
Multiplex CRISPR/Cas9 Mutagenesis of BrVRN1 Delays Flowering Time in Chinese Cabbage (Brassica rapa L. ssp. pekinensis)


Abstract
:1. Introduction
2. Material and Methods
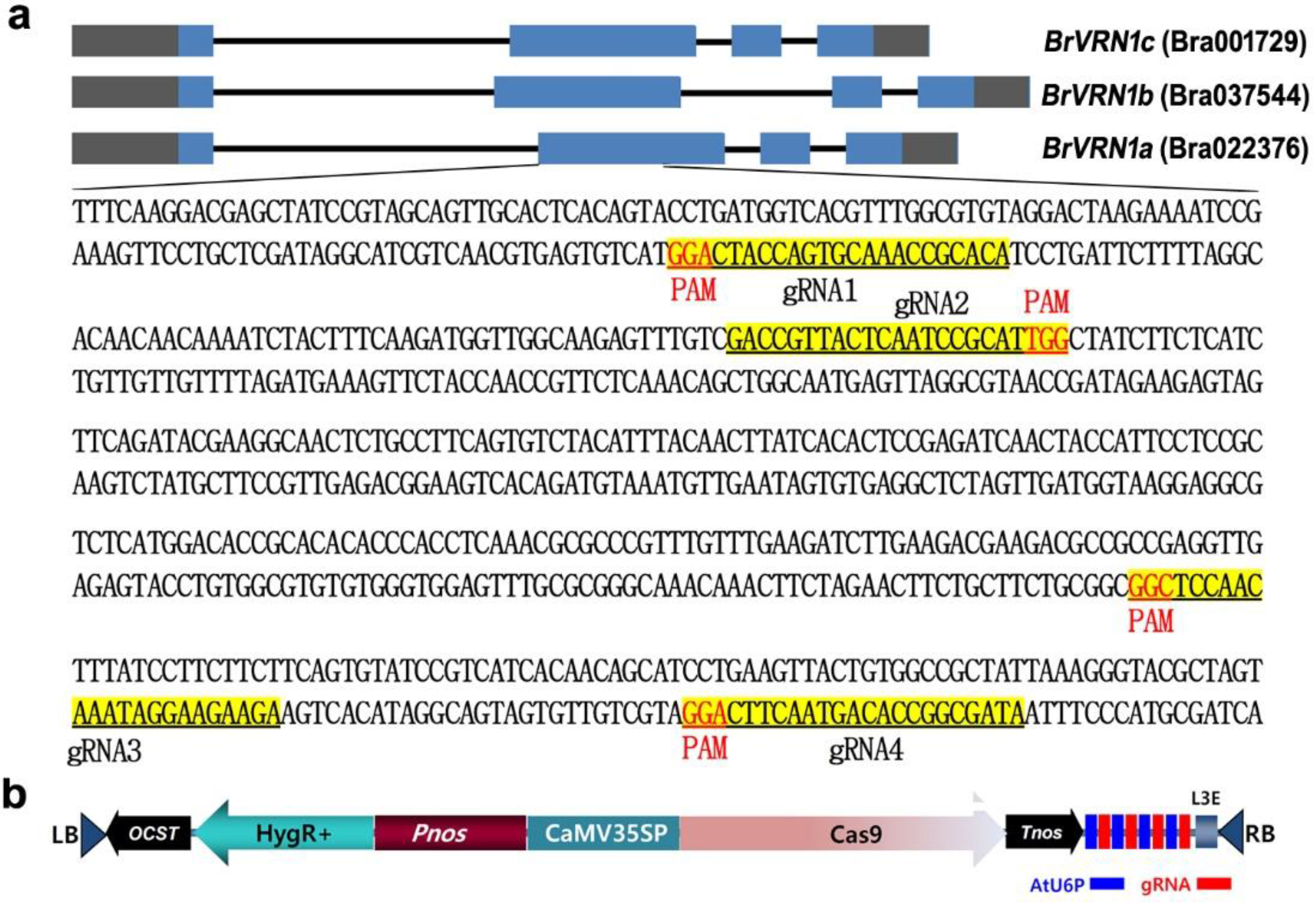
2.1. sgRNA Target Site Design and Vector Construction
2.2. Generation of Transgenic Chinese Cabbage Plants
2.3. Confirmation of the Mutations of Transgenic Plants
2.4. RNA Extraction and Real-Time Reverse Transcription Quantitative PCR (qPCR) Analysis
2.5. Flowering Time Analysis
3. Results
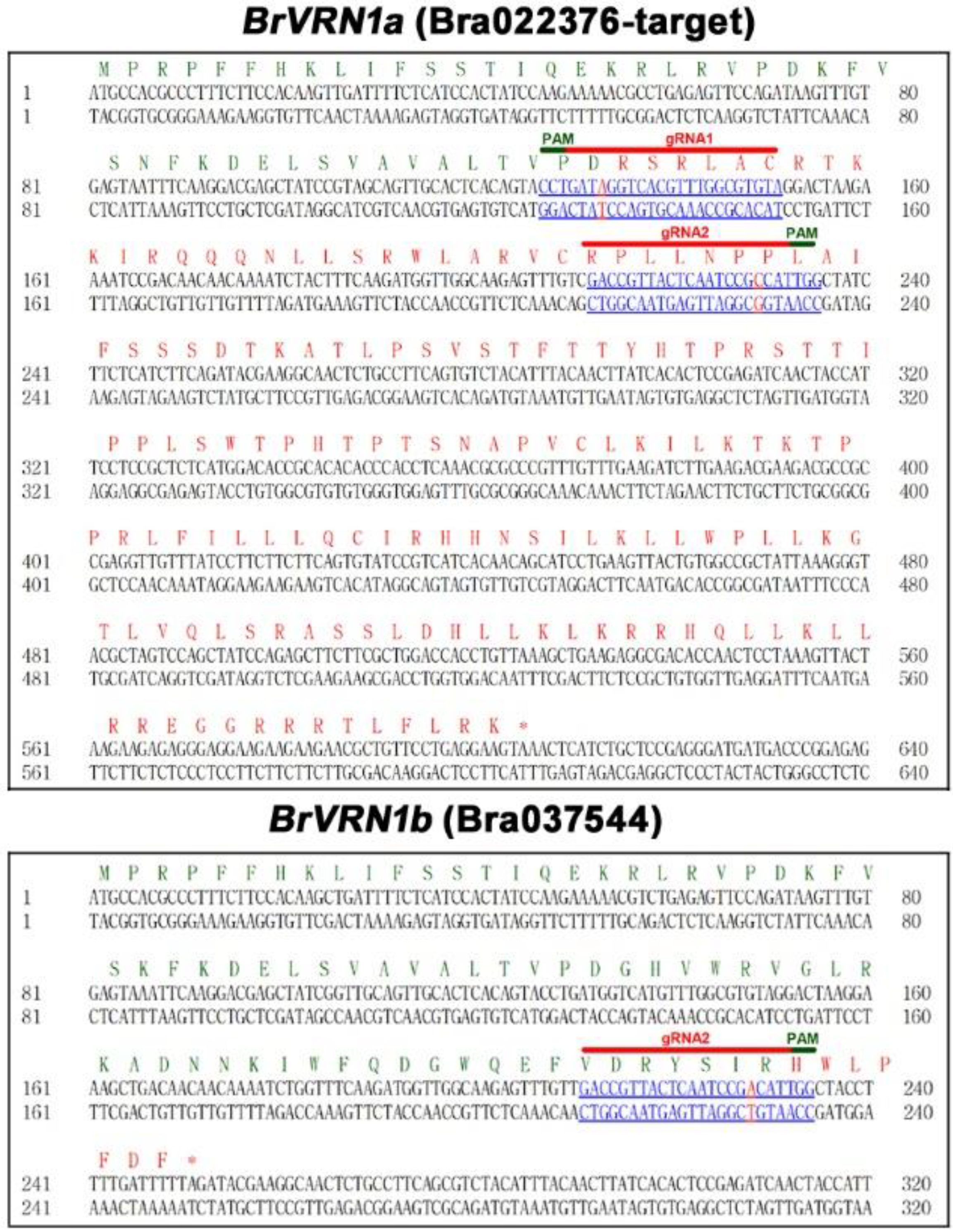
3.1. Identification and CRISPR-Cas9 Mutagenesis of BrVRN1 Genes
3.2. Generation of ‘Transgene-Free’ Mutant Lines
3.3. Stable Inheritance of Induced Mutations and Phenotypes of the Mutants
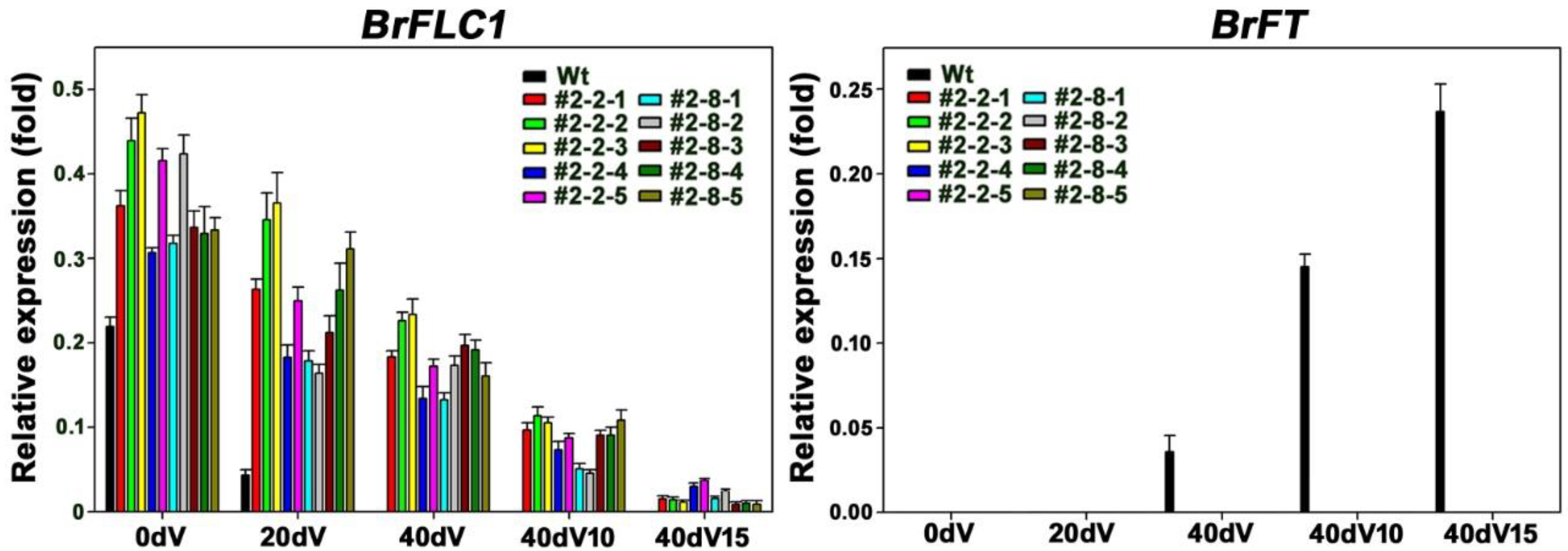
3.4. Expression Patterns of Floral Regulators in GE2 Mutant Plants
3.5. Analysis of Potential Off-Target Changes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jung, H.J.; Dong, X.; Park, J.I.; Thamilarasan, S.K.; Lee, S.S.; Kim, Y.K.; Lim, Y.P.; Nou, I.S.; Hur, Y. Genome-wide transcriptome analysis of two contrasting Brassica rapa doubled haploid lines under cold-stresses using Br135K oligomeric chip. PLoS ONE 2014, 9, e106069. [Google Scholar] [CrossRef]
- Li, J.; Zhao, X.; Nishimura, Y.; Fukumoto, Y. Correlation between bolting and physiological properties in Chinese cabbage (Brassica rapa L. Pekinensis group). J. Jpn. Soc. Hortic. Sci. 2010, 79, 294–300. [Google Scholar] [CrossRef] [Green Version]
- Elers, B.; Wiebe, H.-J. Flower formation of chinese cabbage. I. Response to vernalization and photoperiods. Sci. Hortic. 1984, 22, 219–231. [Google Scholar] [CrossRef]
- Mero, C.E.; Honma, S. Inheritance of bolting resistance in an intraspecific Chinese cabbage x turnip cross. HORTSCIENCE 1985, 20, 881–882. [Google Scholar]
- Jung, C.; Müller, A.E. Flowering time control and applications in plant breeding. Trends Plant Sci. 2009, 14, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Simpson, G.G.; Dean, D. Arabidopsis, the Resetta stone of flowering time? Science 2002, 296, 285–289. [Google Scholar] [CrossRef] [Green Version]
- Baurle, I.; Dean, C. The timing of developmental transitions in plants. Cell 2006, 125, 655–664. [Google Scholar] [CrossRef] [Green Version]
- Amasino, R. Vernalization, competence, and the epigenetic memory of winter. Plant Cell 2004, 16, 2553–2559. [Google Scholar] [CrossRef] [Green Version]
- Amasino, R. Seasonal and developmental timing of flowering. Plant J. 2010, 61, 1001–1013. [Google Scholar] [CrossRef]
- Shea, D.J.; Itabashi, E.; Takada, S.; Fukai, E.; Kakizaki, T.; Fujimoto, R.; Okazaki, K. The role of FLOWERING LOCUS C in vernalization of Brassica: The importance of vernalization research in the face of climate change. Crop Pasture Sci. 2018, 69, 30–39. [Google Scholar] [CrossRef]
- Su, T.; Wang, W.; Li, P.; Zhang, B.; Li, P.; Xin, X.; Sun, H.; Yu, Y.; Zhang, D.; Zhao, X.; et al. A genomic variation map provides insights into the genetic basis of spring Chinese cabbage (Brassica rapa ssp. pekinensis) selection. Mol. Plant 2018, 11, 1360–1376. [Google Scholar] [CrossRef] [Green Version]
- Geraldo, N.; Baurle, I.; Kidou, S.; Hu, X.; Dean, C. FRIGIDA delays flowering in Arabidopsis via a cotranscriptional mechanism involving direct interaction with the nuclear Cap-Binding Complex. Plant Physiol. 2009, 150, 1611–1618. [Google Scholar] [CrossRef] [Green Version]
- Searle, I.; He, Y.; Turck, F.; Vincent, C.; Fornara, F.; Krober, S.; Amasino, R.A.; Coupland, G. The transcription factor FLC confers a flowering response to vernalization by repressing meristem competence and systemic signaling in Arabidopsis. Genes Dev. 2006, 20, 898–912. [Google Scholar] [CrossRef] [Green Version]
- Michaels, S.D.; Amasino, R.M. FLOWERING LOCUS C encodes a novel MADS domain protein that acts as a repressor of flowering. Plant Cell 1999, 11, 949–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blümel, M.; Dally, N.; Jung, C. Flowering time regulation in crops-what did we learn from Arabidopsis? Curr. Opin. Biotechnol. 2015, 32, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, C.C.; Burn, J.E.; Perez, P.P.; Metzger, J.; Edwards, J.A.; Peacock, W.J.; Dennis, E.S. The FLF MADS box gene: A repressor of flowering in Arabidopsis regulated by vernalization and methylation. Plant Cell. 1999, 11, 445–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.; Amasino, R.M. Effect of vernalization, photoperiod, and light quality on the flowering phenotype of Arabidopsis plants containing the FRIGIDA gene. Plant Physiol. 1995, 108, 157–162. [Google Scholar] [CrossRef] [Green Version]
- Greb, T.; Mylne, J.S.; Crevillen, P.; Geraldo, N.; An, H.; Gendall, A.R.; Dean, C. The PHD finger protein VRN5 functions in the epigenetic silencing of Arabidopsis FLC. Curr. Biol. 2007, 17, 73–78. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Amasino, R.M. Role of chromatin modification in flowering time control. Trends Plant Sci. 2005, 10, 30–35. [Google Scholar] [CrossRef]
- Wood, C.C.; Robertson, M.; Tanner, G.; Peacock, W.J.; Dennis, E.S.; Helliwell, C.A. The Arabidopsis thaliana vernalization response requires a polycomb-like protein complex that also includes VERNALIZATION INSENSITIVE 3. Proc. Natl. Acad. Sci. USA 2006, 103, 14631–14636. [Google Scholar] [CrossRef] [Green Version]
- Sung, S.; Amasino, R.M. Vernalization and epigenetics: How plants remember winter. Curr. Opin. Plant Biol. 2004, 7, 4–10. [Google Scholar] [CrossRef]
- Gendall, A.R.; Levy, Y.Y.; Wilson, A.; Dean, C. The VERNALIZATION 2 gene mediates the epigenetic regulation of vernalization in Arabidopsis. Cell 2001, 107, 525–535. [Google Scholar] [CrossRef] [Green Version]
- Levy, Y.Y.; Mesnage, S.; Mylne, J.S.; Gendall, A.R.; Dean, C. Multiple roles of Arabidopsis VRN1 in vernalization and flowering time control. Science 2002, 297, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, C.C.; Finnegan, E.J.; Dennis, E.S.; Peacock, W.J. Quantitative effects of vernalization on FLC and SOC1 expression. Plant J. 2006, 45, 871–883. [Google Scholar] [CrossRef] [PubMed]
- Akter, A.; Miyazaki, J.; Shea, D.J.; Nishida, N.; Takada, S.; Miyaji, N.; Mehraj, H.; Shimizu, M.; Doullah, M.A.; Takasaki-Yasuda, T.; et al. Gene expression analysis in response to vernalization in Chinese cabbage (Brassica rapa L.). Hortic. J. 2020, 89, 268–277. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Duan, W.; Huang, Z.; Liu, G.; Wu, P.; Liu, T.; Li, Y.; Hou, X. Comprehensive analysis of the flowering genes in Chinese cabbage and examination of evolutionary pattern of co-like genes in plant kingdom. Sci. Rep. 2015, 5, 14631. [Google Scholar] [CrossRef]
- Shea, D.J.; Nishida, N.; Takada, S.; Itabashi, E.; Takahashi, S.; Akter, A.; Miyaji, N.; Osabe, K.; Mehraj, H.; Shimizu, M.; et al. Long noncoding RNAs in Brassica rapa L. Following vernalization. Sci. Rep. 2019, 9, 9302. [Google Scholar] [CrossRef]
- Jeong, S.; Ahn, H.; Ryu, J.; Oh, Y.; Sivanandhan, G.; Won, K.-H.; Park, Y.; Kim, J.-S.; Lim, Y.P.; Kim, S.-G. Generation of early-flowering Chinese cabbage (Brassica rapa spp. Pekinensis) through CRISPR/Cas9-mediated genome editing. Plant Biotechnol. Rep. 2019, 13, 491–499. [Google Scholar] [CrossRef]
- Murovec, J.; Gucek, K.; Bohanec, B.; Avbelj, M.; Jerala, R. DNA-free genome editing of Brassica oleracea and B. rapa protoplasts using CRISPR-Cas9 ribonucleoprotein complexes. Front. Plant Sci. 2018, 9, 1594. [Google Scholar] [CrossRef]
- Park, J.; Bae, S.; Kim, J.-S. Cas-Designer: A web-based tool for choice of CRISPR-Cas9 target sites. Bioinformatics 2015, 31, 4014–4016. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Mao, Y.; Ha, S.; Liu, W.; Botella, J.R.; Zhu, J.-K. A multiplex CRISPR/Cas9 platform for fast and efficient editing of multiple genes in Arabidopsis. Plant Cell Rep. 2016, 35, 1519–1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.Y.; Park, B.S.; Kwon, S.J.; Kim, J.; Lim, M.H.; Park, Y.D.; Kim, D.Y.; Suh, S.C.; Jin, Y.M.; Ahn, J.H.; et al. Delayed flowering time in Arabidopsis and Brassica rapa by the overexpression of FLOWERING LOCUS C (FLC) homologs isolated from Chinese cabbage (Brassica rapa L. ssp. pekinensis). Plant Cell Rep. 2007, 26, 327–336. [Google Scholar] [CrossRef]
- Lee, M.-K.; Kim, H.-S.; Kim, J.-S.; Kim, S.-H.; Park, Y.-D. Agrobacterium-mediated transformation system for large-scale production of transgenic Chinese cabbage (Brassica rapa L. ssp. pekinensis) plants for insertional mutagenesis. J. Plant Biol. 2004, 47, 300–306. [Google Scholar] [CrossRef]
- Jung, H.; Lee, A.; Jo, S.H.; Park, H.J.; Jung, W.Y.; Kim, H.-S.; Lee, H.-J.; Jeong, S.-G.; Kim, Y.-S.; Cho, H.S. Nitrogen signaling genes and SOC1 determine the flowering time in a reciprocal negative feedback loop in Chinese cabbage (Brassica rapa L.) based on CRISPR/Cas9-mediated mutagenesis of multiple BrSOC1 homologs. Int. J. Mol. Sci. 2021, 22, 4631. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Cox, M.P.; Peterson, D.A.; Biggs, P.J. SolexaQA: At-a-glance quality assessment of Illumina second-generation sequencing data. BMC Bioinform. 2010, 11, 485. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.E.; Oh, S.K.; Lee, J.H.; Lee, B.M.; Jo, S.H. Genome-wide SNP calling using next generation sequencing data in tomato. Mol. Cells 2014, 37, 36–42. [Google Scholar] [CrossRef]
- Bae, S.; Park, J.; Kim, J.S. Cas-OFFinder: A fast and versatile algorithm that searches for potential off-target sites of Cas9 RNA-guided endonucleases. Bioinformatics 2014, 30, 1473–1475. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Zhang, S.; Sun, X.; Li, G.; Yuan, L.; Li, F.; Zhang, H.; Zhang, S.; Chen, G.; Wang, C.; et al. Comparative transcriptome analysis of gene expression and regulatory characteristics associated with different vernalization periods in Brassica rapa. Genes 2020, 11, 392. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Yi, H.; Do, C.; Han, C.-T.; Nou, I.-S.; Hur, Y. Genome-wide analysis of gene expression to distinguish photoperiod-dependent and -independent flowering in Brassicaceae. Genes Genom. 2017, 39, 207–223. [Google Scholar] [CrossRef]
- Lin, S.I.; Wang, J.G.; Poon, S.Y.; Su, C.L.; Wang, S.S.; Chiou, T.J. Differential regulation of FLOWERING LOCUS C expression by vernalization in cabbage and Arabidopsis. Plant Physiol. 2005, 137, 1037–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, G.J.; Chanson, A.H.; McCallum, E.J.; Ohme-Takagi, M.; Byriel, K.; Hill, J.M.; Martin, J.L.; Mylne, J.S. The Arabidopsis B3 domain protein VERNALIZATION1 (VRN1) is involved in processes essential for development, with structural and mutational studies revealing its DNA-binding surface. J. Biol. Chem. 2013, 288, 3198–3207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Chen, L.; Liu, X.; Guo, C.; Sun, S.; Wu, C.; Jiang, B.; Han, T.; Hou, W. CRISPR/Cas9-mediated targeted mutagenesis of GmFT2a delays flowering time in soya bean. Plant Biotechnol. J. 2018, 16, 176–185. [Google Scholar] [CrossRef] [Green Version]
- Jing, C.; Wei, M.; Fang, P.; Song, R.; Qi, W. Pollen-specific CRISPR/Cas9 system to increase heritable gene mutations in maize. Agriculture 2021, 11, 751. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, J.; Wei, P.; Zhang, B.; Gou, F.; Feng, Z.; Mao, Y.; Yang, L.; Zhang, H.; Xu, N.; et al. The CRISPR/Cas9 system produces specific and homozygous targeted gene editing in rice in one generation. Plant Biotechnol. J. 2014, 12, 797–807. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, Q.; Zhu, Q.; Liu, W.; Chen, Y.; Qiu, R.; Wang, B.; Yang, Z.; Li, H.; Lin, Y.; et al. A robust CRISPR/Cas9 system for convenient, high-efficiency multiplex genome editing in monocot and dicot plants. Mol. Plant 2015, 8, 1274–1284. [Google Scholar] [CrossRef]
- Gao, X.; Chen, J.; Dai, X.; Zhang, D.; Zhao, Y. An effective strategy for reliably isolating heritable and Cas9-free Arabidopsis mutants generated by CRISPR/Cas9-mediated genome editing. Plant Physiol. 2016, 171, 1794–1800. [Google Scholar] [CrossRef] [Green Version]
- Pyott, D.E.; Sheehan, E.; Molnar, A. Engineering of CRISPR/Cas9-mediated potyvirus resistance in transgene-free Arabidopsis plants. Mol. Plant Pathol. 2016, 17, 1276–1288. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Li, X.; Zhou, Z.; Wu, P.; Fang, M.; Pan, X.; Lin, Q.; Luo, W.; Wu, G.; Li, H. Reassessment of the Four Yield-related Genes Gn1a, DEP1, GS3, and IPA1 in Rice Using a CRISPR/Cas9 System. Front. Plant Sci. 2016, 7, 377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; He, M.; Li, J.; Chen, L.; Huang, Z.; Zheng, S.; Zhu, L.; Ni, E.; Jiang, D.; Zhao, B.; et al. Development of commercial thermo-sensitive genic male sterile rice accelerates hybrid rice breeding using the CRISPR/Cas9-mediated TMS5 editing system. Sci. Rep. 2016, 6, 37395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Liang, Z.; Zong, Y.; Wang, Y.; Liu, J.; Chen, K.; Qiu, J.-L.; Gao, C. Efficient and transgene-free genome editing in wheat through transient expression of CRISPR/Cas9 DNA or RNA. Nat. Commun. 2016, 7, 12617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soyk, S.; Muller, N.A.; Park, S.J.; Schmalenbach, I.; Jiang, K.; Hayama, R.; Zhang, L.; Van Eck, J.; Jimenez-Gomez, J.M.; Lippman, Z.B. Variation in the flowering gene SELF PRUNING 5G promotes day-neutrality and early yield in tomato. Nat. Genet. 2017, 49, 162–168. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vernalization Period | Target Gene | GE1 Line | Days to Visible Flowering Bud in Plants |
|---|---|---|---|
| 12 June–12 October 2020 (60 days) | No gene targeted | Wt-1~10 | 9 (9 ± 0) a |
| BrVRN1 (multi-target) | #1-1~19 | 19-20 (19.4 ± 0.116 **) | |
| BrVRN1 (multi-target) | #2-1~13 | 19-20 (19.2 ± 0.104 **) | |
| 28 September–10 November 2020 (40 days) | No gene targeted | Wt-1~5 | 15-16 (15.6 ± 0.245) |
| BrVRN1 (multi-target) | #7-1~5 | 23-27 (25.4 ± 0.748 **) |
| Vernalization Period | Target Gene | GE2 Line | Days to Visible Flowering Bud in Plants |
|---|---|---|---|
| 21 January–2 March 2021 (40 days) | No gene targeted | Wt-1~10 | 16 (16 ± 0) a |
| BrVRN1 (multi-target, transgene-free) | #2-2-1~10 | 37–59 (52.7 ± 2.395 **) | |
| BrVRN1 (multi-target, transgene-free) | #2-8-1~10 | 38–56 (48.3 ± 2.408 **) |
| Target | Target Sequence | Mismatch | Number of Found Targets | Number of SNP Positions | Number of In/Del Positions |
|---|---|---|---|---|---|
| gRNA1 | ACACGCCAAACGTGACCATCNGG | 1 | 0 | 0 | 0 |
| 2 | 0 | 0 | 0 | ||
| 3 | 1 | 0 | 0 | ||
| gRNA2 | GACCGTTACTCAATCCGCATNGG | 1 | 0 | 0 | 0 |
| 2 | 0 | 0 | 0 | ||
| 3 | 0 | 0 | 0 | ||
| gRNA3 | AGAAGAAGGATAAACAACCTNGG | 1 | 0 | 0 | 0 |
| 2 | 0 | 0 | 0 | ||
| 3 | 6 | 1 | 0 | ||
| gRNA4 | ATAGCGGCCACAGTAACTTCNGG | 1 | 0 | 0 | 0 |
| 2 | 0 | 0 | 0 | ||
| 3 | 0 | 0 | 0 | ||
| total | 7 | 1 | 0 |
| Line | No. of Total Trimmed Reads *1 | No. of Mapped Reads *2 | Mapped Region *3 (%) |
|---|---|---|---|
| WT | 143,832,424 | 24 (0.00%) | 374 (2.81%) |
| #2-2-1 | 110,834,798 | 1 (0.00%) | 0 (0.00%) |
| #2-2-2 | 144,407,538 | 13 (0.00%) | 0 (0.00%) |
| #2-2-3 | 129,575,654 | 6 (0.00%) | 0 (0.00%) |
| #2-2-4 | 160,433,726 | 8 (0.00%) | 0 (0.00%) |
| #2-2-5 | 120,246,026 | 4 (0.00%) | 0 (0.00%) |
| #2-8-1 | 130,463,146 | 4 (0.00%) | 0 (0.00%) |
| #2-8-2 | 153,956,546 | 7 (0.00%) | 0 (0.00%) |
| #2-8-3 | 151,410,950 | 2 (0.00%) | 0 (0.00%) |
| #2-8-4 | 160,952,626 | 6 (0.00%) | 0 (0.00%) |
| #2-8-5 | 126,426,790 | 12 (0.00%) | 0 (0.00%) |
| Line | Target | Type of Mutation | BrVRN1a | BrVRN1b | BrVRN1c | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Reads | Read Rate (%) | Zygosity | Reads | Read Rate (%) | Zygosity | Reads | Read Rate (%) | Zygosity | |||
| #2-2-1~5 | gRNA1 | Wt | 10 | Homozygous | 115 | Wt | 171 | 0 | Wt | ||
| +A | 103 | 91.15 | - | 0 | - | ||||||
| Total | 113 | 115 | 171 | ||||||||
| gRNA2 | Wt | 11 | Homozygous | 4 | Homozygous | 165 | 0 | Wt | |||
| +C/+A | 110 | 90.90 | 93 | 95.87 | - | ||||||
| Total | 121 | - | 97 | 165 | |||||||
| gRNA3 | Wt | 147 | Wt | - | - | - | - | - | |||
| SNP/indel | - | 0 | - | - | - | ||||||
| Total | 147 | - | - | ||||||||
| gRNA4 | Wt | 124 | Wt | - | - | - | - | ||||
| SNP/indel | - | 0 | - | - | - | - | |||||
| Total | 124 | - | - | ||||||||
| #2-8-1~5 | gRNA1 | Wt | 5 | Homozygous | 108 | Wt | 171 | 0 | Wt | ||
| +A | 115 | 95.83 | - | 0 | - | ||||||
| Total | 120 | 108 | 171 | ||||||||
| gRNA2 | Wt | 11 | Homozygous | 8 | Homozygous | 155 | 0 | Wt | |||
| +C/+A | 135 | 92.46 | 121 | 93.80 | - | ||||||
| Total | 146 | 129 | 155 | ||||||||
| gRNA3 | Wt | 151 | Wt | - | - | - | - | - | |||
| SNP/indel | - | 0 | - | - | - | ||||||
| Total | 151 | - | - | ||||||||
| gRNA4 | Wt | 149 | Wt | - | - | - | - | - | |||
| SNP/indel | - | 0 | - | - | - | ||||||
| Total | 149 | - | - | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, J.K.; Suh, E.J.; Park, S.R.; Park, J.; Lee, Y.-H. Multiplex CRISPR/Cas9 Mutagenesis of BrVRN1 Delays Flowering Time in Chinese Cabbage (Brassica rapa L. ssp. pekinensis). Agriculture 2021, 11, 1286. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11121286
Hong JK, Suh EJ, Park SR, Park J, Lee Y-H. Multiplex CRISPR/Cas9 Mutagenesis of BrVRN1 Delays Flowering Time in Chinese Cabbage (Brassica rapa L. ssp. pekinensis). Agriculture. 2021; 11(12):1286. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11121286
Chicago/Turabian StyleHong, Joon Ki, Eun Jung Suh, Sang Ryeol Park, Jihee Park, and Yeon-Hee Lee. 2021. "Multiplex CRISPR/Cas9 Mutagenesis of BrVRN1 Delays Flowering Time in Chinese Cabbage (Brassica rapa L. ssp. pekinensis)" Agriculture 11, no. 12: 1286. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11121286