
Recurrent Selection with Low Herbicide Rates and Salt Stress Decrease Sensitivity of Echinochloa colona to Imidazolinone

 , , ,
, , ,  ,
,  and
and
Abstract
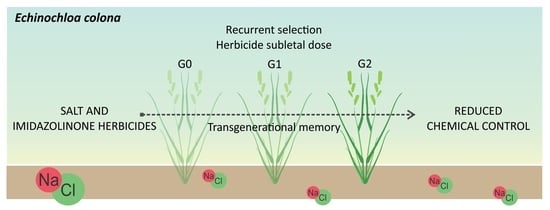
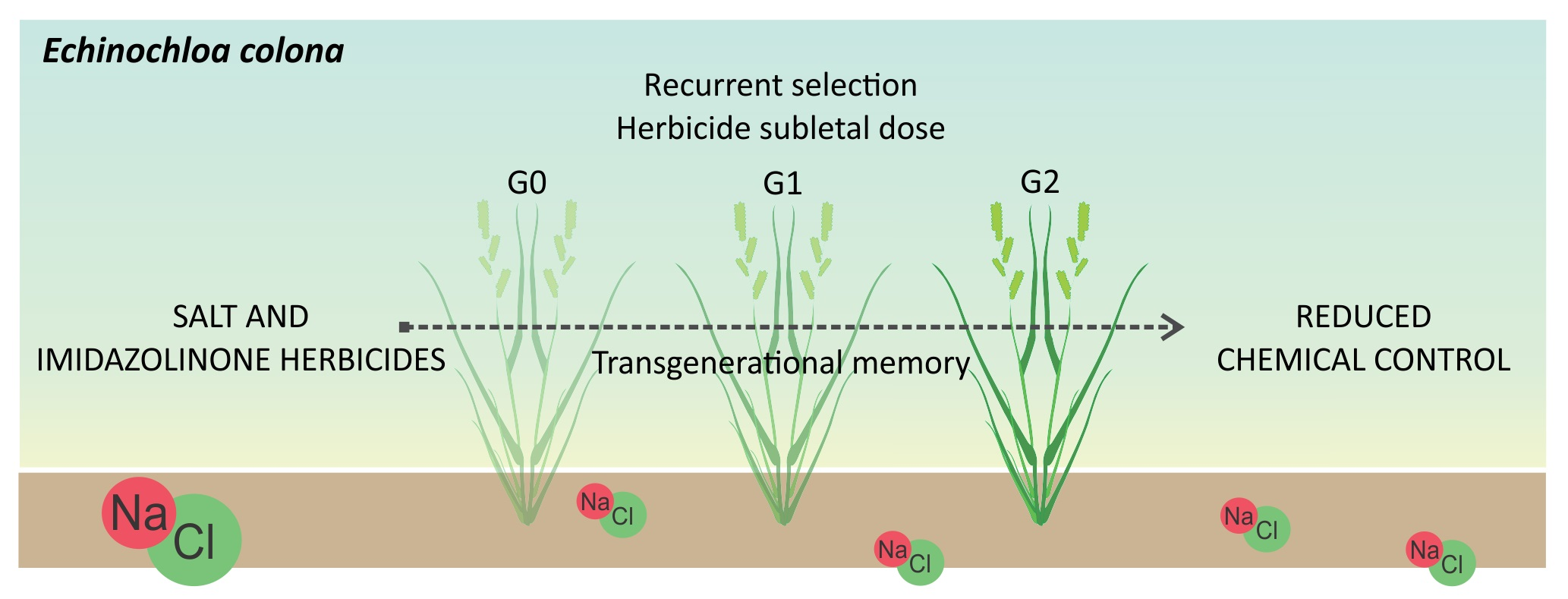
:
1. Introduction
2. Materials and Methods
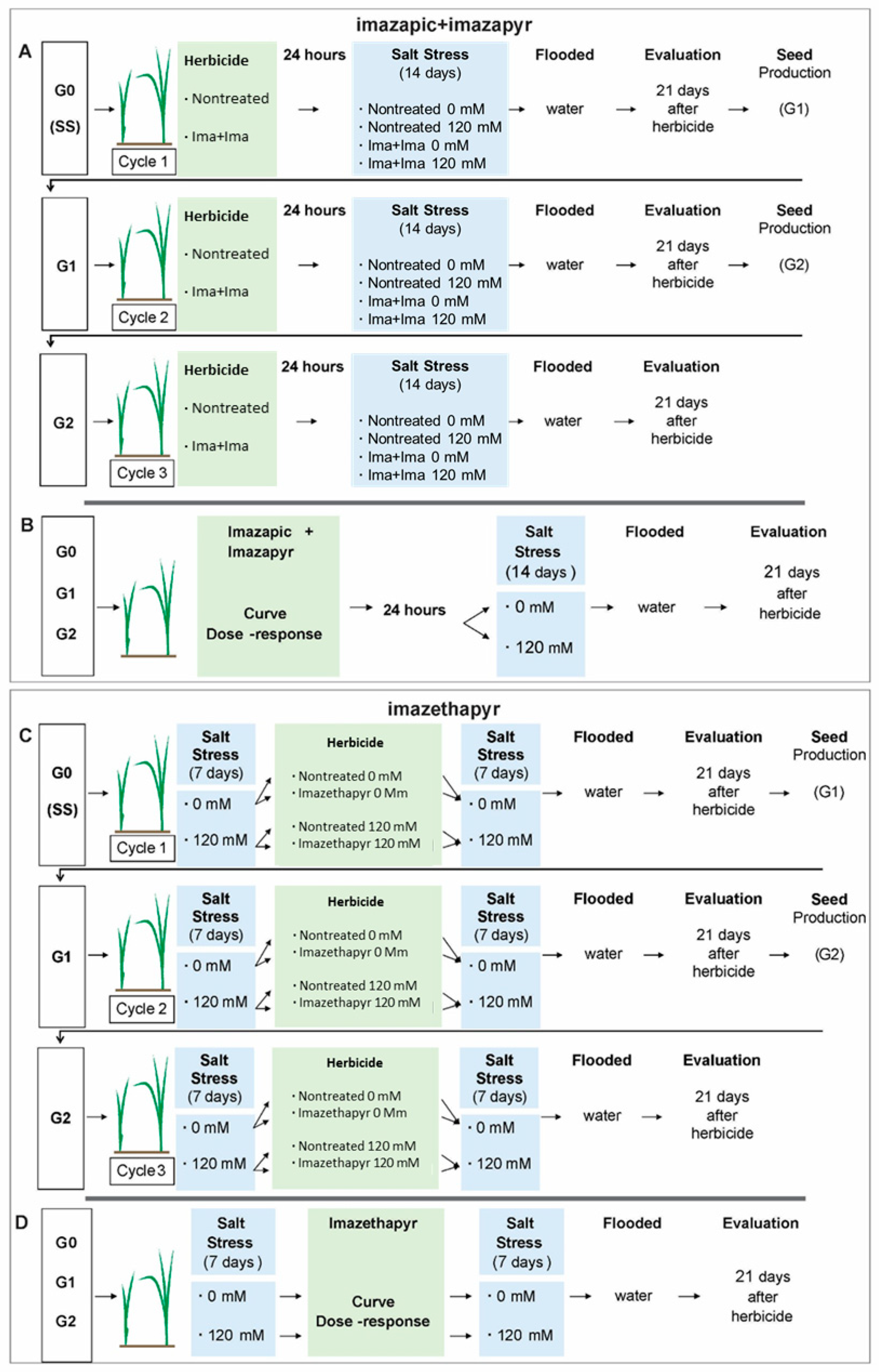
2.1. Experiment I—Effect of Salt Stress and Recurrent Selection With a Sublethal Dose of Imazapic + Imazapyr on the Herbicide Sensitivity of the E. colona Population from Southern Brazil
2.1.1. Plant Material
2.1.2. General Procedure for Population Generation
2.1.3. Determination of Sensitivity Level to Imazapic + Imazapyr
2.2. Experiment II—Effect of Salt Stress and Recurrent Selection With a Sublethal Dose of Imazethapyr on the Herbicide Sensitivity of an E. colona Population from Arkansas, USA
2.2.1. Plant Material
2.2.2. General Procedure for Recurrent Selection
2.2.3. Evaluation of Sensitivity to Imazethapyr
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Desa, U. World Population Projected to Reach 9.8 Billion in 2050, and 11.2 Billion in 2100. Available online: https://www.un.org/development/desa/en/news/population/world-population-prospects-2017.html (accessed on 4 July 2020).
- Croxall, H.E.; Smith, L.P. The Fight for Food Factors Limiting Agricultural Production; Routledge: New York, NY, USA, 2019. [Google Scholar]
- Ziska, L.H.; Gealy, D.R.; Burgos, N.; Caicedo, A.L.; Gressel, J.; Lawton-Rauh, A.L.; Avila, L.A.; Theisen, G.; Norsworthy, J.; Ferrero, A.; et al. Weedy (Red) Rice. An Emerging Constraint to Global Rice Production; Elsevier Ltd.: Amsterdam, The Netherlands, 2015; Volume 129, ISBN 9780128021385. [Google Scholar]
- Jabran, K.; Uludag, A.; Chauhan, B.S. Sustainable Weed Control in Rice. In Weed Control: Sustainability, Hazards, and Risks in Cropping Systems Worldwide; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2018; p. 276. [Google Scholar]
- Peerzada, A.M.; Bajwa, A.A.; Ali, H.H.; Chauhan, B.S. Biology, Impact, and Management of Echinochloa colona (L.) Link. Crop Prot. 2016, 83, 56–66. [Google Scholar] [CrossRef]
- Sudianto, E.; Beng-Kah, S.; Ting-Xiang, N.; Saldain, N.E.; Scott, R.C.; Burgos, N.R. Clearfield® Rice: Its Development, Success, and Key Challenges on a Global Perspective. Crop Prot. 2013, 49, 40–51. [Google Scholar] [CrossRef]
- Merotto, A.; Goulart, I.C.G.R.; Nunes, A.L.; Kalsing, A.; Markus, C.; Menezes, V.G.; Wander, A.E. Evolutionary and Social Consequences of Introgression of Nontransgenic Herbicide Resistance from Rice to Weedy Rice in Brazil. Evol. Appl. 2016, 9, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Bonow, J.F.L.; Lamego, F.P.; Andres, A.; Avila, L.A.; Teló, G.M.; Egewarth, K. Resistance of Echinochloa Crusgalli Var. Mitis to Imazapyr+imazapic Herbicide and Alternative Control in Irrigated Rice. Planta Daninha 2018, 36, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Heap, I. The International Herbicide-Resistant Weed Database. Available online: www.weedscience.org (accessed on 16 July 2020).
- Délye, C.; Jasieniuk, M.; Le Corre, V. Deciphering the Evolution of Herbicide Resistance in Weeds. Trends Genet. 2013, 29, 649–658. [Google Scholar] [CrossRef]
- Zhao, N.; Yan, Y.; Luo, Y.; Zou, N.; Liu, W.; Wang, J. Unravelling Mesosulfuron-Methyl Phytotoxicity and Metabolism-Based Herbicide Resistance in Alopecurus Aequalis: Insight into Regulatory Mechanisms Using Proteomics. Sci. Total Environ. 2019, 670, 486–497. [Google Scholar] [CrossRef]
- Rangani, G.; Salas-Perez, R.A.; Aponte, R.A.; Knapp, M.; Craig, I.R.; Mietzner, T.; Langaro, A.C.; Noguera, M.M.; Porri, A.; Roma-Burgos, N. A Novel Single-Site Mutation in the Catalytic Domain of Protoporphyrinogen Oxidase Ix (PPO) Confers Resistance to PPO-Inhibiting Herbicides. Front. Plant Sci. 2019, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Burgos, N.R.; Heap, I.M.; Rouse, C.E.; Lawton-Rauh, A.L. Evolution of Herbicide-Resistant Weeds. In Weed Control Sustainability, Hazards and Risks in Cropping Systems Worldwide; CRC Press: Boca Raton, FL, USA, 2019; ISBN 9781498719087. [Google Scholar]
- Busi, R.; Girotto, M.; Powles, S.B. Response to Low-Dose Herbicide Selection in Self-Pollinated Avena Fatua. Pest Manag. Sci. 2016, 72, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, L.; Rangani, G.; Ebeling Viana, V.; Carvalho-Moore, P.; Merotto, A.; Rabaioli Camargo, E.; Antonio de Avila, L.; Roma-Burgos, N. Rapid Reduction of Herbicide Susceptibility in Junglerice by Recurrent Selection with Sublethal Dose of Herbicides and Heat Stress. Agronomy 2020, 10, 1761. [Google Scholar] [CrossRef]
- Varanasi, A.; Prasad, P.V.V.; Jugulam, M. Impact of Climate Change Factors on Weeds and Herbicide Efficacy; Elsevier Inc.: Amsterdam, The Netherlands, 2016; Volume 135, ISBN 9780128046937. [Google Scholar]
- Ashworth, M.B.; Walsh, M.J.; Flower, K.C.; Powles, S.B. Recurrent Selection with Reduced 2,4-D Amine Doses Results in the Rapid Evolution of 2,4-D Herbicide Resistance in Wild Radish (Raphanus raphanistrum, L.). Pest Manag. Sci. 2016, 72, 2091–2098. [Google Scholar] [CrossRef] [PubMed]
- Busi, R.; Powles, S.B. Evolution of Glyphosate Resistance in a Lolium Rigidum Population by Glyphosate Selection at Sublethal Doses. Heredity (Edinb.) 2009, 103, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Tehranchian, P.; Norsworthy, J.K.; Powles, S.; Bararpour, M.T.; Bagavathiannan, M.V.; Barber, T.; Scott, R.C. Recurrent Sublethal-Dose Selection for Reduced Susceptibility of Palmer Amaranth (Amaranthus Palmeri) to Dicamba. Weed Sci. 2017, 65, 206–212. [Google Scholar] [CrossRef]
- Vieira, B.C.; Luck, J.D.; Amundsen, K.L.; Gaines, T.A.; Werle, R.; Kruger, G.R. Response of Amaranthus Spp. Following Exposure to Sublethal Herbicide Rates via Spray Particle Drift. PLoS ONE 2019, 14, e0220014. [Google Scholar] [CrossRef] [Green Version]
- Benedetti, L.; Rangani, G.; Viana, V.E.; Carvalho-Moore, P.; Camargo, E.R.; de Avila, L.A.; Roma-Burgos, N. Recurrent Selection by Herbicide Sublethal Dose and Drought Stress Results in Rapid Reduction of Herbicide Sensitivity in Junglerice. Agronomy 2020, 10, 1619. [Google Scholar] [CrossRef]
- Phogat, V.; Pitt, T.; Cox, J.W.; Šimůnek, J.; Skewes, M.A. Soil Water and Salinity Dynamics under Sprinkler Irrigated Almond Exposed to a Varied Salinity Stress at Different Growth Stages. Agric. Water Manag. 2018, 201, 70–82. [Google Scholar] [CrossRef] [Green Version]
- Machado, R.; Serralheiro, R. Soil Salinity: Effect on Vegetable Crop Growth. Management Practices to Prevent and Mitigate Soil Salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- Shahid, S.A.; Zaman, M.; Heng, L. Soil salinity: Historical perspectives and a world overview of the problem. In Guideline for Salinity Assessment, Mitigation and Adaptation Using Nuclear and Related Techniques; Springer: Cham, Switzerland, 2018; pp. 43–53. [Google Scholar]
- Wang, W.; Vinocur, B.; Altman, A. Plant Responses to Drought, Salinity and Extreme Temperatures: Towards Genetic Engineering for Stress Tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef]
- Qadir, M.; Quillérou, E.; Nangia, V.; Murtaza, G.; Singh, M.; Thomas, R.J.; Drechsel, P.; Noble, A.D. Economics of Salt-Induced Land Degradation and Restoration. Nat. Resour. Forum 2014, 38, 282–295. [Google Scholar] [CrossRef]
- Wicke, B.; Smeets, E.; Dornburg, V.; Vashev, B.; Gaiser, T.; Turkenburg, W.; Faaij, A. The Global Technical and Economic Potential of Bioenergy from Salt-Affected Soils. Energy Environ. Sci. 2011, 4, 2669–2681. [Google Scholar] [CrossRef] [Green Version]
- Daliakopoulos, I.N.; Tsanis, I.K.; Koutroulis, A.; Kourgialas, N.N.; Varouchakis, A.E.; Karatzas, G.P.; Ritsema, C.J. The Threat of Soil Salinity: A European Scale Review. Sci. Total Environ. 2016, 573, 727–739. [Google Scholar] [CrossRef]
- Benedetti, L.; Scherner, A.C.; Cuchiara, C.; Moraes, Í.L.A.; Avila, L.; Deuner, S. Soybean Plant Osmotic and Oxidative Stress as Affected by Herbicide and Salinity Levels in Soil. Planta Daninha 2020, 38, 1–7. [Google Scholar] [CrossRef]
- Dahal, K.; Li, X.Q.; Tai, H.; Creelman, A.; Bizimungu, B. Improving Potato Stress Tolerance and Tuber Yield under a Climate Change Scenario—A Current Overview. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant Salt-Tolerance Mechanism: A Review. Biochem. Biophys. Res. Commun. 2018, 495, 286–291. [Google Scholar] [CrossRef]
- Cirillo, V.; Masin, R.; Maggio, A.; Zanin, G. Crop-Weed Interactions in Saline Environments. Eur. J. Agron. 2018, 99, 51–61. [Google Scholar] [CrossRef]
- Bagavathiannan, M.; Singh, V.; Govindasamy, P.; Abugho, S.B.; Liu, R. Impact of concurrent weed or herbicide stress with other biotic and abiotic stressors on crop production. In Plant Tolerance to Individual and Concurrent Stresses; Springer: New Delhi, India, 2017; pp. 33–45. [Google Scholar]
- Islam, F.; Xie, Y.; Farooq, M.A.; Wang, J.; Yang, C.; Gill, R.A.; Zhu, J.; Zhou, W. Salinity Reduces 2,4-D Efficacy in Echinochloa Crusgalli by Affecting Redox Balance, Nutrient Acquisition, and Hormonal Regulation. Protoplasma 2018, 255, 785–802. [Google Scholar] [CrossRef] [PubMed]
- Kollist, H.; Zandalinas, S.I.; Sengupta, S.; Nuhkat, M.; Kangasjärvi, J.; Mittler, R. Rapid Responses to Abiotic Stress: Priming the Landscape for the Signal Transduction Network. Trends Plant Sci. 2019, 24, 25–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markus, C.; Pecinka, A.; Karan, R.; Barney, J.N.; Merotto, A. Epigenetic Regulation-Contribution to Herbicide Resistance in Weeds? Pest Manag. Sci. 2018, 74, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Rouse, C.E. Characterization of Multiple-Herbicide-Resistant Echinochloa Colona from Arkansas. Theses and Dissertations. Available online: https://scholarworks.uark.edu/etd/2582 (accessed on 26 June 2020).
- Ritz, C.; Streibig, J. Analysis of Dose-Response Curve Data. Package ‘Drc’. Available online: https://cran.r-project.org/web/packages/drc/drc.pdf (accessed on 24 June 2020).
- Seefeldt, S.S.; Jensen, J.E.; Feurst, E.P. Log-Logistic Analysis of Herbicide Dose-Response Relationships. Weed Technol. 1995, 9, 218–227. [Google Scholar] [CrossRef]
- BASF Label Kifix®- MAPA Reg. No. Available online: http://www.adapar.pr.gov.br/sites/adapar/arquivos_restritos/files/documento/2020-10/kifix300718.pdf (accessed on 19 June 2020).
- Refatti, J.P.; de Avila, L.A.; Noldin, J.A.; Pacheco, I.; Pestana, R.R. Leaching and Residual Activity of Imidazolinone Herbicides in Lowland Soils. Ciência Rural 2017, 47, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Shaner, L.D. Herbicide Handbook (No. 632.954 W394h10); Weed Science Society of America: Lawrence, KS, USA, 2014. [Google Scholar]
- BASF Label Newpath®-EPA Reg. No. Available online: http://www.cdms.net/ldat/ld4FL012.pdf (accessed on 19 June 2020).
- Concenço, G.; Andres, A.; Schreiber, F.; Moisinho, I.S.; Martins, M.B. Control of Jointvetch (Aeschynomene spp.), Establishment and Productivity of Rice as a Function of [Imazapic + Imazapyr] Doses. J. Agric. Sci. 2018, 10, 287. [Google Scholar] [CrossRef]
- Scherner, A.; Schreiber, F.; Andres, A.; Concenço, G.; Martins, M.B.; Pitol, A. Rice Crop Rotation: A Solution for Weed Management. Rice Crop Curr. Dev. 2018. [Google Scholar] [CrossRef]
- Dyer, W.E. Stress-Induced Evolution of Herbicide Resistance and Related Pleiotropic Effects. Pest Manag. Sci. 2018, 74, 1759–1768. [Google Scholar] [CrossRef]
- Korres, N.E.; Varanasi, V.K.; Slaton, N.A.; Price, A.J.; Bararpour, T. Effects of Salinity on Rice and Rice Weeds: Short-and Long-Term Adaptation Strategies and Weed Management. In Advances in Rice Research for Abiotic Stress Tolerance; Woodhead Publishing: Sawston, UK, 2019; pp. 159–176. [Google Scholar]
- Munns, R.; Day, D.A.; Fricke, W.; Watt, M.; Arsova, B.; Barkla, B.J.; Bose, J.; Byrt, C.S.; Chen, Z.H.; Foster, K.J.; et al. Energy costs of salt tolerance in crop plants. New Phytol. 2020, 225, 1072–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeed, M. Abiotic Stress Tolerance in Rice (Oryza sativa, L.): A Genomics Perspective of Salinity Tolerance. Rice Crop-Curr. Dev. 2018. [Google Scholar] [CrossRef] [Green Version]
- Wani, S.H.; Choudhary, M.; Kumar, P.; Akram, N.A.; Surekha, C.; Ahmad, P.; Gosal, S.S. Marker-assisted breeding for abiotic stress tolerance in crop plants. In Biotechnologies of Crop Improvement; Springer: Cham, Switzerland, 2018; pp. 1–23. [Google Scholar]
- Islam, M.O.; Kato, H.; Shima, S.; Tezuka, D.; Matsui, H.; Imai, R. Functional Identification of a Rice Trehalase Gene Involved in Salt Stress Tolerance. Gene 2019, 685, 42–49. [Google Scholar] [CrossRef]
- Tang, Y.; Bao, X.; Zhi, Y.; Wu, Q.; Guo, Y.; Yin, X.; Zeng, L.; Li, J.; Zhang, J.; He, W.; et al. Overexpression of a Myb Family Gene, Osmyb6, Increases Drought and Salinity Stress Tolerance in Transgenic Rice. Front. Plant Sci. 2019, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, N.M.; Fernandes, T.; Nunes, C.; Rosa, M.T.G.; Matiolli, C.C.; Rodrigues, M.A.A.; Barros, P.M.; Oliveira, M.M.; Abreu, I.A. SUMOylation of Rice DELLA SLR1 Modulates Transcriptional Responses and Improves Yield under Salt Stress. bioRxiv 2020. [Google Scholar] [CrossRef]
- Rastogi, A.; Kovar, M.; He, X.; Zivcak, M.; Kataria, S.; Kalaji, H.M.; Skalicky, M.; Ibrahimova, U.F.; Hussain, S.; Mbarki, S.; et al. JIP-Test as a Tool to Identify Salinity Tolerance in Sweet Sorghum Genotypes. Photosynthetica 2020, 58, 518–528. [Google Scholar] [CrossRef] [Green Version]
- Fogliatto, S.; Serra, F.; Patrucco, L.; Milan, M.; Vidotto, F. Effect of Different Water Salinity Levels on the Germination of Imazamox-Resistant and Sensitive Weedy Rice and Cultivated Rice. Agronomy 2019, 9, 658. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, B.S.; Abugho, S.B.; Amas, J.C.; Gregorio, G.B. Effect of Salinity on Growth of Barnyardgrass (Echinochloa Crus-Galli), Horse Purslane (Trianthema portulacastrum), Junglerice (Echinochloa colona), and Rice. Weed Sci. 2013, 61, 244–248. [Google Scholar] [CrossRef]
- Formentin, E.; Sudiro, C.; Perin, G.; Riccadonna, S.; Barizza, E.; Baldoni, E.; Lavezzo, E.; Stevanato, P.; Sacchi, G.A.; Fontana, P.; et al. Transcriptome and Cell Physiological Analyses in Different Rice Cultivars Provide New Insights into Adaptive and Salinity Stress Responses. Front. Plant Sci. 2018, 9, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrestha, A.; Desouza, L.L.; Yang, P.; Sosnoskie, L.; Hanson, B.D. Differential Tolerance of Glyphosate-Susceptible and Glyphosate-Resistant Biotypes of Junglerice (Echinochloa colona) to Environments during Germination, Growth, and Intraspecific Competition. Weed Sci. 2018, 66, 340–346. [Google Scholar] [CrossRef]
- Serra, F.; Fogliatto, S.; Vidotto, F. Effect of Salinity on Echinochloa Crus-Galli Germination as Affected by Herbicide Resistance. Ital. J. Agron. 2018, 13. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Active Ingredient | Trade Name | Recommended Rate (g·ai·ha−1) | Application Rate a (g·ai·ha−1) |
|---|---|---|---|
| Imazapic + Imazapyr | KifixTM | 24.5 + 73.5 | 3.06 + 9.19 |
| Imazethapyr | NewpathTM | 211 | 26.37 |
| Log-Logistic Regression Estimates a | |||||
|---|---|---|---|---|---|
| Treatments b | B | D | ED50 | p-Value c | SI d |
| Imazapic | (g·ai·ha−1) | ||||
| G0∙0 mM | −3.96 (0.61) | 101.47 (2.18) | 6.92 (0.23) | <0.05 | - |
| G0∙120 mM | −3.63 (0.39) | 100.78 (1.88) | 4.49 (0.16) | <0.05 | 0.65 |
| G1∙0 mM | −4.06 (0.92) | 101.24 (3.21) | 6.92 (0.34) | <0.05 | - |
| G1∙120 mM | −2.74 (0.46) | 99.36 (3.41) | 5.22 (0.35) | <0.05 | 0.75 |
| G2∙0 mM | −4.06 (0.61) | 101.24 (2.14) | 6.92 (0.22) | <0.05 | - |
| G2∙120 mM | −3.01 (0.35) | 102.62 (2.19) | 6.43 (0.24) | <0.05 | 0.93 |
| Imazethapyr | (g·ai·ha−1) | ||||
| G0∙0 mM | −2.21 (0.25) | 103.41 (3.41) | 61.41 (3.90) | <0.05 | - |
| G0∙120 mM | −1.78 (0.18) | 104.71 (3.03) | 37.25 (2.58) | <0.05 | 0.61 |
| G1∙0 mM | −2.53 (0.21) | 102.72 (2.11) | 61.09 (2.30) | <0.05 | - |
| G1∙120 mM | −2.15 (0.16) | 103.64 (1.92) | 43.03 (1.73) | <0.05 | 0.70 |
| G2∙0 mM | −2.62 (0.27) | 102.74 (2.44) | 61.35 (2.62) | <0.05 | - |
| G2∙120 mM | −2.20 (0.21) | 104.37 (2.47) | 53.38 (2.50) | <0.05 | 0.87 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benedetti, L.; Viana, V.E.; Carvalho-Moore, P.; Gehrke, V.R.; Souza, G.M.; Camargo, E.R.; de Avila, L.A.; Roma-Burgos, N. Recurrent Selection with Low Herbicide Rates and Salt Stress Decrease Sensitivity of Echinochloa colona to Imidazolinone. Agriculture 2021, 11, 187. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11030187
Benedetti L, Viana VE, Carvalho-Moore P, Gehrke VR, Souza GM, Camargo ER, de Avila LA, Roma-Burgos N. Recurrent Selection with Low Herbicide Rates and Salt Stress Decrease Sensitivity of Echinochloa colona to Imidazolinone. Agriculture. 2021; 11(3):187. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11030187
Chicago/Turabian StyleBenedetti, Lariza, Vívian Ebeling Viana, Pâmela Carvalho-Moore, Vinicios Rafael Gehrke, Gustavo Maia Souza, Edinalvo Rabaioli Camargo, Luis Antonio de Avila, and Nilda Roma-Burgos. 2021. "Recurrent Selection with Low Herbicide Rates and Salt Stress Decrease Sensitivity of Echinochloa colona to Imidazolinone" Agriculture 11, no. 3: 187. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11030187