
Spatial Distribution and Influencing Factors of Soil Fungi in a Degraded Alpine Meadow Invaded by Stellera chamaejasme

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Soil Sampling and Processing
2.3. DNA Extraction and Sequencing Analysis
2.4. Ecological and Statistical Analysis
2.5. Geostatistics and Interpolation Mapping
3. Results
3.1. Plant Cover, Soil Property, and Soil Fungal Characteristics
3.1.1. Plant Cover and Soil Property
3.1.2. Soil Fungal Characteristics
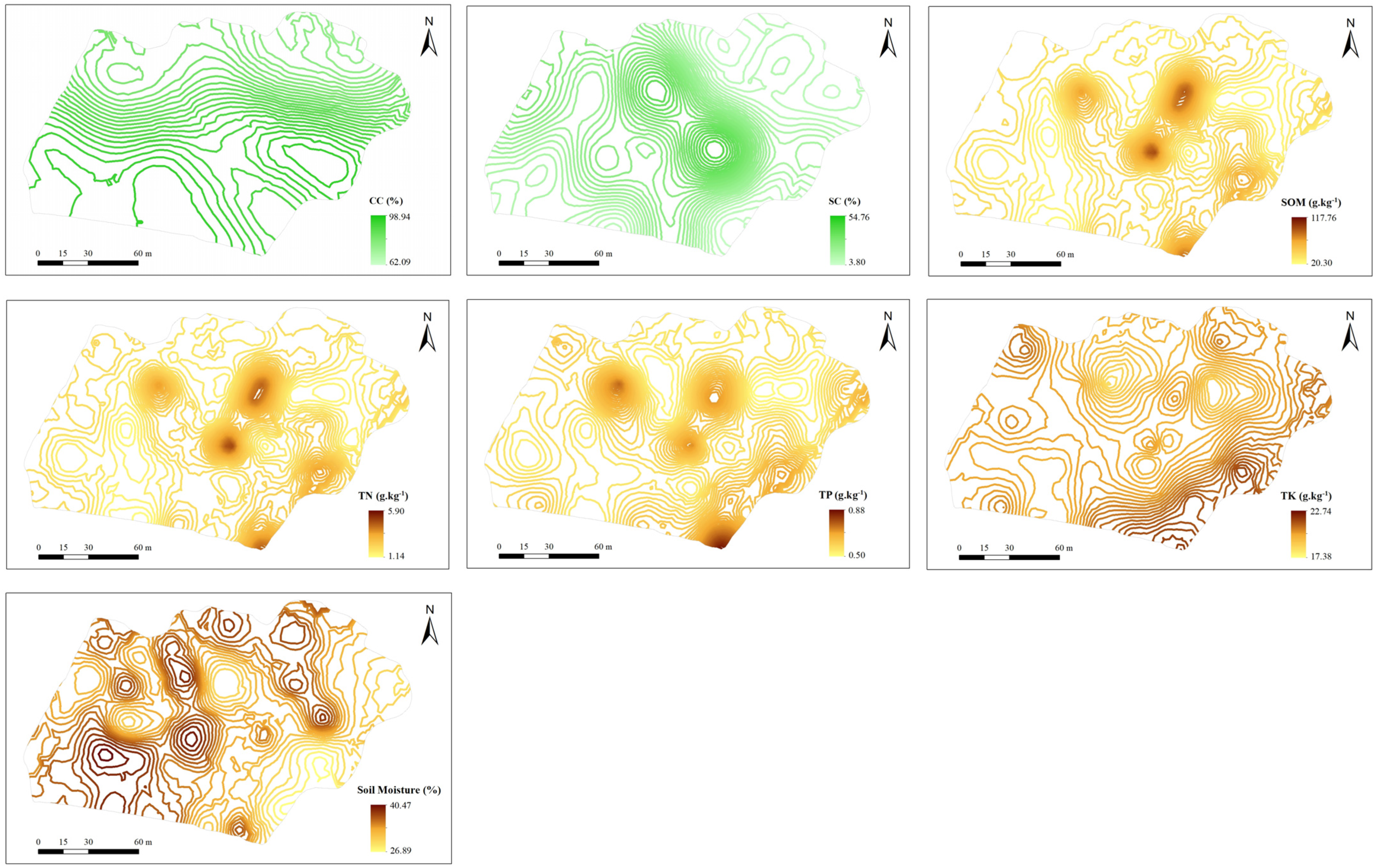
3.2. Mapping of Plant Cover and Soil Property
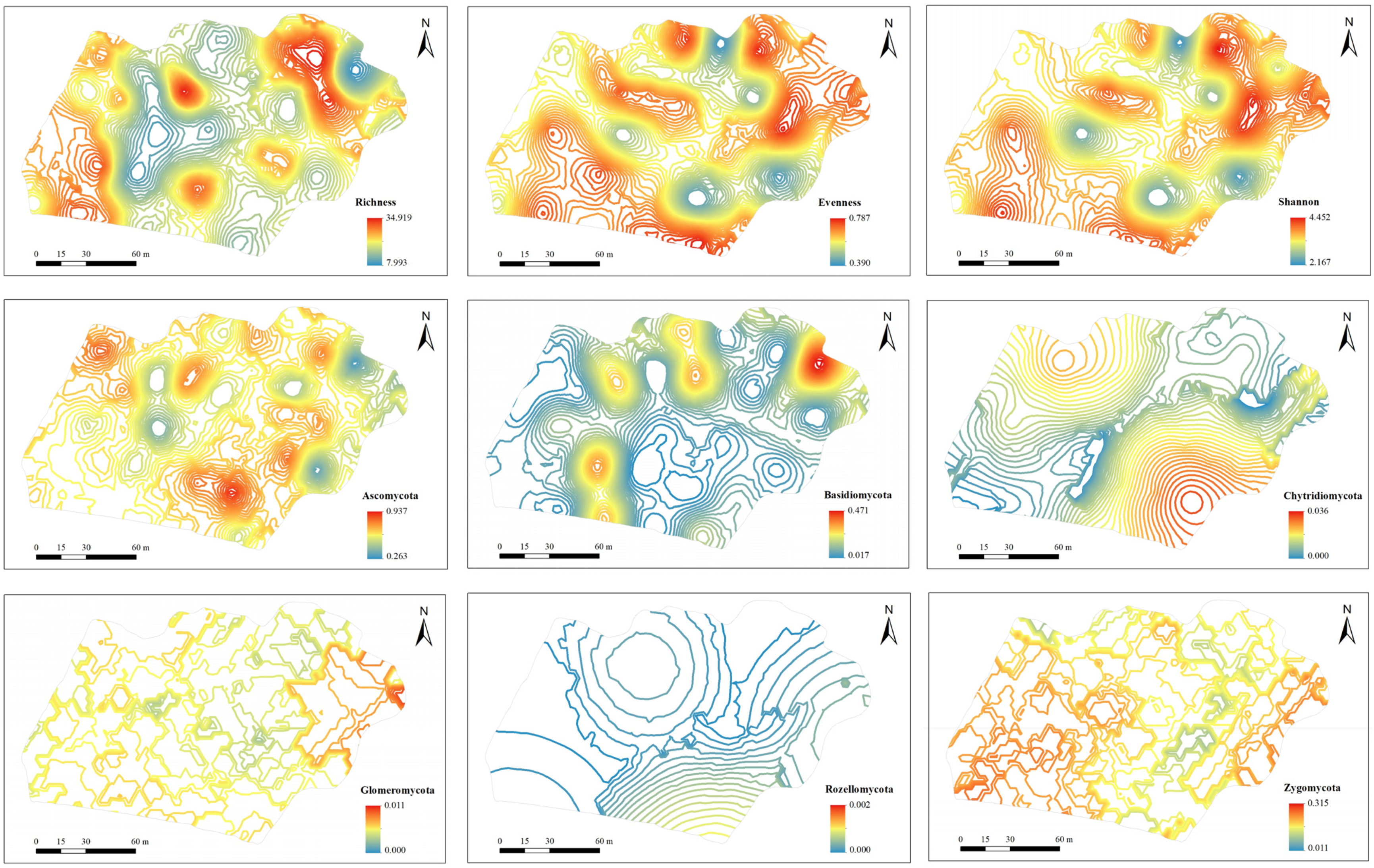
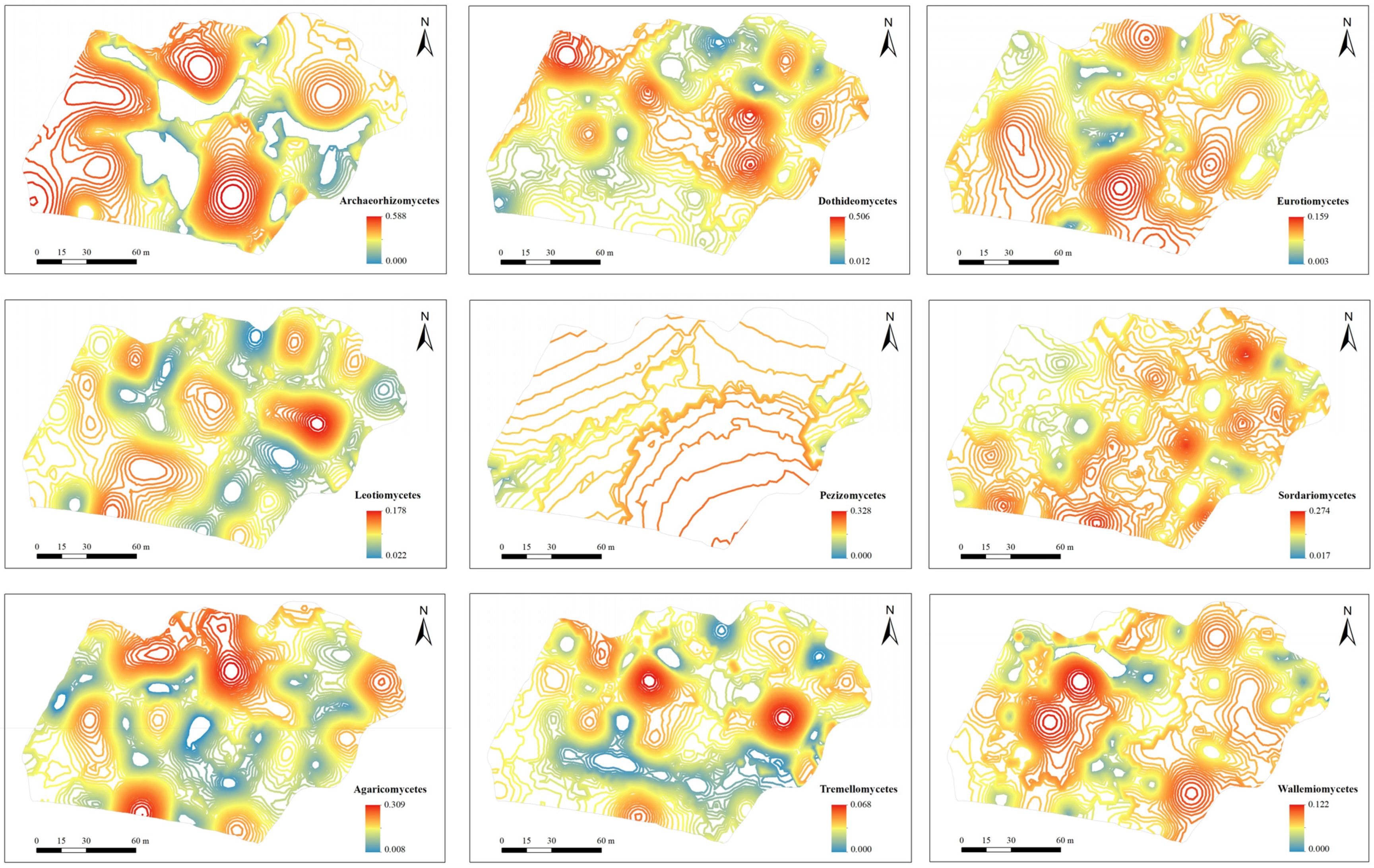
3.3. Mapping of Soil Fungal Diversity and Taxonomic Composition
3.4. Correlation between Plant–Soil Parameters and Soil Fungal Characteristics
4. Discussion
4.1. Spatial Distribution of Soil Fungal Characteristics and Linkage with Plant–Soil Parameters
4.2. Environmental Drivers of Soil Fungal Characteristics and Potential Impact of S. chamaejasme
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, Y.L.; Deng, Y.; Ding, J.Z.; Hu, H.; Xu, T.; Li, F.; Yang, G.B.; Yang, Y.H. Distinct microbial communities in the active and permafrost layers on the Tibetan Plateau. Mol. Ecol. 2017, 26, 6608–6620. [Google Scholar] [CrossRef] [PubMed]
- Martiny, J.B.H.; Bohannon, B.J.M.; Brown, J.H.; Colwell, R.K.; Fuhrman, J.A.; Green, J.L.; Horner-Devine, M.C.; Kane, M.; Krumins, J.A.; Kuske, C.R.; et al. Microbial biogeography: Putting microorganisms on the map. Nat. Rev. Microbiol. 2006, 4, 102–112. [Google Scholar] [CrossRef]
- Constancias, F.; Terrat, S.; Saby, N.P.A.; Horrigue, W.; Villerd, J.; Jean-Philippe, G.; Biju-Duval, L.; Nowak, V.; Dequiedt, S.; Ranjard, L.; et al. Mapping and determinism of soil microbial community distribution across an agricultural landscape. MicrobiologyOpen 2015, 4, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Karimi, B.; Terrat, S.; Dequiedt, S.; Saby, N.P.A.; Horrigue, W.; Lelievre, M.; Nowak, V.; Jolivet, C.; Arrouays, D.; Wincker, P.; et al. Biogeography of soil bacteria and archaea across France. Sci. Adv. 2018, 4, eaat1808. [Google Scholar] [CrossRef] [Green Version]
- Talbot, J.M.; Bruns, T.D.; Taylor, J.W.; Smith, D.P.; Branco, S.; Glassman, S.I.; Erlandson, S.; Vilgalys, R.; Liao, H.L.; Smith, M.E.; et al. Endemism and functional convergence across the North American soil mycobiome. Proc. Natl. Acad. Sci. USA 2014, 111, 6341–6346. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Guo, Z.Y.; Xu, A.A.; Wang, C.K.; Wu, S.W.; Pan, K.; Zhang, F.F.; Pan, X.Z. Spatial differences in bacterial communities preserved in soils archived for a decade. Appl. Soil Ecol. 2019, 142, 64–71. [Google Scholar] [CrossRef]
- Wang, C.T.; Long, R.J.; Wang, Q.J.; Ding, L.M.; Wang, M.P. Effects of altitude on plant-species diversity and productivity in an alpine meadow, Qinghai-Tibetan plateau. Aust. J. Bot. 2007, 55, 110–117. [Google Scholar] [CrossRef] [Green Version]
- Song, M.H.; Xu, X.L.; Hu, Q.W.; Tian, Y.Q.; OuYang, H.; Zhou, C.P. Interactions of plant species mediated plant competition for inorganic nitrogen with soil microorganisms in an alpine meadow. Plant Soil 2007, 297, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Adams, J.M.; Shi, Y.; He, J.S.; Jing, X.; Chen, L.T.; Tedersoo, L.; Chu, H.Y. Soil fungal diversity in natural grasslands of the Tibetan Plateau: Associations with plant diversity and productivity. New Phytol. 2017, 215, 756–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Che, R.X.; Wang, S.P.; Wang, Y.F.; Xu, Z.H.; Wang, W.J.; Rui, Y.C.; Wang, F.; Hu, J.M.; Tao, J.; Cui, X.Y. Total and active soil fungal community profiles were significantly altered by six years of warming but not by grazing. Soil Biol. Biochem. 2019, 139, 107611. [Google Scholar] [CrossRef]
- Wang, C.T.; Zhao, X.Q.; Zi, H.B.; Hu, L.; Ade, L.; Wang, G.X.; Lerdau, M. The effect of simulated warming on root dynamics and soil microbial community in an alpine meadow of the Qinghai-Tibet Plateau. Appl. Soil Ecol. 2017, 116, 30–41. [Google Scholar] [CrossRef]
- Yu, C.Q.; Han, F.S.; Fu, G. Effects of 7 years experimental warming on soil bacterial and fungal community structure in the Northern Tibet alpine meadow at three elevations. Sci. Total Environ. 2019, 655, 814–822. [Google Scholar] [CrossRef] [PubMed]
- Xue, R.; Yang, Q.; Miao, F.H.; Wang, X.Z.; Shen, Y.Y. Slope aspect influences plant biomass, soil properties and microbial composition in alpine meadow on the Qinghai-Tibetan Plateau. J. Soil Sci. Plant Nutr. 2018, 18, 1–12. [Google Scholar] [CrossRef]
- Che, R.X.; Wang, Y.F.; Li, K.X.; Xu, Z.H.; Hu, J.M.; Wang, F.; Rui, Y.C.; Li, L.F.; Pang, Z.; Cui, X.Y. Degraded patch formation significantly changed microbial community composition in alpine meadow soils. Soil Tillage Res. 2019, 195, 104426. [Google Scholar] [CrossRef]
- Liu, S.B.; Zamanian, K.; Schleuss, P.M.; Zarebanadkouki, M.; Kuzyakov, Y. Degradation of Tibetan grasslands: Consequences for carbon and nutrient cycles. Agric. Ecosyst. Environ. 2018, 252, 93–104. [Google Scholar] [CrossRef]
- He, S.Y.; Richards, K. Impact of meadow degradation on soil water status and pasture management-a case study in Tibet. Land Degrad. Dev. 2015, 26, 468–479. [Google Scholar]
- Che, R.X.; Wang, F.; Wang, W.J.; Zhang, J.; Zhao, X.; Rui, Y.C.; Xu, Z.H.; Wang, Y.F.; Hao, Y.B.; Cui, X.Y. Increase in ammonia-oxidizing microbe abundance during degradation of alpine meadows may lead to greater soil nitrogen loss. Biogeochemistry 2017, 136, 341–352. [Google Scholar] [CrossRef]
- Lin, L.; Li, Y.K.; Xu, X.L.; Zhang, F.W.; Du, Y.G.; Liu, S.L.; Guo, X.W.; Cao, G.M. Predicting parameters of degradation succession processes of Tibetan Kobresia grasslands. Solid Earth 2015, 6, 1237–1246. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, X.T.; Liu, G.B.; Zhang, C.; Wang, G.L. Bacterial richness is negatively related to potential soil multifunctionality in a degraded alpine meadow. Ecol. Indic. 2021, 121, 106996. [Google Scholar] [CrossRef]
- Li, Y.M.; Wang, S.P.; Jiang, L.L.; Zhang, L.R.; Cui, S.J.; Meng, F.D.; Wang, Q.; Li, X.N.; Zhou, Y. Changes of soil microbial community under different degraded gradients of alpine meadow. Agric. Ecosyst. Environ. 2016, 222, 213–222. [Google Scholar] [CrossRef]
- Li, S.X.; Wang, Y.L.; Wang, Y.Q.; Yin, Y.L. Response of soil bacterial community characteristics to alpine meadow degradation. Biodivers. Sci. 2021, 29, 53–64. (In Chinese) [Google Scholar]
- Ghorbanzadeh, N.; Salehi, A.; Pourbabaei, H.; Tolarod, A.A.S.; Alavi, S.J. Spatial variability of soil microbial indices in common alder COMMON ALDER (Alnus glutinosa) stands using a geostatistical approach in northern Iran. J. For. Res. 2019, 30, 679–688. [Google Scholar] [CrossRef]
- Harris, R.B. Rangeland degradation on the Qinghai-Tibetan plateau: A review of the evidence of its magnitude and causes. J. Arid Environ. 2010, 74, 1–12. [Google Scholar] [CrossRef]
- Shi, Z.C. The Poisonous Plants of Chinese Grassland; China Agriculture Press: Beijing, China, 1997; pp. 140–150. (In Chinese) [Google Scholar]
- Lu, R.K. Analytical Methods of Soil Agricultural Chemistry; China Agricultural Science Press: Beijing, China, 1999. (In Chinese) [Google Scholar]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.Q.; Zhu, S.L.; Zhou, C.L. Characteristics of spatial variability of soil thickness in China. Geogr. Res. 2001, 20, 161–169. (In Chinese) [Google Scholar]
- Zhou, J.; Jiang, X.; Zhou, B.; Zhao, B.; Ma, M.; Guan, D.; Li, J.; Chen, S.F.; Cao, F.M.; Shen, D.L.; et al. Thirty four years of nitrogen fertilization decreases fungal diversity and alters fungal community composition in black soil in NorthEast China. Soil Biol. Biochem. 2016, 57, 135–143. [Google Scholar] [CrossRef]
- Cambardella, C.A.; Moorman, T.B.; Novak, J.M.; Parkin, T.B.; Karlen, D.L.; Turco, R.F. Field-scale variability of soil properties in central lowa soils. Soil Sci. Soc. Am. J. 1994, 58, 1501–1511. [Google Scholar] [CrossRef]
- Shange, R.S.; Ankumah, R.O.; Ibekwe, A.M.; Zabawa, R.; Dowd, S.E. Distinct soil bacterial communities revealed under a diversely managed agroecosystem. PLoS ONE 2012, 7, e0040338. [Google Scholar] [CrossRef] [Green Version]
- He, D.; Xiang, X.J.; He, J.S.; Wang, C.; Cao, G.M.; Adams, J.; Chu, H.Y. Composition of the soil fungal community is more sensitive to phosphorus than nitrogen addition in the alpine meadow on the Qinghai-Tibetan Plateau. Biol. Fertil. Soils 2016, 52, 1059–1072. [Google Scholar] [CrossRef]
- Weber, C.F.; Vilgalys, R.; Kuske, C.R. Changes in fungal community composition in response to elevated atmospheric CO2 and nitrogen fertilization varies with soil horizon. Front Microbiol. 2013, 4, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, R.C.; Balasch, M.M.; Kuske, C.R. Contrasting soil fungal community responses to experimental nitrogen addition using the large subunit rRNA taxonomic marker and cellobiohydrolase I functional marker. Mol. Ecol. 2014, 23, 4406–4417. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Bahram, M.; Polme, S.; Koljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Fungal biogeography. Global diversity and geography of soil fungi. Science 2014, 346, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, G.S.; Wang, Y.Q.; Song, M.L.; Wang, H.S.; Yin, Y.L.; Liu, S.C.; Yang, Y.W.; Yang, M. Effects of Stellera chamaejasme patches on the surrounding grassland community and on soil physiochemical properties in degraded grasslands susceptible to S. chamaejasme invasion. Acta Prataculturae Sin. 2019, 28, 51–61. (In Chinese) [Google Scholar]
- Sun, G.; Luo, P.; Qiu, P.F.; Gao, Y.H.; Chen, H.; Shi, F.S. Stellera chamaejasme L. increases soil N availability, turnover rates and microbial biomass in an alpine meadow ecosystem on the eastern Tibetan Plateau of China. Soil Biol. Biochem. 2009, 41, 86–91. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Variable Type | Mean | SD | CV | Model | Nugget C0 | Sill C0+C | Nugget/Sill C0/(C0+C)/% | Range /m |
|---|---|---|---|---|---|---|---|---|
| plant cover | ||||||||
| CC (%) | 87.37 | 8.585 | 0.098 | G | 0.004 | 0.007 | 50.00 | 62.2 |
| SC (%) | 20.33 | 11.518 | 0.567 | S | 0.005 | 0.015 | 31.82 | 47.8 |
| soil property | ||||||||
| SOM (g.kg−1) | 40.20 | 19.823 | 0.493 | S | 0.001 | 0.691 | 0.15 | 27.3 |
| TN (g.kg−1) | 2.16 | 0.996 | 0.169 | S | 0.001 | 0.627 | 0.16 | 29.5 |
| TP (g.kg−1) | 0.59 | 0.073 | 0.124 | S | 0.004 | 0.112 | 3.41 | 24.9 |
| TK (g.kg−1) | 20.07 | 1.204 | 0.060 | E | 0.337 | 1.760 | 19.15 | 58.5 |
| SM (%) | 34.98 | 3.120 | 0.089 | S | 0.000 | 0.001 | 5.05 | 25.1 |
| Variable Type | Mean | SD | CV | Model | Nugget C0 | Sill C0+C | Nugget/Sill C0/(C0+C)/% | Range /m | |
|---|---|---|---|---|---|---|---|---|---|
| Alpha diversity | |||||||||
| richness | 23.927 | 5.988 | 0.250 | S | 0.5000 | 36.1633 | 1.38 | 27.0 | |
| evenness | 0.638 | 0.097 | 0.152 | S | 0.0001 | 0.0090 | 1.11 | 29.5 | |
| Shannon index | 3.528 | 0.578 | 0.164 | S | 0.0010 | 0.3367 | 0.30 | 29.2 | |
| Relative abundance of taxonomic composition | |||||||||
| Phylum | Class | ||||||||
| Ascomycota | 0.612 | 0.150 | 0.245 | S | 0.0015 | 0.0235 | 6.38 | 21.3 | |
| Archaeorhizomycetes | 0.064 | 0.128 | 2.001 | G | 0.0000 | 0.0168 | 0.00 | 27.7 | |
| Dothideomycetes | 0.137 | 0.114 | 0.832 | S | 0.0002 | 0.0130 | 1.54 | 23.7 | |
| Eurotiomycetes | 0.029 | 0.031 | 1.043 | S | 0.0001 | 0.0010 | 10.0 | 30.9 | |
| Leotiomycetes | 0.076 | 0.029 | 0.387 | G | 0.0000 | 0.0009 | 0.00 | 22.9 | |
| Pezizomycetes | 0.012 | 0.049 | 4.255 | G | 0.0017 | 0.0090 | 18.89 | 318.0 | |
| Sordariomycetes | 0.113 | 0.071 | 0.627 | S | 0.0000 | 0.0049 | 0.00 | 20.2 | |
| Basidiomycota | 0.119 | 0.103 | 0.866 | G | 0.0003 | 0.0104 | 2.88 | 25.3 | |
| Agaricomycetes | 0.066 | 0.073 | 1.109 | G | 0.0000 | 0.0058 | 0.00 | 22.7 | |
| Tremellomycetes | 0.012 | 0.014 | 1.185 | G | 0.0000 | 0.0002 | 0.00 | 18.9 | |
| Wallemiomycetes | 0.011 | 0.024 | 2.157 | G | 0.0000 | 0.0007 | 0.00 | 19.2 | |
| Chytridiomycota | 0.002 | 0.006 | 3.717 | E | 0.0000 | 0.0001 | 25.04 | 224.4 | |
| Glomeromycota | 0.001 | 0.002 | 1.287 | G | 0.0000 | 0.0000 | 61.34 | 26.3 | |
| Rozellomycota | 0.000 | 0.000 | 3.831 | G | 0.0000 | 0.0000 | 53.14 | 126.3 | |
| Zygomycota | 0.117 | 0.076 | 0.643 | G | 0.0000 | 0.0057 | 1.58 | 20.6 | |
| Variable Type | CC | SC | SOM | TN | TP | TK | SM |
|---|---|---|---|---|---|---|---|
| CC | 1 | ||||||
| SC | 0.22 | 1 | |||||
| SOM | 0.04 | −0.11 | 1 | ||||
| TN | 0.05 | −0.13 | 0.99 ** | 1 | |||
| TP | 0.13 | −0.18 | 0.85 ** | 0.88 ** | 1 | ||
| TK | 0.18 | −0.35 * | 0.47 ** | 0.51 ** | 0.66 ** | 1 | |
| SM | 0.06 | 0.13 | −0.27 | −0.27 | −0.34 * | −0.36 * | 1 |
| Variable Type | CC | SC | SOM | TN | TP | TK | SM |
|---|---|---|---|---|---|---|---|
| Alpha diversity | |||||||
| richness | 0.20 * | −0.06 | 0.06 | 0.03 | 0.05 | −0.01 | −0.05 |
| evenness | 0.02 | −0.05 | 0.13 | 0.14 | 0.15 | 0.17 * | 0.05 |
| Shannon diversity | 0.07 | −0.03 | 0.10 | 0.11 | 0.11 | 0.18 * | 0.05 |
| Relative abundance of soil fungal phylum | |||||||
| Ascomycota | 0.16 * | −0.09 | 0.20 * | 0.19 * | 0.16 | 0.07 | 0.04 |
| Basidiomycota | 0.26 * | −0.08 | 0.10 | 0.07 | 0.06 | −0.10 | −0.04 |
| Chytridiomycota | −0.05 | −0.05 | 0.04 | 0.05 | 0.19 | 0.20 | 0.17 |
| Glomeromycota | −0.10 | −0.09 | −0.10 | −0.10 | −0.08 | −0.12 | −0.01 |
| Rozellomycota | −0.07 | 0.05 | −0.06 | −0.06 | 0.12 | 0.22 | −0.03 |
| Zygomycota | −0.07 | −0.07 | −0.01 | −0.01 | −0.05 | −0.02 | 0.01 |
| Relative abundance of soil fungal class | |||||||
| Archaeorhizomycetes | 0.03 | 0.03 | −0.11 | −0.11 | −0.06 | −0.09 | −0.11 |
| Dothideomycetes | −0.03 | 0.01 | −0.13 | −0.13 | −0.11 | 0.11 | −0.08 |
| Eurotiomycetes | −0.04 | −0.02 | −0.10 | −0.11 | −0.08 | −0.07 | −0.02 |
| Leotiomycetes | −0.01 | −0.07 | 0.01 | 0.01 | −0.05 | −0.10 | −0.01 |
| Pezizomycetes | −0.38 | −0.07 | 0.07 | 0.07 | 0.20 | 0.22 * | 0.04 |
| Sordariomycetes | −0.02 | 0.08 | 0.01 | 0.02 | −0.04 | −0.03 | −0.01 |
| Agaricomycetes | −0.10 | −0.03 | −0.08 | −0.09 | −0.12 | −0.04 | −0.01 |
| Tremellomycetes | −0.06 | 0.13 | 0.02 | 0.03 | −0.07 | 0.16 | 0.04 |
| Wallemiomycetes | 0.01 | −0.11 | 0.52 ** | 0.49 ** | 0.50 ** | −0.01 | 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Zhao, F.; Wang, L.; He, W.; Liu, J.; Long, Y. Spatial Distribution and Influencing Factors of Soil Fungi in a Degraded Alpine Meadow Invaded by Stellera chamaejasme. Agriculture 2021, 11, 1280. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11121280
Liu Y, Zhao F, Wang L, He W, Liu J, Long Y. Spatial Distribution and Influencing Factors of Soil Fungi in a Degraded Alpine Meadow Invaded by Stellera chamaejasme. Agriculture. 2021; 11(12):1280. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11121280
Chicago/Turabian StyleLiu, Yongmei, Fan Zhao, Lei Wang, Wei He, Jianhong Liu, and Yongqing Long. 2021. "Spatial Distribution and Influencing Factors of Soil Fungi in a Degraded Alpine Meadow Invaded by Stellera chamaejasme" Agriculture 11, no. 12: 1280. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11121280