Effects of Different Storage Conditions on the Browning Degree, PPO Activity, and Content of Chemical Components in Fresh Lilium Bulbs (Liliumbrownii F.E.Brown var. viridulum Baker.)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Instruments
2.2. Plant Material
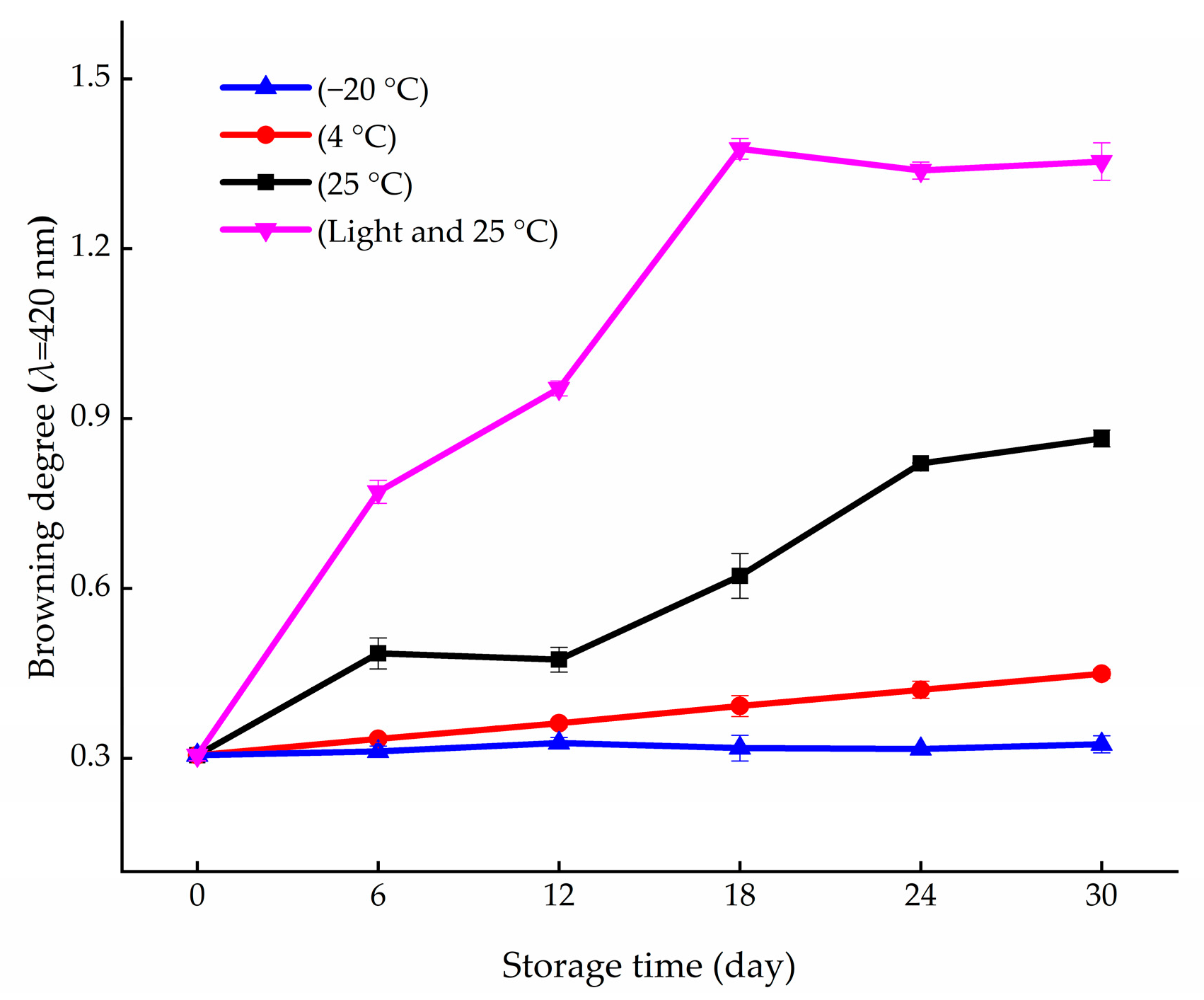
2.3. Measurement of Browning Degree
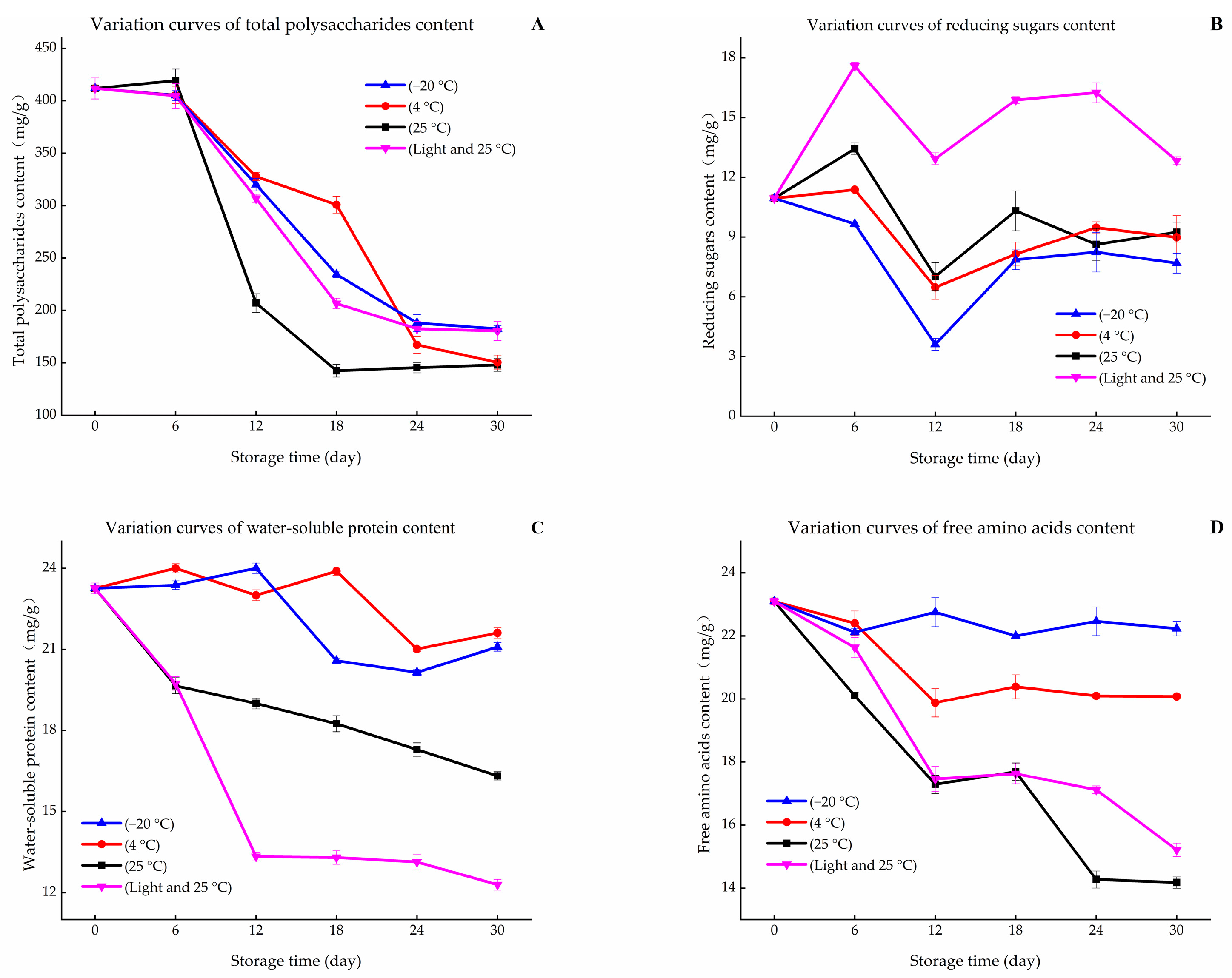
2.4. Measurement of Total Carbohydrates
2.5. Measurement of Total Reducing Sugars
2.6. Measurement the Content of Water-Soluble Protein
2.7. Measurement of Free Amino Acids Content
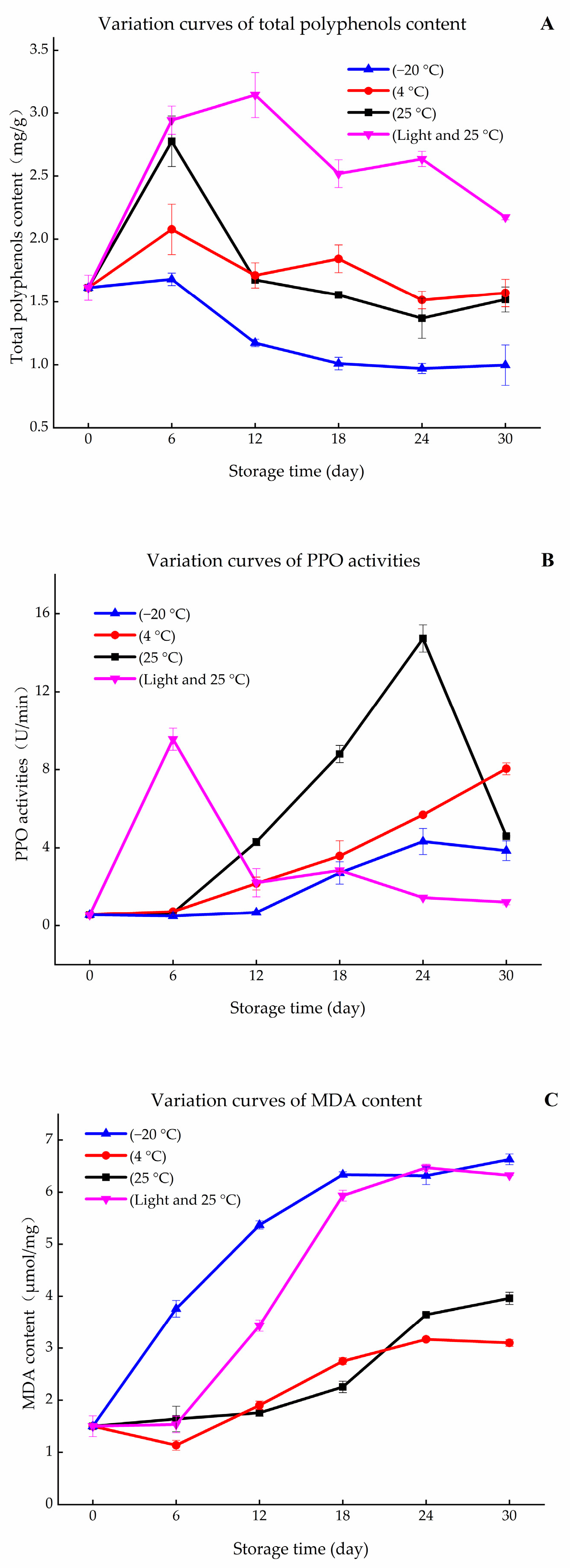
2.8. Measurement of Total Phenolics Content
2.9. Measurement of Polyphenol Oxidase Activity
2.10. Measurement of MDA Content
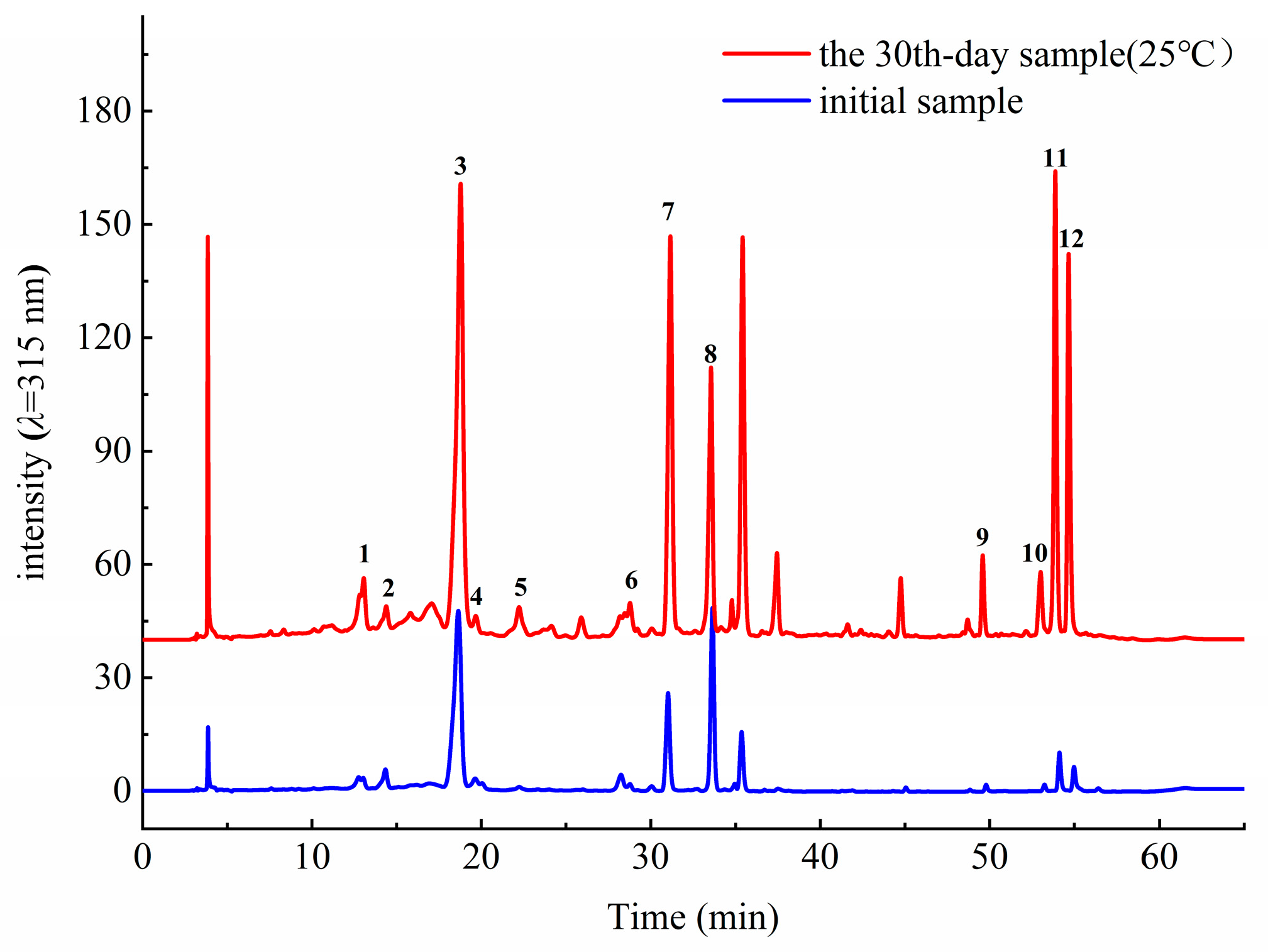
2.11. High-Performance Liquid Chromatography Quadrupole Time-of-Flight Mass Spectrometry (HPLC-Q-TOF-MS) Conditions
2.12. Statistical Analysis
3. Results and Discussion
3.1. Variation in Browning and Contents of Primary Metabolites during Storage Periods
3.2. Variation in PPO Activity and Contents of MDA and Total Polyphenols during Storage Periods
3.3. Identification of Phenolic Compounds Via High-Performance Liquid Chromatography Quadrupole Time-of-Flight Mass Spectrometry (HPLC-Q-TOF-MS)
3.4. Analysis of the Relative Contents of the 12 Compounds via HPLC
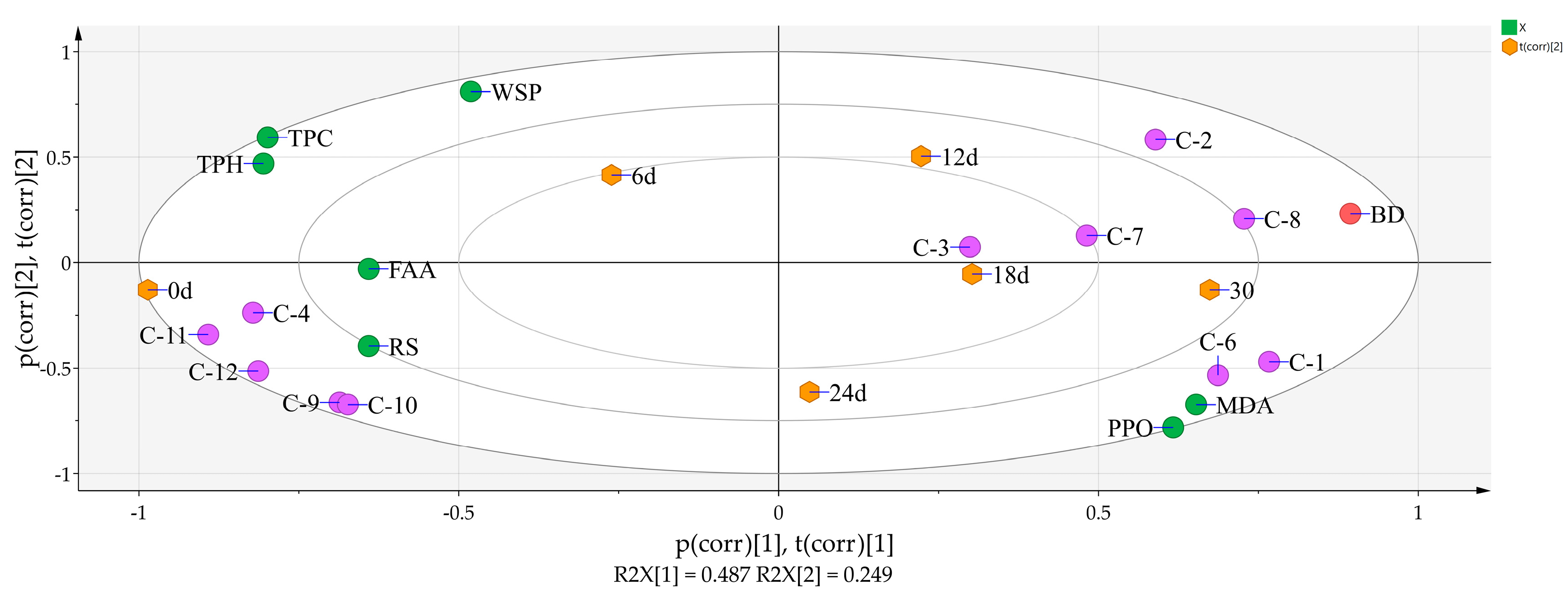
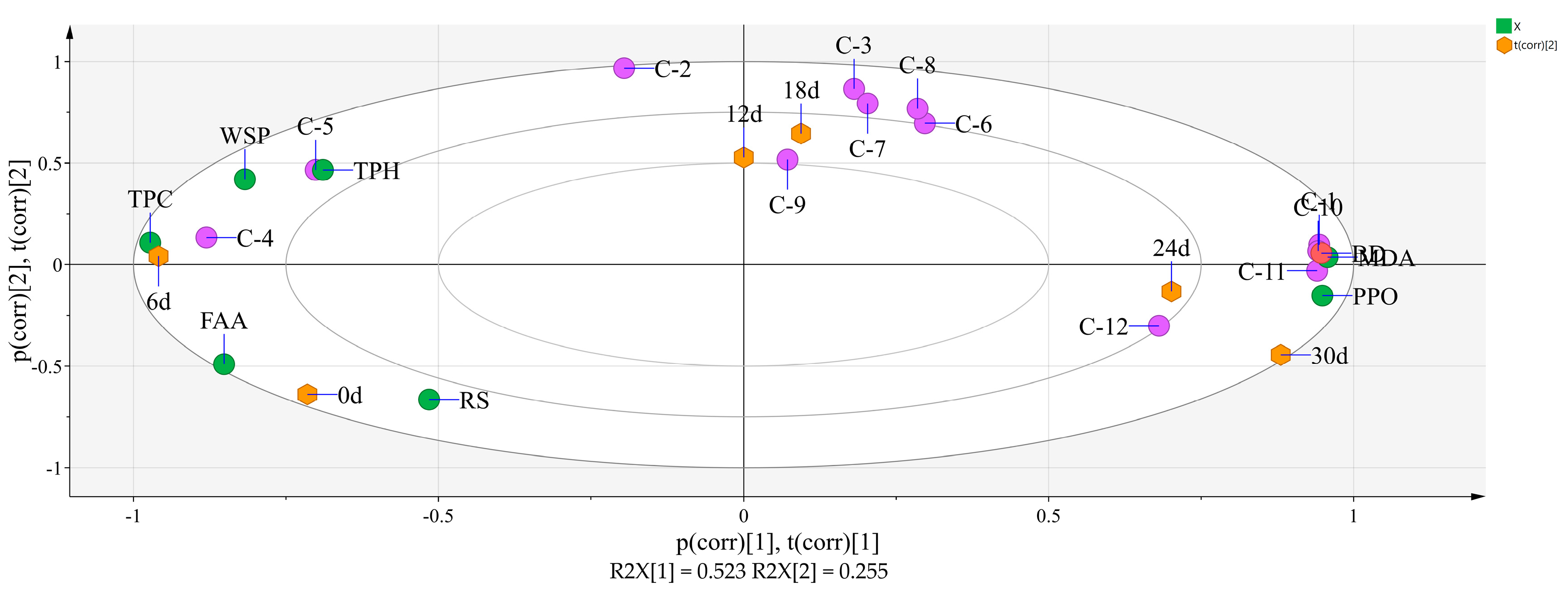
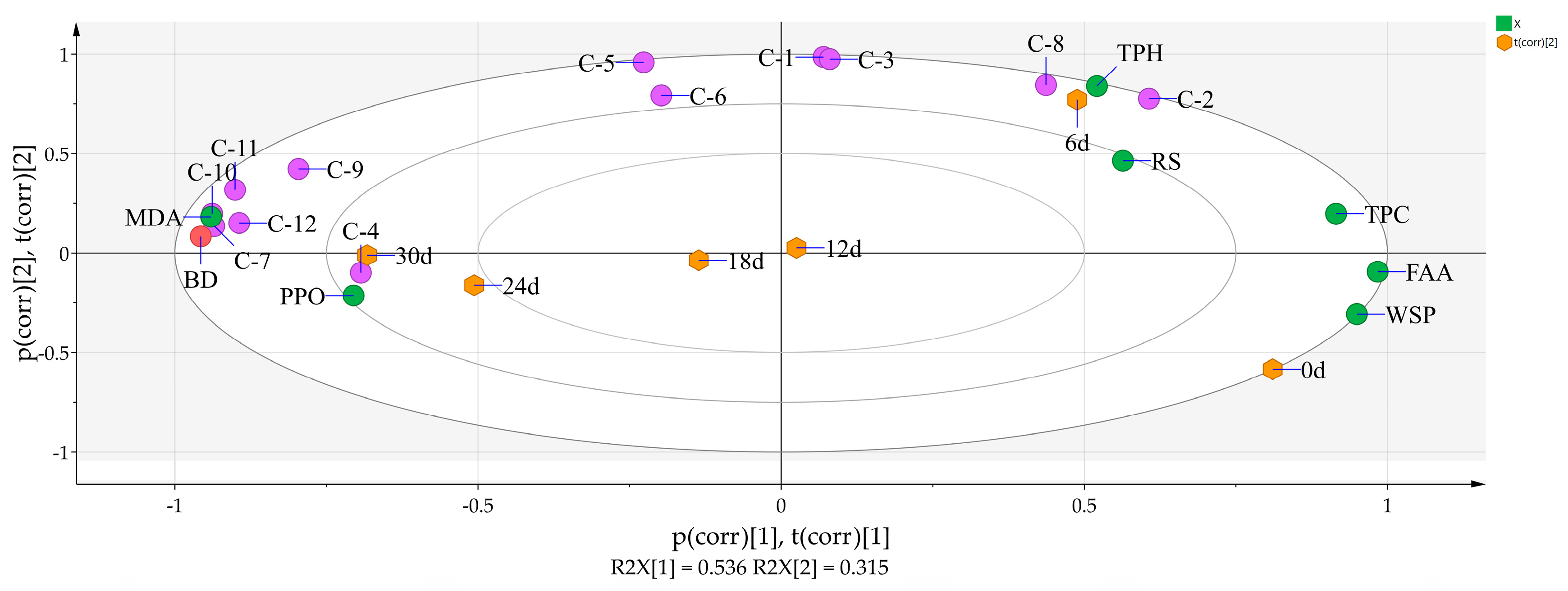
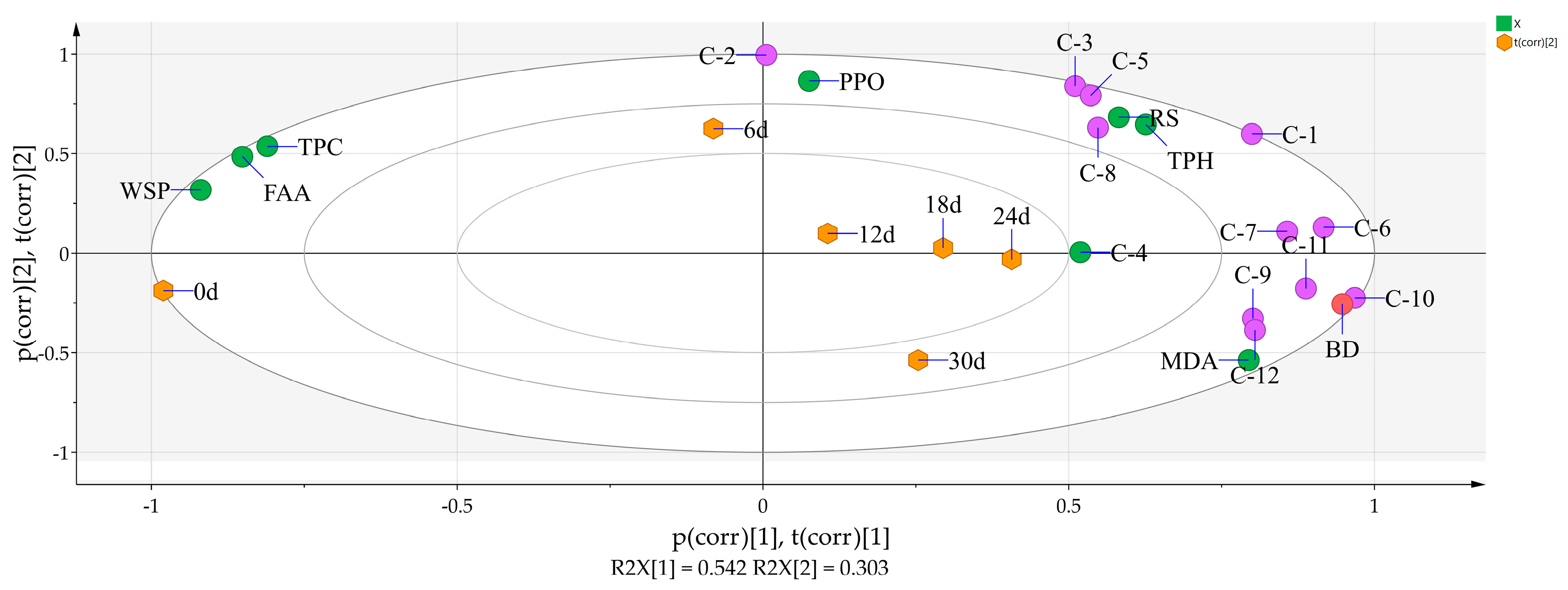
3.5. Principal Component Analysis of 8 Physicochemical Parameters, the 12 Phenolic Compounds, and Storage Time
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, T.; Huang, H.; Zhang, Y.; Li, X.; Li, H.; Jiang, Q.; Gao, W. Role of effective composition on antioxidant, anti-inflammatory, sedative-hypnotic capacities of 6 common edible Lilium varieties. J. Food Sci. 2015, 80, H857–H868. [Google Scholar] [CrossRef] [PubMed]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People Republic of China; Chinese Medical Science and Technology Press: Beijing, China, 2020; pp. 132–133. [Google Scholar]
- Hou, Y.; Jiang, J.G. Origin and concept of medicine food homology and its application in modern functional foods. Food Funct. 2013, 4, 1727–1741. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Wang, Z.; Zhang, Y.M.; Luo, J.G.; Kong, L.Y. Bioassay-guided isolation of anti-inflammatory components from the bulbs of Lilium brownii var. viridulum and identifying the underlying mechanism through acting on the NF-kappaB/MAPKs Pathway. Molecules 2017, 22, 506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, X.X.; Luo, J.G.; Guo, C.; Kong, L.Y. New steroidal saponins from the bulbs of Lilium brownii var. viridulum. Carbohyd. Res. 2012, 361, 19–26. [Google Scholar] [CrossRef]
- Pan, G.; Xie, Z.; Huang, S.; Tai, Y.; Cai, Q.; Jiang, W.; Sun, J.; Yuan, Y. Immune-enhancing effects of polysaccharides extracted from Lilium lancifolium Thunb. Int. Immunopharmacol. 2017, 52, 119–126. [Google Scholar] [CrossRef]
- Munafo, J.P., Jr.; Gianfagna, T.J. Quantitative analysis of phenylpropanoid glycerol glucosides in different organs of Easter Lily (Lilium longiflorum Thunb.). J. Agric. Food Chem. 2015, 63, 4836–4842. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Li, Z.Y.; Zhao, H.Q.; Gao, C.; Xiao, M.W.; Jiang, X.M.; Zhu, J.P.; Huang, H.Y.; Xu, G.M.; Xie, M.Z. Effects of different drying methods on the chemical constituents of Lilium lancifolium Thunb. based on UHPLC-MS analysis and antidepressant activity of the main chemical component regaloside A. J. Sep. Sci. 2020. [Google Scholar] [CrossRef]
- Azadi, P.; Khosh-Khui, M. Micropropagation of Lilium ledebourii (Baker) Boiss as affected by plant growth regulator, sucrose concentration, harvesting season and cold treatments. Electron. J. Biotechnol. 2007, 10, 582–591. [Google Scholar] [CrossRef]
- Aydin, B.; Gulcin, I.; Alwasel, S.H. Purification and characterization of polyphenol oxidase from Hemsin Apple (Malus communis L.). Int. J. Food Prop. 2015, 18, 2735–2745. [Google Scholar] [CrossRef] [Green Version]
- Segovia-Bravo, K.A.; Jarén-Galán, M.; García-García, P.; Garrido-Fernández, A. Browning reactions in olives: Mechanism and polyphenols involved. Food Chem. 2009, 114, 1380–1385. [Google Scholar] [CrossRef]
- Krzyzanowski, A.; Saleeb, M.; Elofsson, M. Synthesis of Indole-, Benzo[b]thiophene-, and Benzo[b] selenophene-Based Analogues of the Resveratrol Dimers Viniferifuran and (+/−)-Dehydroampelopsin B. Org. Lett. 2018, 20, 6650–6654. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, R.F.; Biais, B.; Richard, T.; Puertas, B.; Waffo-Teguo, P.; Merillon, J.M.; Cantos-Villar, E. Grapevine cane’s waste is a source of bioactive stilbenes. Ind. Crop. Prod. 2016, 94, 884–892. [Google Scholar] [CrossRef]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Saura, D.; Vegara, S.; Marti, N.; Valero, M.; Laencina, J. Non-enzymatic browning due to storage is reduced by using clarified lemon juice as acidifier in industrial-scale production of canned peach halves. J. Food Sci. Technol. 2017, 54, 1873–1881. [Google Scholar] [CrossRef]
- Nooshkam, M.; Varidi, M.; Bashash, M. The Maillard reaction products as food-born antioxidant and antibrowning agents in model and real food systems. Food Chem. 2019, 275, 644–660. [Google Scholar] [CrossRef]
- Cha, J.; Debnath, T.; Lee, K.G. Analysis of alpha-dicarbonyl compounds and volatiles formed in Maillard reaction model systems. Sci. Rep. 2019, 9, 5325. [Google Scholar] [CrossRef] [Green Version]
- Farcuh, M.; Copes, B.; Le-Navenec, G.; Marroquin, J.; Cantu, D.; Bradford, K.J.; Guinard, J.X.; Van Deynze, A. Sensory, physicochemical and volatile compound analysis of short and long shelf-life melon (Cucumis melo L.) genotypes at harvest and after postharvest storage. Food Chem. 2020, 8, 100107. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Feng, L.; Zhang, C.; Bao, Y.; He, Y. Identifying freshness of Spinach leaves stored at different temperatures using hyperspectral imaging. Foods 2019, 8, 356. [Google Scholar] [CrossRef] [Green Version]
- Kan, J.; Xie, W.J.; Wan, B.; Huo, T.B.; Lin, X.P.; Liu, J.; Jin, C.H. Heat-induced tolerance to browning of fresh-cut lily bulbs (Lilium lancifolium Thunb.) under cold storage. J. Food Biochem. 2019, 43, e12816. [Google Scholar] [CrossRef] [PubMed]
- Hoch, W.A.; Singsaas, E.L.; McCown, B.H. Resorption protection. Anthocyanins facilitate nutrient recovery in autumn by shielding leaves from potentially damaging light levels. Plant Physiol. 2003, 133, 1296–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logan, B.A.; Stafstrom, W.C.; Walsh, M.J.L.; Reblin, J.S.; Gould, K.S. Examining the photoprotection hypothesis for adaxial foliar anthocyanin accumulation by revisiting comparisons of green- and red-leafed varieties of coleus (Solenostemon scutellarioides). Photosynth. Res. 2015, 124, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.S.; Moon, K.D. Comparison of image analysis methods to evaluate the degree of browning of fresh-cut lettuce. Food Sci. Biotechnol. 2014, 23, 1043–1048. [Google Scholar] [CrossRef]
- Zhang, J.; Gao, Y.; Zhou, X.; Hu, L.; Xie, T. Chemical characterisation of polysaccharides from Lilium davidii. Nat. Prod. Res. 2010, 24, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wu, S.S.; Liang, R.H.; Liu, W.; Liu, C.M.; Shuai, X.X.; Wang, Z.J. The effect of high speed shearing on disaggregation and degradation of pectin from creeping fig seeds. Food Chem. 2014, 165, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Song, W.; Zhao, J.; Zhang, Z.; Zhang, Y. Some physical properties of protein moiety of Alkali-extracted tea polysaccharide conjugates were shielded by its polysaccharide. Molecules 2017, 22, 914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Institute of Measurement and Testing Technology. Tea-Determination of Free Amino Acids Content; AQSIQ: Beijing, China, 2014; Volume GB/T 8314-2013. [Google Scholar]
- Jin, L.; Zhang, Y.; Yan, L.; Guo, Y.; Niu, L. Phenolic compounds and antioxidant activity of bulb extracts of six Lilium species native to China. Molecules 2012, 17, 9361–9378. [Google Scholar] [CrossRef] [PubMed]
- Siddiq, M.; Dolan, K.D. Characterization of polyphenol oxidase from blueberry (Vaccinium corymbosum L.). Food Chem. 2017, 218, 216–220. [Google Scholar] [CrossRef]
- Janero, D.R. Malondialdehyde and thiobarbituric acid-reactivity as diagnostic indices of lipid peroxidation and peroxidative tissue injury. Free Radic. Biol. Med. 1990, 9, 515–540. [Google Scholar] [CrossRef]
- Zhao, K.H.; Zhou, F.; Yan, S.E.; Liu, D.B.; Xie, H.Q. The analysis of compounds from Longya Lilium via HPLC-Q-TOF-MS and HS-SPME-GC-MS. Nat. Prod. Res. Dev. 2020, 32, 1331–1342. [Google Scholar] [CrossRef]
- Narbona, E.; Jaca, J.; del Valle, J.C.; Valladares, F.; Buide, M.L. Whole-plant reddening in Silene germana is due to anthocyanin accumulation in response to visible light. Plant Biol. 2018, 20, 968–977. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Ali, S.; Zandi, P.; Mehmood, A.; Ullah, S.; Ikram, M.; Ismail, M.A.S.; Babar, A. Role of Sugars, Amino acids and organic acids in improving plant abiotic stress tolerance. Pak. J. Bot. 2020, 52, 355–363. [Google Scholar] [CrossRef]
- Sun, Y.J.; Shi, Z.D.; Jiang, Y.P.; Zhang, X.H.; Li, X.A.; Li, F.J. Effects of preharvest regulation of ethylene on carbohydrate metabolism of apple (Malus domestica Borkh cv. Starkrimson) fruit at harvest and during storage. Sci. Hortic. 2021, 276, 109748. [Google Scholar] [CrossRef]
- Saddhe, A.A.; Manuka, R.; Penna, S. Plant sugars: Homeostasis and transport under abiotic stress in plants. Physiol. Plant. 2020. [Google Scholar] [CrossRef] [PubMed]
- Das, B.; Sahoo, R.N.; Pargal, S.; Krishna, G.; Verma, R.; Chinnusamy, V.; Sehgal, V.K.; Gupta, V.K.; Dash, S.K.; Swain, P. Quantitative monitoring of sucrose, reducing sugar and total sugar dynamics for phenotyping of water-deficit stress tolerance in rice through spectroscopy and chemometrics. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2018, 192, 41–51. [Google Scholar] [CrossRef]
- Furtauer, L.; Weiszmann, J.; Weckwerth, W.; Nagele, T. Dynamics of plant metabolism during cold acclimation. Int. J. Mol. Sci. 2019, 20, 5411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.H.; Picha, D.H.; Kilili, A.W.; Johnson, C.E. Changes in invertase activities and reducing sugar content in sweetpotato stored at different temperatures. J. Agric. Food Chem. 1999, 47, 4927–4931. [Google Scholar] [CrossRef]
- Hierl, G.; Vothknecht, U.; Gietl, C. Programmed cell death in Ricinus and Arabidopsis: The function of KDEL cysteine peptidases in development. Physiol. Plant. 2012, 145, 103–113. [Google Scholar] [CrossRef]
- Chodankar, S.; Aswal, V.K.; Kohlbrecher, J.; Vavrin, R.; Wagh, A.G. Small-angle neutron scattering study of structure and kinetics of temperature-induced protein gelation. Phys. Rev. E 2009, 79, 021912. [Google Scholar] [CrossRef] [Green Version]
- Goldford, J.E.; Hartman, H.; Marsland, R.; Segre, D. Environmental boundary conditions for the origin of life converge to an organo-sulfur metabolism. Nat. Ecol. Evol. 2019, 3, 1715–1724. [Google Scholar] [CrossRef] [PubMed]
- Park, J.E.; Kim, J.; Purevdorj, E.; Son, Y.J.; Nho, C.W.; Yoo, G. Effects of long light exposure and drought stress on plant growth and glucosinolate production in pak choi (Brassica rapa subsp. chinensis). Food Chem. 2021, 340, 128167. [Google Scholar] [CrossRef]
- Di Gioia, M.L.; Leggio, A.; Malagrino, F.; Romio, E.; Siciliano, C.; Liguori, A. N-methylated α-amino acids and peptides: Synthesis and biological activity. Mini. Rev. Med. Chem. 2016, 16, 683–690. [Google Scholar] [CrossRef]
- Yu, H.; Seow, Y.X.; Ong, P.K.C.; Zhou, W. Kinetic study of high-intensity ultrasound-assisted Maillard reaction in a model system of d-glucose and glycine. Food Chem. 2018, 269, 628–637. [Google Scholar] [CrossRef]
- Quan, W.; Wu, Z.L.; Jiao, Y.; Liu, G.P.; Wang, Z.J.; He, Z.Y.; Tao, G.J.; Qin, F.; Zeng, M.M.; Chen, J. Exploring the relationship between potato components and Maillard reaction derivative harmful products using multivariate statistical analysis. Food Chem. 2021, 339, 127853. [Google Scholar] [CrossRef] [PubMed]
- Aubert, C.; Chalot, G.; Lurol, S.; Ronjon, A.; Cottet, V. Relationship between fruit density and quality parameters, levels of sugars, organic acids, bioactive compounds and volatiles of two nectarine cultivars, at harvest and after ripening. Food Chem. 2019, 297, 124954. [Google Scholar] [CrossRef]
- Shi, L.Y.; Cao, S.F.; Shao, J.R.; Chen, W.; Yang, Z.F.; Zheng, Y.H. Chinese bayberry fruit treated with blue light after harvest exhibit enhanced sugar production and expression of cryptochrome genes. Postharvest Biol. Technol. 2016, 111, 197–204. [Google Scholar] [CrossRef]
- Yonny, M.E.; Torressi, A.R.; Nazareno, M.A.; Cerutti, S. Development of a novel, sensitive, selective, and fast methodology to determine malondialdehyde in leaves of melon plants by ultra-high-performance liquid chromatography-tandem mass spectrometry. J. Anal. Methods Chem. 2017, 2017, 4327954. [Google Scholar] [CrossRef]
- Yan, S.L.; Yang, T.B.; Luo, Y.G. The mechanism of ethanol treatment on inhibiting lettuce enzymatic browning and microbial growth. LWT-Food Sci. Technol. 2015, 63, 383–390. [Google Scholar] [CrossRef]
- Brillante, L.; De Rosso, M.; Dalla Vedova, A.; Maoz, I.; Flamini, R.; Tomasi, D. Insights on the stilbenes in Raboso Piave grape (Vitis vinifera L.) as a consequence of postharvest vs on-vine dehydration. J. Sci. Food Agric. 2018, 98, 1961–1967. [Google Scholar] [CrossRef] [PubMed]
- Alfeo, V.; Bravi, E.; Ceccaroni, D.; Sileoni, V.; Perretti, G.; Marconi, O. Effect of baking time and temperature on nutrients and phenolic compounds content of fresh sprouts breadlike product. Foods 2020, 9, 1447. [Google Scholar] [CrossRef] [PubMed]
- Suleman, P.; Redha, A.; Afzal, M.; Al-Hasan, R. Temperature-induced changes of malondialdehyde, heat-shock proteins in relation to chlorophyll fluorescence and photosynthesis in Conocarpus lancifolius (Engl.). Acta Physiol. Plant. 2013, 35, 1223–1231. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Jin, H.B.; Suo, J.W.; Yu, W.Y.; Zhou, M.Y.; Dai, W.S.; Song, L.L.; Hu, Y.Y.; Wu, J.S. Effect of temperature and humidity on oil quality of harvested Torreya grandis cv. Merrillii nuts during the after-ripening stage. Front. Plant. Sci. 2020, 11, 573681. [Google Scholar] [CrossRef]
- Peng, X.Y.; Du, C.; Yu, H.Y.; Zhao, X.Y.; Zhang, X.Y.; Wang, X.Y. Purification and characterization of polyphenol oxidase (PPO) from water yam (Dioscorea alata). CyTA J. Food 2019, 17, 676–684. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.H.; Li, C.Y.; Wei, M.L.; Ge, Y.H.; Tang, Q.; Xue, W.J.; Zhang, S.Y.; Wang, W.H.; Lv, J.Y. Effects of trisodium phosphate treatment after harvest on storage quality and sucrose metabolism in jujube fruit. J. Sci. Food Agric. 2019, 99, 5526–5532. [Google Scholar] [CrossRef]
- Balasooriya, H.N.; Dassanayake, K.B.; Seneweera, S.; Ajlouni, S. Impact of elevated carbon dioxide and temperature on strawberry polyphenols. J. Sci. Food Agric. 2019, 99, 4659–4669. [Google Scholar] [CrossRef]
- Simsek, M.; Quezada-Calvillo, R.; Nichols, B.L.; Hamaker, B.R. Phenolic compounds increase the transcription of mouse intestinal maltase-glucoamylase and sucrase-isomaltase. Food Funct. 2017, 8, 1915–1924. [Google Scholar] [CrossRef] [PubMed]
- Mo, E.J.; Ahn, J.H.; Jo, Y.H.; Kim, S.B.; Hwang, B.Y.; Lee, M.K. Inositol derivatives and phenolic compounds from the roots of Taraxacum coreanum. Molecules 2017, 22, 1349. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Li, L.; Kong, L. Preparative separation of phenylpropenoid glycerides from the bulbs of Lilium lancifolium by high-speed counter-current chromatography and evaluation of their antioxidant activities. Food Chem. 2012, 131, 1056–1062. [Google Scholar] [CrossRef]
- Yin, L.B.; Liao, C.; Yang, A.L.; Liu, D.; He, P.; Liu, Y.L.; Li, L.L. Effect of the controlled atmosphere storage on the quality of Lilium brownii bulbs. Int. J. Agric. Biol. 2020, 24, 1607–1613. [Google Scholar] [CrossRef]
- Li, X.D.; Wu, B.H.; Wang, L.J.; Zheng, X.B.; Yan, S.T.; Li, S.H. Changes in trans-resveratrol and other phenolic compounds in grape skin and seeds under low temperature storage after post-harvest UV-irradiation. J. Hortic. Sci. Biotechnol. 2009, 84, 113–118. [Google Scholar] [CrossRef]
- Billet, K.; Houille, B.; Besseau, S.; Melin, C.; Oudin, A.; Papon, N.; Courdavault, V.; Clastre, M.; Giglioli-Guivarc’h, N.; Lanoue, A. Mechanical stress rapidly induces E-resveratrol and E-piceatannol biosynthesis in grape canes stored as a freshly-pruned byproduct. Food Chem. 2018, 240, 1022–1027. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Storage Time (d) | 0 | 6 | 12 | 18 | 24 | 30 |
|---|---|---|---|---|---|---|
| Compound-1 | 107.06 ± 1.12 | 104.74 ± 4.01 | 134.35 ± 3.37 a | 143.99 ± 3.24 | 173.91 ± 0.91a | 232.56 ± 14.82 a |
| Compound-2 | 269.81 ± 0.77 | 366.66 ± 21.92 a | 468.30 ± 12.85 a | 370.04 ± 25.44 a | 350.94 ± 18.63 | 350.21 ± 9.19 |
| Compound-3 | 3792.36 ± 10.64 | 3328.57 ± 41.58 | 4510.88 ± 37.21 b | 3879.37 ± 29.17 a | 3959.79 ± 31.76 | 3818.84 ± 44.48 |
| Compound-4 | 82.43 ± 1.90 | 50.01 ± 2.33 a | 44.24 ± 5.14 | 48.79 ± 2.23 | 49.45 ± 2.00 | 51.25 ± 2.33 |
| Compound-5 | 138.55 ± 8.09 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Compound-6 | 38.17 ± 6.24 | 58.65 ± 6.74 | 123.35 ± 8.91 a | 225.67 ± 13.11 a | 224.93 ± 6.27 | 153.44 ± 0.67 a |
| Compound-7 | 65.15 ± 1.94 | 663.53 ± 11.77 b | 110.70 ± 0.19 | 179.63 ± 10.95 | 124.48 ± 7.34 | 104.61 ± 27.98 |
| Compound-8 | 1902.94 ± 20.62 | 1920.21 ± 34.19 | 2844.26 ± 27.14 a | 2431.70 ± 21.32 | 1731.18 ± 19.50 a | 1741.15 ± 25.31 |
| Compound-9 | 57.18 ± 3.13 | 0.00 | 0.00 | 0.00 | 49.11 ± 2.13 b | 0.00 |
| Compound-10 | 57.13 ± 2.75 | 0.00 | 0.00 | 0.00 | 51.14 ± 4.41 b | 0.00 |
| Compound-11 | 318.89 ± 10.10 | 165.03 ± 7.99 b | 144.05 ± 11.36 | 124.77 ± 18.62 | 246.62 ± 9.93 a | 59.35 ± 4.04 b |
| Compound-12 | 245.41 ± 4.59 | 108.06 ± 9.68b | 102.88 ± 4.40 | 108.51 ± 3.90 | 200.88 ± 6.75 a | 68.01 ± 4.62 b |
| Stor age Time (d) | 0 | 6 | 12 | 18 | 24 | 30 |
|---|---|---|---|---|---|---|
| Compound-1 | 107.06 ± 1.12 | 123.90 ± 6.10 | 243.12 ± 7.70 a | 338.23 ± 9.65 a | 332.02 ± 5.49 | 462.35 ± 2.02 a |
| Compound-2 | 269.81 ± 0.77 | 433.41 ± 16.06 b | 464.93 ± 23.19 | 489.99 ± 8.06 | 331.14 ± 28.23 b | 300.34 ± 11.62 a |
| Compound-3 | 3792.36 ± 10.64 | 4467.63 ± 79.88 a | 8097.52 ± 46.85 b | 6808.66 ± 35.17 b | 4548.36 ± 69.95 b | 4867.97 ± 63.97 |
| Compound-4 | 82.43 ± 1.90 | 126.53 ± 9.02 a | 71.00 ± 7.95 a | 78.89 ± 2.82 | 60.55 ± 4.70 a | 53.73 ± 3.47 a |
| Compound-5 | 138.55 ± 8.09 | 593.93 ± 65.00 b | 350.00 ± 44.35 b | 310.76 ± 7.62 | 182.95 ± 3.43 b | 137.18 ± 6.71 a |
| Compound-6 | 38.17 ± 6.24 | 50.49 ± 2.46 | 106.06 ± 8.87 a | 344.63 ± 9.50 b | 126.13 ± 6.70 b | 88.22 ± 2.24 a |
| Compound-7 | 65.15 ± 1.94 | 1065.39 ± 22.50 b | 1175.18 ± 86.93 | 979.67 ± 18.98 | 979.87 ± 55.00 | 676.12 ± 52.29 b |
| Compound-8 | 1902.94 ± 20.62 | 2011.81 ± 39.71 | 3834.91 ± 57.62 b | 3175.96 ± 78.49 a | 3276.26 ± 15.89 | 1822.14 ± 78.29 b |
| Compound-9 | 57.18 ± 3.13 | 51.12 ± 3.40 | 40.71 ± 3.45 | 142.48 ± 8.46 b | 77.15 ± 7.65 b | 37.51 ± 7.52 b |
| Compound-10 | 57.13 ± 2.75 | 99.40 ± 6.42 | 148.64 ± 8.96 a | 137.33 ± 9.57 | 206.54 ± 4.25 b | 219.61 ± 13.09 |
| Compound-11 | 318.89 ± 10.10 | 312.24 ± 9.63 | 436.58 ± 32.42 b | 466.44 ± 41.59 | 695.25 ± 5.25 b | 588.63 ± 55.47 a |
| Compound-12 | 245.41 ± 4.59 | 295.42 ± 9.24 | 295.20 ± 11.78 | 320.14 ± 6.19 a | 322.86 ± 4.01 | 656.28 ± 20.75 b |
| Storage Time (d) | 0 | 6 | 12 | 18 | 24 | 30 |
|---|---|---|---|---|---|---|
| Compound-1 | 107.06 ± 1.12 | 806.38 ± 83.77 b | 418.99 ± 50.57 b | 390.23 ± 44.67 | 354.21 ± 70.30 | 312.80 ± 26.72 a |
| Compound-2 | 269.81 ± 0.77 | 507.73 ± 63.91 b | 280.24 ± 23.62 b | 221.42 ± 17.38 a | 208.48 ± 10.41 | 203.13 ± 9.90 |
| Compound-3 | 3792.36 ± 10.64 | 10447.69 ± 90.12 b | 7129.24 ± 54.92 b | 6726.39 ± 62.35 a | 6118.67 ± 39.33 | 5647.84 ± 7.67 a |
| Compound-4 | 82.43 ± 1.90 | 86.91 ± 5.02 | 88.66 ± 2.03 | 95.75 ± 3.13 | 115.36 ± 9.63 a | 92.61 ± 3.20a |
| Compound-5 | 138.55 ± 8.09 | 501.10 ± 25.35 b | 327.10 ± 7.31 a | 323.20 ± 7.74 | 332.48 ± 8.64 | 385.84 ± 13.15 |
| Compound-6 | 38.17 ± 6.24 | 231.16 ± 10.56 b | 210.94 ± 14.86 | 200.58 ± 9.78 | 164.02 ± 7.95 a | 116.74 ± 7.85 a |
| Compound-7 | 65.15 ± 1.94 | 1079.15 ± 62.58 b | 1166.15 ± 48.29 | 1170.79 ± 22.29 | 2527.22 ± 14.50 b | 3082.76 ± 36.60 b |
| Compound-8 | 1902.94 ± 20.62 | 3532.78 ± 15.72 b | 2792.35 ± 24.80 a | 2293.82 ± 63.50 a | 1922.79 ± 41.07 a | 1772.28 ± 10.83 |
| Compound-9 | 57.18 ± 3.13 | 231.57 ± 14.57 b | 235.06 ± 5.83 | 246.37 ± 14.22 | 211.59 ± 8.64 | 405.21 ± 10.64 b |
| Compound-10 | 57.13 ± 2.75 | 188.33 ± 9.84 b | 221.71 ± 18.02 a | 245.49 ± 16.89 | 294.58 ± 7.97 | 442.28 ± 14.73 b |
| Compound-11 | 318.89 ± 10.10 | 1298.38 ± 77.36 b | 1372.30 ± 85.52 | 1462.44 ± 28.18 | 1630.94 ± 34.05 a | 2534.24 ± 23.97 b |
| Compound-12 | 245.41 ± 4.59 | 731.93 ± 33.76 b | 879.45 ± 13.08 a | 878.87 ± 23.88 | 1167.70 ± 34.13 a | 2034.22 ± 9.88 b |
| Storage Time (d) | 0 | 6 | 12 | 18 | 24 | 30 |
|---|---|---|---|---|---|---|
| Compound-1 | 107.06 ± 1.12 | 1508.49 ± 5.82 b | 1220.13 ± 18.94 a | 1240.64 ± 3.16 | 1352.80 ± 7.60 | 856.40 ± 12.81 b |
| Compound-2 | 269.81 ± 0.77 | 557.64 ± 8.25 b | 398.62 ± 4.02 a | 365.16 ± 3.84 | 318.01 ± 8.09 | 131.95 ± 3.58 b |
| Compound-3 | 3792.36 ± 10.64 | 19552.51 ± 66.80 b | 14076.69 ± 58.97 a | 12216.46 ± 21.20 | 15310.31 ± 28.22 | 6343.77 ± 22.20 b |
| Compound-4 | 82.43 ± 1.90 | 168.95 ± 25.18 a | 172.03 ± 6.17 | 184.10 ± 8.10 | 738.79 ± 13.22 b | 160.86 ± 4.11 |
| Compound-5 | 138.55 ± 8.09 | 761.45 ± 17.77 b | 565.31 ± 14.00 a | 528.32 ± 11.05 | 476.00 ± 9.13 | 396.46 ± 4.66 a |
| Compound-6 | 38.17 ± 6.24 | 232.56 ± 13.55 b | 319.51 ± 9.52 a | 427.66 ± 5.58 a | 345.16 ± 7.80 | 258.99 ± 6.78 a |
| Compound-7 | 65.15 ± 1.94 | 96.74 ± 4.62 a | 106.42 ± 2.64 | 118.95 ± 5.44 | 146.34 ± 5.92 a | 97.07 ± 2.11 a |
| Compound-8 | 1902.94 ± 20.62 | 5711.26 ± 24.24 b | 5912.23 ± 30.81 | 7707.86 ± 63.75 b | 5743.15 ± 16.20 b | 1777.04 ± 11.74 b |
| Compound-9 | 57.18 ± 3.13 | 305.86 ± 28.12 b | 347.67 ± 14.84 a | 337.14 ± 8.86 | 369.30 ± 11.72 | 604.24 ± 21.24 b |
| Compound-10 | 57.13 ± 2.75 | 251.67 ± 2.22 b | 330.60 ± 18.74 a | 368.88 ± 20.24 | 399.97 ± 15.89 | 441.66 ± 6.57 a |
| Compound-11 | 318.89 ± 10.10 | 1918.76 ± 6.46 b | 2182.49 ± 46.31 | 2000.44 ± 18.32 | 2225.78 ± 35.09 | 3083.81 ± 28.07 b |
| Compound-12 | 245.41 ± 4.59 | 1173.02 ± 28.34 b | 1429.78 ± 37.72 a | 1448.31 ± 28.67 | 1528.76 ± 9.81 | 2548.10 ± 16.24 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, K.; Xiao, Z.; Zeng, J.; Xie, H. Effects of Different Storage Conditions on the Browning Degree, PPO Activity, and Content of Chemical Components in Fresh Lilium Bulbs (Liliumbrownii F.E.Brown var. viridulum Baker.). Agriculture 2021, 11, 184. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11020184
Zhao K, Xiao Z, Zeng J, Xie H. Effects of Different Storage Conditions on the Browning Degree, PPO Activity, and Content of Chemical Components in Fresh Lilium Bulbs (Liliumbrownii F.E.Brown var. viridulum Baker.). Agriculture. 2021; 11(2):184. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11020184
Chicago/Turabian StyleZhao, Kanghong, Zhengpeng Xiao, Jianguo Zeng, and Hongqi Xie. 2021. "Effects of Different Storage Conditions on the Browning Degree, PPO Activity, and Content of Chemical Components in Fresh Lilium Bulbs (Liliumbrownii F.E.Brown var. viridulum Baker.)" Agriculture 11, no. 2: 184. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11020184