
Biostimulant Effects of an Aqueous Extract of Duckweed (Lemna minor L.) on Physiological and Biochemical Traits in the Olive Tree

 , , ,
, , ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Olive Material and Growing Conditions
2.2. Lemna Minor Growth Conditions and Preparation of the Extract
2.3. Olive Treatments with Duckweed Extract
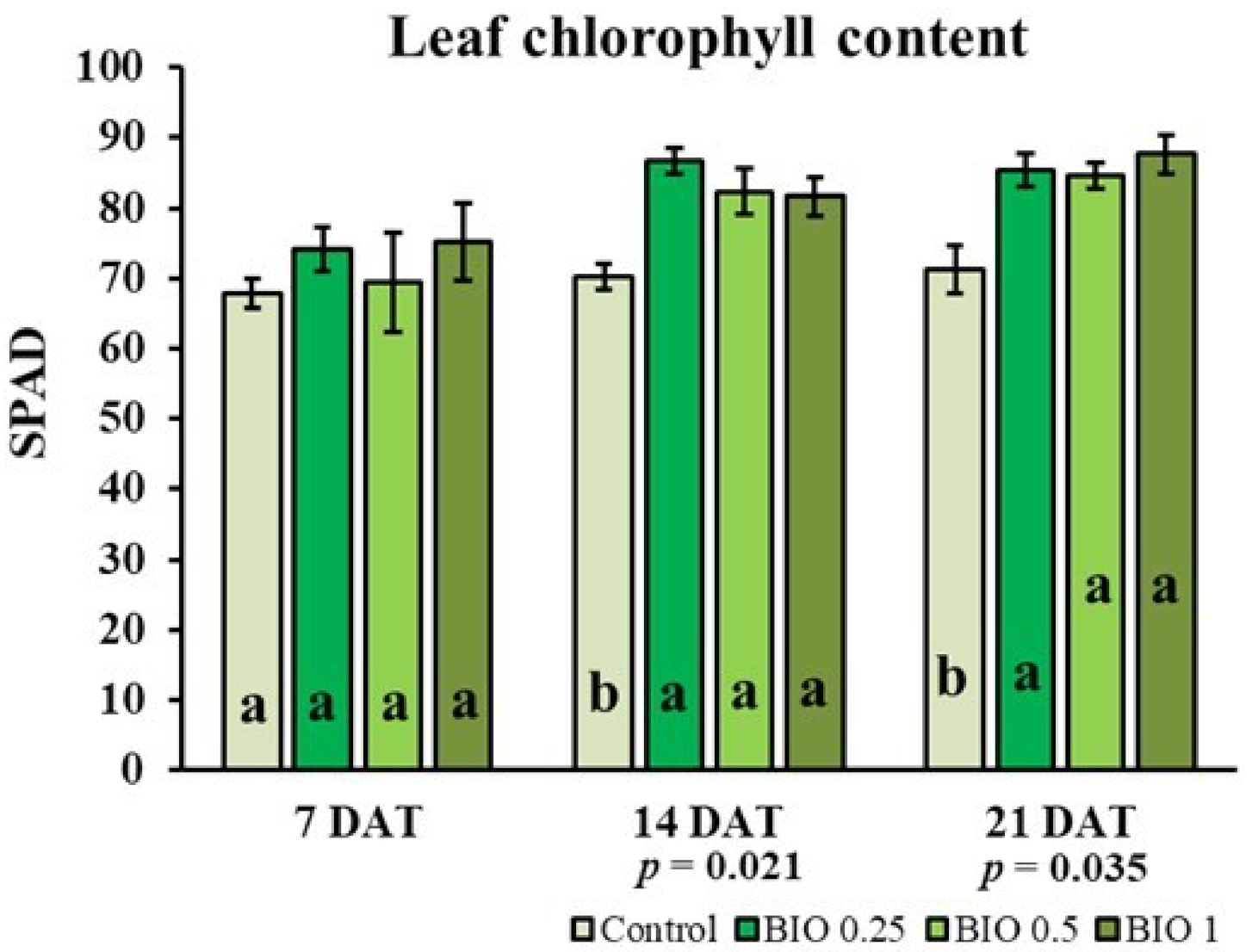
2.4. Olive Leaf Gas Exchange, Chlorophyll Content and Growth
2.5. Nutrient Determination in Olive Leaves
2.6. Duckweed Extract Profiling
2.7. Statistical Analysis
3. Results
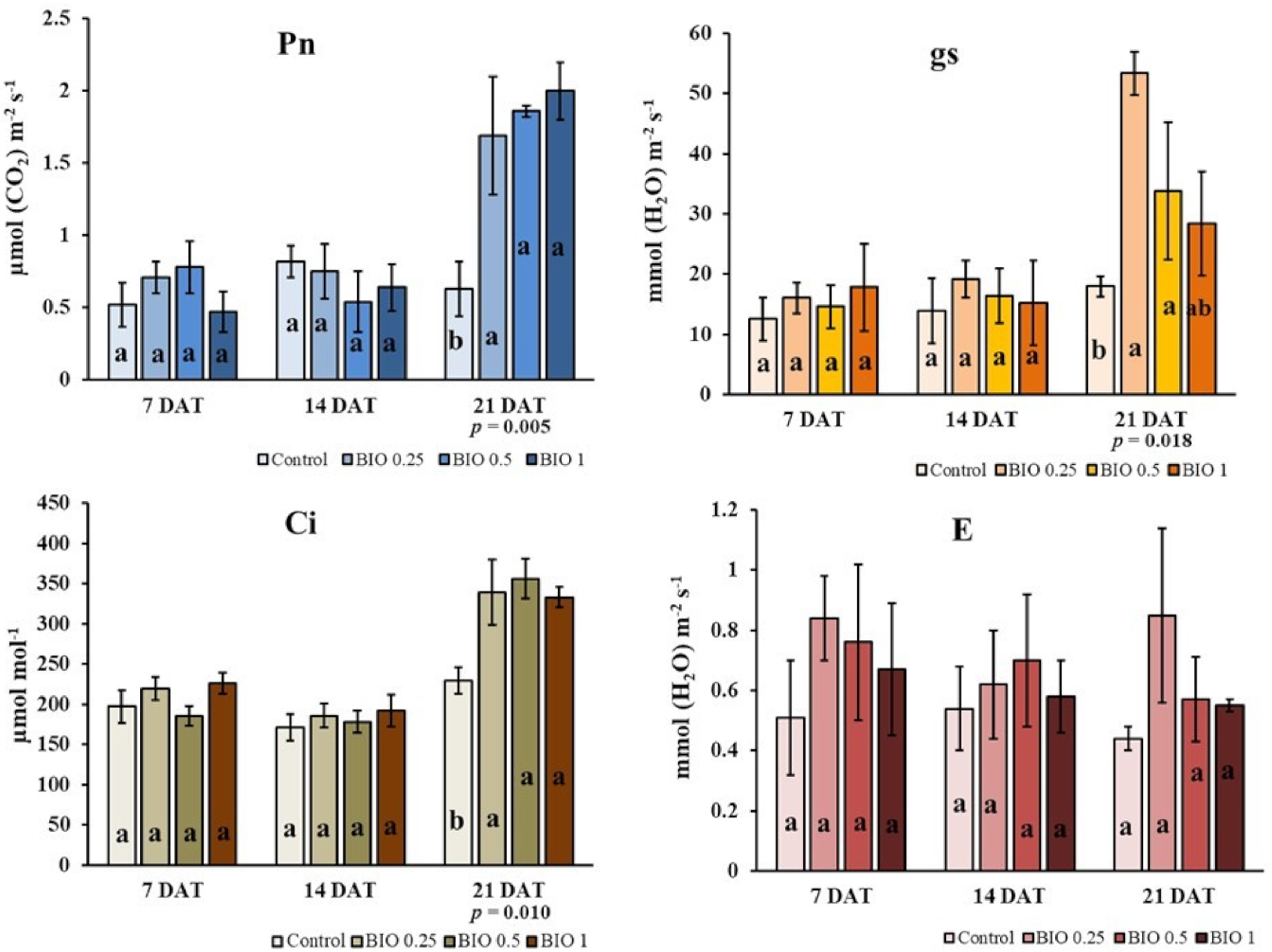
3.1. Leaf Net Photosynthesis (Pn), Leaf Transpiration Rate (E), Stomatal Conductance (gs) and Sub-Stomatal CO2 Concentration (Ci)
3.2. Plant Growth and Biomass Development
3.3. Effect of the Duckweed Extract, Applied at the Three Different Concentrations, on Olive Nutrient Content
3.4. Duckweed Extract Phytochemical Profile
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Du Jardin, P. Plant Biostimulants: Definition, Concept, Main Categories and Regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein Hydrolysates as Biostimulants in Horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Del Buono, D. Can Biostimulants Be Used to Mitigate the Effect of Anthropogenic Climate Change on Agriculture? It Is Time to Respond. Sci. Total Environ. 2021, 751, 141763. [Google Scholar] [CrossRef] [PubMed]
- Agliassa, C.; Mannino, G.; Molino, D.; Cavalletto, S.; Contartese, V.; Bertea, C.M.; Secchi, F. A New Protein Hydrolysate-Based Biostimulant Applied by Fertigation Promotes Relief from Drought Stress in Capsicum Annuum L. Plant Physiol. Biochem. 2021, 166, 1076–1086. [Google Scholar] [CrossRef]
- Malik, A.; Mor, V.S.; Tokas, J.; Punia, H.; Malik, S.; Malik, K.; Sangwan, S.; Tomar, S.; Singh, P.; Singh, N.; et al. Biostimulant-Treated Seedlings under Sustainable Agriculture: A Global Perspective Facing Climate Change. Agronomy 2021, 11, 14. [Google Scholar] [CrossRef]
- Kocira, S. Effect of Amino Acid Biostimulant on the Yield and Nutraceutical Potential of Soybean. Chil. J. Agric. Res. 2019, 79, 17–25. [Google Scholar] [CrossRef]
- Toscano, S.; Ferrante, A.; Branca, F.; Romano, D. Enhancing the Quality of Two Species of Baby Leaves Sprayed with Moringa Leaf Extract as Biostimulant. Agronomy 2021, 11, 1399. [Google Scholar] [CrossRef]
- Mannino, G.; Campobenedetto, C.; Vigliante, I.; Contartese, V.; Gentile, C.; Bertea, C.M. The Application of a Plant Biostimulant Based on Seaweed and Yeast Extract Improved Tomato Fruit Development and Quality. Biomolecules 2020, 10, 1662. [Google Scholar] [CrossRef] [PubMed]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural Uses of Plant Biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [Green Version]
- Caradonia, F. Plant Biostimulant Regulatory Framework: Prospects in Europe and Current Situation at International Level. J. Plant Growth Regul. 2019, 38, 438–448. [Google Scholar] [CrossRef]
- Nardi, S.; Pizzeghello, D.; Reniero, F.; Rascio, N. Chemical and Biochemical Properties of Humic Substances Isolated from Forest Soils and Plant Growth. Soil Sci. Soc. Am. J. 2000, 64, 639–645. [Google Scholar] [CrossRef]
- Buono, D.D.; Regni, L.; Pino, A.M.D.; Bartucca, M.L.; Palmerini, C.A.; Proietti, P. Effects of Megafol on the Olive Cultivar ‘Arbequina’ Grown Under Severe Saline Stress in Terms of Physiological Traits, Oxid. Front. Plant Sci. 2021, 11, 1–12. [Google Scholar]
- Ali, A.H.; Aboohanah, M.A.; Abdulhussein, M.A. Impact of Foliar Application with Dry Yeast Suspension and Amino Acid on Vegetative Growth, Yield and Quality Characteristics of Olive (Olea Europaea L.) Trees. Kufa J. Agric. Sci. 2019, 11, 10. [Google Scholar]
- Chouliaras, V.; Tasioula, M.; Chatzissavvidis, C.; Therios, I.; Tsabolatidou, E. The Effects of a Seaweed Extract in Addition to Nitrogen and Boron Fertilization on Productivity, Fruit Maturation, Leaf Nutritional Status and Oil Quality of the Olive (Olea Europaea L.) Cultivar Koroneiki. J. Sci. Food Agric. 2009, 89, 984–988. [Google Scholar] [CrossRef]
- Soria, C.M. Olive Tree (Olea europaea L.) Response to the Application of Biostimulants; CIHEAM-IAMZ: Zaragoza, Spain, 2006. [Google Scholar]
- Saour, G. Morphological Assessment of Olive Seedlings Treated with Kaolin-Based Particle Film and Biostimulant. Adv. Hortic. Sci. 2005, 19, 193–197. [Google Scholar]
- Almadi, L.; Paoletti, A.; Cinosi, N.; Daher, E.; Rosati, A.; Di Vaio, C.; Famiani, F. A Biostimulant Based on Protein Hydrolysates Promotes the Growth of Young Olive Trees. Agric. Switz. 2020, 10, 618. [Google Scholar] [CrossRef]
- El-Mageed, T.A.A.; Semida, W.M.; Rady, M.M. Moringa Leaf Extract as Biostimulant Improves Water Use Efficiency, Physio-Biochemical Attributes of Squash Plants under Deficit Irrigation. Agric. Water Manag. 2017, 193, 46–54. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Casadesús, A.; Brockman, H.; Munné-Bosch, S. An Overview of Plant-Based Natural Biostimulants for Sustainable Horticulture with a Particular Focus on Moringa Leaf Extracts. Plant Sci. 2020, 295, 110194. [Google Scholar] [CrossRef]
- Caruso, G.; De Pascale, S.; Cozzolino, E.; Giordano, M.; El-Nakhel, C.; Cuciniello, A.; Cenvinzo, V.; Colla, G.; Rouphael, Y. Protein Hydrolysate or Plant Extract-Based Biostimulants Enhanced Yield and Quality Performances of Greenhouse Perennial Wall Rocket Grown in Different Seasons. Plants 2019, 8, 208. [Google Scholar] [CrossRef] [Green Version]
- Saavedra, T.; Gama, F.; Correia, P.J.; Da Silva, J.P.; Miguel, M.G.; de Varennes, A.; Pestana, M. A Novel Plant Extract as a Biostimulant to Recover Strawberry Plants from Iron Chlorosis. J. Plant Nutr. 2020, 43, 2054–2066. [Google Scholar] [CrossRef]
- Gebreluel, T.; He, M.; Zheng, S.; Zou, S.; Woldemicael, A.; Wang, C. Optimization of Enzymatic Degradation of Dealginated Kelp Waste through Response Surface Methodology. J. Appl. Phycol. 2020, 32, 529–537. [Google Scholar] [CrossRef]
- Nanda, S.; Kumar, G.; Hussain, S. Utilization of Seaweed-Based Biostimulants in Improving Plant and Soil Health: Current Updates and Future Prospective. Int. J. Environ. Sci. Technol. 2021, 1–14. [Google Scholar] [CrossRef]
- Mzibra, A.; Aasfar, A.; Khouloud, M.; Farrie, Y.; Boulif, R.; Kadmiri, I.M.; Bamouh, A.; Douira, A. Improving Growth, Yield, and Quality of Tomato Plants (Solanum Lycopersicum L) by the Application of Moroccan Seaweed-Based Biostimulants under Greenhouse Conditions. Agronomy 2021, 11, 1373. [Google Scholar] [CrossRef]
- Del Buono, D.; Bartucca, M.L.; Ballerini, E.; Senizza, B.; Lucini, L.; Trevisan, M. Physiological and Biochemical Effects of an Aqueous Extract of Lemna Minor L. as a Potential Biostimulant for Maize. J. Plant Growth Regul. 2021. [Google Scholar] [CrossRef]
- Lasfar, S.; Monette, F.; Millette, L.; Azzouz, A. Intrinsic Growth Rate: A New Approach to Evaluate the Effects of Temperature, Photoperiod and Phosphorus-Nitrogen Concentrations on Duckweed Growth under Controlled Eutrophication. Water Res. 2007, 41, 2333–2340. [Google Scholar] [CrossRef] [PubMed]
- Panfili, I.; Bartucca, M.L.; Del Buono, D. The Treatment of Duckweed with a Plant Biostimulant or a Safener Improves the Plant Capacity to Clean Water Polluted by Terbuthylazine. Sci. Total Environ. 2019, 646, 832–840. [Google Scholar] [CrossRef] [PubMed]
- Gülçin, I.; Kireçci, E.; Akkemik, E.; Fevzi, T.; Hisar, O. Antioxidant, Antibacterial, and Anticandidal Activities of an Aquatic Plant: Duckweed (Lemna Minor L. Lemnaceae). Turk. J. Biol. 2010, 34, 175–188. [Google Scholar] [CrossRef]
- Del Buono, D.; Di Michele, A.; Costantino, F.; Trevisan, M.; Lucini, L. Biogenic ZnO Nanoparticles Synthesized Using a Novel Plant Extract: Application to Enhance Physiological and Biochemical Traits in Maize. Nanomaterials 2021, 11, 1270. [Google Scholar] [CrossRef]
- Gülçin, I. The Antioxidant and Radical Scavenging Activities of Black Pepper (Piper Nigrum) Seeds. Int. J. Food Sci. Nutr. 2005, 56, 491–499. [Google Scholar] [CrossRef]
- Caruso, T.; Campisi, G.; Marra, F.P.; Camposeo, S.; Vivaldi, G.A.; Proietti, P.; Nasini, L. Growth and Yields of “Arbequina” High-Density Planting Systems in Three Different Olive Growing Areas in Italy. Acta Horticulturae 2014, 1057, 341–348. [Google Scholar] [CrossRef] [Green Version]
- Rosati, A.; Paoletti, A.; Al Harir, R.; Famiani, F. Fruit Production and Branching Density Affect Shoot and Whole-Tree Wood to Leaf Biomass Ratio in Olive. Tree Physiol. 2018, 38, 1278–1285. [Google Scholar] [CrossRef] [PubMed]
- Rosati, A.; Paoletti, A.; Al Hariri, R.; Morelli, A.; Famiani, F. Resource Investments in Reproductive Growth Proportionately Limit Investments in Whole-Tree Vegetative Growth in Young Olive Trees with Varying Crop Loads. Tree Physiol. 2018, 38, 1267–1277. [Google Scholar] [CrossRef] [Green Version]
- Famiani, F.; Farinelli, D.; Gardi, T.; Rosati, A. The Cost of Flowering in Olive (Olea Europaea L.). Sci. Hortic. 2019, 252, 268–273. [Google Scholar] [CrossRef]
- Aquaponics Food Production Systems: Combined Aquaculture and Hydroponic Production Technologies for the Future; Life Sciences; Göddeke, S.; Joyce, A.; Kotzen, B.; Burnell, G.M. (Eds.) Springer: Cham, Switzerland, 2019; ISBN 978-3-030-15943-. [Google Scholar]
- Tabatabai, M.A.; Bremner, J.M. A Simple Turbidimetric Method of Determining Total Sulfur in Plant Materials1. Agron. J. 1970, 62, 805–806. [Google Scholar] [CrossRef]
- Hansen, T.H.; De Bang, T.C.; Laursen, K.H.; Pedas, P.; Husted, S.; Schjoerring, J.K. Multielement Plant Tissue Analysis Using ICP Spectrometry. Methods Mol. Biol. 2013, 953, 121–141. [Google Scholar]
- Lenth, R.V. Least-Squares Means: The R Package Lsmeans. J. Stat. Softw. 2016, 69, 17496. [Google Scholar] [CrossRef] [Green Version]
- Povero, G.; Mejia, J.F.; Di Tommaso, D.; Piaggesi, A.; Warrior, P. A Systematic Approach to Discover and Characterize Natural Plant Biostimulants. Front. Plant Sci. 2016, 7, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulgari, R.; Podetta, N.; Cocetta, G.; Piaggesi, A.; Ferrante, A. The Effect of a Complete Fertilizer for Leafy Vegetables Production in Family and Urban Gardens. Bulg. J. Agric. Sci. 2014, 20, 1361–1367. [Google Scholar]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [Green Version]
- Puglia, D.; Pezzolla, D.; Gigliotti, G.; Torre, L.; Bartucca, M.L.; Del Buono, D. The Opportunity of Valorizing Agricultural Waste, Through Its Conversion into Biostimulants, Biofertilizers, and Biopolymers. Sustainability 2021, 13, 2710. [Google Scholar] [CrossRef]
- Bulgari, R.; Cocetta, G.; Trivellini, A.; Vernieri, P.; Ferrante, A. Biostimulants and Crop Responses: A Review. Biol. Agric. Hortic. 2015, 31, 1–17. [Google Scholar] [CrossRef]
- Abdalla, M.M. The Potential of Moringa Oleifera Extract as a Biostimulant in Enhancing the Growth, Biochemical and Hormonal Contents in Rocket (Eruca Vesicaria Subsp. Sativa) Plants. Int. J. Plant Physiol. Biochem. 2013, 5, 42–49. [Google Scholar] [CrossRef]
- Bulgari, R.; Morgutti, S.; Cocetta, G.; Negrini, N.; Farris, S.; Calcante, A.; Spinardi, A.; Ferrari, E.; Mignani, I.; Oberti, R.; et al. Evaluation of Borage Extracts as Potential Biostimulant Using a Phenomic, Agronomic, Physiological, and Biochemical Approach. Front. Plant Sci. 2017, 8, 935. [Google Scholar] [CrossRef] [PubMed]
- Proietti, P.; Federici, E.; Fidati, L.; Scargetta, S.; Massaccesi, L.; Nasini, L.; Regni, L.; Ricci, A.; Cenci, G.; Gigliotti, G. Effects of Amendment with Oil Mill Waste and Its Derived-Compost on Soil Chemical and Microbiological Characteristics and Olive (Olea Europaea L.) Productivity. Agric. Ecosyst. Environ. 2015, 207, 51–60. [Google Scholar] [CrossRef]
- Kałuzewicz, A.; Krzesiński, W.; Spizewski, T.; Zaworska, A. Effect of Biostimulants on Several Physiological Characteristics and Chlorophyll Content in Broccoli under Drought Stress and Re-Watering. Not. Bot. Horti Agrobot. Cluj-Napoca 2017, 45, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Popescu, G.C.; Popescu, M. Yield, Berry Quality and Physiological Response of Grapevine to Foliar Humic Acid Application. Bragantia 2018, 77, 273–282. [Google Scholar] [CrossRef]
- Carillo, P.; Colla, G.; El-Nakhel, C.; Bonini, P.; D’Amelia, L.; Dell’Aversana, E.; Pannico, A.; Giordano, M.; Sifola, M.I.; Kyriacou, M.C.; et al. Biostimulant Application with a Tropical Plant Extract Enhances Corchorus Olitorius Adaptation to Sub-Optimal Nutrient Regimens by Improving Physiological Parameters. Agronomy 2019, 9, 249. [Google Scholar] [CrossRef] [Green Version]
- Spinelli, F.; Fiori, G.; Noferini, M.; Sprocatti, M.; Costa, G. A Novel Type of Seaweed Extract as a Natural Alternative to the Use of Iron Chelates in Strawberry Production. Sci. Hortic. 2010, 125, 263–269. [Google Scholar] [CrossRef]
- Massa, D.; Prisa, D.; Montoneri, E.; Battaglini, D.; Ginepro, M.; Negre, M.; Burchi, G. Application of Municipal Biowaste Derived Products in Hibiscus Cultivation: Effect on Leaf Gaseous Exchange Activity, and Plant Biomass Accumulation and Quality. Sci. Hortic. 2016, 205, 59–69. [Google Scholar] [CrossRef]
- Fascella, G.; Montoneri, E.; Ginepro, M.; Francavilla, M. Effect of Urban Biowaste Derived Soluble Substances on Growth, Photosynthesis and Ornamental Value of Euphorbia x Lomi. Sci. Hortic. 2015, 197, 90–98. [Google Scholar] [CrossRef]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed Extracts as Biostimulants of Plant Growth and Development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Fan, D.; Hodges, D.M.; Critchley, A.T.; Prithiviraj, B. A Commercial Extract of Brown Macroalga (Ascophyllum Nodosum) Affects Yield and the Nutritional Quality of Spinach In Vitro. Commun. Soil Sci. Plant Anal. 2013, 44, 1873–1884. [Google Scholar] [CrossRef]
- Dudaš, S.; Šola, I.; Sladonja, B.; Erhatić, R.; Ban, D.; Poljuha, D. The Effect of Biostimulant and Fertilizer on “Low Input” Lettuce Production. Acta Bot. Croat. 2016, 75, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Regni, L.; Nasini, L.; Ilarioni, L.; Brunori, A.; Massaccesi, L.; Agnelli, A.; Proietti, P. Long Term Amendment with Fresh and Composted Solid Olive Mill Waste on Olive Grove Affects Carbon Sequestration by Prunings, Fruits, and Soil. Front. Plant Sci. 2017, 7, 2042. [Google Scholar] [CrossRef] [PubMed]
- du Jardin, P.; Xu, L.; Geelen, D. Agricultural Functions and Action Mechanisms of Plant Biostimulants (PBs). In The Chemical Biology of Plant Biostimulants; John Wiley & Sons, Ltd: Hoboken, NJ, USA, 2020; pp. 1–30. ISBN 978-1-119-35725-4. [Google Scholar]
- Halpern, M.; Bar-Tal, A.; Ofek, M.; Minz, D.; Muller, T.; Yermiyahu, U. Chapter Two—The Use of Biostimulants for Enhancing Nutrient Uptake. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2015; Volume 130, pp. 141–174. ISBN 0065-2113. [Google Scholar]
- Rouphael, Y.; Giordano, M.; Cardarelli, M.; Cozzolino, E.; Mori, M.; Kyriacou, M.; Bonini, P.; Colla, G. Plant- and Seaweed-Based Extracts Increase Yield but Differentially Modulate Nutritional Quality of Greenhouse Spinach through Biostimulant Action. Agronomy 2018, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Rouphael, Y.; Canaguier, R.; Svecova, E.; Cardarelli, M. Biostimulant Action of a Plant-Derived Protein Hydrolysate Produced through Enzymatic Hydrolysis. Front. Plant Sci. 2014, 5, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palumbo, G.; Schiavon, M.; Nardi, S.; Ertani, A.; Celano, G.; Colombo, C.M. Biostimulant Potential of Humic Acids Extracted from an Amendment Obtained via Combination of Olive Mill Wastewaters (OMW) and a Pre-Treated Organic Material Derived from Municipal Solid Waste (MSW). Front. Plant Sci. 2018, 9, 1028. [Google Scholar] [CrossRef] [PubMed]
- Goñi, O.; Łangowski, Ł.; Feeney, E.; Quille, P.; O’Connell, S. Reducing Nitrogen Input in Barley Crops While Maintaining Yields Using an Engineered Biostimulant Derived from Ascophyllum Nodosum to Enhance Nitrogen Use Efficiency. Front. Plant Sci. 2021, 12, 789. [Google Scholar] [CrossRef]
- Ertani, A.; Pizzeghello, D.; Altissimo, A.; Nardi, S. Use of Meat Hydrolyzate Derived from Tanning Residues as Plant Biostimulant for Hydroponically Grown Maize. J. Plant Nutr. Soil Sci. 2013, 176, 287–295. [Google Scholar] [CrossRef]
- Zrig, A.; AbdElgawad, H.; Touneckti, T.; Mohamed, H.B.; Hamouda, F.; Khemira, H. Potassium and Calcium Improve Salt Tolerance of Thymus Vulgaris by Activating the Antioxidant Systems. Sci. Hortic. 2021, 277, 109812. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Agarwal, R.M. Potassium Up-Regulates Antioxidant Metabolism and Alleviates Growth Inhibition under Water and Osmotic Stress in Wheat (Triticum Aestivum L). Protoplasma 2017, 254, 1471–1486. [Google Scholar] [CrossRef]
- Rehman, H.U.; Alharby, H.F.; Alzahrani, Y.; Rady, M.M. Magnesium and Organic Biostimulant Integrative Application Induces Physiological and Biochemical Changes in Sunflower Plants and Its Harvested Progeny on Sandy Soil. Plant Physiol. Biochem. 2018, 126, 97–105. [Google Scholar] [CrossRef]
- Barroso, F.D.L.; Milagres, C.D.C.; Fontes, P.C.R.; Cecon, P.R. Magnesium-Influenced Seed Potato Development and Yield. J. Plant Nutr. 2021, 44, 296–308. [Google Scholar] [CrossRef]
- Kroh, G.E.; Pilon, M. Regulation of Iron Homeostasis and Use in Chloroplasts. Int. J. Mol. Sci. 2020, 21, 3395. [Google Scholar] [CrossRef] [PubMed]
- Salama, D.M.; Osman, S.A.; Abd El-Aziz, M.E.; Abd Elwahed, M.S.A.; Shaaban, E.A. Effect of Zinc Oxide Nanoparticles on the Growth, Genomic DNA, Production and the Quality of Common Dry Bean (Phaseolus Vulgaris). Biocatal. Agric. Biotechnol. 2019, 18, 101083. [Google Scholar] [CrossRef]
- Szczepanek, M.; Siwik-Ziomek, A. P and K Accumulation by Rapeseed as Affected by Biostimulant under Different NPK and S Fertilization Doses. Agronomy 2019, 9, 477. [Google Scholar] [CrossRef] [Green Version]
- Jannin, L.; Arkoun, M.; Etienne, P.; Laîné, P.; Goux, D.; Garnica, M.; Fuentes, M.; Francisco, S.S.; Baigorri, R.; Cruz, F.; et al. Brassica Napus Growth Is Promoted by Ascophyllum Nodosum (L.) Le Jol. Seaweed Extract: Microarray Analysis and Physiological Characterization of N, C, and S Metabolisms. J. Plant Growth Regul. 2013, 32, 31–52. [Google Scholar] [CrossRef]
- Li, J.; Guan, Y.; Yuan, L.; Hou, J.; Wang, C.; Liu, F.; Yang, Y.; Lu, Z.; Chen, G.; Zhu, S. Effects of Exogenous IAA in Regulating Photosynthetic Capacity, Carbohydrate Metabolism and Yield of Zizania Latifolia. Sci. Hortic. 2019, 253, 276–285. [Google Scholar] [CrossRef]
- Saini, S.; Sharma, I.; Kaur, N.; Pati, P.K. Auxin: A Master Regulator in Plant Root Development. Plant Cell Rep. 2013, 32, 741–757. [Google Scholar] [CrossRef]
- Zhang, L.; Miras-Moreno, B.; Yildiztugay, E.; Ozfidan-Konakci, C.; Arikan, B.; Elbasan, F.; Ak, G.; Rouphael, Y.; Zengin, G.; Lucini, L. Metabolomics and Physiological Insights into the Ability of Exogenously Applied Chlorogenic Acid and Hesperidin to Modulate Salt Stress in Lettuce Distinctively. Molecules 2021, 26, 6291. [Google Scholar] [CrossRef]
- Zhang, L.; Martinelli, E.; Senizza, B.; Miras-Moreno, B.; Yildiztugay, E.; Arikan, B.; Elbasan, F.; Ak, G.; Balci, M.; Zengin, G.; et al. The Combination of Mild Salinity Conditions and Exogenously Applied Phenolics Modulates Functional Traits in Lettuce. Plants 2021, 10, 1457. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, C.B.; Rouina, B.B.; Sensoy, S.; Boukhriss, M.; Abdullah, F.B. Exogenous Proline Effects on Photosynthetic Performance and Antioxidant Defense System of Young Olive Tree. J. Agric. Food Chem. 2010, 58, 4216–4222. [Google Scholar] [CrossRef] [PubMed]
- Lucini, L.; Miras-Moreno, B.; Rouphael, Y.; Cardarelli, M.; Colla, G. Combining Molecular Weight Fractionation and Metabolomics to Elucidate the Bioactivity of Vegetal Protein Hydrolysates in Tomato Plants. Front. Plant Sci. 2020, 11, 976. [Google Scholar] [CrossRef] [PubMed]
- Sohag, A.A.M.; Tahjib-Ul-Arif, M.; Polash, M.A.S.; Chowdhury, M.B.; Afrin, S.; Burritt, D.J.; Murata, Y.; Hossain, M.A.; Afzal Hossain, M. Exogenous Glutathione-Mediated Drought Stress Tolerance in Rice (Oryza Sativa L.) Is Associated with Lower Oxidative Damage and Favorable Ionic Homeostasis. Iran. J. Sci. Technol. Trans. Sci. 2020, 44, 955–971. [Google Scholar] [CrossRef]
- Zhou, Y.; Diao, M.; Cui, J.-X.; Chen, X.-J.; Wen, Z.-L.; Zhang, J.-W.; Liu, H.-Y. Exogenous GSH Protects Tomatoes against Salt Stress by Modulating Photosystem II Efficiency, Absorbed Light Allocation and H2O2-Scavenging System in Chloroplasts. J. Integr. Agric. 2018, 17, 2257–2272. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Leaves FW | Leaves DW | Roots FW | Roots DW | Stem FW | Stem DW | Lateral Shoots FW | Lateral Shoots DW | Number of Leaves | |
|---|---|---|---|---|---|---|---|---|---|
| (g) | (g) | (g) | (g) | (g) | (g) | (g) | (g) | (n) | |
| Control | 1.19 (0.27) b | 0.59 (0.08) b | 20.61 (3.23) a | 3.19 (0.34) a | 2.46 (0.25) a | 1.37 (0.10) a | 0.28 (0.04) b | 0.09 (0.03) b | 18.4 (3.11) b |
| BIO 0.25 | 3.68 (0.30) a | 1.48 (0.10) a | 19.23 (4.43) a | 3.24 (0.62) a | 2.95 (0.41) a | 1.48 (0.21) a | 0.72 (0.19) a | 0.24 (0.02) a | 46.0 (7.48) a |
| BIO 0.5 | 1.76 (0.27) a | 0.81 (0.10) a | 22.18 (4.93) a | 3.80 (0.85) a | 3.02 (0.30) a | 1.62 (0.13) a | 0.47 (0.04) a | 0.13 (0.03) a | 24.8 (2.52) a |
| BIO 1 | 2.62 (0.58) a | 1.17 (0.21) a | 17.86 (4.00) a | 3.06 (0.54) a | 2.55 (0.10) a | 1.36 (0.08) a | 0.45 (0.09) a | 0.21 (0.04) a | 33.4 (7.32) a |
| p = 0.0015 | p = 0.0013 | p = 0.0034 | p = 0.0042 | p = 0.0027 |
| N | K | Ca | Mg | Na | Fe | Mn | Zn | Cu | |
|---|---|---|---|---|---|---|---|---|---|
| (mg g−1 DW) | (µg g−1 DW) | ||||||||
| Control | 1.84 (0.60) b | 59.8 (7.1) b | 3.68 (0.15) b | 0.52 (0.07) b | 0.50 (0.10) a | 23.8 (2.4) b | 13.7 (3.5) a | 9.1 (0.10) b | 16.8 (4.8) a |
| BIO 0.25 | 3.10 (0.15) a | 78.9 (4.5) a | 3.70 (0.07) b | 0.70 (0.01) a | 0.51 (0.12) a | 25.0 (2.0) b | 17.8 (4.4) a | 8.8 (0.6) b | 15.6 (3.0) a |
| BIO 0.5 | 2.80 (0.10) a | 81.6 (5.7) a | 4.56 (0.86) ab | 0.76 (0.03) a | 0.65 (0.11) a | 43.2 (7.7) a | 12.6 (5.6) a | 9.3 (0.3) b | 18.2 (2.0) a |
| BIO 1 | 3.00 (0.01) a | 95.7 (15.4) a | 5.30 (0.3) a | 0.71 (0.01) a | 0.64 (0.07) a | 51.8 (7.8) a | 14.0 (5.4) a | 11.9 (0.3) a | 17.5 (1.3) a |
| p = 0.0051 | p = 0.0093 | p = 0.0079 | p = 0.0004 | p = 0.0006 | p = 0.00004 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Regni, L.; Del Buono, D.; Miras-Moreno, B.; Senizza, B.; Lucini, L.; Trevisan, M.; Morelli Venturi, D.; Costantino, F.; Proietti, P. Biostimulant Effects of an Aqueous Extract of Duckweed (Lemna minor L.) on Physiological and Biochemical Traits in the Olive Tree. Agriculture 2021, 11, 1299. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11121299
Regni L, Del Buono D, Miras-Moreno B, Senizza B, Lucini L, Trevisan M, Morelli Venturi D, Costantino F, Proietti P. Biostimulant Effects of an Aqueous Extract of Duckweed (Lemna minor L.) on Physiological and Biochemical Traits in the Olive Tree. Agriculture. 2021; 11(12):1299. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11121299
Chicago/Turabian StyleRegni, Luca, Daniele Del Buono, Begoña Miras-Moreno, Biancamaria Senizza, Luigi Lucini, Marco Trevisan, Diletta Morelli Venturi, Ferdinando Costantino, and Primo Proietti. 2021. "Biostimulant Effects of an Aqueous Extract of Duckweed (Lemna minor L.) on Physiological and Biochemical Traits in the Olive Tree" Agriculture 11, no. 12: 1299. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11121299