
Responses of Ammonia-Oxidizing Microorganisms to Intercropping Systems in Different Seasons

1
Department of Horticulture, Northeast Agricultural University, Harbin 150030, China
2
Key Laboratory of Cold Area Vegetable Biology, Northeast Agricultural University, Harbin 150030, China
3
Ministry of Agriculture Key Laboratory of Biology and Germplasm Enhancement of Horticulture Crops in Northeast China, Northeast Agricultural University, Harbin 150030, China
*
Author to whom correspondence should be addressed.
Agriculture 2021, 11(3), 195; https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11030195
Submission received: 17 January 2021
/
Revised: 23 February 2021
/
Accepted: 23 February 2021
/
Published: 27 February 2021
Abstract
:Intercropping plays an essential role in agricultural production, impacting the soil’s physical and chemical properties and microbial communities. However, the responses of ammonia-oxidizing microorganisms in the continuous-cropping soil to different intercropping systems in different growing seasons are still insufficiently studied. Here, we investigated the effects of seven intercropping systems (alfalfa (Medicago sativa L.)/cucumber, trifolium (Trifolium repens L.)/cucumber, wheat (Triticum aestivum L.)/cucumber, rye (Secale cereale L.)/cucumber, chrysanthemum (Chrysanthemum coronrium L.)/cucumber, rape (Brassica campestris L.)/cucumber, mustard (Brassica juncea L.)/cucumber) on soil physical and chemical properties, potential nitrification rate (PNR), soil ammonia-oxidizing archaea (AOA), and ammonia-oxidizing bacteria (AOB) communities in the greenhouse in spring and autumn. The results showed that, compared with cucumber monoculture, intercropping increased the soil -N and -N. The chrysanthemum–cucumber, rape–cucumber, and mustard–cucumber treatments increased soil PNR. Intercropping increased the AOA and AOB abundances in two seasons, especially in rape–cucumber, wheat–cucumber, chrysanthemum–cucumber, and trifolium–cucumber treatments. The ratio of AOA and AOB decreased with seasonal variation. The wheat–cucumber and rape–cucumber treatments increased soil AOA community diversity. Seasonal variation had a significant effect on the relative abundance of the AOB community. Nonmetric multidimensional scaling analysis showed that the AOA and AOB community structures were obviously different from spring to autumn. Redundancy analysis showed that the AOA community was significantly regulated by moisture, –N, and available potassium (AK), while the AOB community was significantly regulated by moisture, available phosphorus (AP), AK, -N, and pH. Network analysis showed that the co-occurrence relationship and complexity of AOA and AOB communities were different in two growing seasons. The AOB community may play a critical role in ammonia oxidation in autumn. Taken together, intercropping improved soil physicochemical state, increased soil PNR and significantly altered soil AOA and AOB communities. Seasonal variation significantly altered the AOA and AOB communities’ structure and interaction between them. The effect of seasonal variation on AOA and AOB communities was greater than intercropping.
1. Introduction
Intercropping is a commonly used agricultural practice, which has been proven to have significant advantages in yield increase, disease control, and resource utilization [1,2]. Simultaneously, intercropping changes the plant–microbe or microbe–microbe interaction, thereby affecting the nitrogen transformation in the soil [3]. Nitrification is a crucial step in the nitrogen cycle and plant nutrition. It is a biology-mediated process and plays a crucial role in nitrogen form transformation and the nitrogen cycle. It directly affects environmental quality and nitrogen utilization efficiency [4,5]. Ammonia oxidation and nitrite oxidation are two critical steps of nitrification. Ammonia oxidation is the rate-limiting step of nitrification, which is mainly completed with the participation of soil ammonia-oxidizing archaea (AOA) and ammonia-oxidizing bacteria (AOB) [6,7]. Ammonia-oxidizing microorganisms can respond to changes in the soil micro-habitat of the intercropping system. For example, maize–faba bean intercropping increased the abundances of total archaea and the AOA and AOB community diversities and structures were affected by plant growth period and competition among plant species, which changed the soil nitrification rate [8]. Pepper–alfalfa intercropping and pepper–maize intercropping stimulated the activity of AOB in the topsoil, while AOA decreased with planting along with the soil profile and this deep and shallow root intercropping system could absorb nitrogen more effectively [9]. Previous studies have also confirmed that seasonal variation can also change the abundance and structure of ammonia-oxidizing microorganisms [10,11]. However, the effects of intercropping on AOA and AOB communities in the continuous-cropping soil are poorly understood in different seasons.
Due to the increase in plant diversity, intercropping often affects soil properties. Many studies have shown that ammonia-oxidizing microorganisms were affected by many soil environmental factors [12,13]. For example, the AOA and AOB community structures were regulated by the soil moisture, pH, and TN in wheat-maize rotation soil [14]. In low-pH and ammonia nitrogen soil environments, AOA were most represented and functionally dominant, while in neutral, alkaline or nitrogen-rich soil, AOB were dominant in nitrification [15,16]. The soil pH directly affects the effectiveness of -N as a substrate for ammonia oxidation. The soil nitrification rate represents the ability of soil nitrification microorganisms to convert into , so it is also an essential indicator of the abundances of nitrification microorganisms [17,18]. The availability of is also an important factor determining the ecological niche of AOA and AOB [6,19]. AOA are more effective in utilizing low concentrations of produced by organic nitrogen mineralization [20]. In the range of high concentrations, the AOB abundance and contribution to ammonia oxidation are positively correlated with the concentration [21,22]. AOA are more widely distributed than AOB in soil, which means that AOA may contribute more to ammonia oxidation [23,24]. However, some studies have also confirmed that AOA were dominant in abundance, while AOB were dominant in function in intercropping systems [14]. Therefore, the effects of different intercropping systems on the continuous-cropping soil ammonia-oxidizing microorganisms need to be studied in depth.
Cucumber (Cucumis sativus L.) is a nitrogen demanding crop. The availability of nitrogen in the soil is critical for the growth of cucumber. Because continuous cropping obstacle is common in facilities, the availability of soil nutrients will be reduced, which is not conducive to the growth of crops. However, intercropping can alleviate the obstacle of continuous cropping, increase the abundances of microorganisms in the soil, improve the community structure, accelerate the transformation of nutrients in the soil, and improve the quality of crop nutrient absorption [25,26,27]. However, the interaction of ammonia-oxidizing microorganisms in the continuous-cropping soil of intercropping system in the greenhouse is still unclear.
In this study, the physical and chemical properties, PNR, and ammonia-oxidizing microbial communities of the continuous-cropping soil were studied in cucumber monoculture and cucumber intercropping with seven crops from the Gramineae, Leguminosae, Cruciferae, and Compositae families in spring and autumn. The abundances, diversities, and composition of soil AOA and AOB communities in monoculture and intercropping systems of cucumber were analyzed by quantitative PCR (qPCR) and Illumina MiSeq sequencing to assess the potential effects of different intercropping systems and planting seasons on soil AOA and AOB communities. Our aim was to determine the effects of seven intercropping systems on soil AOA and AOB in two cropping seasons. We hypothesized that (1) the structure and composition of soil AOA and AOB communities could be changed by intercropping seven crops with cucumber, (2) the difference of physical and chemical factors caused by seasonal variation changed the community structure and coexistence mode of ammonia-oxidizing microorganisms.
2. Materials and Methods
2.1. Experimental Location and Design
The experiment was carried out in a greenhouse at the experimental base of Horticulture of Northeast Agricultural University (45°41′ N, 126°37′ E), Harbin, Heilongjiang Province, China. The soil was a black soil (Mollisol) under continuous cucumber cropping for 3 years with a sandy loam texture, and contained sand, 326 g kg−1; silt, 384 g kg−1; clay, 270 g kg−1; organic matter, 3.51%; pH, 6.64; -N, 13.32 mg kg−1; -N, 253.04 mg kg−1; available potassium (AK), 359.03 mg kg−1; and soil available phosphorus (AP), 277.62 mg kg−1.
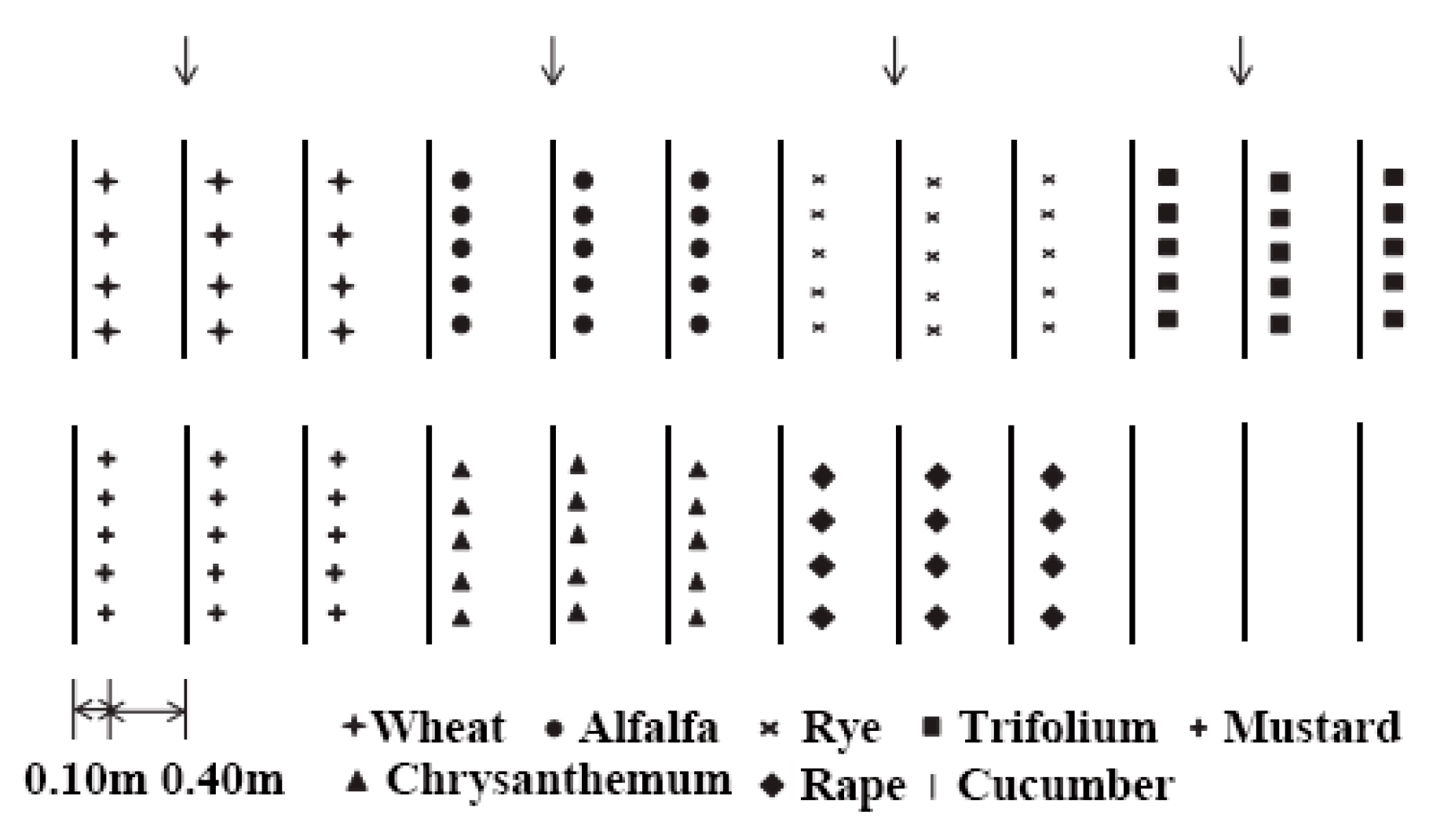
A single-factor random block design was adopted, and each treatment was repeated with three replicates in the greenhouse in spring (April 21–June 30) and autumn (July 29–October 6) in 2015. The size of each plot was 6 × 0.5 m2. Seven crops (alfalfa (A), trifolium (T), wheat (W), rye (Ry), chrysanthemum (C), rape (Ra), and mustard (M)) were intercropped with cucumbers as the experimental group, and a cucumber monoculture (CM) was the control. Each plot consisted of one row of 12 cucumber plants with two protection lines on each side. When cucumbers were planted for 10 days, intercropped seeds were sown approximately 10 cm away from cucumber seedlings (Figure 1). The number of intercrop seedlings for wheat and rye was 30 seedlings, that for trifolium and alfalfa was 40 seedlings, that for rape and mustard was 3 seedlings, and that for chrysanthemum was 5 seedlings. When the intercropping crops reached 20 cm, to avoid an impact on cucumber growth, the intercropping crops were trimmed by 10 cm. Compound NPK fertilizer (16:16:8) was applied to each plot at a rate of 300 kg ha−1.
2.2. Soil Sampling and Analysis
The soil samples were obtained from 24 test districts in the greenhouse on 30 June and 6 October, the collection depth was 0–20 cm, and then they were mixed and sieved through a 2 mm mesh. Afterward, the soil sample was divided into two parts in the laboratory: one was kept at −80 °C for DNA extraction, and the other at −4 °C for chemical analysis. For soil inorganic nitrogen concentration (-N, -N) analysis; each sample was extracted by 2 M KCl, and the soil available phosphorus (AP) and available potassium (AK) were extracted by 0.5 M NaHCO3 (pH = 8.5) and 1 M NH4+OAc (pH = 7.0), respectively. The soil filtrates were measured with a continuous flow analyzer (SAN++, Skalar, Breda, Netherlands). Soil pH was determined by inserting electrodes into a soil–water suspension (1:2.5). The soil moisture content was calculated by drying at 105 °C for 8 h [28].
2.3. Potential Nitrification Rate (PNR)
PNR was analyzed as previously described [29]. Briefly, 5 g soil was put into per conical flask respectively, and 20 mL of phosphate buffer solution (1 mM, pH 7.4) and 0.1 mL of NaClO3 solution (1.5 M) were added. They were gently mixed and sealed with sealing film. The two conical flasks were put onto a shaking table for 5 h (samples). The other sample was stored at −20 °C for 5 h (control). After culturing, the control was dissolved at room temperature, and then 5 mL of 2M KCl solution was added to the sample and control and filtered immediately. Next, 5 mL of filtrate, 3 mL of NH4Cl buffer solution, and 2 mL of color-developing solution were added to the test tube and mixed evenly. After being placed at room temperature for 15 min, the color was determined spectrophotometrically at 520 nm.
2.4. DNA Extraction and Quantitative PCR (qPCR)
According to the manufacturer’s instructions, the total DNA of the soil was extracted from 0.5 g soil using the MoBioPowerSoilTM DNA Isolation Kit (Mobio Laboratories Inc. Carlsbad, CA, USA). The DNA concentration and purity were determined with a NanoDrop 2000 Spectrophotometer (Thermo Scientific, Waltham, MA, USA).
The PCR amplification was performed on an IQ5 real-time PCR system (Bio-Rad Lab, Hercules, CA, USA). The reaction system of real-time PCR was 20 μL, including 10 μL 2 × SYBR® Green qPCR Master Mix (Applied Biosystems, Foster City, CA, USA), 1 μL each of the front and back primers, 2 μL DNA extract (2 ng·μL−1), and 6 μL ddH2O. The primers for AOA amoA gene amplification were Arch-amoAF (5′-STAATGGTCTGGCTTAGACG-3′) and Arch-amoAR (5′-GCGGCCATCCATCTGTATGT-3′) [30]. The primers for AOB amoA gene amplification were amoA1F (5′-GGGGTTTCTACTGGTGGT-3′) and amoA2R (5′-CCCCTCKGSAAAGCCTTCTTC-3′) [31]. The PCR reaction system was as follows: 95 °C for 5 min, 40 amplification cycles of 95 °C for 10 s, 55 °C for 20 s, 72 °C for 30 s. The standard product was transformed into the DH5 strain by connecting p MD19-T (TaKaRa, Dalian, China) with purified AOA-amoA and AOB-amoA genes, and the recombinant plasmid was extracted. The plasmid was continuously diluted by 6 orders of magnitude as the standard curve, the AOA and AOB amplification efficiency and R2 were 94.7%, 0.9998 and 95.4%, 0.9996, respectively.
2.5. Illumina MiSeq Sequencing and Data Processing
The PCR amplification primers were the same as qPCR, and the Barcode sequence was added at the front of the primer to distinguish samples during amplification. The amplification was carried out in 25 uL volume, including 2 μL DNA extract, 1 μL each of the front and back primers, 5 μL 5 × PCR buffer, 2 μL (2.5 mmol·L−1) dNTP, 5 μL 5 × Q5High GC Enhancer buffer, 0.25 μL (0.02U·μL−1) Q5High-Fidelity DNA polymerase (NEB) and 8.75 μL ddH2O. The reaction conditions were as follows: 98 °C for 5 min, 35 amplification cycles of 98 °C for 30 s, 55 °C for 30 s, 72 °C for 45 s. The amplified PCR products were detected by 2% agarose gel electrophoresis, and AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, Union City, CA, USA) were used to recover the products. The PCR products were analyzed by QuantiFluor™-ST (Promega, Madison, WI, USA) blue fluorescence quantitative system, and the samples were mixed in proportion. Then, the high-throughput amplicon sequencing was carried out on Illumina MiSeq platform (2 × 300) at Majorbio Bio-Pharm Technology Co., Ltd. (Shanghai, China), each treatment was done in triplicate.
The raw sequence files were quality filtered and processed using QIIME (Version 1.8.0). The operational taxonomic units (OTUs) were clustered at 97% sequence similarity using UPARSE (Version 7.1) [32], and the chimeric sequences were detected and removed using UCHIME [33]. The archaeal and bacterial OTUs were classified by using ribosomal database project (RDP) classifier against the functional genomics resource (FGR) functional gene database [34]. To preclude bias due to several sequencing depths, all samples were subsequently subsampled based on the minimum number of soil microbial sequencing depths of this study. All sequences have been deposited in the NCBI SRA database (AOA: SRP 160125; AOB: SRP160403).
2.6. Statistical Analysis
The alpha diversity indices of the samples were generated by QIIME [35]. For beta diversity, the AOA and AOB community structures were analyzed by nonmetric multidimensional scaling (NMDS) based on the Bray–Curtis distance dissimilarity, and ANOSIM and Adonis were used to compare the microbial community differences of two growing seasons with the Bray–Curtis distance and 999 permutations, and the soil physical and chemical factors less than 10 were screened by variance inflation factor (VIF) for redundancy analysis (RDA), and they were performed using the “vegan” package in “R” (Version 3.3.1, R Foundation for Statistical Computing, Vienna, Austria). All data were checked for normality and homogeneity of variance. Network analysis based on spearman’s rank analysis was performed using the 50 most abundant OTUs of AOA and AOB communities. Based on strong (ρ > 0.8) and significant correlations (p < 0.01), the symbiotic patterns of ammonia oxidizing microorganisms in different seasons were visualized by Gephi (Version 0.9.2). The correlation and analysis of variance were performed at 0.05 probability level in SPSS (Version 17.0).
3. Results
3.1. Soil Physical and Chemical Properties and PNR
The physical and chemical properties of the different treatments were illustrated in Table 1. Compared with cucumber monoculture, all intercropping treatments significantly reduced the soil AP in spring (p < 0.05) but not in autumn. The rye–cucumber treatment significantly increased the soil -N in spring (p < 0.05), and the alfalfa–cucumber and chrysanthemum–cucumber treatments significantly increased the soil -N in autumn (p < 0.05). There was no significant difference in the soil moisture, pH, or AK between intercropping and monoculture.
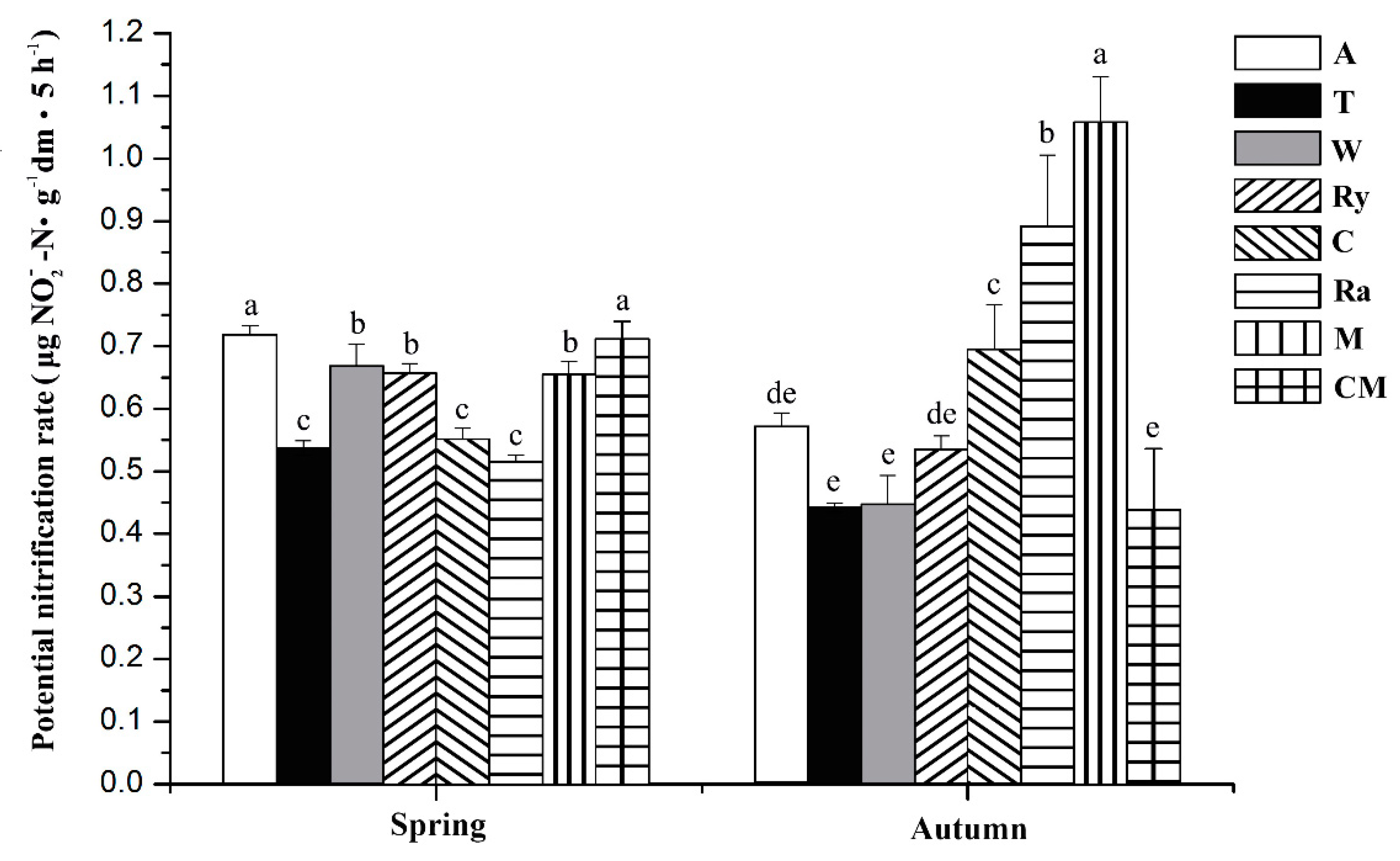
The PNR of different treatments was shown in Figure 2. Except for the PNR of the alfalfa–cucumber treatment, other intercropping treatments were significantly lower than the cucumber monoculture in spring (p < 0.05). The PNR of the mustard–cucumber, rape–cucumber, and chrysanthemum–cucumber treatments were significantly higher than the cucumber monoculture in autumn (p < 0.05).
3.2. Soil AOA and AOB Abundances
The AOA abundance of the wheat–cucumber, chrysanthemum–cucumber, rape–cucumber, and mustard–cucumber treatments were significantly higher than the cucumber monoculture (p < 0.05), but that of the rye–cucumber treatment was significantly lower compared to the cucumber monoculture in spring (p < 0.05). The AOA abundance of the alfalfa–cucumber, trifolium–cucumber, and wheat–cucumber treatments were significantly higher than the cucumber monoculture in autumn (p < 0.05). The AOA abundance of the wheat–cucumber treatment was the highest in both growing seasons (Figure 3a).
The AOB abundance of the chrysanthemum–cucumber, rye–cucumber, and alfalfa–cucumber treatments were significantly higher than the cucumber monoculture in spring (p < 0.05). The AOB abundance of the trifolium–cucumber treatment was significantly higher than the cucumber monoculture (p < 0.05), while that of the rye–cucumber, chrysanthemum–cucumber, rape–cucumber, and mustard–cucumber treatments were significantly lower than the cucumber monoculture in autumn (p < 0.05) (Figure 3b).
In addition, the soil AOA abundance of all treatments was significantly higher than the AOB in both growing seasons (p < 0.05), and the ratio AOA/AOB was 9.16 to 58.35 in spring and 9.63 to 31.50 in autumn (Figure 3c,d).
The correlation analysis showed that the AOA abundance was negatively correlated to the moisture and positively correlated to the -N (p < 0.05), and the AOB abundance was negatively correlated to the pH and positively correlated to the -N in spring (p < 0.01). The AOA abundance was negatively correlated to the -N, AP, and AK (p < 0.05), and the AOB abundance was positively correlated to the -N in autumn (p < 0.05) (Table 2).
3.3. Soil AOA and AOB Community Alpha and Beta Diversities
The alpha-diversity indices of AOA and AOB communities were observed for Table 3. The sequence numbers of amoA genes in AOA and AOB ranged from 11,467 to 24,485, and the coverage ranged from 99.90% to 99.96%. At the 97% similarity level, AOA and AOB sequences can be divided into 50 to 57 and 51 to 57 OTUs. The average fragment lengths of AOA and AOB were 272 bp and 473 bp, respectively. The AOA Simpson index values of the wheat–cucumber, rye–cucumber, and rape–cucumber treatments were significantly higher than the cucumber monoculture, and the AOB Simpson index value of the rape–cucumber treatment was significantly higher than the cucumber monoculture in spring (p < 0.05). Moreover, the AOA Simpson index values of the wheat–cucumber, chrysanthemum–cucumber, and mustard–cucumber treatments were significantly higher than the cucumber monoculture in autumn (p < 0.05). The AOA Chao 1 index values of the alfalfa–cucumber, trifolium–cucumber, wheat–cucumber, and chrysanthemum–cucumber treatments were significantly higher than the cucumber monoculture in spring (p < 0.05). The AOA Chao 1 index values of all treatments were significantly higher than the cucumber monoculture except for the wheat–cucumber treatment. The AOB Chao 1 index value of the alfalfa–cucumber treatment was significantly higher than the cucumber monoculture in autumn (p < 0.05).
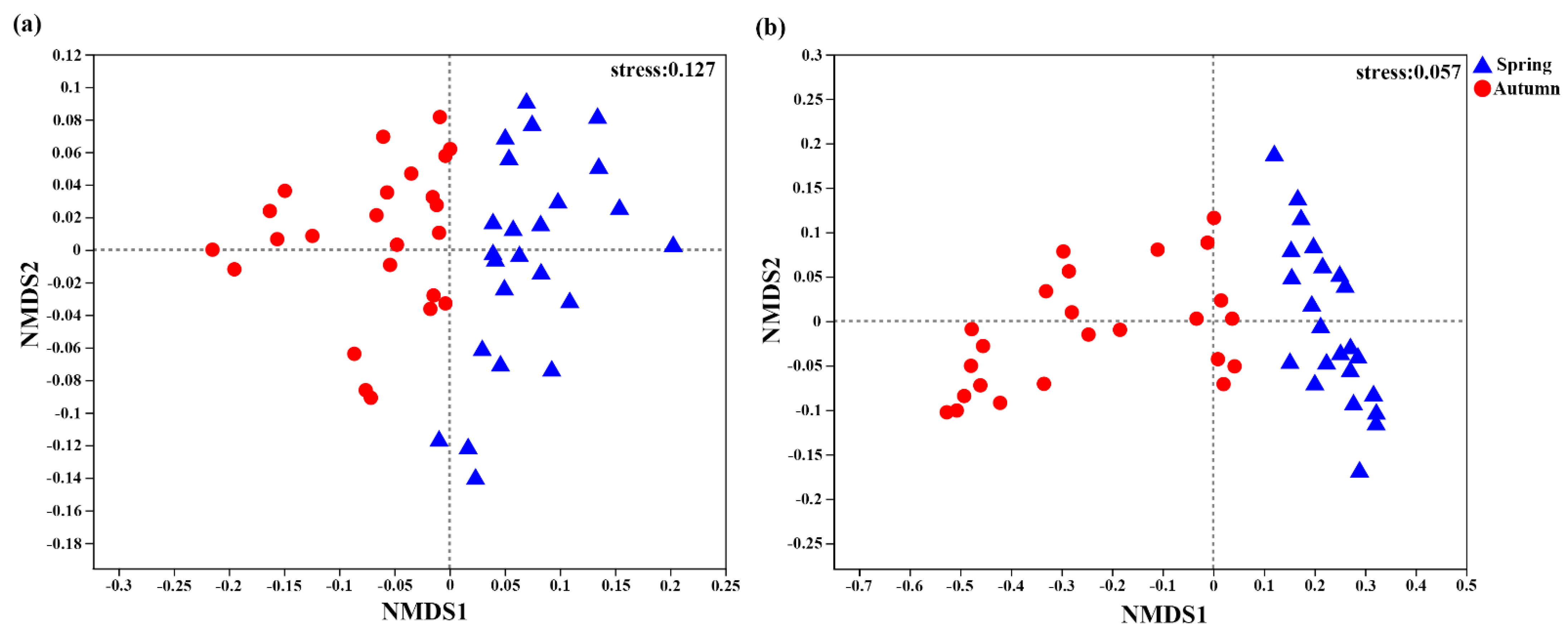
The AOA and AOB community structures from spring to autumn were analyzed by NMDS (Figure 4). Non-parametric multivariate statistical analysis showed that the community structures of different treatments were similar (date not shown). However, the soil AOA (ANOSIM, R = 0.1778, p = 0.001; Adonis, R2= 0.1418, p = 0.001) and AOB (ANOSIM, R = 0.6588, p = 0.001; Adonis, R2= 0.5151, p = 0.001) community structures of all samples were different from spring to autumn.
3.4. Soil AOA and AOB Community Composition
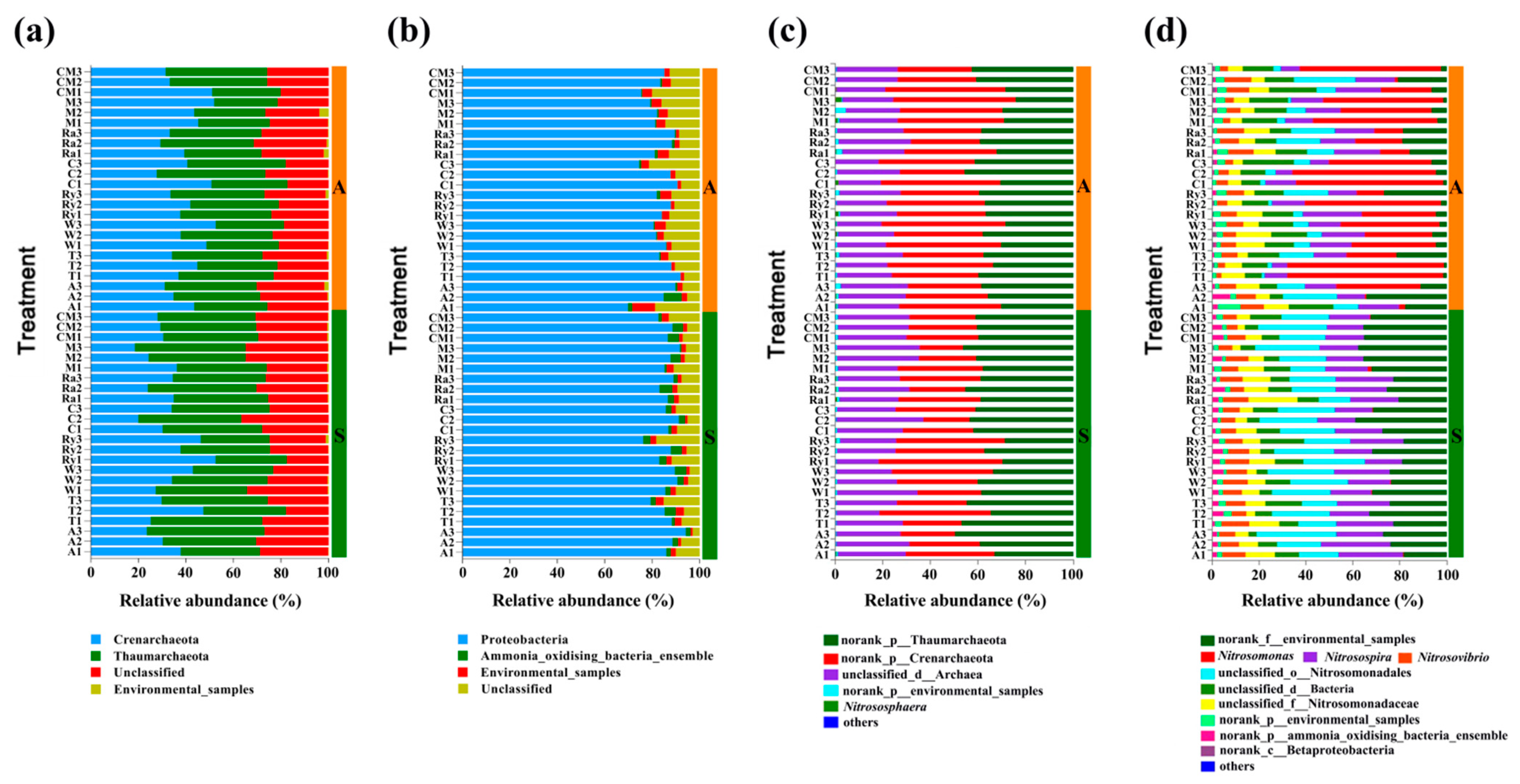
The predominant phyla of the AOA community were Crenarchaeota and Thaumarchaeota, occupying more than 75% of the total sequences (Figure 5a). The genus Nitrososphaera (AOA) was only represented 0.31% and 0.61% in two growing seasons. (Figure 5c). The predominant phyla of the AOB community were Proteobacteria and a group of unidentified bacteria found in the environment, occupying more than 95% of the total sequences in spring, while Proteobacteria and, the group of unidentified bacteria found in the environment, and a group of unidentified bacteria occupied more than 90% in autumn (Figure 5b). The predominant genera of the AOB community were the group of bacteria from the norank in the environment, Nitrosospira, group of unidentified from Nitrosomonadales, and Nitrosovibrio, and the known genera accounted for 8.1% (Nitrosovibrio) and 19.8% (Nitrosospira) of the total sequences in spring. The predominant genera of the AOB community were Nitrosomonas, group of bacteria from the norank in the environment, Nitrosospira and, Nitrosovibrio, and the known genera accounted for 6.5% (Nitrosovibrio), 14.2% (Nitrosospira) and, 35.0% (Nitrosomonas) of the total sequences in autumn. Interestingly, Nitrosomonas was the rarest genus in all treatments in spring, while Nitrosomonas increased significantly in autumn. In contrast, the group of bacteria from the norank in the environment decreased significantly from spring to autumn. This indicated that seasonal variation had a significant effect on Nitrosomonas and group of bacteria from the norank in the environment (Figure 5d).
3.5. Relationships between Soil AOA and AOB Communities and Soil Physical and Chemical Properties
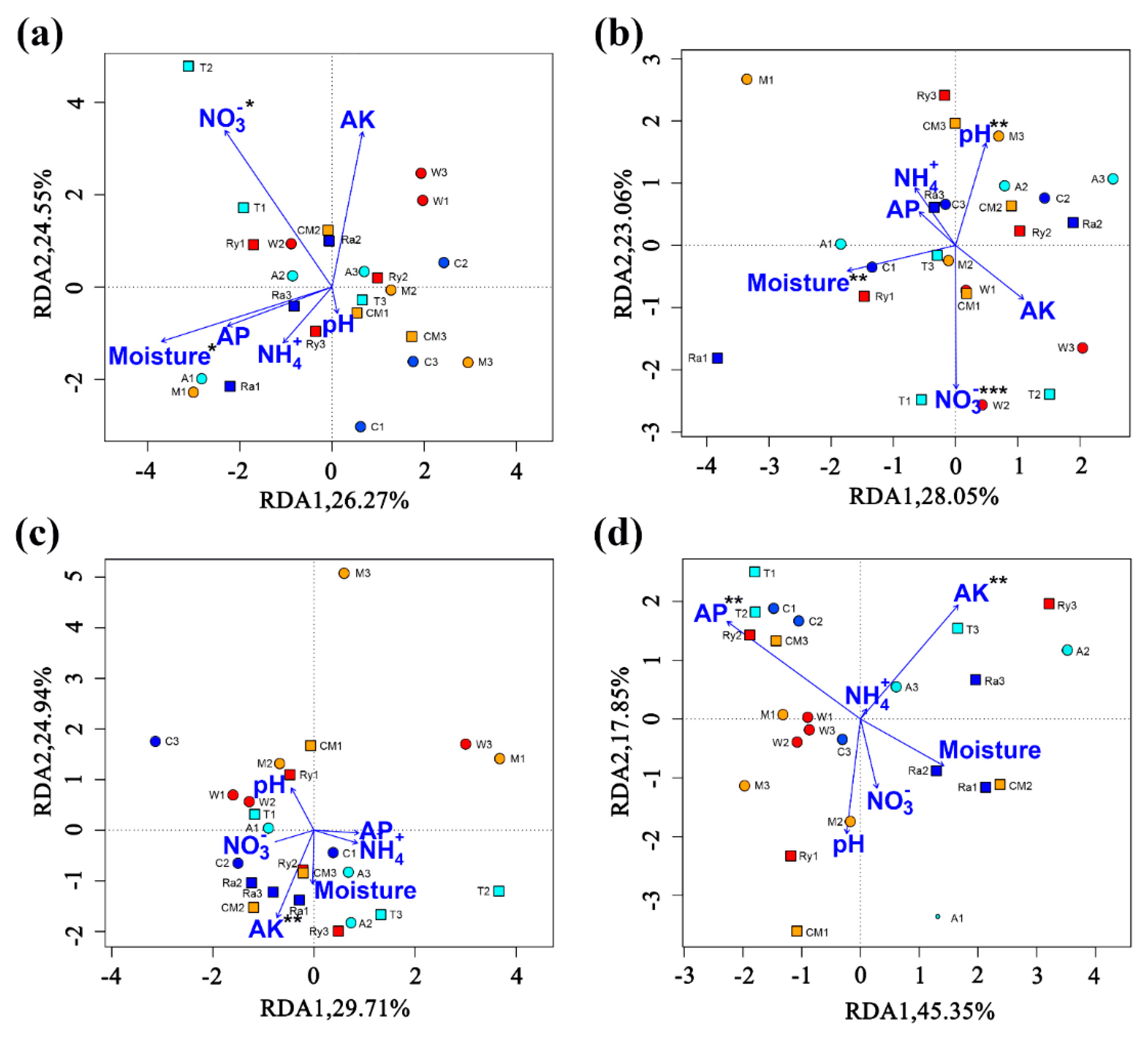
The RDA analysis showed that the soil AOA community was significantly affected by moisture (p < 0.05) and -N (p < 0.05), and the soil AOB community was significantly affected by moisture (p < 0.01), pH (p < 0.01), and -N (p <0.01) in spring (Figure 6a,b). The soil AOA community was significantly affected by AK (p < 0.01), and the soil AOB community was significantly affected by soil AP (p <0.01) and AK (p < 0.01) in autumn (Figure 6c,d).
3.6. Co-Occurrence Network Analysis
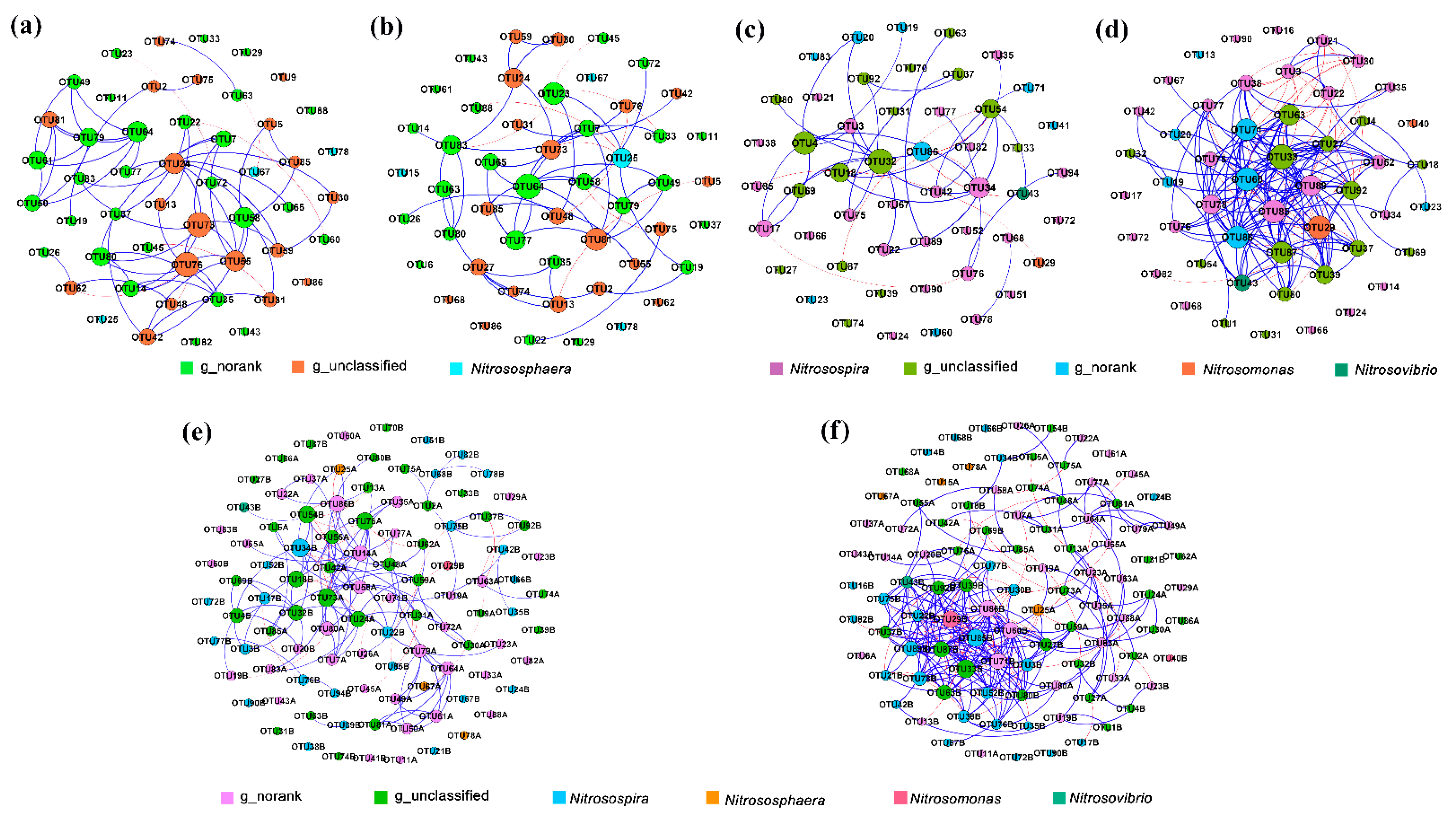
We used the network graph to compare the co-occurrence relationship and community complexity of ammonia-oxidizing bacteria communities between spring and autumn (Figure 7). The network topological properties of AOA and AOB were shown in Table 4. The results showed that the modularity index of each process was greater than 0.4, indicating that the co-occurrence network of each process was a typical modular structure. There were significant differences in the size of the AOA and AOB community networks in different seasons. The total number of edges, positive correlation edges, and graph density of the AOA network in spring were significantly higher than those in autumn (Figure 7a,b), In comparison, the total number of edges, positive, and negative correlation edges and graph density of the AOB network in autumn were significantly higher than those in spring (Figure 7c,d). The analysis of the network graph between AOA and AOB showed that the total number of edges, positive correlation edges, and graph density of AOA and AOB in autumn were significantly higher than those in spring. The relationship between AOA and AOB was relatively simple to analyze in spring, but in autumn, the relationship between AOA and AOB was more complex, the AOB community played a leading role in the whole network (Figure 7e,f). These results indicated that seasonal variation regulated the co-occurrence relationship, correlation, and complexity of AOA and AOB, and changed their complex interactions.
4. Discussion
Long-term continuous cropping results in excessive soil nutrient consumption and the deterioration of soil physical and chemical properties, while reasonable intercropping can effectively improve soil physical and chemical properties and promote nutrient recycling and transformation in agricultural production [36,37]. Previous studies have shown that intercropping system can affect soil physical and chemical properties [38,39]. Our results showed that compared with the cucumber monoculture, the rye–cucumber treatment significantly increased the soil -N in spring, which could be due to the increase of soil nitrogen content by pruning rye stubble [40]. The alfalfa–cucumber and chrysanthemum–cucumber treatment -N contents were significantly increased in autumn, which could be due to the solid water in autumn that slowed moisture evaporation and reduced the soil -N leaching degree. On the other hand, soil nitrifying bacteria converted the unused -N into -N and stored it in the soil under the appropriate soil temperature and pH rang, causing some intercropping systems to increase in -N contents in autumn [41]. In the intercropping systems, the AP content significantly decreased, which is consistent with previous research results [27], this may be due to the increase of plant diversity in intercropping, activate the difficult soluble phosphorus in soil, beneficial to crops on the absorption of phosphorus, thus resulting in a decrease of soil in AP [8], but the intercropping treatments did not significantly change the water, pH and AK (Table 1), this may be caused by the experiment of short-term [42,43].
The ammonia-oxidizing process is mainly conducted by ammonia-oxidizing microorganisms such as AOA and AOB, and PNR is one of the critical indicators of soil nitrification activity. We found that most intercropping treatments significantly reduced soil PNR in spring (Figure 2), which may be due to interspecific competition and synergy. Some studies have found that not all intercropping systems can improve soil ammonia-oxidizing microorganisms. Previous studies have found that compared with monoculture, intercropping of broad bean and maize significantly reduced soil AOA and AOB abundances and rhizosphere microbial abundances. The change of nitrification rate may be related to the competition among species, which may affect the nitrification rate of soil [10,28]. The PNR values of the mustard–cucumber, rape–cucumber, and chrysanthemum–cucumber intercropping treatments were significantly higher than the cucumber monoculture in autumn. This could be justified by the fact that most intercropping treatments have better water retention in autumn, and the soil moisture content of the cucumber monoculture was lower than the intercropping treatments. A previous study found that the soil moisture content was significantly positively correlated with the PNR, and a decreased moisture content can lead to a decrease of PNR [1].
Previous studies have shown that intercropping system can significantly change the abundances of ammonia-oxidizing microorganisms in soil [3,8,9]. Those findings are consistent with the results of this study. This may be related to the improvement of the soil environment by increasing plant diversity [1,2]. In addition, AOB were dominant players in the ammonia-oxidizing process until AOA were discovered and proven to be more abundant and diverse in population compared to AOB in some soils, and it was also found that AOA could play a leading role in nitrification [44,45]. According to the qPCR results, the AOA abundance was significantly higher than AOB in all treatments in the two growing seasons, which was consistent with most findings in agricultural soils [46]. In the comparison of the same treatment, the ratio of AOA to AOB in spring was 9.16 and 58.35, while in autumn, it was 9.63 and 31.50. With the change of seasons, the rate of AOA:AOB showed a downward trend, indicating that AOB rather than AOA may play a leading role in ammonia oxidation [47]. We also found that different growing seasons had a significant effect on AOA and AOB abundances. From spring to autumn, AOB abundance increased in all treatments (Figure 3). The results of this study are consistent with other results, these studies indicate that seasonal climate change and increased plant diversity caused by intercropping may be the reasons for the change of soil AOA and AOB abundances [48,49,50], indicating that different intercropping patterns and seasonal variation could cause changes in AOA and AOB abundances. However, a study has shown that seasonal heterogeneity had a significant impact on the AOA abundance of the rhizosphere of Phragmites communis but had no significant effect on the AOB abundance [51], suggesting that nitrifying microbial functional genes have different responses to different ecosystems and land management practices.
A variety of environmental factors could influence aerobic ammonia-oxidizing bacteria through complex mechanisms of action, among which pH, moisture, -N, and -N have significant effects on the abundances of AOA and AOB [7,15]. In this study, the AOA and AOB abundances were closely related to soil properties. For example, the diffusion capacity of O2 decreased with an increase in the soil moisture, which significantly affected the microbial activities of AOA [52]. This may be the cause of the decrease in AOA abundance of in soil. Similarly, the factor analysis driving the variation in soil AOA and AOB abundances showed that the moisture and AK were negatively correlated to the AOA abundance, and the pH was negatively correlated to the AOB abundance in spring (Table 2). The AP was negatively correlation to the AOA abundance in autumn. Studies have shown that the favorable pH for the growth and proliferation of AOB is 7.0-7.40 in soil, while nitrification is inhibited in soils at low pH [53,54]. In our study, the pH in spring ranged from 7.10–7.32, which was higher than that in autumn, ranging from 6.75–6.91. In addition, previous studies also showed that AOA generally dominate nitrification in acidic soils, while AOB dominate nitrification in alkaline soils [55,56], which was contrary to our results. Our results showed that the abundance of AOB decreased with the increase of pH, which may be because a suitable alkaline soil is conducive to the survival of AOB, and excessively high pH reduced AOB activity [57]. Previously, it was shown that only AOB was found to be involved in 13CO2 fixation and ammonia-oxidizing processes in the alkaline tidal soils of Fengqiu city in China, and AOB was the dominant microorganisms in the soil nitrification. [58]. Therefore, nitrification may have been dominated by AOB in our study since pH was the key environmental factor regulating the amount of AOB in spring. The reason why AOB cannot grow in a low pH environment may be the low affinity of AOB to substrates, their inability to adapt to an oligotrophic environment and their inability to metabolize in an environment with a pH lower than 6.50 [59]. However, recent studies have found that some strains and species of AOB have also been isolated from acidic agricultural soils with pH values between 5.0–7.50, and the Nitrosospira cluster 3a.2 of AOB isolated from acidic woodland soil could be self-oxidized with ammonia [60,61]. These results showed that the contribution of AOB to ammonia-oxidation in acidic soils could not be ignored and further research to understand its unknown function. AOB could survive well in grass soils with high-dose substrates, while AOA usually survive in soils with low substrate concentrations, proving that AOA are more suitable for growth under oligotrophic conditions and that high substrate concentrations could inhibit their abundance [62]. This could be why the AOA abundance was significantly negatively correlated with the soil AP and AK nutrients in our study.
The soil microbial diversity and ecosystem stability are closely related to nutrient transformation, and intercropping can affect various agro–ecosystem services by improving the soil microecology [36]. Our study showed that the results showed that the diversity index of AOA and AOB was increased by intercropping systems (Table 3), indicating that intercropping improved the diversity of microorganisms involved in the ammonia oxidation reaction in continuous-cropping soil, which may contribute to the enhancement of ammonia oxidation in agro–ecosystems. NMDS and Non-parametric multivariate statistical analysis showed that the three replicates of AOA and AOB were closely distributed, and there was no apparent separation between the treatments, and the community composition of different treatments was similar. However, the AOA and AOB community structures changed significantly from spring to autumn (Figure 4). Studies have shown that changes in seasons had significant effects on the community structures and activity of AOA and AOB in different soil types [10,63]. These results proved that AOA and AOB community structures were susceptible to seasonal variation, and the effect of seasonal variation was greater than intercropping.
The wheat–cucumber and alfalfa–cucumber treatments significantly increased the relative abundance of Nitrososphaera and Nitrosospira for AOA and AOB at the genus level in spring, respectively. From a seasonal perspective, the relative abundance of Nitrosospira and Nitrosovibrio decreased significantly in most treatments from spring to autumn. Interestingly, the most abundant Nitrosomonas in all systems increased significantly, but the group of bacteria from the norank in the environment decreased significantly at the genus level when the season changed from spring to autumn (Figure 5). This phenomenon may have been caused by the large temperature difference between the two growing seasons in northeast China, which affected the availability of soil substrates and changed the activity of Nitrosomonas and other bacteria; therefore, the AOA and AOB community composition was changed in the two growing seasons [64]. Our results showed that the change in the AOA community was relatively stable compared with the cucumber monoculture, the composition of the AOB community was mainly affected by seasonal variation, and this also further proves that the ammonia-oxidation process of the soil might have been dominated by the AOB community.
Moisture, AP, AK, -N, and pH were related to alterations in the nitrifying bacterial community structure (Figure 6). The AOA community was mainly affected by the moisture, -N, and AK. The AOB community was affected primarily by the moisture, AP, AK, -N, and pH, which also indicated that AOA were more tolerant to different environmental factors compared to AOB. A previous study has shown that moisture was the main factor affecting AOA and AOB community structures, which is consistent with the results of our study [65], however -N did not cause changes in community structure, which is different from our results [14]. The community structure of AOB was regulated by soil pH. Previous studies have shown that the soil pH is a crucial factor influencing changes in the AOB community structure [14,66]. Notably, the correlation between -N and the AOB community was not significant, which is contrary to the results of our study, which indicated that the ammonia concentration had a significant effect on the AOB community [53]. Soil AOA and AOB communities were correlated with the soil nutrients, which may have been caused by the fact that AP and AK changed the proportions of the nutrient matrix in the soil, thus affecting the flora structure of ammonia-oxidizing bacteria, which further indicates that AOA and AOB had a strong dependence on the soil nutrients [67,68].
The complex interactions between species are one of the crucial indicators of community biodiversity [69]. The significant changes observed in the network of two seasons in this study showed that the intraspecific and interspecific interactions of AOA and AOB varied with seasons. The intraspecies interactions of AOA and AOB communities were stronger in spring than in autumn. The results of the interspecific co-occurrence network showed that the interaction (edges) in autumn was significantly stronger than that in spring, indicating that the interaction between AOA and AOB communities was stronger in autumn, and the whole network was dominated by the AOB community (Figure 7, Table 4). In spring and autumn, the difference of soil ammonia-oxidizing microorganisms and their interaction intensity may be due to the lower soil temperature in spring and the lower activity of ammonia-oxidizing microorganisms in the soil. The relationship between the microbial communities was relatively simple, and the interaction between the microorganisms was less. However, the soil temperature in autumn was higher, and the activity of ammonia-oxidizing microorganisms was higher, which leads to more complex interaction between microbial communities. The results of this study on the interaction between communities caused by seasonal variation are similar to previous studies [70,71]. This result also further proved that with the seasonal variation, the ammonia oxidation in the soil might be dominated by the AOB community.
5. Conclusions
In summary, this study showed that intercropping significantly changed the soil -N and -N contents, and most intercropping systems significantly increased soil PNR, AOA, and AOB abundances. The AOA abundance in all treatments was significantly higher than AOB, and the ratio between them decreased as the seasonal variation. The wheat–cucumber and rape–cucumber treatments increased AOA community diversity. Seasonal variation was an important factor affecting the relative abundance of AOB. Seasonal variation had greater influence on the community structures of AOA and AOB than intercropping. Moisture, pH, -N, AP, and AK were essential factors to regulate the AOA and AOB communities. The regulatory effects of AP and AK on the AOA and AOB communities should not be ignored. The interaction between the AOA and AOB communities was simple in spring, but on the contrary in autumn, the AOB community might be the main driver of ammoxidation in autumn.
Author Contributions
All authors declare that everyone contributed adequately to the experimental procedures and writing of the manuscript. F.W. conceived and designed the study. X.H. and S.L. performed the experiments. X.H. analyzed the data and wrote the manuscript. All the authors have read and approved the final manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the China Agricultural Research System (CARS-23-C-10).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request. The raw data have been deposited in the NCBI SRA database (AOA: SRP 160125; AOB: SRP160403). Compliance with ethical standards.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fan, F.L.; Yang, Q.B.; Li, Z.J.; Wei, D.; Cui, X.A.; Liang, Y.C. Impacts of organic and inorganic fertilizers on nitrification in a cold climate soil are linked to the bacterial ammonia oxidizer community. Microb. Ecol. 2011, 62, 982–990. [Google Scholar] [CrossRef] [PubMed]
- Li, N.H.; Gao, D.M.; Zhou, X.G.; Chen, S.C.; Li, C.X.; Wu, F.Z. Intercropping with Potato-Onion Enhanced the Soil Microbial Diversity of Tomato. Microorganisms 2020, 8, 834. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Jones, C.M.; Meijer, J.; Lundquista, P.O.; Franssonb, P.; Carlssonc, G.; Hallin, S. Intercropping affects genetic potential for inorganic nitrogen cycling by root -associated microorganisms in Medicago sativa and Dactylis glomerata. Appl. Soil Ecol. 2017, 119, 260–266. [Google Scholar] [CrossRef]
- Chu, H.Y.; Fujii, T.; Morimoto, S.; Lin, X.G.; Yagi, K. Population size and specific nitrification potential of soil ammonia-oxidizing bacteria under long-term fertilizer management. Soil Biol. Biochem. 2008, 40, 1960–1963. [Google Scholar] [CrossRef]
- Philippot, L.; Hallin, S. Towards food, feed and energy crops mitigating climate change. Trends Plant Sci. 2011, 16, 476–480. [Google Scholar] [CrossRef]
- Yao, H.Y.; Gao, Y.M.; Nicol, G.W.; Campbell, C.D.; Prosser, J.I.; Zhang, L.M. Links between ammonia oxidizer community structure, abundance, and nitrification potential in acidic soil. Appl. Environ. Microbiol. 2011, 77, 4618–4625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.L.; Liu, X.R.; Zhan, Q.W. Effects of combined biochar and organic fertilizer on nitrous oxide fluxes and the related nitrifier and denitrifier communities in a saline-alkali soil. Sci. Total Environ. 2019, 686, 199–211. [Google Scholar] [CrossRef]
- Zhang, N.N.; Sun, Y.M.; Wang, E.T.; Yang, J.S.; Yuan, H.L.; Scow, K.M. Effects of intercropping and Rhizobial inoculation on the ammonia-oxidizing microorganisms in rhizospheres of maize and faba bean plants. Appl. Soil Ecol. 2015, 85, 76–85. [Google Scholar] [CrossRef]
- Ding, Y.; Huang, X.; Li, Y.; Liu, H.Y.; Zhang, Q.C.; Liu, X.M. Nitrate leaching losses mitigated with intercropping of deep-rooted and shallow-rooted plants. J. Soils Sediments 2021, 21, 364–375. [Google Scholar] [CrossRef]
- Chen, Y.L.; Hu, H.W.; Han, H.Y.; Du, Y.; Wn, S.Q.; Xu, Z.W.; Chen, B.D. Abundance and community structure of ammonia-oxidizing Archaea and Bacteria in response to fertilization and mowing in a temperate steppe in Inner Mongolia. FEMS Microbiol. Ecol. 2014, 89, 67–79. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Zhang, L.M.; Yuan, C.; Zheng, Y.; Wang, J.; Chen, D.; He, J. The large-scale distribution of ammonia oxidizers in paddy soils is driven by soil pH, geographic distance, and climatic factors. Front. Microbiol. 2015, 6, 938. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, X.; Li, Y.; Ran, Y.; Liu, Y.; Zhang, Q. High temperatures inhibited the growth of soil bacteria and archaea but not that of fungi and altered nitrous oxide production mechanisms from different nitrogen sources in an acidic soil. Soil Biol. Biochem. 2017, 107, 168–179. [Google Scholar] [CrossRef]
- Ming, H.X.; Zhang, H.Z.; Chen, Q.R.; Wang, Y.T.; Su, J.; Zhao, X.H. Abundance and community structure of ammonium monooxygenase (amoA) genes in the wet season of liaohe estuary sediments. Cont. Shelf Res. 2020, 209, 10425. [Google Scholar] [CrossRef]
- Yang, Y.D.; Ren, Y.F.; Wang, X.Q.; Hu, Y.G.; Wang, Z.M.; Zeng, Z.H. Ammonia-oxidizing archaea and bacteria responding differently to fertilizer type and irrigation frequency as revealed by Illumina Miseq sequencing. J. Soil Sediment 2018, 18, 1029–1040. [Google Scholar] [CrossRef]
- Pereira e Silva, M.C.; Poly, F.; Guillaumaud, N.; van Elsas, J.D.; Salles, J.F. Fluctuations in ammonia oxidizing communities across agricultural soils are driven by soil structure and pH. Front. Microbiol. 2012, 3, 77. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.M.; Alves, R.J.E.; Zhang, D.D.; Han, L.L.; He, J.Z.; Zhang, L.M. Time-dependent shifts in populations and activity of bacterial and archaeal ammonia oxidizers in response to liming in acidic soils. Soil Biol. Biochem. 2017, 112, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Verhamme, D.T.; Prosser, J.I.; Nicol, G.W. Ammonia concentration determines differential growth of ammonia-oxidising archaea and bacteria in soil microcosms. ISME J. 2011, 5, 1067–1071. [Google Scholar] [CrossRef] [Green Version]
- Li, J.L.; Nedwell, D.B.; Beddow, J.; Dumbrell, A.J.; McKew, B.A.; Thorpe, E.L. amoA gene abundances and nitrification potential rates suggest that benthic ammonia-oxidizing bacteria and not archaea dominate N cycling in the Colne Estuary, United Kingdom. Appl. Environ. Microbiol. 2015, 81, 159–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di, H.J.; Cameron, K.C.; Shen, J.P.; Winefield, C.S.; O’Callaghan, M.; Bowatte, S. Nitrification driven by bacteria and not archaea in nitrogen-rich grassland soils. Nat. Geosci. 2009, 2, 621–624. [Google Scholar] [CrossRef]
- Shen, J.P.; Zhang, L.M.; Di, H.J.; He, J.Z. A review of ammonia-oxidizing bacteria and archaea in Chinese soils. Front. Microbiol. 2012, 3, 296–303. [Google Scholar] [CrossRef] [Green Version]
- Van Kessel, M.A.H.J.; Speth, D.R.; Albertsen, M.; Nielsen, P.H.; Op den Camp, H.J.M.; Kartal, B. Complete nitrification by a single microorganism. Nature 2015, 528, 555–559. [Google Scholar] [CrossRef] [Green Version]
- Daims, H.; Lebedeva, E.V.; Pjevac, P.; Han, P.; Herbold, C.; Albertsen, M. Complete nitrifi-cation by Nitrospira bacteria. Nature 2015, 528, 504–509. [Google Scholar] [CrossRef]
- Meng, H.; Katayama, Y.; Gu, J.D. More wide occurrence and dominance of ammonia-oxidizing archaea than bacteria at three Angkor sandstone temples of Bayon, Phnom Krom and Wat Athvea in Cambodia. Int. Biodeterior. Biodegrad. 2017, 117, 78–88. [Google Scholar] [CrossRef]
- Tang, H.M.; Xiao, X.P.; Li, C.; Shi, L.H.; Cheng, K.K.; Wen, L. Influences of different manure N input on soil ammonia-oxidizing archaea and bacterial activity and community structure in a double-cropping rice field. J. Appl. Microbiol. 2020. [Google Scholar] [CrossRef]
- Fu, X.; Wu, X.; Zhou, X.; Liu, S.; Shen, Y.; Wu, F. Companion cropping with potato onion enhances the disease resistance of tomato against Verticillium dahliae. Front. Plant Sci. 2015, 6, 726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Wu, F.; Zhou, X.; Fu, X.; Tao, Y.; Xu, W.; Pan, K.; Liu, S. Effects of Intercropping with Potato Onion on the Growth of Tomato and Rhizosphere Alkaline Phosphatase Genes Diversity. Front. Plant Sci. 2016, 7, 846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.L.; Fu, X.P.; Zhou, X.G.; Guo, M.Y.; Wu, F.Z. Effects of seven different companion plants on cucumber productivity, soil chemical characteristics and Pseudominas community. J. Integr. Agric. 2017, 16, 2206–2214. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Wu, F.Z. Diversity and Co-occurrence patterns of soil bacterial and fungal communities in seven intercropping systems. Front. Microbiol. 2018, 9, 1521. [Google Scholar] [CrossRef]
- Schinner, F.; Öhliger, R.; Kandeler, E.; Margesin, R. Methods in Soil Biology; Springer: Berlin/Heidelberg, Germany, 1996. [Google Scholar]
- Francis, C.A.; Roberts, K.J.; Beman, J.M.; Santoro, A.E.; Oakley, B.B. Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean. Proc. Natl. Acad. Sci. USA 2005, 102, 14683–14688. [Google Scholar] [CrossRef] [Green Version]
- Rotthauwe, J.H.; Witzel, K.P.; Liesack, W. The ammonia monooxygenase structural gene amoA as a functional marker: Molecular fine-scale analysis of natural ammonia-oxidizing populations. Appl. Environ. Microbiol. 1997, 63, 4704–4712. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.N.; Xu, H.S.; Jiang, Y.; Zhang, K.; Hu, Y.G.; Zeng, Z.H. Methane emissions and microbial communities as influenced by dual cropping of Azolla along with early rice. Sci. Rep. 2017, 7, 40635. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ni, L.; Song, Y.; Rhodes, G.; Li, J.; Huang, Q.; Shen, Q. Dynamic response of ammonia-oxidizers to four fertilization regimes across a wheat-rice rotation system. Front. Microbiol. 2017, 8, 630. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cong, W.F.; Hoffland, E.; Li, L.; Six, J.; Sun, J.H.; Bao, X.G. Intercropping enhances soil carbon and nitrogen. Glob. Chang. Biol. 2015, 21, 1715–1726. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.J.; Chen, S.C.; Zhou, X.G.; Wu, F.Z. Root interactions and tomato growth in tomato/potato onion companion-cropping system under different phosphorus levels. J. Plant Interact. 2017, 12, 438–466. [Google Scholar] [CrossRef] [Green Version]
- Bedoussac, L.; Justes, E. The efficiency of a durum wheat-winter pea intercrop to improve yield and wheat grain protein concentration depends on N availability during early growth. Plant Soil 2010, 330, 19–35. [Google Scholar] [CrossRef]
- Sun, R.; Myrold, D.D.; Wang, D.Z.; Guo, X.S.; Chu, H.Y. AOA and AOB communities respond differently to changes of soil pH under long-term fertilization. Soil Ecol. Lett. 2019, 1, 126–135. [Google Scholar] [CrossRef] [Green Version]
- Lawson, A.; Fortuna, A.M.; Cogger, C.; Bary, A.; Stubbs, T. Nitrogen contribution of rye-hairy vetch cover crop mixtures to organically grown sweet corn. Renew. Agric. Food Syst. 2013, 28, 59–69. [Google Scholar] [CrossRef]
- Taylor, A.E.; Zeglin, L.H.; Wanzek, T.A.; Myrold, D.D.; Bottomley, P.J. Dynamics of ammonia-oxidizing archaea and bacteria populations and contributions to soil nitrification potentials. ISME J. 2012, 6, 2024–2032. [Google Scholar] [CrossRef]
- Goldberg, D.E. Competitive ability: Definitions, contingency and correlated traits. Philos. Trans. R. Soc. B 1996, 351, 1377–1385. [Google Scholar]
- Zhang, S.; Lamb, E.G. Plant competitive ability and the transitivity of competitive hierarchies change with plant age. Plant Ecol. 2011, 213, 15–23. [Google Scholar] [CrossRef]
- Könneke, M.; Bernhard, A.E.; Torre, J.R.; Walker, C.B.; Waterbury, J.B.; Stahl, D.A. Isolation of an autotrophic ammonia-oxidizing marine archaeon. Nature 2005, 437, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Leininger, S.; Urich, T.; Schloter, M.; Schwark, L.; Qi, J.; Nicol, G.W. Archaea predominate among ammonia-oxidizing prokaryotes in soils. Nature 2006, 442, 806–809. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.F.; Shi, X.J.; Zheng, Y.; Qin, Z.X.; Xie, D.T.; Li, Z.L. Abundance and community composition of ammonia-oxidizing bacteria and archaea in purple soil under long-term fertilization. Eur. J. Soil Biol. 2014, 60, 24–33. [Google Scholar] [CrossRef]
- Wang, Y.F.; Feng, Y.Y.; Ma, X.J.; Gu, J.D. Seasonal dynamics of ammonia/ammonium-oxidizing prokaryotes in oxic and anoxic wetland sediments of subtropical coastal mangrove. Appl. Microbiol. Biotechnol. 2013, 97, 7919–7934. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, C.A.; Wakelin, S.A.; Fillery, I.R.P.; Roper, M.M. Factors affecting ammonia-oxidising microorganisms and potential nitrification rates in southern Australian agricultural soils. Soil Res. 2013, 51, 240–252. [Google Scholar] [CrossRef]
- Zhou, G.; Zhang, J.; Zhang, C.; Feng, Y.Z.; Chen, L.; Yu, Z.H. Effects of changes in straw chemical properties and alkaline soils on bacterial communities engaged in straw decomposition at different temperature. Sci. Rep. 2016, 6, 22186–22198. [Google Scholar] [CrossRef] [Green Version]
- Roy, D.; McEvoy, J.; Blonigen, M.; Amundson, M.; Khan, E. Seasonal variation and ex-situ nitrification activity of ammonia oxidizing archaea in biofilm based wastewater treatment processes. Bioresour. Technol. 2017, 244, 850–859. [Google Scholar] [CrossRef]
- Wang, S.Y.; Wang, W.D.; Liu, L.; Zhuang, L.J.; Zhao, S.Y.; Su, Y. Microbial nitrogen cycle hotspots in the plant-bed/ditch system of a constructed wetland with N2O mitigation. Environ. Sci. Technol. 2018, 52, 6226–6236. [Google Scholar] [CrossRef]
- Szukics, U.; Hackl, E.; Boltenstern, S.Z.; Sessitsch, A. Rapid and dissimilar response of ammonia oxidizing archaea and bacteria to nitrogen and water amendment in two temperate forest soils. Microbiol. Res. 2012, 167, 103–109. [Google Scholar] [CrossRef]
- Nicol, G.W.; Leininger, S.; Schleper, C.; Prosser, J.I. The influence of soil pH on the diversity, abundance and transcriptional activity of ammonia oxidizing archaea and bacteria. Environ. Microbiol. 2008, 10, 2966–2978. [Google Scholar] [CrossRef]
- Hayden, H.L.; Drake, J.; Imhof, M.; Oxley, A.P.A.; Norng, S.; Mele, P.M. The abundance of nitrogen cycle genes amoA and nifH depends on land-uses and soil types in South-Eastern Australia. Soil Biol. Biochem. 2010, 42, 1774–1783. [Google Scholar] [CrossRef]
- Jiang, X.; Hou, X.; Zhou, X.; Xin, X.; Wright, A.; Jia, Z. pH regulates key players of nitrification in paddy soils. Soil Biol. Biochem. 2015, 81, 9–16. [Google Scholar] [CrossRef]
- Luo, Y.; Yu, Z.; Zhang, K.; Xu, J.; Brookes, P.C. The properties and functions of biochars in forest ecosystems. J. Soils Sediments 2016, 16, 2005–2020. [Google Scholar] [CrossRef]
- Xiao, H.F.; Schaefer, D.A.; Yang, X.D. pH drives ammonia oxidizing bacteria rather than archaea thereby stimulate nitrification under Ageratina adenophora colonization. Soil Biol. Biochem. 2017, 114, 12–19. [Google Scholar] [CrossRef]
- Xia, W.W.; Zhang, C.X.; Zeng, X.W.; Feng, Y.Z.; Weng, J.H.; Lin, X.G. Autotrophic growth of nitrifying community in an agricultural soil. ISME J. 2011, 5, 1226–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allison, S.M.; Prosser, J.I. Ammonia oxidation at low pH by attached populations of nitrifying bacteria. Soil Biol. Biochem. 1993, 25, 935–941. [Google Scholar] [CrossRef]
- Hayatsu, M.; Tago, K.; Uchiyama, I.; Toyoda, A.; Wang, Y.; Shimomura, Y. An acid-tolerant ammonia-oxidizing γ-proteobacterium from soil. ISME J. 2017, 11, 1130–1141. [Google Scholar] [CrossRef]
- Huang, X.; Zhao, J.; Su, J.; Jia, Z.; Shi, X.; Wright, A.L. Neutrophilic bacteria are responsible for autotrophic ammonia oxidation in an acidic forest soil. Soil Biol. Biochem. 2018, 119, 83–89. [Google Scholar] [CrossRef]
- Di, H.J.; Cameron, K.C.; Shen, J.P.; Winefield, C.S.; O’Callaghan, M.; Bowatte, S. Ammonia-oxidizing bacteria and archaea grow under contrasting soil nitrogen conditions. FEMS Microbiol. Ecol. 2010, 72, 386–394. [Google Scholar] [CrossRef] [Green Version]
- Wertz, S.; Goyer, C.; Zebarth, B.J.; Burton, D.L.; Tatti, E.; Chantigny, M.H. Effects of temperatures near the freezing point on N2O emissions, denitrification and on the abundance and structure of nitrifying and denitrifying soil communities. FEMS Microbiol. Ecol. 2013, 83, 242–254. [Google Scholar] [CrossRef] [Green Version]
- Avrahami, S.; Bohannan, B.J.M. Response of Nitrosospira sp. strain AF-like ammonia oxidizers to changes in temperature, soil moisture content, and fertilizer concentration. Appl. Environ. Microbiol. 2007, 73, 1166–1173. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zhang, L.M.; Shen, J.P.; Wei, W.X.; He, J.Z. Abundance and community composition of ammonia-oxidizing archaea and bacteria in an acid paddy soil. Biol. Fert. Soils 2011, 47, 323–331. [Google Scholar] [CrossRef]
- Stopnišek, N.; Rangin, G.C.; Höfferle, S.; Nicol, G.W.; Mulec, I.M.; Prosser, J.I. Thaumarchaeal ammonia oxidation in an acidic forest peat soil is not influenced by ammonium amendment. Appl. Environ. Microbiol. 2010, 76, 7626–7634. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.L.; Hou, L.J.; Newell, S.; Liu, M.; Zhou, J.L.; Zhao, H. Community dynamics and activity of ammonia-oxidizing prokaryotes in intertidal sediments of the Yangtze Estuary. Appl. Environ. Microbiol. 2014, 80, 408–419. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Zhang, X.; Li, D.; Wang, H.; Chen, F.; Fu, X. Impacts of nitrogen and phosphorus additions on the abundance and community structure of ammonia oxidizers and denitrifying bacteria in Chinese fir plantations. Soil Biol. Biochem. 2016, 103, 284–293. [Google Scholar] [CrossRef]
- Olesen, J.M.; Bascompte, J.; Dupont, Y.L.; Jordano, P. The modularity of pollination networks. Proc. Natl. Acad. Sci. USA 2007, 104, 19891–19896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.Q.; Zhao, D.Y.; Zeng, J.; Cao, X.Y.; Jiao, C.C. Network Analysis Reveals Seasonal Patterns of Bacterial Community Networks in Lake Taihu under Aquaculture Conditions. Water 2019, 11, 1868. [Google Scholar] [CrossRef] [Green Version]
- Siles, J.A.; Margesin, R. Seasonal soil microbial responses are limited to changes in functionality at two Alpine forest sites differing in altitude and vegetation. Sci. Rep. 2017, 7, 2204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
The different intercropping systems and the cucumber monocultured system in the greenhouse experiment.
Figure 1.
The different intercropping systems and the cucumber monocultured system in the greenhouse experiment.

Figure 2.
Effects of different intercropping systems on the potential nitrification rate of cucumber in spring and autumn. A—alfalfa; T—trifolium; W—wheat; Ry—rye; C—chrysanthemum; Ra—rape; M—mustard; CM—cucumber monoculture. Columns with different letters are significantly different at 0.05 probability level.
Figure 2.
Effects of different intercropping systems on the potential nitrification rate of cucumber in spring and autumn. A—alfalfa; T—trifolium; W—wheat; Ry—rye; C—chrysanthemum; Ra—rape; M—mustard; CM—cucumber monoculture. Columns with different letters are significantly different at 0.05 probability level.

Figure 3.
The abundances of soil ammonia-oxidizing archaea (AOA) (a), ammonia-oxidizing bacteria (AOB) (b), and ratio AOA/AOB during spring (c) and autumn (d) in different treatments. A—alfalfa; T—trifolium; W—wheat; Ry—rye; C—chrysanthemum; Ra—rape; M—mustard; CM—cucumber monoculture. Columns with different letters are significantly different at 0.05 probability level.
Figure 3.
The abundances of soil ammonia-oxidizing archaea (AOA) (a), ammonia-oxidizing bacteria (AOB) (b), and ratio AOA/AOB during spring (c) and autumn (d) in different treatments. A—alfalfa; T—trifolium; W—wheat; Ry—rye; C—chrysanthemum; Ra—rape; M—mustard; CM—cucumber monoculture. Columns with different letters are significantly different at 0.05 probability level.

Figure 4.
Nonmetric multidimensional scaling (NMDS) based on Bray–Curtis distances at the operational taxonomic unit (OTU) level of soil AOA (a) and AOB (b) communities in spring and autumn.
Figure 4.
Nonmetric multidimensional scaling (NMDS) based on Bray–Curtis distances at the operational taxonomic unit (OTU) level of soil AOA (a) and AOB (b) communities in spring and autumn.

Figure 5.
Changes in the relative abundance at phyla (a,b) and genus (c,d) levels of AOA and AOB in different treatments in spring (S) and autumn (A). A—alfalfa; T—trifolium; W—wheat; Ry—rye; C—chrysanthemum; Ra—rape; M—mustard; CM—cucumber monoculture.
Figure 5.
Changes in the relative abundance at phyla (a,b) and genus (c,d) levels of AOA and AOB in different treatments in spring (S) and autumn (A). A—alfalfa; T—trifolium; W—wheat; Ry—rye; C—chrysanthemum; Ra—rape; M—mustard; CM—cucumber monoculture.

Figure 6.
Redundancy analysis (RDA) demonstrating the relationships between soil environmental factors and soil AOA and AOB communities in spring (a,b) and autumn (c,d). A—alfalfa; T—trifolium; W—wheat; Ry—rye; C—chrysanthemum; Ra—rape; M—mustard; CM—cucumber monoculture. * p < 0.05, ** p < 0.01, *** p < 0.001.
Figure 6.
Redundancy analysis (RDA) demonstrating the relationships between soil environmental factors and soil AOA and AOB communities in spring (a,b) and autumn (c,d). A—alfalfa; T—trifolium; W—wheat; Ry—rye; C—chrysanthemum; Ra—rape; M—mustard; CM—cucumber monoculture. * p < 0.05, ** p < 0.01, *** p < 0.001.

Figure 7.
Co-occurrence network of ammonia oxidizing bacteria communities in different seasons. The AOA community in spring (a) and autumn (b), AOB community in spring (c) and autumn (d), AOA and AOB communities in spring (e) and autumn (f). A link represents a significant correlation (Spearman, |ρ| > 0.8, p < 0.05). The blue line represents positive correlation and the red line represents negative correlation. The node size is proportional to the number of connections. A and B after OTUs represent AOA and AOB, respectively. Only the groups with the top 50 average relative abundance of AOA and AOB were shown.
Figure 7.
Co-occurrence network of ammonia oxidizing bacteria communities in different seasons. The AOA community in spring (a) and autumn (b), AOB community in spring (c) and autumn (d), AOA and AOB communities in spring (e) and autumn (f). A link represents a significant correlation (Spearman, |ρ| > 0.8, p < 0.05). The blue line represents positive correlation and the red line represents negative correlation. The node size is proportional to the number of connections. A and B after OTUs represent AOA and AOB, respectively. Only the groups with the top 50 average relative abundance of AOA and AOB were shown.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Effects of different intercropping systems on soil physicochemical properties at two sampling times.
Table 1.
Effects of different intercropping systems on soil physicochemical properties at two sampling times.
| Season | Treatment | Moisture (%) | pH | -N (mg·kg−1) | -N (mg·kg−1) | AP (mg·kg−1) | AK (mg·kg−1) |
|---|---|---|---|---|---|---|---|
| Spring | A | 24.94 ± 5.44 a | 7.32 ± 0.05 a | 27.66 ± 8.17 b | 35.13 ± 9.37 a | 467.14 ± 53.44 b | 533.20 ± 110.66 a |
| T | 23.44 ± 0.41 a | 7.18 ± 0.05 a | 15.33 ± 6.96 b | 63.61 ± 26.32 a | 427.53 ± 51.22 b | 561.11 ± 78.41 a | |
| W | 22.47 ± 0.71 a | 7.16 ± 0.09 a | 16.51 ± 2.76 b | 62.78 ± 24.81 a | 308.54 ± 83.92 c | 566.91 ± 49.71 a | |
| Ry | 22.54 ± 4.01 a | 7.22 ± 0.15 a | 56.31 ± 28.18 a | 54.58 ± 25.28 a | 276.72 ± 66.06 c | 460.27 ± 21.98 a | |
| C | 19.51 ± 1.40 a | 7.22 ± 0.17 a | 30.37 ± 11.53 b | 41.65 ± 14.32 a | 237.93 ± 49.81 c | 589.08 ± 49.52 a | |
| Ra | 21.00 ± 4.73 a | 7.17 ± 0.09 a | 20.39 ± 9.53 b | 37.18 ± 15.57 a | 430.65 ± 50.62 b | 619.80 ± 160.82 a | |
| M | 24.71 ± 2.66 a | 7.20 ± 0.21 a | 35.92 ± 6.91 a b | 33.29 ± 14.66 a | 505.27 ± 111.20 b | 503.53 ± 54.84 a | |
| CM | 23.41 ± 1.31 a | 7.10 ± 0.06 a | 26.77 ± 9.94 b | 44.96 ± 21.57 a | 621.85 ± 41.44 a | 617.54 ± 36.36 a | |
| Autumn | A | 25.50 ± 2.86 a | 6.81 ± 0.14 a | 13.15 ± 3.86 a | 43.64 ± 2.99 a | 388.36 ± 40.70 a | 546.25 ± 37.31 a |
| T | 26.68 ± 1.36 a | 6.78 ± 0.05 a | 12.82 ± 2.92 a | 36.51 ± 3.58 a b | 386.53 ± 37.81 a | 517.75 ± 26.07 b | |
| W | 19.98 ± 8.27 a | 6.91 ± 0.05 a | 22.25 ± 8.56 a | 34.14 ± 3.03 b | 390.63 ± 19.06 a | 431.84 ± 38.31 b | |
| Ry | 26.49 ± 2.73 a | 6.85 ± 0.05 a | 17.63 ± 3.03 a | 34.25 ± 7.82 b | 427.94 ± 12.86 a | 530.36 ± 70.57 a b | |
| C | 25.82 ± 1.76 a | 6.80 ± 0.06 a | 19.60 ± 7.38 a | 43.91 ± 3.41 a | 358.98 ± 26.13 a | 525.23 ± 56.65 a b | |
| Ra | 24.39 ± 0.88 a | 6.84 ± 0.08 a | 19.36 ± 8.49 a | 39.16 ± 8.27 a b | 368.92 ± 59.10 a | 506.17 ± 95.25 a b | |
| M | 25.87 ± 2.33 a | 6.86 ± 0.11 a | 21.33 ± 3.87 a | 32.78 ± 1.70 b | 393.76 ± 34.35 a | 515.80 ± 40.82 a b | |
| CM | 19.11 ± 9.03 a | 6.75 ± 0.08 a | 18.81 ± 4.15 a | 33.66 ± 1.72 b | 425.29 ± 30.81 a | 521.20 ± 22.51 a b |
A—alfalfa; T—trifolium; W—wheat; Ry—rye; C—chrysanthemum; Ra—rape; M—mustard; CM—cucumber monoculture. AP, AK indicate soil available phosphorus and available potassium, respectively. Columns with different letters are significantly different at 0.05 probability level.
Table 2.
Pearson correlation coefficients between soil properties and AOA and AOB abundances.
| Season | Abundance | Moisture | pH | -N | -N | AP | AK |
|---|---|---|---|---|---|---|---|
| Spring | AOA | −0.464 * | −0.069 | −0.201 | 0.475 * | −0.087 | −0.278 |
| AOB | −0.253 | −0.576 ** | 0.331 | 0.598 ** | −0.153 | −0.152 | |
| Autumn | AOA | −0.139 | 0.201 | 0.167 | 0.467 * | −0.459* | −0.464 * |
| AOB | −0.018 | −0.213 | 0.369 | 0.452 * | 0.135 | 0.004 |
* p < 0.05; ** p < 0.01.
Table 3.
Illumina MiSeq sequencing results and alpha diversity estimates for soil AOA and AOB communities.
Table 3.
Illumina MiSeq sequencing results and alpha diversity estimates for soil AOA and AOB communities.
| Classified | Season | Treatment | Sequences | Number of OTUs | Shannon | Simpson | Chao 1 | Coverage (%) |
|---|---|---|---|---|---|---|---|---|
| AOA | Spring | A | 11,467 ± 903 | 52 ± 2.12 b | 2.35 ± 0.03 a b | 0.1413 ± 0.0010 c d | 69.25 ± 6.01 a | 99.90 |
| T | 11,860 ± 485 | 53 ± 0.71 a b | 2.34 ± 0.06 a b | 0.1408 ± 0.0003 b c | 64.0 ± 1.41 a b | 99.91 | ||
| W | 13,463 ± 2161 | 54 ± 1.41 a b | 2.36 ± 0.16 a b | 0.1668 ± 0.0157 a | 60.60 ± 4.81 b c | 99.92 | ||
| Ry | 11,599 ± 1216 | 50 ± 2.83 a b | 2.22 ± 0.24 b | 0.1593 ± 0.0085 a b | 54.75 ± 3.89 c d | 99.95 | ||
| C | 11,926 ± 2186 | 57 ± 1.41 a | 2.50 ± 0.05 a | 0.1292 ± 0.0029 d | 60.93 ± 1.04 b c | 99.90 | ||
| Ra | 13,620 ± 1645 | 50 ± 0.71 b | 2.30 ± 0.11 a b | 0.1698 ± 0.0042 a | 50.23 ± 0.89 d | 99.96 | ||
| M | 12,758 ± 1706 | 52 ± 4.24 a b | 2.37 ± 0.03 a b | 0.1345 ± 0.0060 c d | 47.38 ± 3.36 d | 99.95 | ||
| CM | 12,400 ± 571 | 52 ± 0.71 b | 2.48 ± 0.11 a | 0.1386 ± 0.0022 c d | 52.29 ± 0.20 d | 99.94 | ||
| Autumn | A | 12,711 ± 1284 | 52 ± 2.12 a b c | 2.35 ± 0.14 a | 0.1403 ± 0.0014 b | 58.80 ± 0.28 a b c | 99.92 | |
| T | 13,821 ± 2446 | 55 ± 1.41 a | 2.34 ± 0.15 a | 0.1672 ± 0.0185 b | 62.00 ± 1.41 a b | 99.92 | ||
| W | 15,714 ± 2920 | 50 ± 0 b c | 2.06 ± 0.09 b | 0.2498 ± 0.0093 a | 52.00 ± 1.42 d | 99.95 | ||
| Ry | 15,114 ± 2652 | 53 ± 1.41 a b | 2.23 ± 0.07 a b | 0.1878 ± 0.0015 b | 56.27 ± 0.09 c | 99.92 | ||
| C | 12,214 ± 1629 | 52 ± 0 c | 2.06 ± 0.15 b | 0.2401 ± 0.0317 a | 62.17 ± 1.18 a | 99.92 | ||
| Ra | 15,193 ± 1976 | 53 ± 1.41 a b | 2.35 ± 0.09 a | 0.1650 ± 0.0067 b | 58.25 ± 2.47 b c | 99.92 | ||
| M | 16,022 ± 2551 | 52 ± 2.83 c | 2.17 ± 0.21 a b | 0.2405 ± 0.0387 a | 60.47 ± 1.60 a b | 99.93 | ||
| CM | 14,474 ± 608 | 51 ± 0 c | 2.20 ± 0.15 a b | 0.1569 ± 0.0159 b | 52.38 ± 2.30 d | 99.93 | ||
| AOB | Spring | A | 18,500 ± 1023 | 54 ± 2.65 a | 2.63 ± 0.22 a | 0.1236 ± 0.0405 a b | 62.14 ± 9.32 a | 99.92 |
| T | 19,921 ± 1211 | 54 ± 7.51 a | 2.74 ± 0.18 a | 0.1181 ± 0.0364 a b | 55.33 ± 8.81 a | 99.96 | ||
| W | 24,485 ± 991 | 53 ± 1.73 a | 2.62 ± 0.10 a | 0.1302 ± 0.0203 a b | 57.25 ± 4.63 a | 99.94 | ||
| Ry | 21,533 ± 599 | 57 ± 4.04 a | 2.65 ± 0.17 a | 0.1201 ± 0.0323 a b | 62.20 ± 10.05 a | 99.92 | ||
| C | 19,429 ± 1106 | 51 ± 2.08 a | 2.46 ± 0.18 a | 0.1612 ± 0.0394 a b | 54.65 ± 4.79 a | 99.93 | ||
| Ra | 21,797 ± 726 | 53 ± 4.62 a | 2.75 ± 0.03 a | 0.1023 ± 0.0050 b | 56.78 ± 3.89 a | 99.94 | ||
| M | 23,097 ± 1326 | 52 ± 1.0 a | 2.44 ± 0.23 a | 0.1688 ± 0.0355 a | 57.19 ± 3.76 a | 99.92 | ||
| CM | 17,184 ± 878 | 52 ± 4.51 a | 2.42 ± 0.12 a | 0.1699 ± 0.0231 a | 58.23 ± 9.46 a | 99.92 | ||
| Autumn | A | 18,457 ± 917 | 57 ± 1.53 a | 2.58 ± 0.27 a b | 0.1387 ± 0.0478 a b | 60.89 ± 1.46 a | 99.91 | |
| T | 17,755 ± 784 | 52 ± 7.0 c | 1.85 ± 0.62 b | 0.3449 ± 0.1940 a | 44.63 ± 8.29 c | 99.93 | ||
| W | 24,315 ± 1284 | 53 ± 1.15 b c | 2.46 ± 0.15 a b | 0.1623 ± 0.0413 a b | 48.58 ± 2.55 b c | 99.94 | ||
| Ry | 21,624 ± 1157 | 52 ± 8.02 a b | 2.41 ± 0.51 a b | 0.1882 ± 0.1410 a b | 57.05 ± 10.15 a b | 99.91 | ||
| C | 21,039 ± 1437 | 51 ± 1.53 b c | 1.85 ± 0.30 b | 0.3428 ± 0.0994 a | 48.76 ± 4.34 b c | 99.92 | ||
| Ra | 21,426 ± 554 | 52 ± 4.04 a b | 2.72 ± 0.16 a | 0.0991 ± 0.0180 b | 56.07 ± 2.69 a b | 99.93 | ||
| M | 24,196 ± 1451 | 51 ± 1.15 a b | 2.13 ± 0.28 a b | 0.2572 ± 0.0700 a b | 55.38 ± 4.89 a b | 99.92 | ||
| CM | 16,464 ± 343 | 51 ± 4.04 a b c | 2.37 ± 0.54 a b | 0.1995 ± 0.1582 a b | 54.67 ± 3.21 b c | 99.92 |
A—alfalfa; T—trifolium; W—wheat; Ry—rye; C—chrysanthemum; Ra—rape; M—mustard; CM—cucumber monoculture. Columns with different letters are significantly different at 0.05 probability level.
Table 4.
Topological properties of co-occurrence network of AOA and AOB in spring and autumn.
| Network Indicators | AOA (S) | AOA (A) | AOB (S) | AOB (A) | AOA and AOB (S) | AOA and AOB (A) |
|---|---|---|---|---|---|---|
| Nodes | 50 | 50 | 50 | 50 | 100 | 100 |
| Edges | 112 | 97 | 88 | 217 | 228 | 341 |
| Positive correlation edge numbers | 106 | 89 | 81 | 180 | 197 | 283 |
| Negative correlation edge numbers | 6 | 8 | 7 | 37 | 28 | 58 |
| Average path length (APL) | 2.875 | 4.226 | 2.891 | 2.580 | 3.607 | 4.174 |
| Graph density (GD) | 0.091 | 0.079 | 0.072 | 0.177 | 0.044 | 0.069 |
| Modularity (MD) | 0.707 | 0.897 | 0.722 | 0.466 | 1.141 | 0.676 |
| Clustering coefficient (CC) | 0.627 | 0.567 | 0.592 | 0.678 | 0.631 | 0.656 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
He, X.; Li, S.; Wu, F. Responses of Ammonia-Oxidizing Microorganisms to Intercropping Systems in Different Seasons. Agriculture 2021, 11, 195. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11030195
AMA Style
He X, Li S, Wu F. Responses of Ammonia-Oxidizing Microorganisms to Intercropping Systems in Different Seasons. Agriculture. 2021; 11(3):195. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11030195
Chicago/Turabian StyleHe, Xingjia, Sen Li, and Fengzhi Wu. 2021. "Responses of Ammonia-Oxidizing Microorganisms to Intercropping Systems in Different Seasons" Agriculture 11, no. 3: 195. https://0-doi-org.brum.beds.ac.uk/10.3390/agriculture11030195
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.