1. Introduction
In many Mediterranean areas, high evapotranspiration rates, increased precipitation variability, and intense summer drought are the main environmental constraints to agricultural management and crop production. Under such conditions, sustainable agricultural practices and the introduction of new crops that are able to diversify cropping systems and to mitigate climate changes are very promising strategies for addressing cropping system sustainability.
Compared to continuous cropping systems, crop diversification involves better utilization of land resources, lower risks from pests and diseases, and greater yield stability. In this context, medicinal and aromatic plants (MAPs) can be included in low-input productive systems: they can enhance the multifunctionality traits of the agricultural sector, produce safe final products, respect the environment, and promote rural areas. Various MAPs find optimal growth conditions in Mediterranean environments and, among these, perennial and semi-perennial species can contribute to agro-ecosystem services.
The cultivation of perennial crops reduces soil erosion, minimizes nutrient leaching, sequesters more C in soils, protects water resources, creates a better pest tolerance, and provides a continuous habitat for wildlife [
1,
2,
3]. In addition, perennial species require less use of farm equipment than annual crops as well as fewer fertilizers and herbicides [
4]. Among perennial MAPs,
Stevia rebaudiana (Bertoni) is grown successfully in a wide range of agro-ecological environments, from semi-humid, subtropical to temperate zones. Thanks to its extreme versatility, it can be grown as a pluriannual crop (three to five years) in temperate to warm climates and, as an annual crop, in colder regions [
5,
6,
7]. Unlike other traditional sweetener crops, such as sugar beet and sugar cane, stevia cultivation reduces agronomic inputs, including nutrients, water, and energy [
8].
Besides these agronomic benefits, stevia has an exceptional phytocomplex composition that can be exploited in several applications, from the pharmaceutical to cosmetic and nutraceutical industry. Metabolic disorders such as type-II diabetes and obesity, associated with an excessive sugar consumption, are becoming more prevalent and stevia is the perfect sugar substitute in foods and drinks, given that its leaves contain non-calorie high-intensive sweetener compounds, namely steviol glycosides (SVglys) [
9,
10,
11]. Its leaves also contain a complex mixture of triterpenoids, sterols, essential oils, phenols, and flavonoids, with functional and health-promoting properties [
12,
13,
14,
15,
16,
17]. Thanks to its positive agronomic and phytochemical characteristics, stevia could thus offer new solutions in terms of cropping system diversification and marketing opportunities.
Stevia cultivation is widespread all over the world and it has been introduced as a commercial crop in several countries [
8,
18]. Experimental cultivations are increasing in Europe aimed at producing higher-performing and yielding stevia genotypes [
19,
20,
21,
22,
23]. Access to genotypes that can adapt to different environmental conditions is important for the selection of those characterized by both high levels of secondary metabolites and biomass yield.
To improve the competitiveness of stevia production in the Mediterranean region, it is important to identify higher-performing genotypes in terms of yield and quality, resource use efficiency, and resistance/tolerance to a wide range of biotic/abiotic stress combinations. Therefore, the aim of this study was to determine the best performing genotypes suitable for developing site-specific recommendations for stevia cultivation under the Mediterranean climate of central Italy. Nine stevia genotypes and two harvest times were compared for two consecutive years, exploring differences in plant growth traits (growth cycle, biometric characteristics, leaf yield) and in desired compounds (SVglys content and profile, total phenol and flavonoid accumulation, and antioxidant activity).
4. Discussion
We assessed the agronomic and phytochemical performances of nine S. rebaudiana (Bertoni) genotypes for two consecutive growing seasons. The aim was to select the best productive genetic resources to develop new and improved cultivars for the Mediterranean area. To improve the competitiveness of stevia production as a semi-perennial crop in this environment, it is important to produce higher-performing crops in terms of yield and quality. Therefore, the identification of new varieties/cultivars of S. rebaudiana with long stand duration, consistently high and stable leaf and SVglys yields, as well as a high level of other beneficial substances, is the top priority for the successful introduction of stevia into Mediterranean cropping systems.
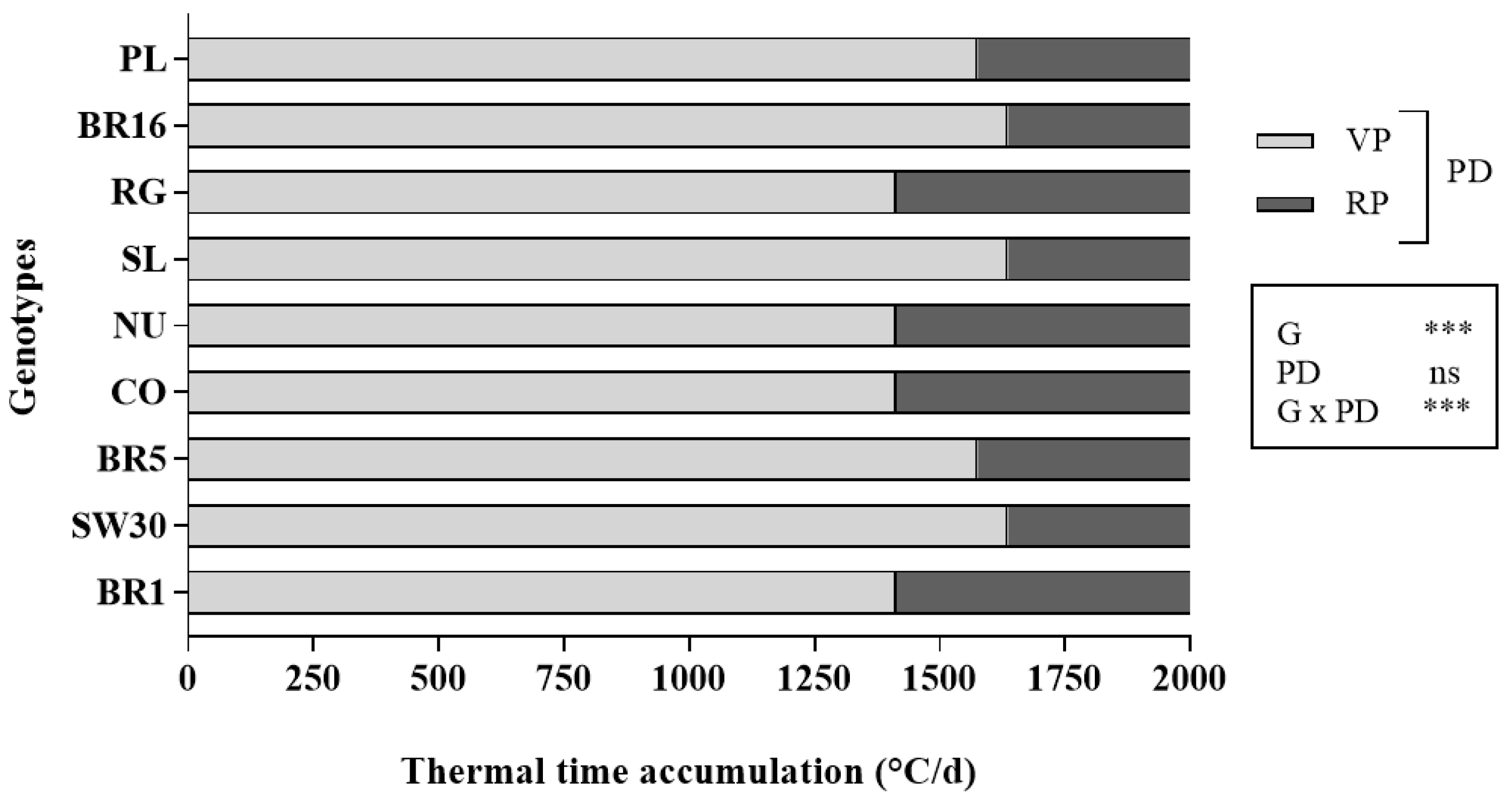
Our results showed that some of the genotypes tested were characterized by a longer vegetative period before flowering, as demonstrated by the higher thermal requirements (
Figure 1). This behavior promotes both higher leaf yield and steviol glycoside concentration, which in turn determine higher SVglys yield. This is important given the time required for the plant to synthesize and accumulate the steviol glycosides in the leaves. In fact, the maximum accumulation of these compounds, which depends on several environmental, agronomic, and physiological factors, is reached when the plant moves from the budding phase to an initial flowering stage with less than 10% flowers [
31]. Therefore, the greater the vegetative period, the greater the accumulation of steviol glycosides in the leaves. When the plant starts to flower, nutrients accumulate in the reproductive organs and, as a result, vegetative growth declines.
Given that the leaves are the commercially important part of stevia, a delay in flowering can enhance vegetative growth and economic yield. In this regard, Ceunen and Geuns [
31] observed large amounts of SVglys within the upper leaves during the budding phase and, as more and more buds become flowers, leaves contained lower amounts of SVglys. This is important for the choice of best harvesting time, since the maximum SVglys concentration is reached in the physiological stage of flowering-bud formation [
32,
33]. Flowering and, in general, plant growth are also affected by the photoperiod and temperature. Stevia is, in fact, a short-day plant with a critical photoperiod of between 12–13 h of day length [
34,
35]. Regarding air temperature, in our environment vegetative growth was lower when the maximum day temperature was below 10 °C or over 35 °C.
Overall, our results showed that genotype, crop age and harvest time represented key pre-harvest factors for defining the morphological, phenological, and quanti-qualitative traits in stevia.
Regarding differences in plant morphology and canopy architecture, we confirmed previous findings [
21,
36,
37], which reported a very high variability, due to genetic characteristics, has been reported for plant height, basal stem diameter, branching, specific leaf weight, and leaf yield. In order to investigate this great variability among stevia genotypes and populations, some authors [
21,
22] have observed that increased yields were correlated to a high leaf area index (LAI), which is responsible for a greater light interception and, in turn, for a higher leaf photosynthesis.
We assessed the specific leaf weight (SLW), a leaf thickness index that is positively correlated with leaf photosynthesis: leaves with high SLW values are thicker and generally have a higher chlorophyll density per area unit (µg cm
−2) and, therefore, greater photosynthetic capacities than thinner leaves [
38]. SLW significantly changed depending on genotype, crop age and harvest time. In particular, in 2-year-old plants, an increase in SLW was accompanied by increased leaf dry yields, suggesting that, thanks to a greater SLW leaf photosynthesis may have been enhanced with a consequent increase in crop yield [
39].
Plants with a high SLW have leaves with a low surface/volume ratio, which is more efficient in terms of water use [
40]. This ratio plays an important role in leaf functioning and is related to the strategies for acquiring and using the energy resources [
41], as well as a tool to evaluate plant productivity [
42] with a view to sustainable agriculture. For a better comprehension of the relationships between morphotype and light interception and photosynthetic activity, future studies are needed in order to improve leaf yield in
S. rebaudiana.
Regarding stevia productivity, we found that the least productive genotypes are those with more branching, as reported by Tateo et al. [
43]. In addition, our findings suggested a positive relation between plant height and leaf dry yield in agreement with previous studies [
36,
37]. Finally, the aerial biomass variability among genotypes exhibited high heritability, and the differences can also be partly attributed to crop age, pedo-climatic conditions of the cultivation site and plant development stage, as already observed in previous studies [
19,
44,
45].
In our environment, stevia productivity significantly increased from the first to the second year of cultivation, with a very high winter survival rate. This suggests that stevia could grow as a semi-perennial crop in central Italy. This could bring positive agronomic advantages, such as, reductions in soil erosion and nutrient leaching, C soil sequestration, which in turn have important implications in multifunctional and sustainable agriculture.
Our findings confirmed previous results [
5,
6] in which in the temperate area of central Italy, the cultivation of stevia is long-term (5–7 years), with a vegetative period from April–May to September-October. In the tested climatic conditions, during winter, above-ground parts of the plant became dry, and there was regrowth through new shoots in the buried rhizome the next spring, thus producing a new crop without replanting. On the other hand, at higher latitudes as in central Europe (such as Belgium and Germany) and Canada [
46,
47,
48], with cooler winters, stevia behaves like an annual crop with annual replanting.
We thus believe that stevia can be cultivated as a promising new crop in the Mediterranean climate of central Italy, though the SVgly content and composition still need to be optimized.
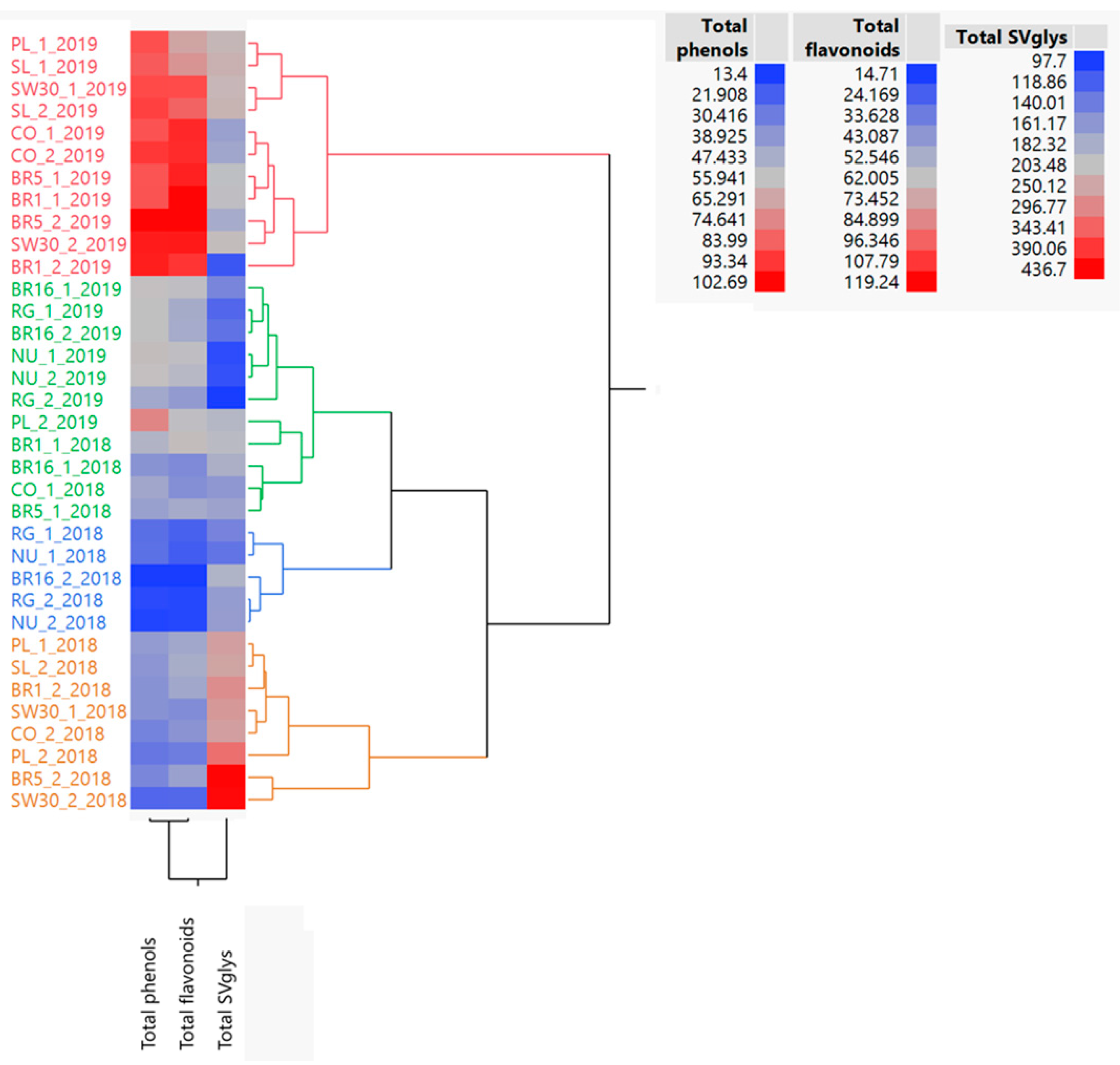
We found that the content and composition in bioactive compounds varied greatly depending on the genotype, crop age and harvest time. Two-year-old plants generally exhibited the highest content of total phenols and flavonoids, as well as the antioxidant activities; conversely, total SVglys content peaked in the establishment year. The sampling carried out at the beginning of the reproductive phase only improved total SVglys, again confirming that, in stevia, plant development is a primary factor in influencing the biosynthetic pathway.
Harvesting when plants were in full vegetative development (July harvest) maximized the content of polyphenol compounds and improved the antioxidant activities. Similar results have been obtained in a previous work [
49] highlighting how the identification of the optimal harvest time was able to maximize the bioactive compounds of interest and, consequently, the health-promoting properties of stevia leaves. However, since the biosynthesis of secondary metabolites is a plant defense mechanism against biotic and abiotic stresses, through many physiological, biochemical, and molecular changes in plant metabolism, it is not always easy to identify the optimal time in which the different metabolites are maximally expressed.
The accumulation of polyphenols during the beginning of flowering stage could be related to the ecological roles of these compounds, such as intensifying antifungal defenses and attracting pollinators [
32]. In addition, since different interactions among environmental and agronomic factors may occur, it is difficult to select individual stimuli that can influence a single metabolic pathway.
Of the secondary metabolites that have been synthesized and accumulated in stevia leaves, the presence of phenolic compounds is the subject of increasing interest because of their significant practical use for nutritional and medicinal applications. In fact, there are important implications for the growing market of natural stevia products [
50], where they are employed as natural preservatives, thanks to their capacity to delay the oxidative degradation of lipids and to improve the shelf life of foods and beverages. They are also involved in the prevention of oxidative stress in humans, thanks to the hydroxyl groups in their molecules which have antioxidant anti-inflammatory properties [
51,
52].
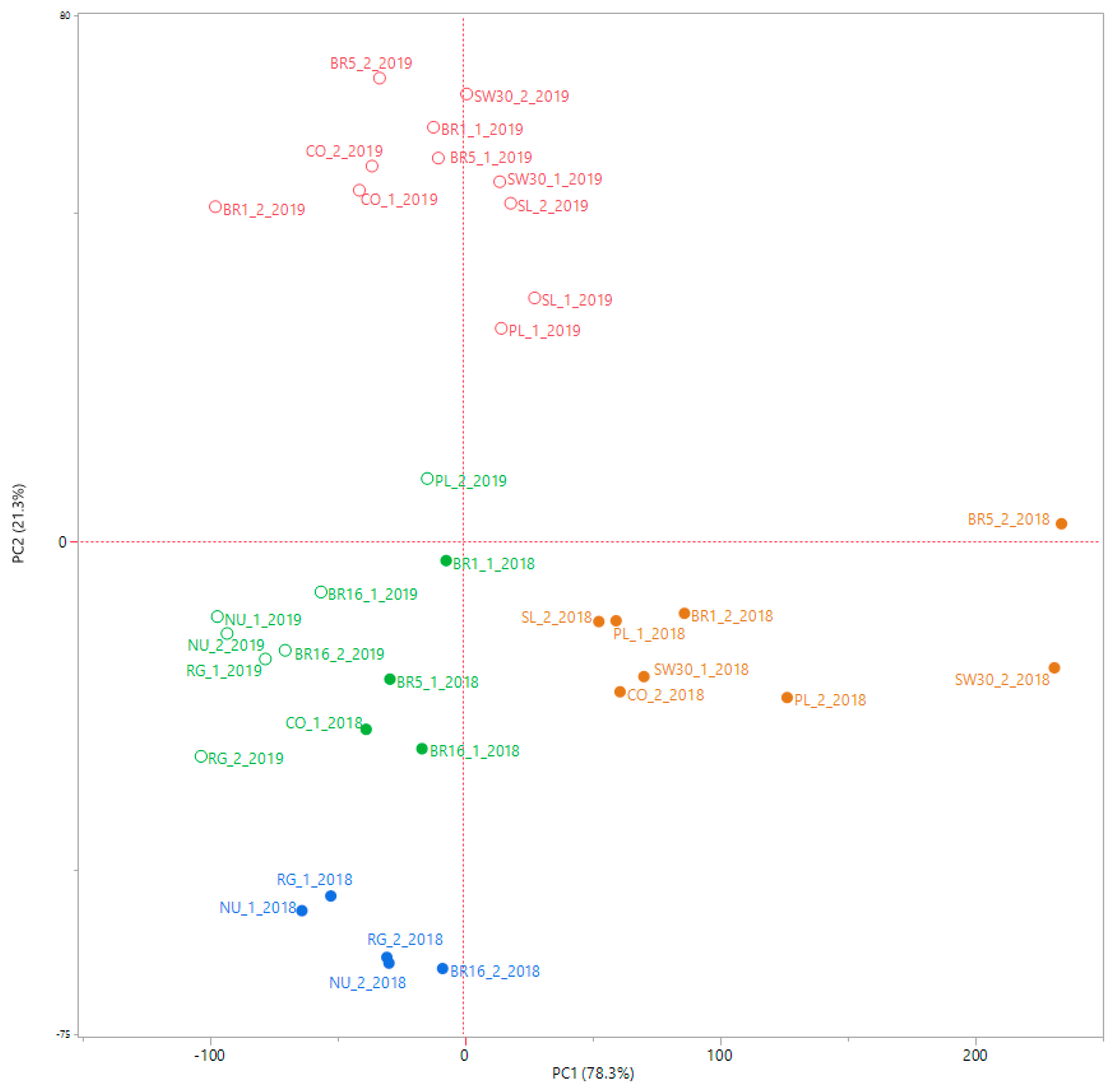
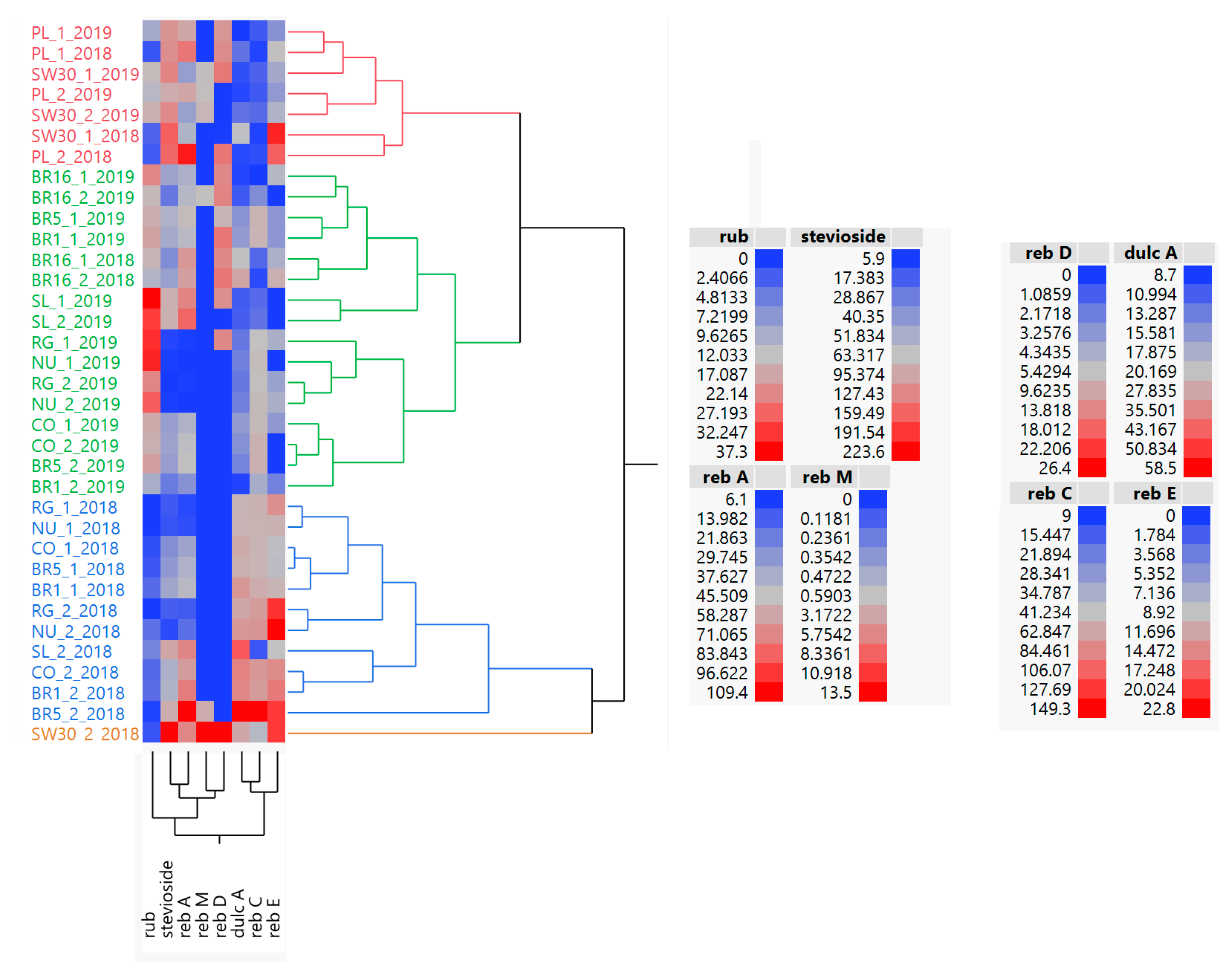
Our findings provide new knowledge about the dissimilarity among genotypes, which was above all generated by the total SVglys content with respect to total phenols and flavonoids. Previous studies have investigated the effect of farming practices and genotype × environment interaction on stevia crop productivity, Reb A and Stev accumulation and on their reciprocal ratio (Reb A/Stev) [
19,
21,
22,
53,
54,
55]. Our results showed that the differences in the steviol glycoside profile are less due to crop age and more to genetics, as suggested by the dendrogram of the SVglys profile. It is well-known that, while the SVglys content in the leaves can vary depending on environmental and agronomic factors [
56,
57], the SVglys qualitative profile remains quite stable, indicating the high genotypic determinism of this trait [
19].
We thus believe that our findings are a good starting point to screen the best genotypes in terms of SVglys profile, taking into account that breeding programs for stevia are increasingly aimed at varieties with an optimal Reb A/Stev ratio, which is considered a good qualitative measure of sweetness, or with a high content of Reb M and Reb D, characterized by a very sweet taste with no liquorice-like taste. We found that genotypes, such as SL, BR5, PL, and SW30 accumulated significantly high amounts of Reb A, Reb M, and Reb D. Among these genotypes, SL and BR5 were characterized by relatively low content of Stevioside. Conversely, PL and SW30 exhibited high contents of Stevioside, which negatively affect the Reb A/Stev ratio, lowering it below 1.
In our study, we try to identify the stevia ideotype, deriving from the combination of morphological, productive, and phytochemical traits. Consequently, considering not only the SVglys profile, but also leaf yield, growth crop cycle and the composition of their whole phytocomplex (polyphenols, total SVglys and their related antioxidant activities), SL, BR5, PL, and SW30 seemed to be the best performing genotypes, in the given environment, and thus suitable as starting point for future breeding programs.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}