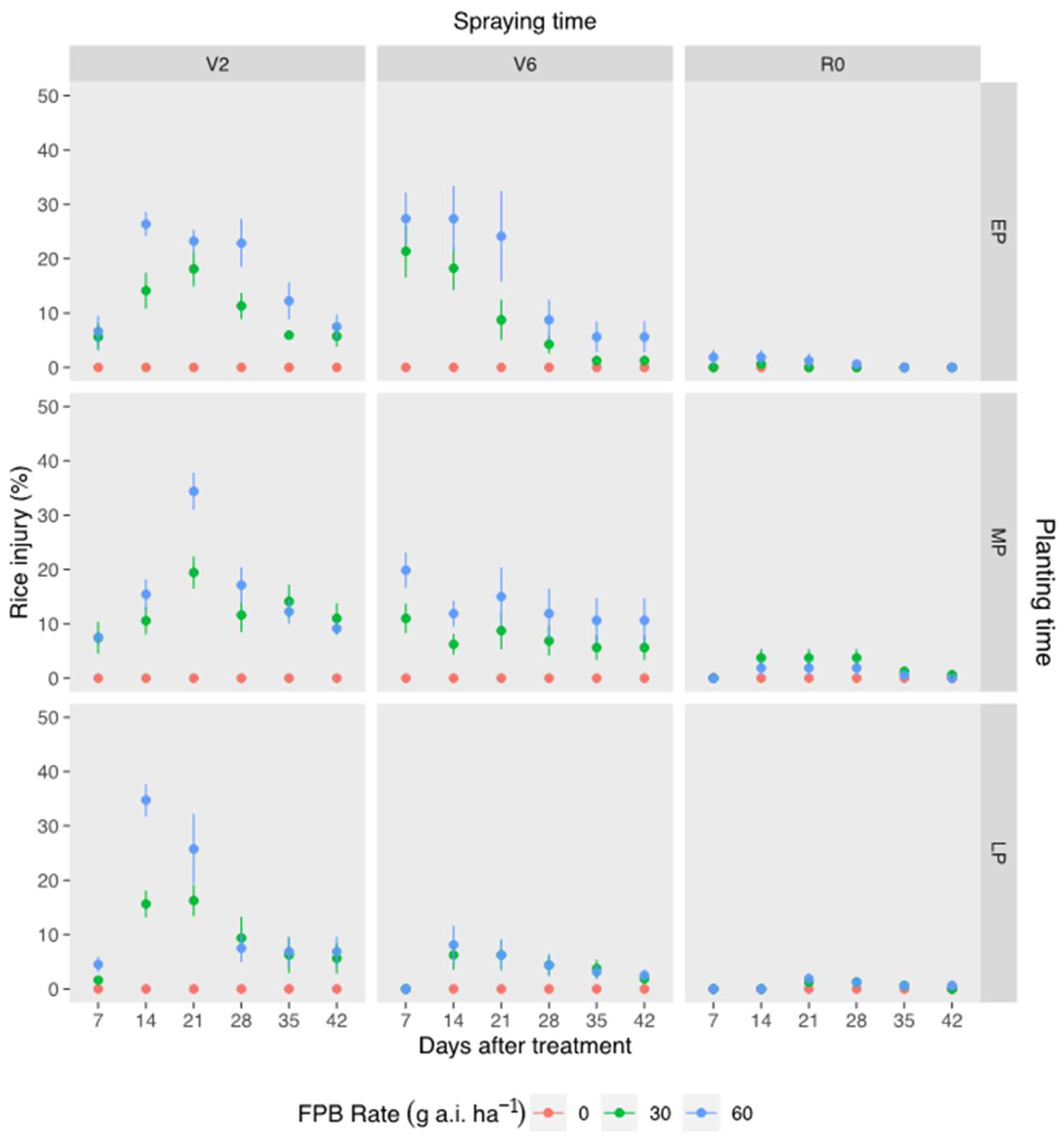
3.1. Florpyrauxifen-Benzyl Selectivity to Rice, as Affected by Planting Time, Stage, and Rate of Application
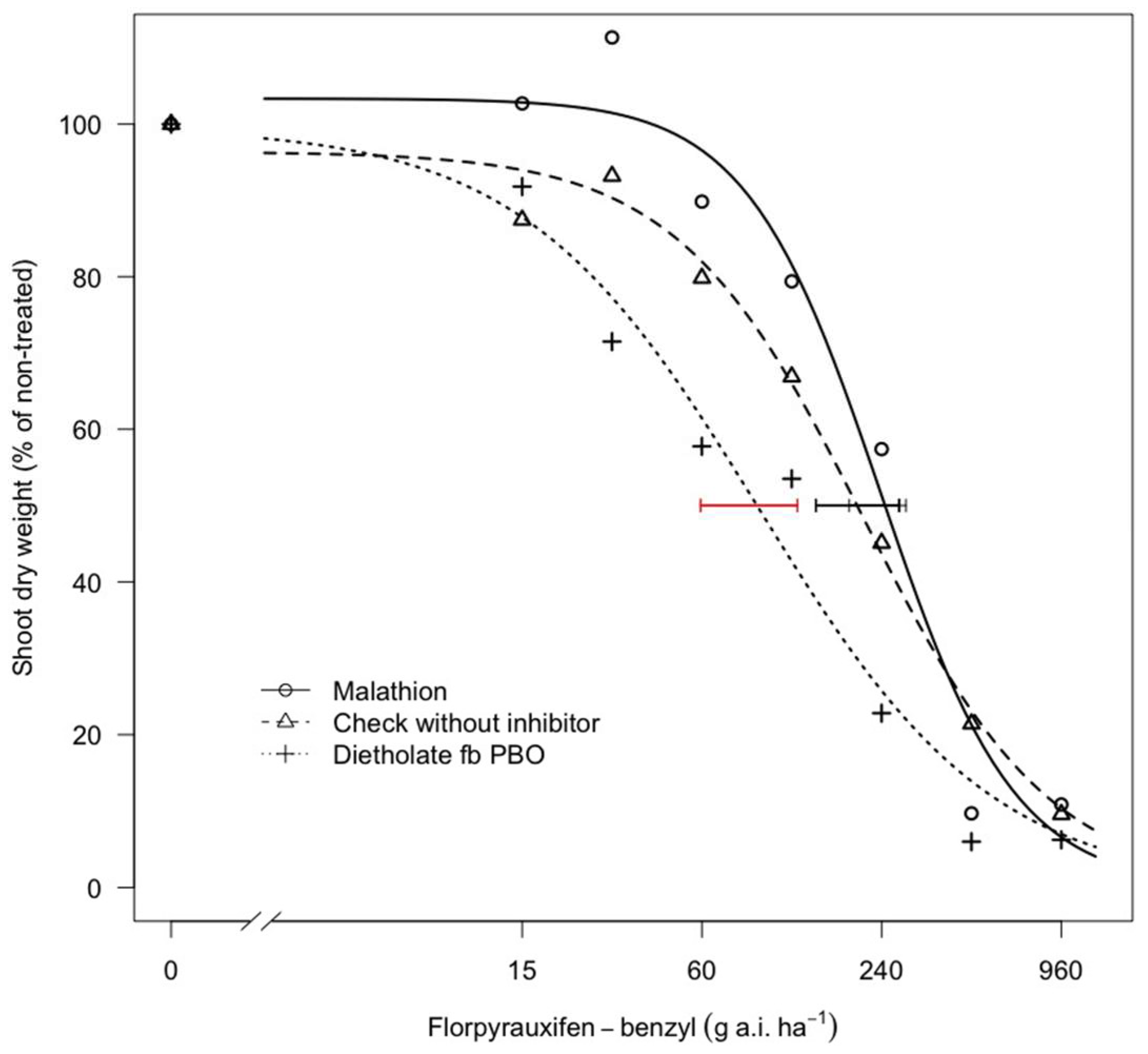
3.2. Florpyrauxifen-Benzyl Selectivity to Rice, as Affected by P450 Inhibitors, and the Tolerance of Two Cultivars
3.2.1. Effect of P450 Inhibitors on Rice Response to Florpyrauxifen-Benzyl
3.2.2. Rice Cultivar Response to Florpyrauxifen-Benzyl Application
3.3. Florpyrauxifen-Benzyl Selectivity to Rice, as Affected by Temperature Regimen and the Expression of Target Candidate Genes
3.3.1. Temperature and Rice Injury
3.3.2. Cytochrome P450 Monooxygenase Expression in Rice as Affected by Florpyrauxifen-Benzyl and Temperature
3.3.3. Glutathione S-Transferase Expression in Rice as Affected by Florpyrauxifen-Benzyl and Temperature
3.3.4. OsWAKL21.2 Expression in Rice as Affected by Florpyrauxifen-Benzyl and Temperature
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Brim-DeForest, W.B.; Al-Khatib, K.; Fischer, A.J. Predicting Yield Losses in Rice Mixed-Weed Species Infestations in California. Weed Sci. 2017, 65, 61–72. [Google Scholar] [CrossRef] [Green Version]
- Kraehmer, H.; Jabran, K.; Mennan, H.; Chauhan, B.S. Global distribution of rice weeds—A review. Crop Prot. 2016, 80, 73–86. [Google Scholar] [CrossRef]
- Peterson, M.A.; McMaster, S.A.; Riechers, D.E.; Skelton, J.; Stahlman, P.W. 2,4-D Past, Present, and Future: A Review. Weed Technol. 2016, 30, 303–345. [Google Scholar] [CrossRef]
- Grossmann, K.; Kwiatkowski, J. Selective induction of ethylene and cyanide biosynthesis appears to be involved in the selectivity of the herbicide quinclorac between rice and barnyardgrass. J. Plant Physiol. 1993, 142, 457–466. [Google Scholar] [CrossRef]
- Heap, I.M. International Survey of Herbicide Resistant Weeds. Available online: www.weedscience.org (accessed on 11 March 2021).
- Pannell, D.J.; Tillie, P.; Rodríguez-Cerezo, E.; Ervin, D.; Frisvold, G.B. Herbicide resistance: Economic and environmental challenges. AgBioForum 2016, 19, 136–155. [Google Scholar]
- Epp, J.B.; Alexander, A.L.; Balko, T.W.; Buysse, A.M.; Brewster, W.K.; Bryan, K.; Daeuble, J.F.; Fields, S.C.; Gast, R.E.; Green, R.A.; et al. The discovery of ArylexTM active and RinskorTM active: Two novel auxin herbicides. Bioorg. Med. Chem. 2016, 24, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Ryan Miller, M.; Norsworthy, J.K.; Scott, R.C. Evaluation of florpyrauxifen-benzyl on herbicide-resistant and herbicide-susceptible barnyardgrass accessions. Weed Technol. 2018, 32, 126–134. [Google Scholar] [CrossRef]
- Travlos, I.; Kanatas, P.; Tsekoura, A.; Gazoulis, I.; Papastylianou1, P.; Kakabouki, I.; Antonopoulos, N. Effcacy of Different Herbicides on Echinochloa colona (L.) Link Control and the First Case of Its Glyphosate Resistance in Greece. Agronomy 2020, 10, 1056. [Google Scholar] [CrossRef]
- Grossmann, K. Auxin herbicides: Current status of mechanism and mode of action. Pest Manag. Sci. 2009, 66, 113–120. [Google Scholar] [CrossRef]
- Hagen, G.; Guilfoyle, T. Auxin-responsive gene expression: Genes, promoters and regulatory factors. Plant Mol. Biol. 2002, 49, 373–385. [Google Scholar] [CrossRef]
- Kraft, M.; Kuglitsch, R.; Kwiatkowski, J.; Frank, M.; Grossmann, K. Indole-3-acetic acid and auxin herbicides up-regulate 9-cis-epoxycarotenoid dioxygenase gene expression and abscisic acid accumulation in cleavers (Galium aparine): Interaction with ethylene. J. Exp. Bot. 2007, 58, 1497–1503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Sundaram, S.; Armitage, L.; Evans, J.P.; Hawkes, T.; Kepinski, S.; Ferro, N.; Napier, R.M. Defining binding efficiency and specificity of auxins for SCF TIR1/AFB-Aux/IAA Co-receptor complex formation. ACS Chem. Biol. 2014, 9, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Prigge, M.J.; Greenham, K.; Zhang, Y.; Santner, A.; Castillejo, C.; Mutka, A.M.; O’Malley, R.C.; Ecker, J.R.; Kunkel, B.N.; Estelle, M. The Arabidopsis auxin receptor F-box proteins AFB4 and AFB5 are required for response to the synthetic auxin picloram. G3 Genes Genomes Genet. 2016, 6, 1383–1390. [Google Scholar] [CrossRef] [Green Version]
- Bell, J.; Schmitzer, P.; Weimer, M.; Napier, R.; Prusinska, J. Mode-of-action analysis of a new arylpicolinate herbicide from Dow AgroSciences. In Proceedings of the Weed Science Society of America Annual Meeting; Weed Science Society of America: Lexington, KY, USA, 2015. [Google Scholar]
- de Carvalho, S.J.P.; Nicolai, M.; Ferreira, R.R.; Figueira, A.V.d.O.; Christoffoleti, P.J. Herbicide selectivity by differential metabolism: Considerations for reducing crop damages. Sci. Agric. 2009, 66, 136–142. [Google Scholar] [CrossRef] [Green Version]
- Beste, C.E. Herbicide Handbook of the Weed Science Society of America, 5th ed.; Weed Science Society of America: Lexington, KY, USA, 1983. [Google Scholar]
- Jacobsen, R.E.; Fantke, P.; Trapp, S. Analysing half-lives for pesticide dissipation in plants. SAR QSAR Environ. Res. 2015, 26, 325–342. [Google Scholar] [CrossRef] [Green Version]
- Vijay, N.; Riechers, D.E.; Ferhatoglu, Y.; Barrett, M.; Duke, S.O.; Dayan, F.E.; Goldberg-Cavalleri, A.; Tétard-Jones, C.; Wortley, D.J.; Onkokesung, N.; et al. Herbicide Metabolism: Crop Selectivity, Bioactivation, Weed Resistance, and Regulation. Weed Sci. 2019, 67, 149–175. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.R.; Norsworthy, J.K. Influence of Soil Moisture on Absorption, Translocation, and Metabolism of Florpyrauxifen-benzyl. Weed Sci. 2018, 66, 418–423. [Google Scholar] [CrossRef]
- Todd, O.E.; Figueiredo, M.R.A.; Morran, S.; Soni, N.; Preston, C.; Kubeš, M.F.; Napier, R.; Gaines, T.A. Synthetic auxin herbicides: Finding the lock and key to weed resistance. Plant Sci. 2020, 300, 110631. [Google Scholar] [CrossRef]
- Arena, M.; Auteri, D.; Barmaz, S.; Brancato, A.; Brocca, D.; Bura, L.; Carrasco Cabrera, L.; Chaideftou, E.; Chiusolo, A.; Civitella, C.; et al. Peer review of the pesticide risk assessment of the active substance florpyrauxifen (variant assessed florpyrauxifen-benzyl). EFSA J. 2018, 16, 5378. [Google Scholar] [CrossRef]
- Mora, A.D.; Rosario, J.; Rojano-Delgado, A.M.; Palma-Bautista, C.; Torra, J.; Alcántara-De La Cruz, R.; De Prado, R. Multiple Resistance to Synthetic Auxin Herbicides and Glyphosate in Parthenium hysterophorus Occurring in Citrus Orchards. J. Agric. Food Chem. 2019, 67, 10010–10017. [Google Scholar] [CrossRef]
- Torra, J.; Rojano-Delgado, A.M.; Rey-Caballero, J.; Royo-Esnal, A.; Salas, M.L.; De Prado, R. Enhanced 2,4-D metabolism in two resistant papaver rhoeas populations from Spain. Front. Plant Sci. 2017, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Linscott, D.L.; Hagin, R.D.; Dawson, J.E. Conversion of 4- (2,4-Dichlorophenoxy) Butyric Acid to Homologs by Alfalfa. J. Agric. Food Chem. 1966, 16, 2–6. [Google Scholar]
- Anonymous Specimen Label Loyant 2021. Available online: https://www.corteva.us/products-and-solutions/crop-protection/loyant.html (accessed on 1 June 2021).
- Wells, G.S.; Taylor, M. RinskorTM Active—A new herbicide for rice weed control in Australia. In Proceedings of the Twentieth Australasian Weeds Conference; Randall, R., Lloyd, S., Borger, C., Eds.; Weeds Society of Western Australia: Perth, Australia, 2016; pp. 269–271. [Google Scholar]
- Hapukotuwa, N.K.; Perea, M.S.K.K.; Abeysekara, A.S.K.; Weligamage, S.S.; Piyatissa, U.B. Weed control efficacy of a new post-emergence herbicide ‘florpyrauxifen-benzyl’ 2.5 ec (Rinskor) in wet-seeded rice in Sri Lanka. Trop. Agric. 2018, 166, 141–152. [Google Scholar]
- Valle, N.D.; Carone, D.C.; Mascanzoni, E.; Baino, M. Florpyrauxifen-benzyl (Rinskor Active): New post-emergence herbicide for rice. J. Chem. Inf. Model. 2012, 53, 1689–1699. [Google Scholar] [CrossRef]
- Rubin, R.S.; Bundt, A.D.C.; Neves, R.; Kalsing, A. Controle de ciperáceas em arroz irrigado, utilizando novo herbicida RinskorTM Active. In Proceedings of the X CBAI Manejo de Plantas Daninhas; Sociedade Brasileira da Ciência das Plantas Daninhas: Balneario Camboriú, Brazil, 2017. [Google Scholar]
- Bundt, A.D.C.; Rubin, R.d.S.; Kalsing, A.; Neves, R.; Carranza, N. Eficiência do ingrediente ativo RinskorTM para controle de capim arroz na cultura do arroz irrigado. In Proceedings of the X CBAI Manejo de Plantas Daninhas; Sociedade Brasileira da Ciência das Plantas Daninhas: Balneario Camboriú, Brazil, 2017. [Google Scholar]
- Rubin, R.S.; Bundt, A.D.C.; Kalsing, A.; Neves, R.; Santos, W. Eficiência do herbicida RinskorTM active no controle de Sagittaria montevidensis e Aeschynomene denticulata na cultura do arroz irrigado. In Proceedings of the X CBAI Manejo de Plantas Daninhas; Sociedade Brasileira da Ciência das Plantas Daninhas: Balneario Camboriú, Brazil, 2017. [Google Scholar]
- Bundt, A.D.C.; Rubin, R.d.S.; Daltro, F.; Neves, R.; Menezes, V. Eficácia do ingrediente ativo RinskorTM em diferentes manejos de irrigação. In Proceedings of the X CBAI Manejo de Plantas Daninhas; Sociedade Brasileira da Ciência das Plantas Daninhas: Balneario Camboriú, Brazil, 2017. [Google Scholar]
- Wright, H.E.; Norsworthy, J.K.; Roberts, T.L.; Scott, R.; Hardke, J.; Gbur, E.E. Characterization of Rice Cultivar Response to Florpyrauxifen-benzyl. Weed Technol. 2020, 35, 1–35. [Google Scholar] [CrossRef]
- Wright, H.E.; Norsworthy, J.K.; Roberts, T.L.; Scott, R.; Hardke, J.; Gbur, E.E. Rice Cultivar Response to Florpyrauxifen-Benzyl When Applied with Imazethapyr and a Cytochrome P450 Inhibitor. Int. J. Agron. 2020, 2020, 2769610. [Google Scholar] [CrossRef] [PubMed]
- Counce, P.A.; Keisling, T.C.; Mitchell, A.J. A uniform, objectives, and adaptive system for expressing rice development. Crop Sci. 2000, 40, 436–443. [Google Scholar] [CrossRef] [Green Version]
- Royston, P. Approximating the Shapiro-Wilk W-test for Non-normality. Stat. Comput. 1992, 2, 117–119. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Package Lme4: Linear Mixed-Effects Models Using Eigen and S4. R Package Version. 67. Available online: https://www.researchgate.net/publication/279236477_Package_Lme4_Linear_Mixed-Effects_Models_Using_Eigen_and_S4 (accessed on 10 December 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 10 December 2021).
- Oliveira, M.C. Mixed Model ANOVA. Available online: https://esalq-weed-r.netlify.app/aulas/anova/anova.html (accessed on 5 July 2021).
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-response analysis using R. PLoS ONE 2015, 10, e0146021. [Google Scholar] [CrossRef] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Jain, M.; Nijhawan, A.; Tyagi, A.K.; Khurana, J.P. Validation of housekeeping genes as internal control for studying gene expression in rice by quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2006, 345, 646–651. [Google Scholar] [CrossRef]
- Pantone, D.J.; Baker, J.B. Varietal Tolerance of Rice (Oryza sativa) to Bromoxynil and Triclopyr at Different Growth Stages. Weed Technol. 1992, 6, 968–974. [Google Scholar] [CrossRef]
- Johnston, C.R.; Malladi, A.; Vencill, W.K.; Grey, T.L.; Stanley Culpepper, A.; Henry, G.; Czarnota, M.A.; Randell, T.M. Investigation of physiological and molecular mechanisms conferring diurnal variation in auxinic herbicide efficacy. PLoS ONE 2020, 15, e0238144. [Google Scholar] [CrossRef]
- Xu, J.; Wang, X.Y.; Guo, W.Z. The cytochrome P450 superfamily: Key players in plant development and defense. J. Integr. Agric. 2015, 14, 1673–1686. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, R.P.; Luster, D.G. Multiple forms of plant cytochromes P-450. Plant Physiol. 1991, 96, 669–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bond, J.A.; Walker, T.W. Differential Tolerance of Clearfield Rice Cultivars to Imazamox. Weed Technol. 2011, 25, 192–197. [Google Scholar] [CrossRef]
- Hirose, S.; Kawahigashi, H.; Tagiri, A.; Imaishi, H.; Ohkawa, H.; Ohkawa, Y. Tissue-specific expression of rice CYP72A21 induced by auxins and herbicides. Plant Biotechnol. Rep. 2007, 1, 27–36. [Google Scholar] [CrossRef]
- Rouse, C.E. Characterization of Multiple-Herbicide-Resistant Echinochloa colona from Arkansas; University of Arkansas: Fayetteville, Arkansas, 2017. [Google Scholar]
- Benedetti, L.; Rangani, G.; Ebeling Viana, V.; Carvalho-Moore, P.; Merotto, A.; Rabaioli Camargo, E.; Antonio de Avila, L.; Roma-Burgos, N. Rapid Reduction of Herbicide Susceptibility in Junglerice by Recurrent Selection with Sublethal Dose of Herbicides and Heat Stress. Agronomy 2020, 10, 1761. [Google Scholar] [CrossRef]
- Benedetti, L.; Rangani, G.; Viana, V.E.; Carvalho-Moore, P.; Camargo, E.R.; de Avila, L.A.; Roma-Burgos, N. Recurrent selection by herbicide sublethal dose and drought stress results in rapid reduction of herbicide sensitivity in junglerice. Agronomy 2020, 10, 1619. [Google Scholar] [CrossRef]
- Hu, T. A glutathione s-transferase confers herbicide tolerance in rice. Crop Breed. Appl. Biotechnol. 2014, 14, 76–81. [Google Scholar] [CrossRef] [Green Version]
- DeRidder, B.P.; Goldsbrough, P.B. Organ-specific expression of glutathione S-transferases and the efficacy of herbicide safeners in Arabidopsis. Plant Physiol. 2006, 140, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Baek, Y.S.; Goodrich, L.V.; Brown, P.J.; James, B.T.; Moose, S.P.; Lambert, K.N.; Riechers, D.E. Transcriptome Profiling and Genome-Wide Association Studies Reveal GSTs and Other Defense Genes Involved in Multiple Signaling Pathways Induced by Herbicide Safener in Grain Sorghum. Front. Plant Sci. 2019, 10, 3800–3805. [Google Scholar] [CrossRef]
- Chen, W.; Wu, L.; Wang, J.; Yu, Q.; Bai, L.; Pan, L. Quizalofop-p-ethyl resistance in Polypogon fugax involves glutathione S-transferases. Pest Manag. Sci. 2020, 76, 3800–3805. [Google Scholar] [CrossRef]
- Nishimura, K.; Nakagawa, R.; Hachisuga, C.; Munekage, Y.N. Deciphering the proteotoxic stress responses triggered by the perturbed thylakoid proteostasis in arabidopsis. Plants 2021, 10, 519. [Google Scholar] [CrossRef] [PubMed]
- Kohorn, B.D. Cell wall-associated kinases and pectin perception. J. Exp. Bot. 2016, 67, 489–494. [Google Scholar] [CrossRef] [Green Version]
- Malukani, K.K.; Ranjan, A.; Jyothi, H.S.; Patel, H.K.; Sonti1, R.V. The dual function receptor kinase, OsWAKL21.2, is involved in elaboration of lipaseA/esterase induced immune responses in rice. bioRxiv 2019. [Google Scholar] [CrossRef]
- Ruiz-Santaella, J.P.; Heredia, A.; De Prado, R. Basis of selectivity of cyhalofop-butyl in Oryza sativa L. Planta 2006, 223, 191–199. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Factor | Pr (>F) | |||||
|---|---|---|---|---|---|---|
| 7 DAA 1 | 14 DAA | 21 DAA | 28 DAA | 35 DAA | 42 DAA | |
| Year | 0.1325 | 0.2345 | 4 × 10−11 | 0.2744 | 0.0953 | 0.0928 |
| Planting time | 1 × 10−11 | 0.070 | 0.044 | 0.010 | 0.002 | 0.004 |
| Spraying time | 2 × 10−16 | 2 × 10−16 | 2 × 10−16 | 2 × 10−13 | 3 × 10−12 | 1 × 10−10 |
| Rate | 2 × 10−15 | 2 × 10−16 | 2 × 10−16 | 1 × 10−15 | 4 × 10−12 | 1 × 10−10 |
| Planting time × Spraying time | 2 × 10−12 | 3 × 10−4 | 0.028 | 0.080 | 0.225 | 0.194 |
| Planting time × Rate | 1 × 10−5 | 0.393 | 0.267 | 0.022 | 0.039 | 0.169 |
| Spraying time × Rate | 3 × 10−10 | 5 × 10−12 | 8 × 10−15 | 2 × 10−7 | 3 × 10−6 | 9 × 10−6 |
| Planting time × Spraying time × Rate | 7 × 10−6 | 0.058 | 0.027 | 0.238 | 0.338 | 0.539 |
| Treatments | B 1 | SE 2 | Dry Shoot Weight (g plant−1) | GR50 4 | CI 5 95% | Inhibition Ratio (%) 6 | p-Value 7 | |
|---|---|---|---|---|---|---|---|---|
| D 3 | SE | g a.i. ha−1 | ||||||
| Check without inhibitor | 1.4 | (0.3) | 4.4 | (0.2) | 216.9 | (144.5–274.7) | ||
| Malathion | 2.0 | (0.4) | 4.0 | (0.2) | 243.0 | (186.9–289.6) | 0.0 | 0.573 |
| Dietholate fb PBO | 1.1 | (0.2) | 3.5 | (0.2) | 89.6 | (59.5–125.4) | 63.1 | 0.031 |
| Cultivar | B 1 | SE 2 | Rice Plant Injury (%) | ED50 4 | CI 95% 5 | p-Value 6 | |
|---|---|---|---|---|---|---|---|
| D 3 | SE | g a.i. ha−1 | |||||
| IRGA 424 RI | −2.1 | (0.2) | 100 | (3.7) | 205.5 | (181.2–229.8) | |
| BRS Pampeira | −1.6 | (0.1) | 100 | (3.2) | 88.0 | (75.4–100.5) | 0.000 |
| Dry shoot weight (g) | GR50 5 | CI 95% | |||||
| IRGA 424 RI | 2.1 | (0.5) | 4.6 | (0.2) | 267.2 | (204.9–329.4) | |
| BRS Pampeira | 1.3 | (0.2) | 4.5 | (0.3) | 88.3 | (54.2–122.5) | 0.004 |
| Treatment 1 | Rate 2 | Rice Plant Injury (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 DAA 3 | 7 DAA | 14 DAA | 21 DAA | 28 DAA | |||||||
| Non-treated | 0 | 0 | h | 0 | h | 0 | e | 0 | d | 0 | f |
| T1 (all medium) | 30 | 3.3 | fg | 5.5 | efg | 2.8 | de | 4.0 | c | 1.8 | f |
| 60 | 6.5 | cde | 11.3 | b | 11.0 | bc | 10.4 | b | 2.3 | f | |
| T2 (med-low-med) | 30 | 3.5 | f | 3.9 | fg | 6.1 | cde | 2.6 | cd | 1.9 | f |
| 60 | 4.0 | ef | 7.2 | cdef | 5.3 | cde | 1.7 | cd | 1.7 | f | |
| T3 (high-low-med) | 30 | 2.4 | fgh | 4.0 | efg | 8.1 | cd | 4.6 | c | 3.3 | ef |
| 60 | 5.3 | def | 6.6 | defg | 10.6 | bc | 4.3 | c | 2.5 | f | |
| T4 (med-high-high) | 30 | 8.9 | bc | 7.6 | cde | 16.5 | b | 10.0 | b | 6.9 | de |
| 60 | 14.4 | a | 20.5 | a | 24.2 | a | 12.4 | ab | 11.9 | b | |
| T5 (med-high-med) | 30 | 8.2 | bcd | 3.2 | gh | 8.0 | cd | 10.8 | b | 7.8 | cd |
| 60 | 14.5 | a | 4.7 | efg | 5.6 | cde | 11.1 | b | 10.9 | bc | |
| T6 (med-high-low) | 30 | 7.8 | bcd | 10.8 | bc | 11.3 | bc | 13.5 | ab | 10.9 | bc |
| 60 | 10.0 | b | 9.4 | bcd | 16.4 | b | 14.6 | a | 21.9 | a | |
| Pr (>F) | |||||||||||
| Rate | 2.20 × 10−16 | 2.20 × 10−16 | 2.20 × 10−16 | 2.20 × 10−16 | 2.20 × 10−16 | ||||||
| Temperature treatment | 2.20 × 10−16 | 3.84 × 10−8 | 1.05 × 10−8 | 2.20 × 10−16 | 2.20 × 10−16 | ||||||
| Run | 2.20 × 10−16 | 0.0253 | 0.7306 | 0.4464 | 1.23 × 10−5 | ||||||
| Rate: Temperature treatment | 5.62 × 10−9 | 3.83 × 10−8 | 0.000121 | 3.50 × 10−12 | 2.20 × 10−16 | ||||||
| Florpyrauxifen-Benzyl | Log2-Fold Change of Gene Expression | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| T1 All Medium | T2 Med-Low | T3 High-Low | T4 Med-High | ||||||||
| Gene | Doses | 6 HAS 1 | 12 HAS | 24 HAS | 12 HAS | 24 HAS | 6 HAS | 12 HAS | 24 HAS | 12 HAS | 24 HAS |
| CYP71A21 | 0 | 3.4 (0.32) | −0.6 (0.02) | −3.1 (0.17) | 1.9 (1.06) | −1.9 (0.14) | −0.8 (0.35) | −4.3 (0.81) | −4.4 (1.66) | −3.4 (1.32) | −3.0 (0.46) |
| 30 | −0.9 (0.07) | 1.6 (0.21) | −2.1 (0.84) | −0.1 (0.08) | −4.0 (0.63) | 2.5 (0.25) | −7.6 (1.82) | −5.1 (1.22) | −4.6 (1.94) | −2.7 (0.91) | |
| 60 | 0.7 (0.13) | 1.8 (0.55) | −4.3 (1.00) | −4.9 (1.32) | −3.0 (1.28) | 0.6 (0.83) | −5.0 (1.12) | −3.4 (0.64) | −3.3 (0.66) | −4.6 (0.81) | |
| OsGSTL3 | 0 | 0.20 (0.37) | 0.3 (1.15) | −0.8 (0.44) | 0.6 (0.02) | −2.2 (0.32) | −2.3 (0.87) | −0.5 (0.68) | −3.3 (0.33) | −1.0 (0.67) | −6.1 (1.91) |
| 30 | −0.7 (0.18) | 1.1 (0.96) | −2.4 (1.31) | 0.4 (0.50) | −3.3 (1.12) | 0.9 (0.00) | −4.0 (1.35) | −3.6 (1.79) | −1.6 (0.30) | −5.4 (1.65) | |
| 60 | 2.9 (1.01) | 0.2 (0.18) | −3.0 (0.66) | −2.7 (0.59) | −2.7 (1.13) | −0.9 (0.16) | −2.0 (0.88) | −3.5 (0.85) | −1.5 (0.62) | −6.2 (1.37) | |
| WAKL21.2 | 0 | 6.9 (0.16) | 6.6 (0.81) | 4.9 (0.04) | 3.1 (0.10) | 4.5 (0.20) | 5.0 (0.41) | 4.6 (0.48) | −1.0 (0.98) | 5.7 (0.92) | 1.7 (0.73) |
| 30 | 3.4 (0.28) | 5.4 (0.89) | 2.7 (0.70) | 5.6 (1.16) | 0.5 (0.01) | 0.1 (0.00) | −1.2 (0.09) | −2.0 (0.51) | 3.5 (0.77) | 2.6 (0.34) | |
| 60 | 2.9 (0.90) | 5.4 (0.39) | 3.3 (0.78) | 5.0 (0.32) | 2.4 (0.65) | 1.7 (0.64) | 0.0 (0.56) | 3.0 (0.36) | 3.6 (1.20) | 0.6 (0.33) | |
| Scale | |||||||||||
| Expression scale (Log2-fold change) | <−8 | −8 to −4 | −4 to −2 | −2 to 0 | 0 to 2 | 2 to 4 | >4 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velásquez, J.C.; Bundt, A.D.C.; Camargo, E.R.; Andres, A.; Viana, V.E.; Hoyos, V.; Plaza, G.; de Avila, L.A. Florpyrauxifen-Benzyl Selectivity to Rice. Agriculture 2021, 11, 1270. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11121270
Velásquez JC, Bundt ADC, Camargo ER, Andres A, Viana VE, Hoyos V, Plaza G, de Avila LA. Florpyrauxifen-Benzyl Selectivity to Rice. Agriculture. 2021; 11(12):1270. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11121270
Chicago/Turabian StyleVelásquez, Juan Camilo, Angela Das Cas Bundt, Edinalvo Rabaioli Camargo, André Andres, Vívian Ebeling Viana, Verónica Hoyos, Guido Plaza, and Luis Antonio de Avila. 2021. "Florpyrauxifen-Benzyl Selectivity to Rice" Agriculture 11, no. 12: 1270. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11121270